
Changes in Plasma Metabolic Signature upon Acute and Chronic Morphine Administration in Morphine-Tolerant Mice
 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Material and Methods
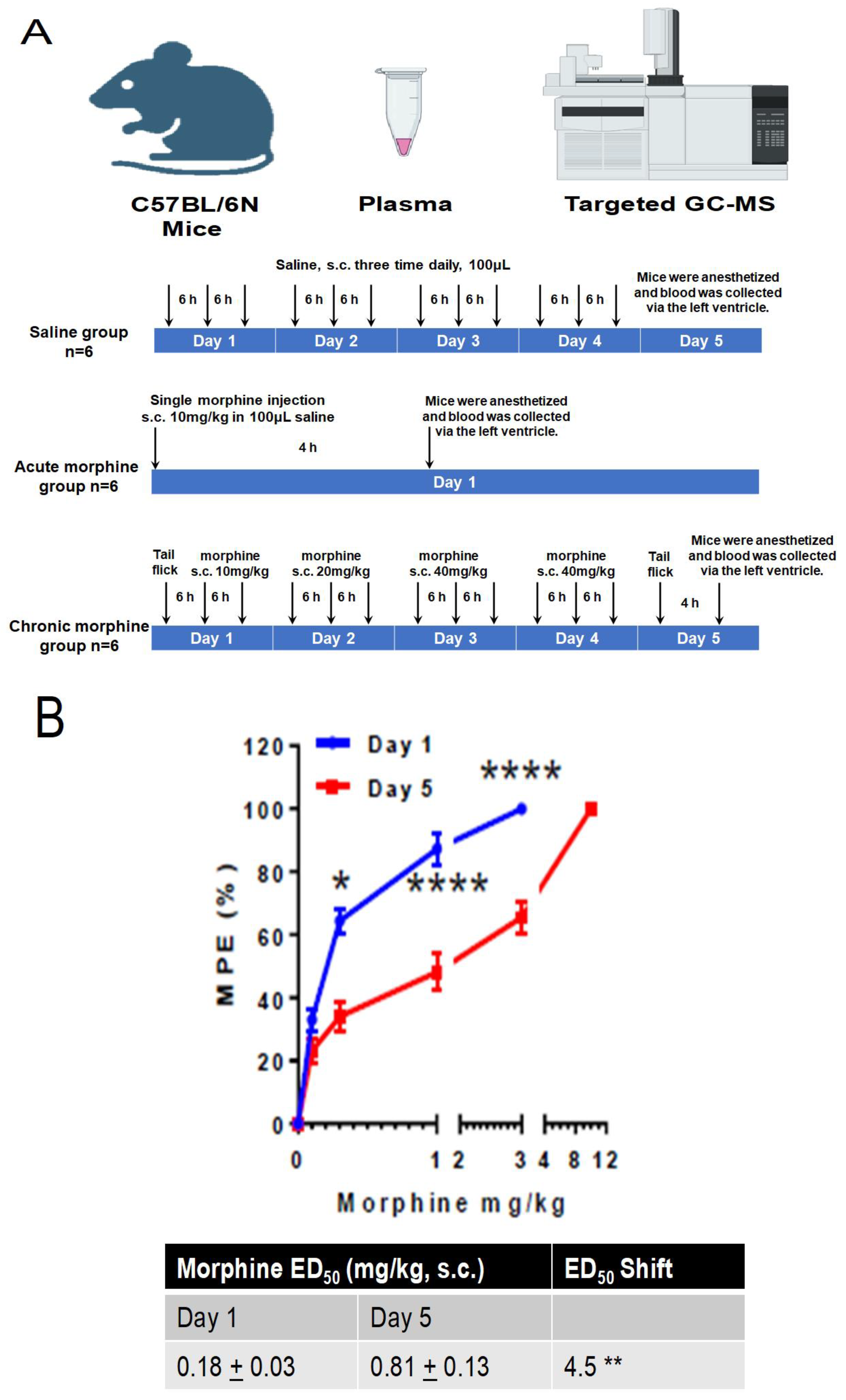
2.1. Study Design
2.2. Animals
2.3. Morphine Analgesia and Tolerance
2.4. Metabolite Profiling
2.5. Data Analysis
3. Results
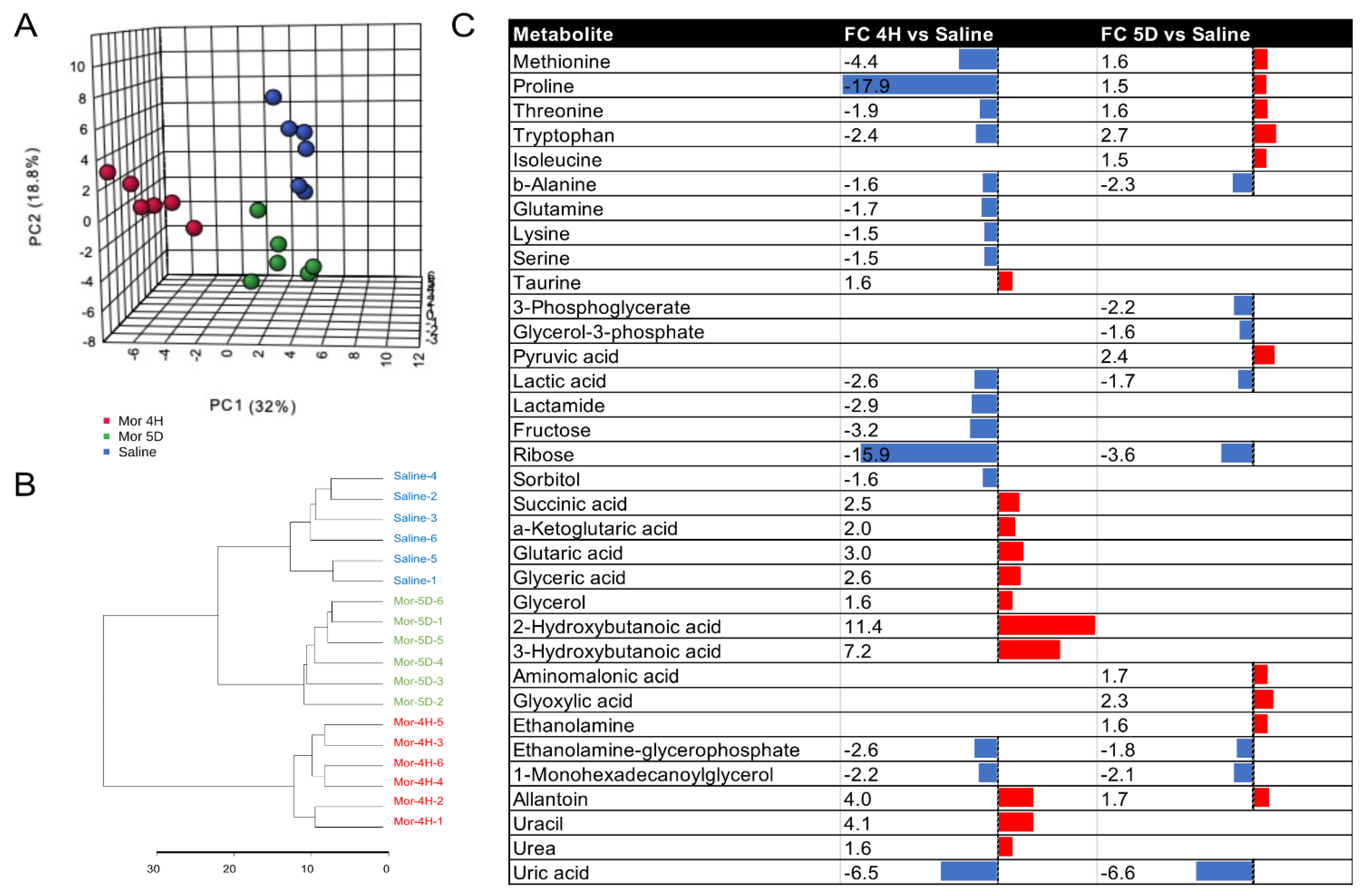
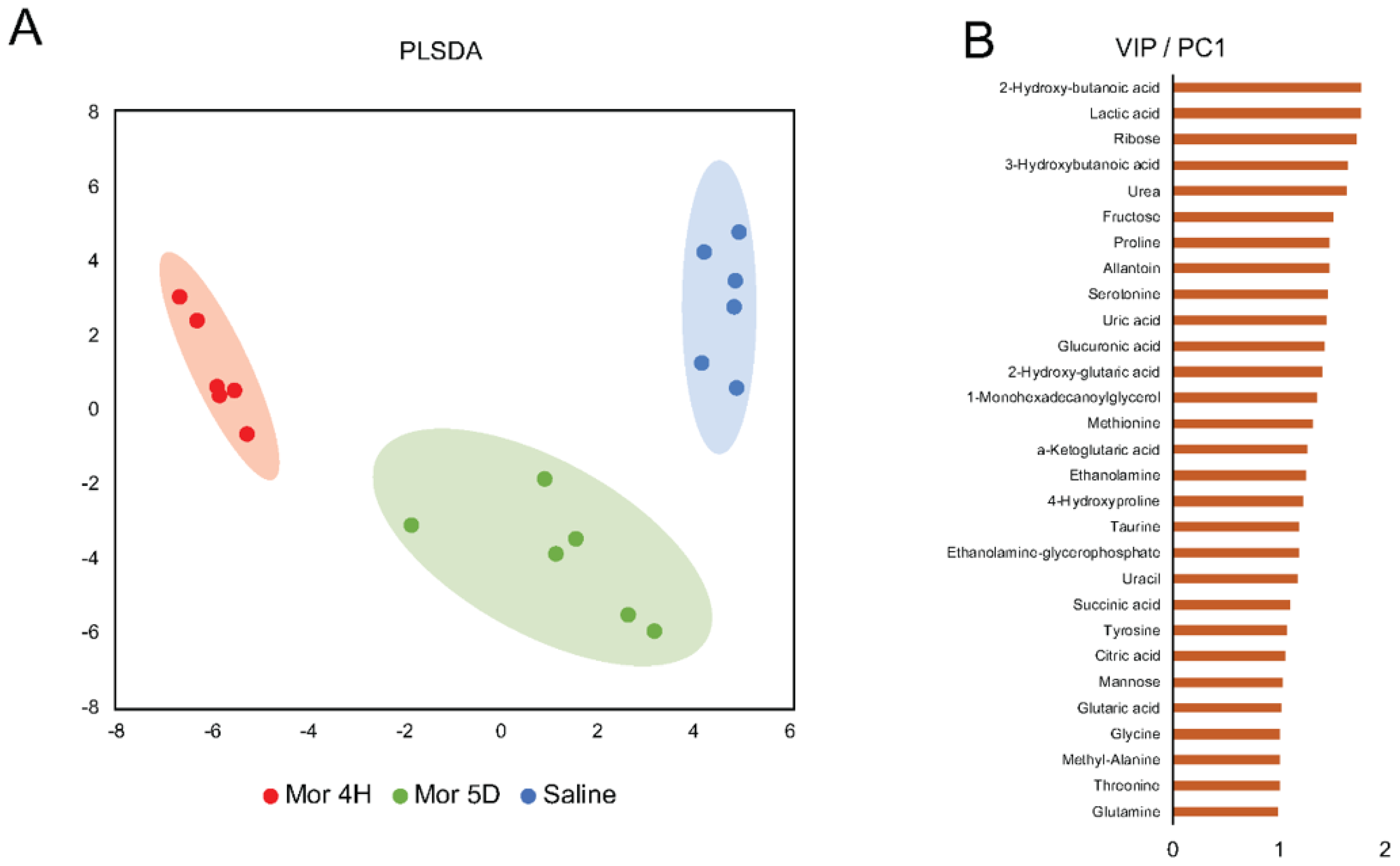
3.1. Metabolite Changes in Plasma Postacute or Chronic Morphine Intake in Tolerant Mice
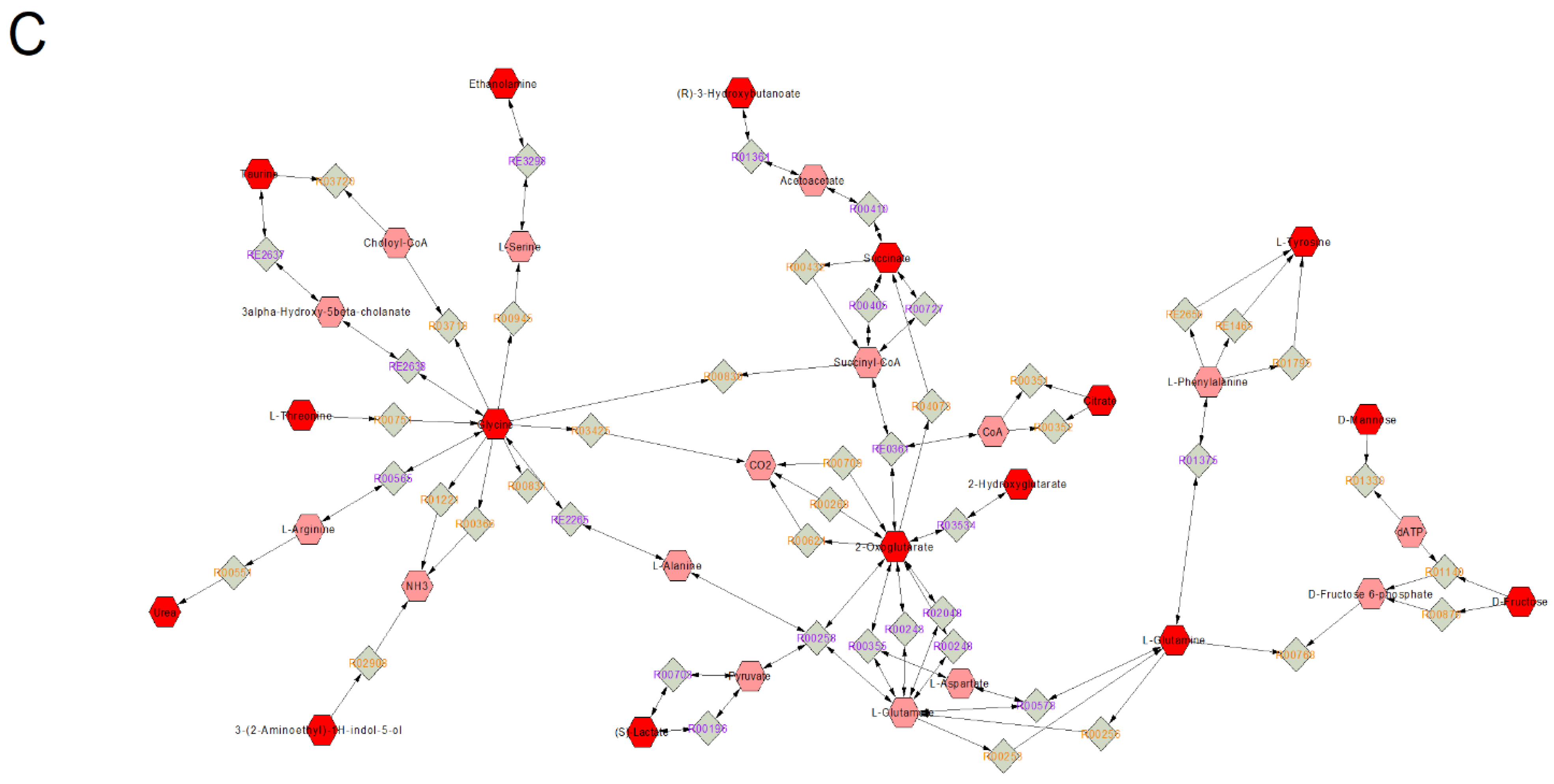
3.2. Pathways and Networks of Plasma Metabolites in Acute versus Chronic Intake
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wang, J.; Sun, H.; Sun, W.T.; Sun, H.P.; Tian, T.; Sun, J. Efficacy and safety of intrathecal morphine for pain control after spinal surgery: A systematic review and meta-analysis. Eur. Rev. Med. Pharmacol. Sci. 2021, 25, 2674–2684. [Google Scholar] [CrossRef]
- Tang, J.Z.J.; Weinberg, L. A Literature Review of Intrathecal Morphine Analgesia in Patients Undergoing Major Open Hepato-Pancreatic-Biliary (HPB) Surgery. Anesthesiol. Pain Med. 2019, 9, e94441. [Google Scholar] [CrossRef] [Green Version]
- Afsharimani, B.; Cabot, P.J.; Parat, M.O. Morphine use in cancer surgery. Front. Pharmacol. 2011, 2, 46. [Google Scholar] [CrossRef] [Green Version]
- Portenoy, R.K.; Thaler, H.T.; Inturrisi, C.E.; Friedlander-Klar, H.; Foley, K.M. The metabolite morphine-6-glucuronide contributes to the analgesia produced by morphine infusion in patients with pain and normal renal function. Clin. Pharmacol. Ther. 1992, 51, 422–431. [Google Scholar] [CrossRef] [PubMed]
- Osborne, R.; Thompson, P.; Joel, S.; Trew, D.; Patel, N.; Slevin, M. The analgesic activity of morphine-6-glucuronide. Br. J. Clin. Pharmacol. 1992, 34, 130–138. [Google Scholar] [CrossRef]
- Paul, D.; Standifer, K.M.; Inturrisi, C.E.; Pasternak, G.W. Pharmacological characterization of morphine-6 beta-glucuronide, a very potent morphine metabolite. J. Pharmacol. Exp. Ther. 1989, 251, 477–483. [Google Scholar] [PubMed]
