


Glycerol Monolaurate Alleviates Oxidative Stress and Intestinal Flora Imbalance Caused by Salinity Changes for Juvenile Grouper
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Diets Preparation
2.2. Experimental Design
2.3. Sample Collection
2.4. Liver Antioxidant Index
2.5. Real-Time Quantitative RT-PCR Analysis of Gene Expression
2.6. Intestinal Flora Analysis
2.6.1. Extraction of Genomic DNA
2.6.2. PCR Amplification of Target Regions
2.6.3. PCR Product Purification, Library Construction and Sequencing
2.6.4. Data Analysis and Functional Prediction
2.7. Statistical Analysis
3. Results
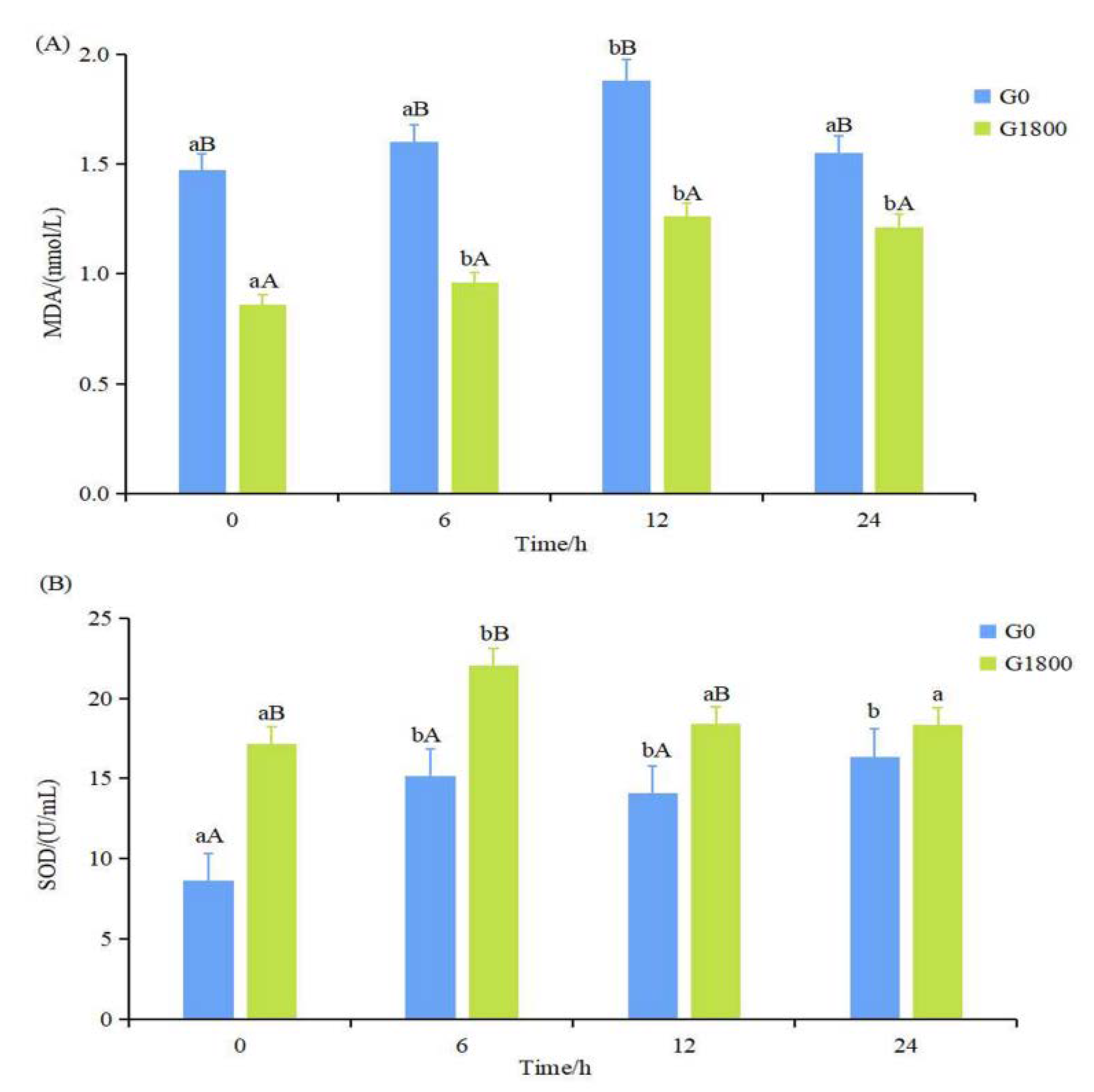
3.1. Effects of GML on Antioxidant Enzyme Activities Caused by Salinity Changes for Grouper
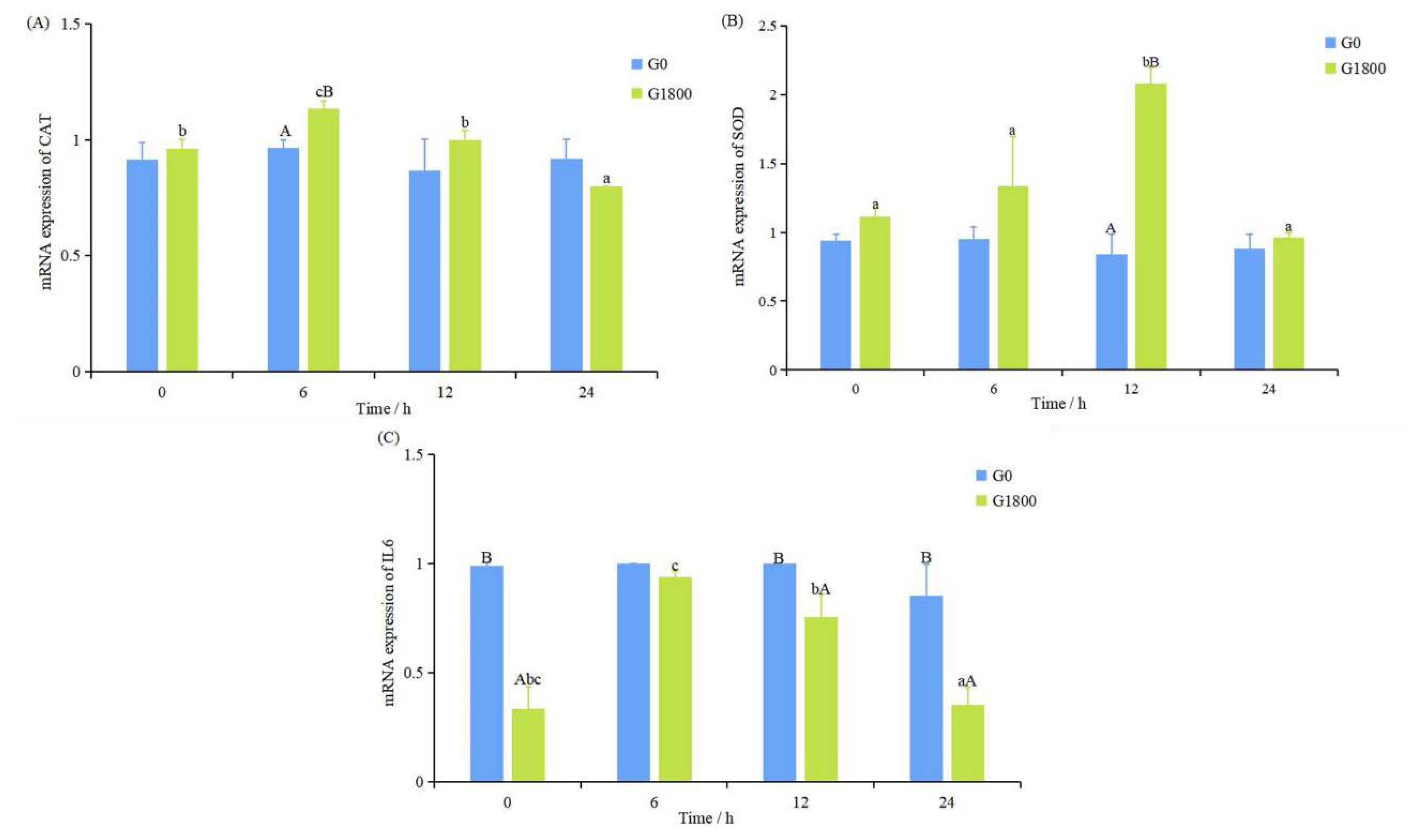
3.2. Effects of GML on Gene Expression Caused by Salinity Changes in Grouper
3.3. Effects of GML on Intestinal Flora Caused by Salinity Changes in Grouper
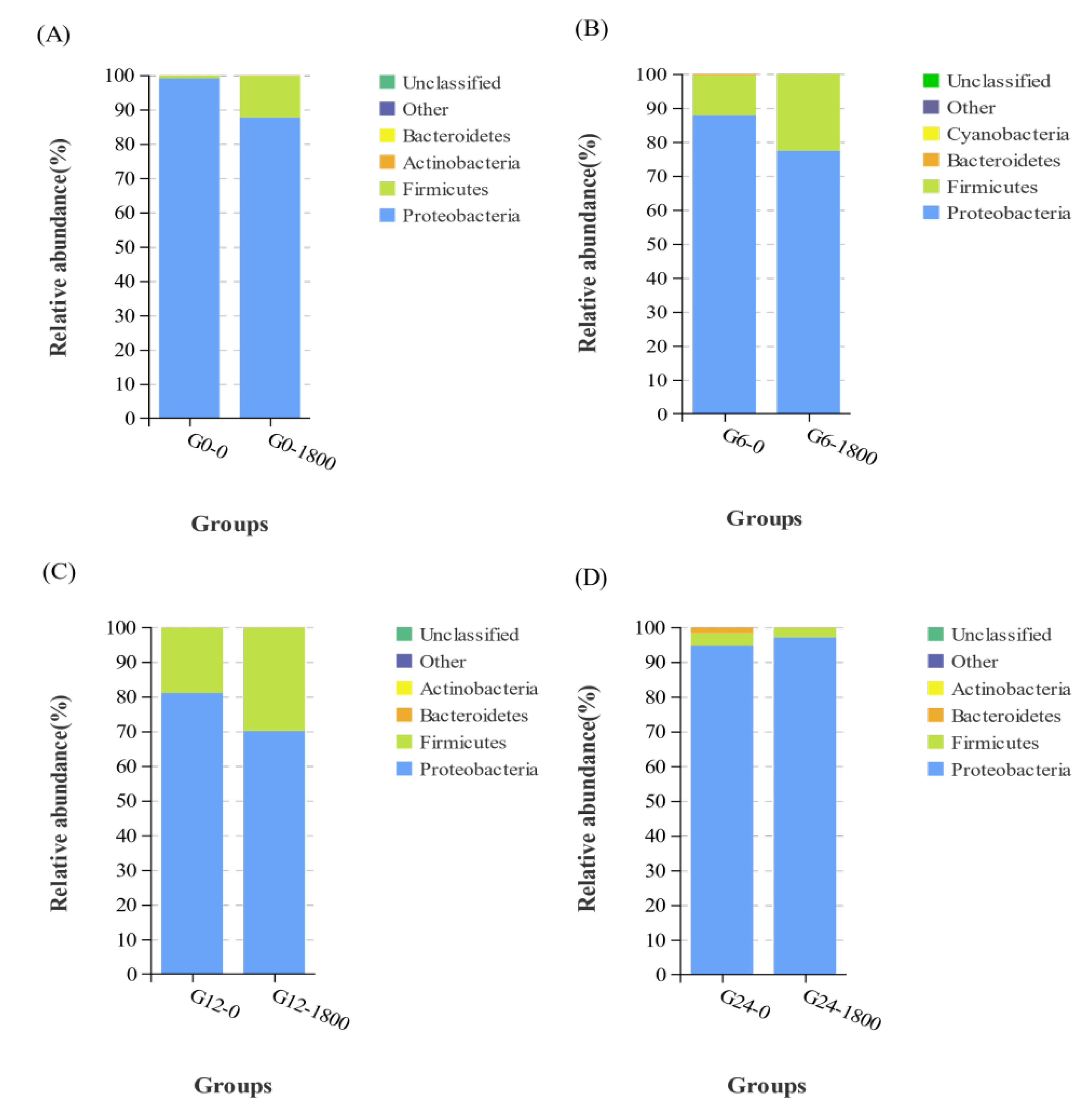
3.3.1. Effects of GML on Relative Abundance at the Phylum Level Caused by Salinity Changes in Grouper
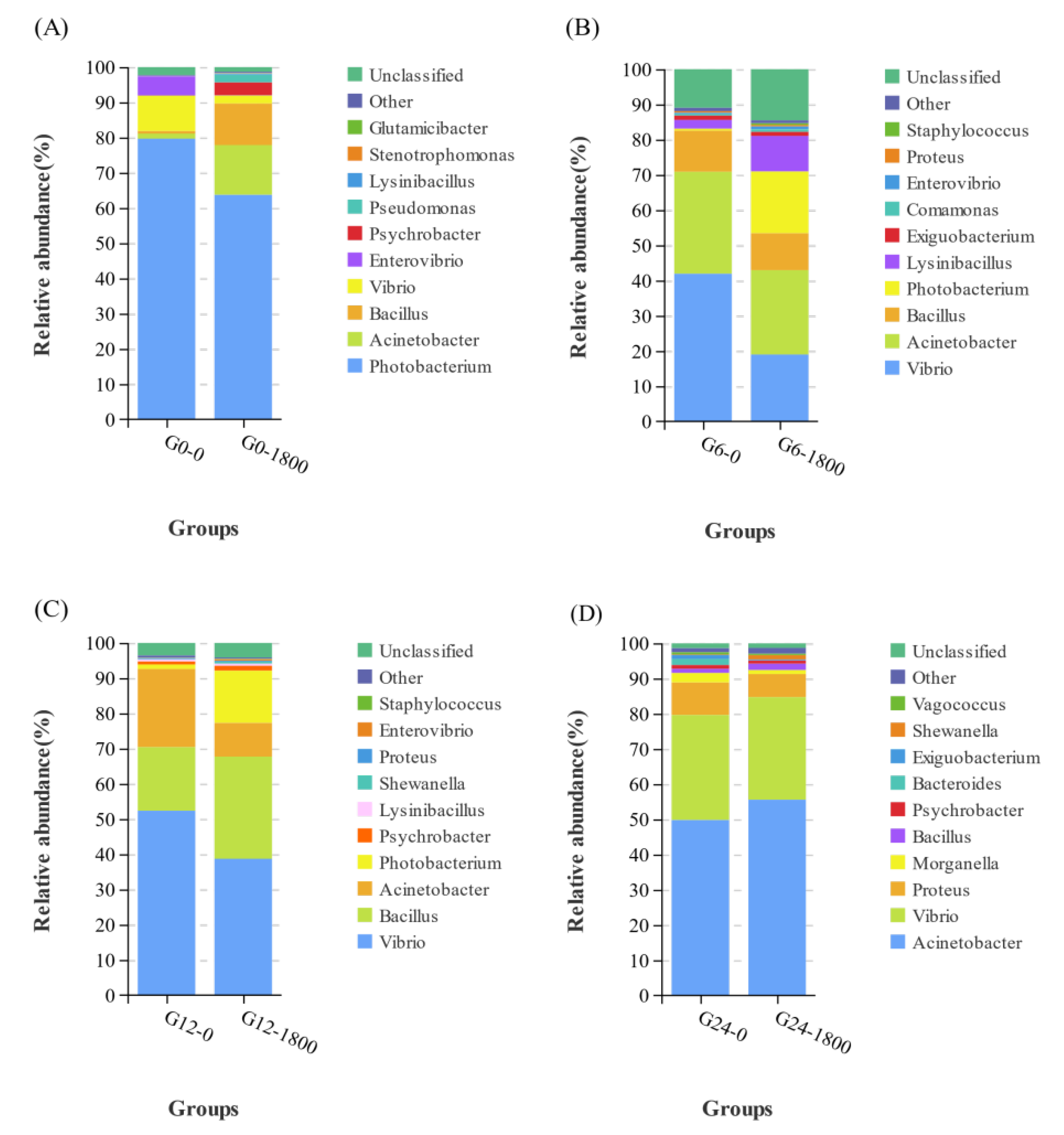
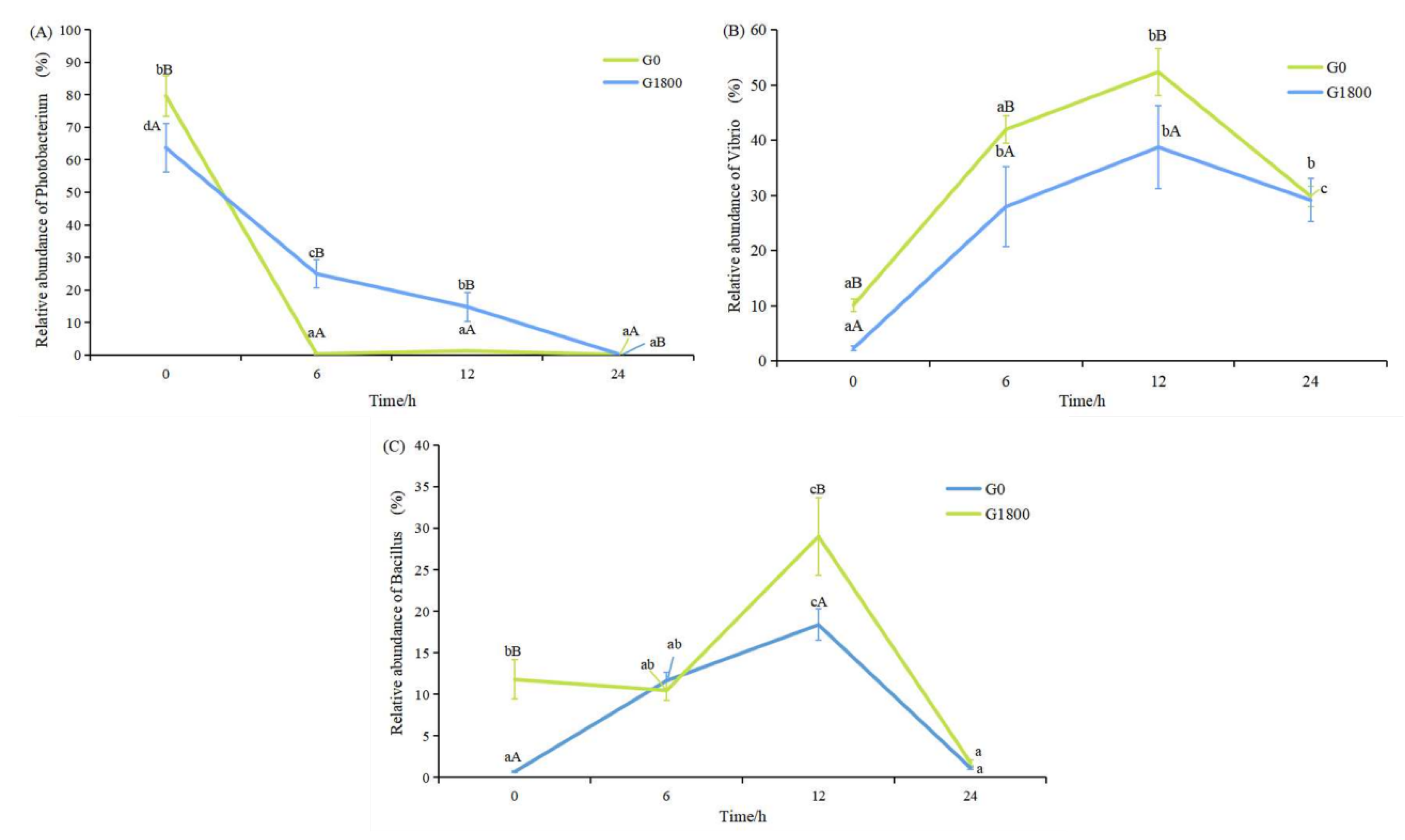
3.3.2. Effects of GML on Relative Abundance at the Genus Level Caused by Salinity Changes in Grouper
3.3.3. Effects of GML on Indicator Species Caused by Salinity Changes in Grouper
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Guo, Z.X.; Zeng, Z.Q.; Huang, J.S.; Wang, W.Z.; Li, H.J.; Chen, G. Effects of acute hypoxia on oxidative stress, energy utilization and carbohydrate metabolism in liver of large-sized juvenile cobia (Rachycentron canadum). J. Guangdong Ocean. Univ. 2020, 40, 134–140. [Google Scholar]
- Hou, S.; Jin, Z.; Jiang, W.; Chi, L.; Xia, B.; Chen, J. Physiological and immunological responses of sea cucumber Apostichopus japonicus during desiccation and subsequent resubmersion. PeerJ 2019, 7, e7427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tuerxun, M.; Muhda, A.; Yin, L. The molecular mechanisms of signal pathway activating effect of E2F-1/NF-kappaB/GSK-3beta on cognitive dysfunction of alzheimer rats. Bioengineered 2021, 12, 10000–10008. [Google Scholar] [CrossRef]
