1. Introduction
Protein accounts for a significant portion of the cost of a formula diet. In addition to protein, carbohydrates and lipids are key nutrients in fish that have a significant function in the growth, development and immunity of fish [
1,
2]. Like other all animals, fish depend on energy from their foodstuff to fulfil their normal growth requirements. However, in contrast to other land animals, particularly when there is not enough energy in the diet, fish obtain their total energy requirements by the breakdown of proteins into amino acids [
3]. With the development of intensive farming and the soaring price of fishmeal, research on the substitution and cost-effectiveness of protein in feed has been continuously conducted. A number of studies have demonstrated that an appropriate CHO:L ratio in feed can improve growth, feed efficiency, enzyme activity, protein conservation and reduce nitrogen and ammonia excretion, promoting an improved farming environment [
4,
5]. Most fish make good use of lipids, but at high dietary levels, they may lead to fatty liver in fish [
6]; similarly, the amount of lipids added is also detrimental to the lipid deposition of fish, which may affect the organoleptic characteristics of the fillet, such as its flavor [
7,
8]. On the one hand, carnivorous fish have a lower carbohydrate utilization than omnivorous and herbivorous fish. On the other hand, carbohydrates are more readily available and cheaper relative to lipids, but excess addition of carbohydrates in feed may inhibit their growth and increase glycogen accumulation in the liver [
9,
10]. Therefore, maintaining an optimum carbohydrate and lipid (CHO/L) balance will be beneficial for fish growth performance, feed utilization, nutrient absorption, and deposition.
The secretion of certain specific digestive enzymes in fish is so low that many nutrients in plant materials cannot be easily digested [
11]. A trend towards increased energy and nitrogen-free leachate digestibility was observed with exogenous enzyme supplementation. The addition of glycosidase complexes to rapeseed meal feeds also significantly increased feed digestibility and improve the growth performance of spot prawns
(Pandalus platyceros) [
12]. It is generally accepted that an important role in glucose-sensitive mechanisms is played by
glut2, and that it is expressed in the sinusoidal membrane of hepatocytes, which execute bidirectional glucose transport in response to dietary and hormonal conditions [
13]. It has been previously shown that
glut2 expression in the liver of Atlantic cod (
Gadus morhua) diminished during starvation and was boosted on refeeding; thus,
glut2 is thought to reflect the transport of glucose once the glycogen has been depleted [
14]. At physiological conditions, the glucose transport rate of
glut2 varies with glucose concentration even under diabetic conditions [
15]. Earlier studies reported that glycosylase supplementation in a high-carbohydrate diet significantly promoted the expression of
glut2 gene in liver and intestine [
16]. From the current research situation, different CHO:L ratios have been studied in many species of fish [
1,
2], but there are few studies on how to mitigate the adverse effects on high CHO:L ratio. In our research, we investigated the effects on
glut2 gene expression at different CHO:L ratios and the supplementation of carbohydrase at different CHO:L ratios.
Hybrid grouper (
Epinephelus fuscoguttatus♀ ×
E. lanceolatus♂) possesses the advantages of excellent growth, high illness resistance and commercial value, and represents a typical representative of a carnivorous fish. More recently, it has emerged as the dominant species of grouper farmed in southeast China [
17]. In order to maximize profits, diets are often formulated to contain high levels of CHO:L ratio to reduce feed costs. Although the optimal glycolipid ratio requirements at the juvenile stage of grouper have been studied [
18,
19,
20], there is little research on the addition of carbohydrase based on different glycolipid ratios. With this in mind, the purpose of this study was to assess the influence of carbohydrase supplementation on growth performance, carbohydrate metabolic enzyme and
glut2 gene expression in the Hybrid Grouper-fed diets with different CHO:L ratios.