- Pasternak, G.W.; Bodnar, R.J.; Clark, J.A.; Inturrisi, C.E. Morphine-6-glucuronide, a potent mu agonist. Life Sci. 1987, 41, 2845–2849. [Google Scholar] [CrossRef] [PubMed]
- Glare, P.A.; Walsh, T.D. Clinical pharmacokinetics of morphine. Ther. Drug Monit. 1991, 13, 1–23. [Google Scholar] [CrossRef]
- Frances, B.; Gout, R.; Campistron, G.; Panconi, E.; Cros, J. Morphine-6-glucuronide is more mu-selective and potent in analgesic tests than morphine. Prog. Clin. Biol. Res. 1990, 328, 477–480. [Google Scholar] [PubMed]
- Hucks, D.; Thompson, P.I.; McLoughlin, L.; Joel, S.P.; Patel, N.; Grossman, A.; Rees, L.H.; Slevin, M.L. Explanation at the opioid receptor level for differing toxicity of morphine and morphine 6-glucuronide. Br. J. Cancer 1992, 65, 122–126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghanbari, R.; Li, Y.; Pathmasiri, W.; McRitchie, S.; Etemadi, A.; Pollock, J.D.; Poustchi, H.; Rahimi-Movaghar, A.; Amin-Esmaeili, M.; Roshandel, G.; et al. Metabolomics reveals biomarkers of opioid use disorder. Transl. Psychiatry 2021, 11, 103. [Google Scholar] [CrossRef]
- Wang, F.; Meng, J.; Zhang, L.; Johnson, T.; Chen, C.; Roy, S. Morphine induces changes in the gut microbiome and metabolome in a morphine dependence model. Sci. Rep. 2018, 8, 3596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palermo, A.; Botre, F.; de la Torre, X.; Zamboni, N. Non-targeted LC-MS based metabolomics analysis of the urinary steroidal profile. Anal. Chim. Acta 2017, 964, 112–122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iannone, M.; Palermo, A.; de la Torre, X.; Romanelli, F.; Sansone, A.; Sansone, M.; Lenzi, A.; Botre, F. Effects of transdermal administration of testosterone gel on the urinary steroid profile in hypogonadal men: Implications in antidoping analysis. Steroids 2019, 152, 108491. [Google Scholar] [CrossRef]
- Mazzarino, M.; Buccilli, V.; de la Torre, X.; Fiacco, I.; Palermo, A.; Ughi, D.; Botre, F. Characterization of the phase I and phase II metabolic profile of tolvaptan by in vitro studies and liquid chromatography-mass spectrometry profiling: Relevance to doping control analysis. J. Pharm. Biomed. Anal. 2017, 145, 555–568. [Google Scholar] [CrossRef] [Green Version]