- Famurewa, A.C.; Aja, P.M.; Maduagwuna, E.K.; Ekeleme-Egedigwe, C.A.; Ufebe, O.G.; Azubuike-Osu, S.O. Antioxidant and anti-inflammatory effects of virgin coconut oil supplementation abrogate acute chemotherapy oxidative nephrotoxicity induced by anticancer drug methotrexate in rats. Biomed. Pharmacother. 2017, 96, 905–911. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Zhou, Q.L.; Liang, H.L.; Sun, C.X.; Zheng, X.C.; Lin, Y.; Ren, M.C.; Zhu, L.; Ge, X.P. Advances in aquatic animal nutrition and immunity research. J. Fish. China 2022, 46, 1761–1775. [Google Scholar]
- Zhang, C.; Zhang, Q.; Song, X.; Pang, Y.; Song, Y.; Cheng, Y.; Yang, X. Dietary L-tryptophan modulates the hematological immune and antibacterial ability of the chinese mitten crab, Eriocheir sinensis, under cheliped autotomy stress. Front. Immunol. 2018, 9, 2744. [Google Scholar] [CrossRef]
- Higgs, D.A.; McBride, J.R.; Markert, J.R.; Dosanjh, B.S.; Plotnikoff, M.D.; Clarke, W.C. Evaluation of tower and candle rapeseed (canola) meal and bronowski rapeseed protein concentrate as protein supplements in practical dry diets for juvenile chinook salmon (Oncorhynchus tshawytscha). Aquaculture 1982, 29, 1–31. [Google Scholar] [CrossRef]
- Kong, L.; Wang, Z.; Xiao, C.; Zhu, Q.; Song, Z. Glycerol monolaurate attenuated immunological stress and intestinal mucosal injury by regulating the gut microbiota and activating AMPK/Nrf2 signaling pathway in lipopolysaccharide-challenged broilers. Anim. Nutr. 2022, 10, 347–359. [Google Scholar] [CrossRef]
- Wang, Y.; Abdullah; Zhong, H.; Wang, J.; Feng, F. Dietary glycerol monolaurate improved the growth, activity of digestive enzymes and gut microbiota in zebrafish (Danio rerio). Aquacult. Rep. 2021, 20, 100670. [Google Scholar] [CrossRef]