4. Discussion
Carbohydrates are considered to provide energy at a low cost, while lipids are essential for animal growth and development, and the provision of an appropriate amount of CHO: L ratios in the diet can have beneficial effect on animal growth and spare the utilization of fishmeal protein [
2,
26,
27,
28]. With the scarcity of fishmeal resources, the impact of different levels of dietary CHO: L ratios on growth have been assessed in a number of aquaculture species with varying results, such as grass carp (
Ctenopharyngodon idellus) [
5], Nile tilapia (
Oreochromis niloticus) [
2], juvenile yellow catfish (
Pelteobagrus fulvidraco) [
26], and juvenile Senegalese sole (
Solea senegalensis, Kaup) [
4]. In recent years, exogenous enzymes have been extensively studied in livestock and poultry as functional additives to improve the nutritional digestibility of plant-based raw materials. Therefore, in the present study, we not only investigated the impact of CHO: L ratios, but also explored the impact of carbohydrase. Our results showed that fish fed CHO:L ratio of 1.92 achieved the best growth performance compared to fish fed CHO:L ratios of 0.91 and 3.91. Previous studies on groupers [
18,
29] have also shown that the best growth was achieved at medium CHO:L ratio, which was similar to our results. A significant increase in WG and SGR and a significant decrease in FCR were observed with carbohydrase supplementation at a CHO:L ratio of 3.91, indicating that carbohydrase act as a growth promoter. In studies on Atlantic salmon (
Salmo salar) [
30], tilapia [
31], and large yellow croaker (
Larimichthys crocea) [
28], it was found that appropriate CHO: L ratios improved growth performance and feed digestibility, but higher CHO: L ratio conditions were found to limit carbohydrate absorption in various fish species. The ability of carnivorous fish to utilize carbohydrates in their diet was known to be relatively low compared with grass carp and tilapia, and also to lead to different effects. Hybrid grouper was able to tolerate 28% of dietary starch with no negative impact on growth performance [
32]. The experimental group with a CHO:L ratio of 3.91 contained 33% starch, which may be responsible for the poorer growth of the groupers. A report on rohu (
Labeo rohita) showed a significant improvement in growth and protein utilization after 50 mg/kg α-amylase supplementation in non-gelatinized corn diet [
33]. Amylase is known to hydrolyze large α-linked polysaccharides (such as starch, glycogen, branched-chain starch), while glycosylase and pullulanase are exogenous enzymes that can hydrolyze the glycoside bonds of type α-1,4 and α-1,6, releasing D-glucose by continuously consuming the glucose unit at the non-reduction end of the sugar chain [