- Palermo, A.; Alessi, B.; Botre, F.; de la Torre, X.; Fiacco, I.; Mazzarino, M. In vitro evaluation of the effects of anti-fungals, benzodiazepines and non-steroidal anti-inflammatory drugs on the glucuronidation of 19-norandrosterone: Implications on doping control analysis. Drug Test. Anal. 2016, 8, 930–939. [Google Scholar] [CrossRef] [PubMed]
- Mazzarino, M.; de la Torre, X.; Fiacco, I.; Palermo, A.; Botre, F. Drug-drug interaction and doping, part 1: An in vitro study on the effect of non-prohibited drugs on the phase I metabolic profile of toremifene. Drug Test. Anal. 2014, 6, 482–491. [Google Scholar] [CrossRef]
- Palermo, A.; Botrè, F.; de la Torre, X.; Fiacco, I.; Iannone, M.; Mazzarino, M. Drug-drug interactions and masking effects in sport doping: Influence of miconazole administration on the urinary concentrations of endogenous anabolic steroids. Forensic Toxicol. 2016, 34, 386–397. [Google Scholar] [CrossRef]
- Zaitsu, K.; Miyawaki, I.; Bando, K.; Horie, H.; Shima, N.; Katagi, M.; Tatsuno, M.; Bamba, T.; Sato, T.; Ishii, A.; et al. Metabolic profiling of urine and blood plasma in rat models of drug addiction on the basis of morphine, methamphetamine, and cocaine-induced conditioned place preference. Anal. Bioanal. Chem. 2014, 406, 1339–1354. [Google Scholar] [CrossRef]
- Liu, R.; Cheng, J.; Yang, J.; Ding, X.; Yang, S.; Dong, F.; Guo, N.; Liu, S. GC-MS-based plasma metabolomic investigations of morphine dependent rats at different states of euphoria, tolerance and naloxone-precipitated withdrawal. Metab. Brain Dis. 2015, 30, 767–776. [Google Scholar] [CrossRef]
- Caspani, G.; Sebok, V.; Sultana, N.; Swann, J.R.; Bailey, A. Metabolic phenotyping of opioid and psychostimulant addiction: A novel approach for biomarker discovery and biochemical understanding of the disorder. Br. J. Pharmacol. 2022, 179, 1578–1606. [Google Scholar] [CrossRef] [PubMed]