- Wang, J.; Jiang, H.; Alhamoud, Y.; Chen, Y.; Zhuang, J.; Liu, T.; Cai, L.; Shen, W.; Wu, X.; Zheng, W.; et al. Integrated metabolomic and gene expression analyses to study the effects of glycerol monolaurate on flesh quality in large yellow croaker (Larimichthys crocea). Food Chem. 2022, 367, 130749. [Google Scholar] [CrossRef]
- Sun, C.Y.; Dong, H.B.; Wang, W.H.; Gu, Q.H.; Duan, Y.F.; Zhang, J.S.; Xu, X.D. Effects of glycerol monolaurate on lipid metabolism of Lateolabrax maculatus. South China Fish. Sci. 2021, 17, 67–75. [Google Scholar]
- Wang, Y.; Abdullah; Zhang, C.; Li, Y.; Feng, F. Effects of dietary glycerol monolaurate on the growth performance, digestive enzymes, body composition and non-specific immune response of white shrimp (Litopenaeus vannamei). Aquacult. Rep. 2020, 18, 100535. [Google Scholar] [CrossRef]
- Wang, Y.C.; Du, J.; Li, Y.; Zhang, H.; Feng, F.Q. Effects of glycerol monolaurate on growth, health and nutritional quality of Chinese soft-shelled turtle ( Pelodiscus sinensis). Chin. J. Anim. Nutr. 2019, 31, 428–436. [Google Scholar]
- Yan, X.; Li, Z.; Dong, X.; Tan, B.; Pan, S.; Li, T.; Long, S.; Huang, W.; Suo, X.; Yang, Y. Degradation of Muscle Quality in Hybrid Grouper (female symbol Epinephelus fuscoguttatus x male symbol Epinephelus lanceolatus) due to oxidative damage caused by ingestion of oxidized fish oil. Front. Nutr. 2022, 9, 840535. [Google Scholar] [CrossRef] [PubMed]
- Ministry of Agriculture and Rural Affairs of the Peoples Republic of China; National Fisheries Technology Extension Center; Chinese Society of Fisheries. China Fisheries Statistical Yearbook; China Agricultural Publishing House: Beijing, China, 2022; p. 42. [Google Scholar]
- Wang., D.P.; Cao., Z.W.; Xie., D.X.; Gan., X. Research progress in Epinephelus industry. J. South. Agric. 2018, 43, 1058–1065. [Google Scholar]