34]. Protein-sparing effects and protein-carbohydrate hydrolysis may occur when exogenous enzymes supplemented to the diets [
35]. This may account for better growth performance of grouper-fed carbohydrase supplementation at a CHO:L ratio of 3.91.
There were no significant differences in moisture, ash, and crude protein content between all treatment groups. In contrast, crude lipid decreased significantly with the increase in dietary CHO:L ratio, which was consistent with previous reported on large yellow croaker [
28], hybrid Clarias catfish (
Clarias macrocephalus × C. gariepinus) [
36], and juvenile hybrid grouper [
19]. When nutritional requirements are fulfilled, the appropriate lipid content in the diet is about 10% for hybrid grouper [
37]. Higher lipid content in the diet, which cannot be fully utilized by grouper and tended to cause lipid deposition in different tissues, whereas the amount of lipid content obtained from carbohydrate was probably quite limited. This result confirms a positive correlation between lipid deposition in fish and the lipid content of the diet [
28,
38,
39]. As we all know, the levels of triglycerides, cholesterol, glucose and total protein in blood are generally correlated with animal health and indicate the metabolic and physiological state of the organism, and too high or too low in blood will cause aberrant changes in the body [
40]. It is well-known that changes in tissue cholesterol concentrations depend on the nutritional status of the fish [
41]. Among the plasma biochemical indicators of our research, a significant decrease in TG and TC concentration was observed with the decrease in dietary lipid level, which suggested that endogenous lipid transport is more dynamic in response to higher lipid in the diet. This indicates that lipid transport in the blood is quite efficient in the presence of high lipids, and the results of this study are in consistency with earlier research reported on juvenile yellow catfish [
2,
5]. Contrastingly, plasma insulin and glucose increased with the increase in dietary CHO:L ratio. Long-term adaptation to high carbohydrates can increase plasma insulin concentrations in fish [
42]. In mammals, promoting glucose uptake by the liver and skeletal muscle, increasing glucose uptake and fatty acid influx are the most prominent functions of insulin [
43,
44]. In fact, most dietary carbohydrate is then excreted after digestion, either stored as glycogen in the body or converted to fat [
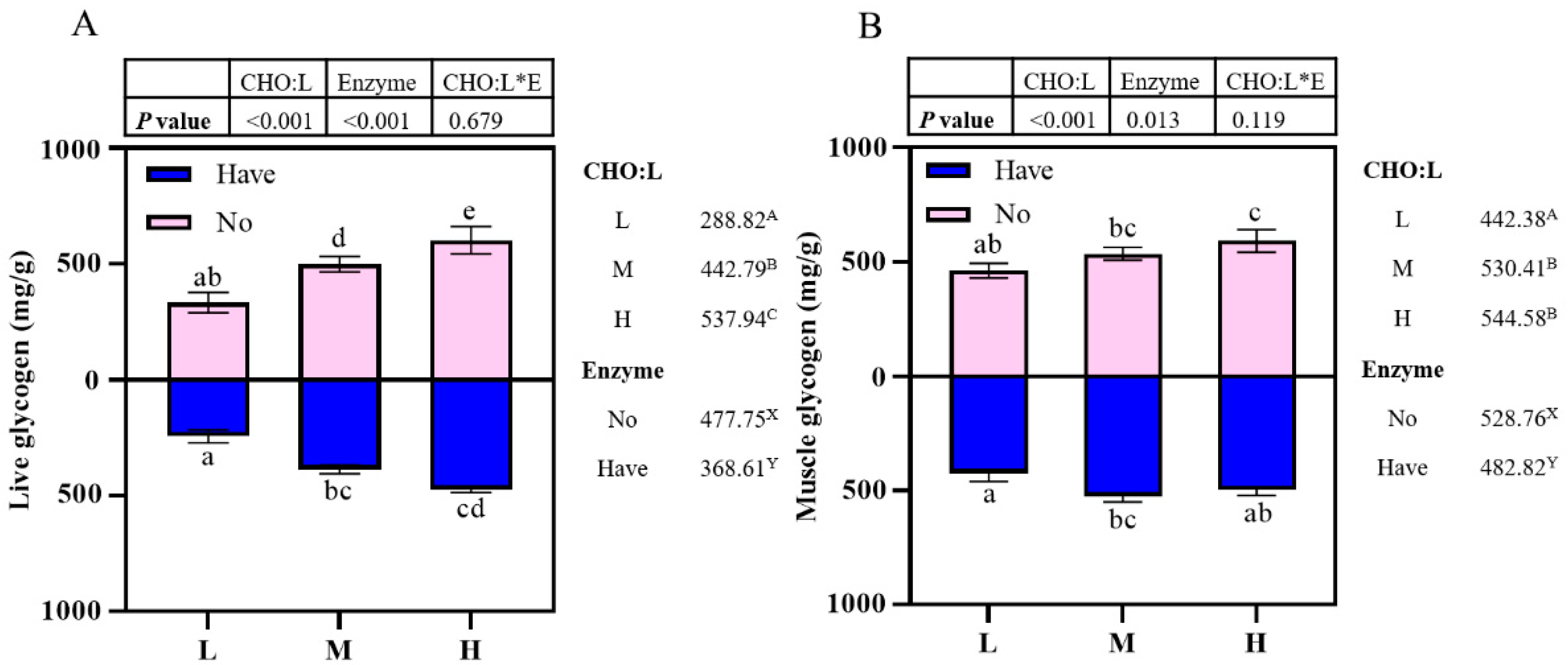
28]. Insulin levels were highest in fish fed a CHO:L ratio of 3.91, and we also found that glycogen content both in liver and muscle increased significantly with the increase in dietary CHO: L ratios, which was a reflection of the self-balancing mechanism of internal environmental homeostasis in fish.