- Kaddurah-Daouk, R.; Weinshilboum, R.M.; Pharmacometabolomics Research, N. Pharmacometabolomics: Implications for clinical pharmacology and systems pharmacology. Clin. Pharmacol. Ther. 2014, 95, 154–167. [Google Scholar] [CrossRef] [PubMed]
- Ma, R.; Kutchy, N.A.; Hu, G. Astrocyte-Derived Extracellular Vesicle-Mediated Activation of Primary Ciliary Signaling Contributes to the Development of Morphine Tolerance. Biol. Psychiatry 2021, 90, 575–585. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Meng, J.; Ban, Y.; Jalodia, R.; Chupikova, I.; Fernandez, I.; Brito, N.; Sharma, U.; Abreu, M.T.; Ramakrishnan, S.; et al. Morphine tolerance is attenuated in germfree mice and reversed by probiotics, implicating the role of gut microbiome. Proc. Natl. Acad. Sci. USA 2019, 116, 13523–13532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zachariou, V.; Georgescu, D.; Sanchez, N.; Rahman, Z.; DiLeone, R.; Berton, O.; Neve, R.L.; Sim-Selley, L.J.; Selley, D.E.; Gold, S.J.; et al. Essential role for RGS9 in opiate action. Proc. Natl. Acad. Sci. USA 2003, 100, 13656–13661. [Google Scholar] [CrossRef] [Green Version]
- Zachariou, V.; Bolanos, C.A.; Selley, D.E.; Theobald, D.; Cassidy, M.P.; Kelz, M.B.; Shaw-Lutchman, T.; Berton, O.; Sim-Selley, L.J.; Dileone, R.J.; et al. An essential role for DeltaFosB in the nucleus accumbens in morphine action. Nat. Neurosci. 2006, 9, 205–211. [Google Scholar] [CrossRef]
- Han, M.H.; Renthal, W.; Ring, R.H.; Rahman, Z.; Psifogeorgou, K.; Howland, D.; Birnbaum, S.; Young, K.; Neve, R.; Nestler, E.J.; et al. Brain region specific actions of regulator of G protein signaling 4 oppose morphine reward and dependence but promote analgesia. Biol. Psychiatry 2010, 67, 761–769. [Google Scholar] [CrossRef] [Green Version]
- Okerman, T.; Jurgenson, T.; Moore, M.; Klein, A.H. Inhibition of the phosphoinositide 3-kinase-AKT-cyclic GMP-c-Jun N-terminal kinase signaling pathway attenuates the development of morphine tolerance in a mouse model of neuropathic pain. Mol. Pain 2021, 17, 17448069211003375. [Google Scholar] [CrossRef]