- He, Y.; Guo, X.; Tan, B.; Dong, X.; Yang, Q.; Liu, H.; Zhang, S.; Chi, S. Replacing fish meal with fermented rice protein in diets for hybrid groupers (Epinephelus fuscoguttatus♀× Epinephelus lanceolatus♂): Effects on growth, digestive and absorption capacities, inflammatory-related gene expression, and intestinal microbiota. Aquacult. Rep. 2021, 19, 100603. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Chen, F.J.; Fu, Y.S.; Ling, X.D.; Chang, L.; Li, X.Y. Effects of hypoxia stress on activities of mitochondrial respiratory chain complexes and antioxidant enzyme in muscle of lake Qinghai Scaleless Carp. J. Guangdong Ocean. Univ. 2021, 41, 118–124. [Google Scholar]
- He, G.; Dong, C.; Luan, Z.; McAllan, B.M.; Xu, T.; Zhao, L.; Qiao, J. Oxygen free radical involvement in acute lung injury induced by H5N1 virus in mice. Influenza Other Respir. Viruses 2013, 7, 945–953. [Google Scholar] [CrossRef] [Green Version]
- Fu, C.; Cui, Z.; Shi, X.; Liu, J.; Jiang, Y.; Zhang, R. Effects of dietary glyceryl monolaurate supplementation on growth performance, non-specific immunity, antioxidant status and intestinal microflora of Chinese mitten crabs. Fish Shellfish Immunol. 2022, 125, 65–73. [Google Scholar] [CrossRef]
- Lin, H.Z.; Gao, J.L.; Liang, Z.Y.; Fan, X.P.; Lin, H.S.; Cao, W.H.; Huang, Y.P.; Qin, X.M. Effect of cold stress methods on antioxidant and energy metabolism of Crassostrea gigas in water-free live storage period. J. Guangdong Ocean. Univ. 2022, 42, 95–103. [Google Scholar]
- Jiang, K.; Cao, S.; Cui, J.Z.; Matsubara, J.A. Immuno-modulatory Effect of IFN-gamma in AMD and its role as a possible target for therapy. J. Clin. Exp. Ophthalmol. 2013, Suppl. 2, 0071–0076. [Google Scholar] [CrossRef] [Green Version]
- Zhao, M.; Jiang, Z.; Cai, H.; Li, Y.; Feng, F. Modulation of the gut microbiota during high-dose glycerol monolaurate-mediated mmelioration of obesity in mice fed a high-fat Diet. mBio 2020, 11, e00190-20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, H.; Li, D.; Guan, X.; Yang, Y.; Yan, J.; Shi, J.; Ma, R.; Shu, Q. MsrA suppresses inflammatory ictivation of microglia and oxidative stress to prevent demyelination via inhibition of the NOX2-MAPKs/NF-kappaB signaling pathway. Drug Des. Dev. Ther. 2020, 14, 1377–1389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Y.; Tang, Z.; Nan, X.; Sun, F.; Xiong, B. Effects of Lonicera japonica extract on performance, blood biomarkers of inflammation and oxidative stress during perinatal period in dairy cows. Asian-Australas. J. Anim. Sci. 2019, 33, 1096. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Federico, A.; Morgillo, F.; Tuccillo, C.; Ciardiello, F.; Loguercio, C. Chronic inflammation and oxidative stress in human carcinogenesis. Int. J. Cancer 2010, 121, 2381–2386. [Google Scholar] [CrossRef] [PubMed]
- Rajkumar, K.; Bhattacharya, A.; David, S.; Balaji, S.H.; Hariharan, R.; Jayakumar, M.; Balaji, N. Socio-demographic study on extent of knowledge, awareness, attitude, and risks of zoonotic diseases among livestock owners in Puducherry region. Vet. World 2016, 9, 1018–1024. [Google Scholar] [CrossRef] [PubMed]
- Luo, X.; Zheng, Y.; Wen, R.; Deng, X.; Zhou, L.; Liao, H. Effects of ceftriaxone induced intestinal dysbacteriosis on lymphocytes in different tissues in mice. Immunobiology 2016, 221, 994–1000. [Google Scholar] [CrossRef]
- Tilocca, B.; Burbach, K.; Heyer, C.M.E.; Hoelzle, L.E.; Mosenthin, R.; Stefanski, V.; Camarinha-Silva, A.; Seifert, J. Dietary changes in nutritional studies shape the structural and functional composition of the pigs’ fecal microbiome-from days to weeks. Microbiome 2017, 5, 144. [Google Scholar] [CrossRef] [Green Version]
- Alkushi, A.G.; Abdelfattah-Hassan, A.; Eldoumani, H.; Elazab, S.T.; Mohamed, S.A.M.; Metwally, A.S.; El-Shetry, E.S.; Saleh, A.A.; ElSawy, N.A.; Ibrahim, D. Probiotics-loaded nanoparticles attenuated colon inflammation, oxidative stress, and apoptosis in colitis. Sci. Rep. 2022, 12, 5116. [Google Scholar] [CrossRef]
- Deng, Y.; Zhang, Y.; Chen, H.; Xu, L.; Wang, Q.; Feng, J. Gut-liver immune response and gut microbiota profiling reveal the pathogenic mechanisms of vibrio harveyi in pearl gentian grouper (Epinephelus lanceolatusmale symbol x E. fuscoguttatus female symbol). Front. Immunol. 2020, 11, 607754. [Google Scholar] [CrossRef] [PubMed]