- Rodgers, H.M.; Lim, S.A.; Yow, J.; Dinkins, M.L.; Patton, R.; Clemens, S.; Brewer, K.L. Dopamine D(1) or D(3) receptor modulators prevent morphine tolerance and reduce opioid withdrawal symptoms. Pharmacol. Biochem. Behav. 2020, 194, 172935. [Google Scholar] [CrossRef]
- Fisher, C.; Johnson, K.; Okerman, T.; Jurgenson, T.; Nickell, A.; Salo, E.; Moore, M.; Doucette, A.; Bjork, J.; Klein, A.H. Morphine Efficacy, Tolerance, and Hypersensitivity Are Altered After Modulation of SUR1 Subtype K(ATP) Channel Activity in Mice. Front. Neurosci. 2019, 13, 1122. [Google Scholar] [CrossRef]
- Shueb, S.S.; Erb, S.J.; Lunzer, M.M.; Speltz, R.; Harding-Rose, C.; Akgun, E.; Simone, D.A.; Portoghese, P.S. Targeting MOR-mGluR(5) heteromers reduces bone cancer pain by activating MOR and inhibiting mGluR5. Neuropharmacology 2019, 160, 107690. [Google Scholar] [CrossRef] [PubMed]
- Cataldo, G.; Erb, S.J.; Lunzer, M.M.; Luong, N.; Akgun, E.; Portoghese, P.S.; Olson, J.K.; Simone, D.A. The bivalent ligand MCC22 potently attenuates hyperalgesia in a mouse model of cisplatin-evoked neuropathic pain without tolerance or reward. Neuropharmacology 2019, 158, 107598. [Google Scholar] [CrossRef] [PubMed]
- Ninkovic, J.; Anand, V.; Dutta, R.; Zhang, L.; Saluja, A.; Meng, J.; Koodie, L.; Banerjee, S.; Roy, S. Differential effects of gram-positive and gram-negative bacterial products on morphine induced inhibition of phagocytosis. Sci. Rep. 2016, 6, 21094. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ninkovic, J.; Roy, S. Morphine decreases bacterial phagocytosis by inhibiting actin polymerization through cAMP-, Rac-1-, and p38 MAPK-dependent mechanisms. Am. J. Pathol. 2012, 180, 1068–1079. [Google Scholar] [CrossRef] [Green Version]
- Psifogeorgou, K.; Papakosta, P.; Russo, S.J.; Neve, R.L.; Kardassis, D.; Gold, S.J.; Zachariou, V. RGS9-2 is a negative modulator of mu-opioid receptor function. J. Neurochem. 2007, 103, 617–625. [Google Scholar] [CrossRef]
- Xu, J.; Faskowitz, A.J.; Rossi, G.C.; Xu, M.; Lu, Z.; Pan, Y.X.; Pasternak, G.W. Stabilization of morphine tolerance with long-term dosing: Association with selective upregulation of mu-opioid receptor splice variant mRNAs. Proc. Natl. Acad. Sci. USA 2015, 112, 279–284. [Google Scholar] [CrossRef] [Green Version]
- Pang, Z.; Zhou, G.; Ewald, J.; Chang, L.; Hacariz, O.; Basu, N.; Xia, J. Using MetaboAnalyst 5.0 for LC-HRMS spectra processing, multi-omics integration and covariate adjustment of global metabolomics data. Nat. Protoc. 2022, 17, 1735–1761. [Google Scholar] [CrossRef]
- Reimand, J.; Isserlin, R.; Voisin, V.; Kucera, M.; Tannus-Lopes, C.; Rostamianfar, A.; Wadi, L.; Meyer, M.; Wong, J.; Xu, C.; et al. Pathway enrichment analysis and visualization of omics data using g:Profiler, GSEA, Cytoscape and EnrichmentMap. Nat. Protoc. 2019, 14, 482–517. [Google Scholar] [CrossRef]
- Zhou, Y.; Zhou, B.; Pache, L.; Chang, M.; Khodabakhshi, A.H.; Tanaseichuk, O.; Benner, C.; Chanda, S.K. Metascape provides a biologist-oriented resource for the analysis of systems-level datasets. Nat. Commun. 2019, 10, 1523. [Google Scholar] [CrossRef]
- Zhang, Y.; Liang, F.; Zhang, D.; Qi, S.; Liu, Y. Metabolites as extracellular vesicle cargo in health, cancer, pleural effusion, and cardiovascular diseases: An emerging field of study to diagnostic and therapeutic purposes. Biomed. Pharmacother. 2023, 157, 114046. [Google Scholar] [CrossRef]
- Williams, C.; Palviainen, M.; Reichardt, N.C.; Siljander, P.R.; Falcon-Perez, J.M. Metabolomics Applied to the Study of Extracellular Vesicles. Metabolites 2019, 9, 276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chivero, E.T.; Dagur, R.S.; Peeples, E.S.; Sil, S.; Liao, K.; Ma, R.; Chen, L.; Gurumurthy, C.B.; Buch, S.; Hu, G. Biogenesis, physiological functions and potential applications of extracellular vesicles in substance use disorders. Cell Mol. Life Sci. 2021, 78, 4849–4865. [Google Scholar] [CrossRef] [PubMed]
- Hu, G.; Liao, K.; Niu, F.; Yang, L.; Dallon, B.W.; Callen, S.; Tian, C.; Shu, J.; Cui, J.; Sun, Z.; et al. Astrocyte EV-Induced lincRNA-Cox2 Regulates Microglial Phagocytosis: Implications for Morphine-Mediated Neurodegeneration. Mol. Ther. Nucleic Acids 2018, 13, 450–463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dinis-Oliveira, R.J. Metabolism and metabolomics of opiates: A long way of forensic implications to unravel. J. Forensic Leg. Med. 2019, 61, 128–140. [Google Scholar] [CrossRef]