- Kolba, N.; Zarei, A.; Cheng, J.; Agarwal, N.; Dadmohammadi, Y.; Khazdooz, L.; Abbaspourrad, A.; Tako, E. Alterations in intestinal brush border membrane functionality and bacterial populations following intra-amniotic administration (Gallus gallus) of nicotinamide riboside and its derivatives. Nutrients 2022, 14, 3130. [Google Scholar] [CrossRef] [PubMed]
- Kaszab, E.; Farkas, M.; Rado, J.; Micsinai, A.; Nyiro-Fekete, B.; Szabo, I.; Kriszt, B.; Urbanyi, B.; Szoboszlay, S. Novel members of bacterial community during a short-term chilled storage of common carp (Cyprinus carpio). Folia Microbiol. 2022, 67, 299–310. [Google Scholar] [CrossRef] [PubMed]
- Hu, L.; Yun, B.; Xue, M.; Wang, J.; Wu, X.; Zheng, Y.; Han, F. Effects of fish meal quality and fish meal substitution by animal protein blend on growth performance, flesh quality and liver histology of Japanese seabass (Lateolabrax japonicus). Aquaculture 2013, 372, 52–61. [Google Scholar] [CrossRef]
- Li, W.; Pan, X.; Cheng, W.; Cheng, Y.; Yin, Y.; Chen, J.; Xu, G.; Xie, L. Serum biochemistry, histology and transcriptomic profile analysis reflect liver inflammation and damage following dietary histamine supplementation in yellow catfish (Pelteobagrus fulvidraco). Fish Shellfish Immunol. 2018, 77, 83–90. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Estes, J.D.; Schlievert, P.M.; Duan, L.; Brosnahan, A.J.; Southern, P.J.; Reilly, C.S.; Peterson, M.L.; Schultz-Darken, N.; Brunner, K.G.; et al. Glycerol monolaurate prevents mucosal SIV transmission. Nature 2009, 458, 1034–1038. [Google Scholar] [CrossRef] [Green Version]
- Kalatzis, P.G.; Castillo, D.; Katharios, P.; Middelboe, M. Bacteriophage Interactions with Marine Pathogenic Vibrios: Implications for Phage Therapy. Antibiotics 2018, 7, 15. [Google Scholar] [CrossRef]
- Takemura, A.F.; Chien, D.M.; Polz, M.F. Associations and dynamics of Vibrionaceae in the environment, from the genus to the population level. Front. Microbiol. 2014, 5, 38. [Google Scholar] [CrossRef] [Green Version]
- Froelich, B.; Bowen, J.; Gonzalez, R.; Snedeker, A.; Noble, R. Mechanistic and statistical models of total Vibrio abundance in the Neuse River Estuary. Water Res. 2013, 47, 5783–5793. [Google Scholar] [CrossRef]
- Oberbeckmann, S.; Fuchs, B.M.; Meiners, M.; Wichels, A.; Wiltshire, K.H.; Gerdts, G. Seasonal dynamics and modeling of a Vibrio community in coastal waters of the North Sea. Microb. Ecol. 2012, 63, 543–551. [Google Scholar] [CrossRef]
- Randa, M.A.; Polz, M.F.; Lim, E. Effects of temperature and salinity on Vibrio vulnificus population dynamics as assessed by quantitative PCR. Appl. Environ. Microbiol. 2004, 70, 5469–5476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buruiană, C.T.; Profir, A.G.; Vizireanu, C. Effects of probiotic Bacillus species in aquaculture—An overview. Food Technol. 2014, 38, 9–17. [Google Scholar]
- Cha, J.; Rahimnejad, S.; Yang, S.; Kim, K.; Lee, K. Evaluations of Bacillus spp. as dietary additives on growth performance, innate immunity and disease resistance of olive flounder (Paralichthys olivaceus) against Streptococcus iniae and as water additives. Aquaculture 2013, 402, 50–57. [Google Scholar] [CrossRef]
- Kuebutornye, F.K.A.; Abarike, E.D.; Lu, Y. A review on the application of Bacillus as probiotics in aquaculture. Fish Shellfish Immunol. 2019, 87, 820–828. [Google Scholar] [CrossRef]
- Nayak, S.K. Probiotics and immunity: A fish perspective. Fish Shellfish Immunol. 2010, 29, 2–14. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredients | G0 | G1800 |
|---|---|---|
| Brown fish meal | 30.00 | 30.00 |
| Soybean meal | 18.00 | 18.00 |
| Corn gluten meal | 9.50 | 9.50 |
| Peanut meal | 10.00 | 10.00 |
| Chicken powder | 6.00 | 6.00 |
| Wheat flour | 18.59 | 18.59 |
| Fish oil | 2.00 | 2.00 |
| Soybean oil | 2.00 | 2.00 |
| Soybean lecithin | 1.00 | 1.00 |
| Ca(H2PO4)2 | 1.00 | 1.00 |
| Premix | 1.00 | 1.00 |
| Choline chloride | 0.50 | 0.50 |
| Vitamin C | 0.05 | 0.05 |
| Microcrystalline cellulose | 0.36 | 0.18 |
| GML | 0.00 | 0.18 |
| Total | 100.00 | 100.00 |