- Meng, J.; Zhang, X.; Wu, H.; Bu, J.; Shi, C.; Deng, C.; Mao, Y. Morphine-induced conditioned place preference in mice: Metabolomic profiling of brain tissue to find "molecular switch" of drug abuse by gas chromatography/mass spectrometry. Anal. Chim. Acta 2012, 710, 125–130. [Google Scholar] [CrossRef]
- Hu, Z.; Deng, Y.; Hu, C.; Deng, P.; Bu, Q.; Yan, G.; Zhou, J.; Shao, X.; Zhao, J.; Li, Y.; et al. 1H NMR-based metabonomic analysis of brain in rats of morphine dependence and withdrawal intervention. Behav. Brain Res. 2012, 231, 11–19. [Google Scholar] [CrossRef]
- Deng, Y.; Bu, Q.; Hu, Z.; Deng, P.; Yan, G.; Duan, J.; Hu, C.; Zhou, J.; Shao, X.; Zhao, J.; et al. 1H-nuclear magnetic resonance-based metabonomic analysis of brain in rhesus monkeys with morphine treatment and withdrawal intervention. J. Neurosci. Res. 2012, 90, 2154–2162. [Google Scholar] [CrossRef]
- Chahkandi, M.; Askari, N.; Asadikaram, G. The Effect of Acute and Chronic Morphine on Some Blood Biochemical Parameters in an Inflammatory Condition in Gonadectomized Male Rats. Addict. Health 2015, 7, 130–139. [Google Scholar]
- Najafipour, H.; Beik, A. The Impact of Opium Consumption on Blood Glucose, Serum Lipids and Blood Pressure, and Related Mechanisms. Front. Physiol. 2016, 7, 436. [Google Scholar] [CrossRef] [Green Version]
- Radosevich, P.M.; Williams, P.E.; Lacy, D.B.; McRae, J.R.; Steiner, K.E.; Cherrington, A.D.; Lacy, W.W.; Abumrad, N.N. Effects of morphine on glucose homeostasis in the conscious dog. J. Clin. Invest. 1984, 74, 1473–1480. [Google Scholar] [CrossRef]
- Hu, G.; Yao, H.; Chaudhuri, A.D.; Duan, M.; Yelamanchili, S.V.; Wen, H.; Cheney, P.D.; Fox, H.S.; Buch, S. Exosome-mediated shuttling of microRNA-29 regulates HIV Tat and morphine-mediated neuronal dysfunction. Cell Death Dis. 2012, 3, e381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghosh, N.; Kesh, K.; Singh, P.K.; Sharma, U.; Chupikova, I.; Ramakrishnan, S.; Roy, S. Morphine use induces gastric microbial dysbiosis driving gastric inflammation through TLR2 signalling which is attenuated by proton pump inhibition. Br. J. Pharmacol. 2022. [Google Scholar] [CrossRef] [PubMed]
- Meng, J.; Abu, Y.F.; Zhang, Y.; Zhou, Y.; Xie, Y.; Yan, Y.; Tao, J.; Ramakrishnan, S.; Chen, C.; Roy, S. Opioid-induced microbial dysbiosis disrupts irinotecan (CPT-11) metabolism and increases gastrointestinal toxicity in a murine model. Br. J. Pharmacol. 2022. [Google Scholar] [CrossRef]
- Jalodia, R.; Kolli, U.; Braniff, R.G.; Tao, J.; Abu, Y.F.; Chupikova, I.; Moidunny, S.; Ramakrishnan, S.; Roy, S. Morphine mediated neutrophil infiltration in intestinal tissue play essential role in histological damage and microbial dysbiosis. Gut Microbes 2022, 14, 2143225. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, N.; Kesh, K.; Ramakrishnan, S.; Roy, S. Opioid Use in Murine Model Results in Severe Gastric Pathology that May Be Attenuated by Proton Pump Inhibition. Am. J. Pathol. 2022, 192, 1136–1150. [Google Scholar] [CrossRef] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kutchy, N.A.; Palermo, A.; Ma, R.; Li, Z.; Ulanov, A.; Callen, S.; Siuzdak, G.; Roy, S.; Buch, S.; Hu, G. Changes in Plasma Metabolic Signature upon Acute and Chronic Morphine Administration in Morphine-Tolerant Mice. Metabolites 2023, 13, 434. https://doi.org/10.3390/metabo13030434
Kutchy NA, Palermo A, Ma R, Li Z, Ulanov A, Callen S, Siuzdak G, Roy S, Buch S, Hu G. Changes in Plasma Metabolic Signature upon Acute and Chronic Morphine Administration in Morphine-Tolerant Mice. Metabolites. 2023; 13(3):434. https://doi.org/10.3390/metabo13030434
Chicago/Turabian StyleKutchy, Naseer A., Amelia Palermo, Rong Ma, Zhong Li, Alexandria Ulanov, Shannon Callen, Gary Siuzdak, Sabita Roy, Shilpa Buch, and Guoku Hu. 2023. "Changes in Plasma Metabolic Signature upon Acute and Chronic Morphine Administration in Morphine-Tolerant Mice" Metabolites 13, no. 3: 434. https://doi.org/10.3390/metabo13030434