| Nutrient levels | ||
| Crude protein | 49.74 | 49.66 |
| Crude lipid | 9.42 | 9.37 |
| Genes | Nucleotide Sequence (5′-3) | Genbank Accession No. |
|---|---|---|
| β-actin | F: GATCTGGCATCACACCTTCT R: CATCTTCTCCCTGTTGGCTT | AY510710.2 |
| SOD | F: GTTGGAGACCTGGGAAATGTGACTG R: CCATTGAGGGTGAGCATCTTGTCC | AY735008.1 |
| CAT | F:GCTCTATCCGCTCCTCTTCTCCTC R: GTAGTTCCTGACGACGGTGATGTG | KT884509.1 |
| IL6 | F: CAATCCCAGCACCTTCCAC R: CCTGACAGCCAGACTTCCTCT | AFE62919.1 |
| Itmes | G0-0 | G0-1800 | G6-0 | G6-1800 | G12-0 | G12-1800 | G24-0 | G24-1800 |
|---|---|---|---|---|---|---|---|---|
| Proteobacteria | 99.072 | 87.608 | 87.832 | 77.375 | 80.998 | 70.044 | 94.666 | 97.028 |
| Firmicutes | 0.794 | 12.205 | 11.683 | 22.513 | 18.841 | 29.836 | 3.524 | 2.908 |
| Actinobacteria | 0.050 | 0.133 | 0.004 | 0.003 | 0.011 | 0.009 | 0.015 | 0.011 |
| Bacteroidetes | 0.052 | 0.029 | 0.403 | 0.083 | 0.112 | 0.086 | 1.749 | 0.027 |
| Cyanobacteria | 0.011 | 0.004 | 0.056 | 0.000 | 0.005 | 0.002 | 0.010 | 0.000 |
| Others | 0.020 | 0.019 | 0.007 | 0.008 | 0.000 | 0.000 | 0.001 | 0.000 |
| Unclassified | 0.002 | 0.002 | 0.016 | 0.017 | 0.033 | 0.022 | 0.035 | 0.026 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, X.; Zhu, D.; Mao, M.; Wu, J.; Yang, Q.; Tan, B.; Chi, S. Glycerol Monolaurate Alleviates Oxidative Stress and Intestinal Flora Imbalance Caused by Salinity Changes for Juvenile Grouper. Metabolites 2022, 12, 1268. https://doi.org/10.3390/metabo12121268
Li X, Zhu D, Mao M, Wu J, Yang Q, Tan B, Chi S. Glycerol Monolaurate Alleviates Oxidative Stress and Intestinal Flora Imbalance Caused by Salinity Changes for Juvenile Grouper. Metabolites. 2022; 12(12):1268. https://doi.org/10.3390/metabo12121268
Chicago/Turabian StyleLi, Xuehe, Dongwenjun Zhu, Minling Mao, Jianwei Wu, Qihui Yang, Beiping Tan, and Shuyan Chi. 2022. "Glycerol Monolaurate Alleviates Oxidative Stress and Intestinal Flora Imbalance Caused by Salinity Changes for Juvenile Grouper" Metabolites 12, no. 12: 1268. https://doi.org/10.3390/metabo12121268