Fish use glucose in a way similar to that of mammals, alternating glycolysis and glycogenesis to maintain glucose balance [
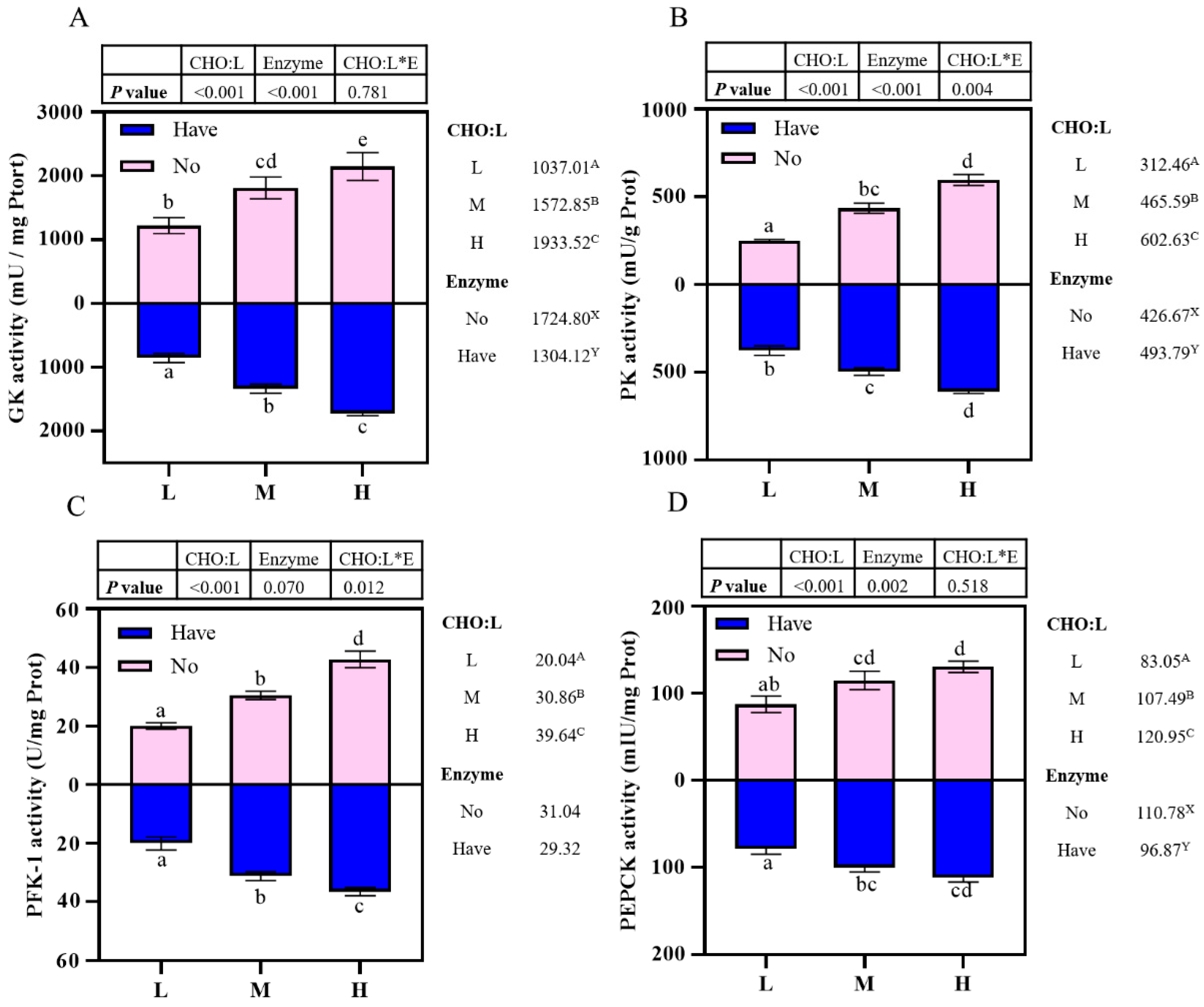
45]. Many studies have found that carbohydrate in the diet can regulate the activity of GK and its gene expression [
46,
47,
48,
49]. The primary function of GK, one of the four hexokinases, is to remove glucose from the blood after meals. Some studies have reported that the GK activity in the liver of European sea bass
(Dicentrarchus labrax) and gilthead seabream (
Sparus aurata) increased significantly with dietary starch level from 10% to 20% [
50,
51,
52]; however, as the starch level increased from 20% to 30%, GK activity in the liver stopped increasing, indicating it may be close to threshold for the ability to utilize glucose effectively [
52]. In this study, GK activity increased significantly with increasing carbohydrate in the diet, which was similar to the studies described above. In contrast, GK activity was significantly reduced by the addition of complex enzymes to the feed that enhanced the hydrolysis of starch, whereas small-molecule carbohydrates are better utilized by fish than large-molecule carbohydrates [
53], which may be responsible for the reduction in GK activity. Besides GK, PK and PFK-1 are also important rate-limiting enzymes in glycolysis. In this study, PFK-1 activity increased significantly with the increase in dietary CHO: L ratios, and there was no significant difference with the addition of carbohydrase. As in mammals, PFK-1 activity in the liver of fish fed a high-carbohydrate, low-protein diet was higher than in those fed a high-protein, low-carbohydrate diet [
54,
55]; PK activity elevated significantly with the increase in dietary CHO: L ratios, and also with carbohydrase supplementation. However, previous studies have shown that the supplementation of 30% starch or glucose in the diet of gibel carp (
Carassius auratus gibelio) has no significant effect on PK activity of livers [
56]. This is due to differences in the regulation of PK activity by the content and source of carbohydrate in the feed. In our research, PEPCK activity followed a similar trend to GK activity and was regulated by CHO: L ratios and carbohydrase. However, the ingestion of digestible sugars can lead to a decrease in PEPCK activity and gene expression reported in common carp (
Cyprinus carpio) [
57]. In contrast, the PEPCK activity and gene expression of Atlantic salmon and rainbow trout (
Salmo gairdneri) were not influenced by the digestible sugar content of the diet [
52,
58,
59]. Therefore, further research is needed to explore this aspect.
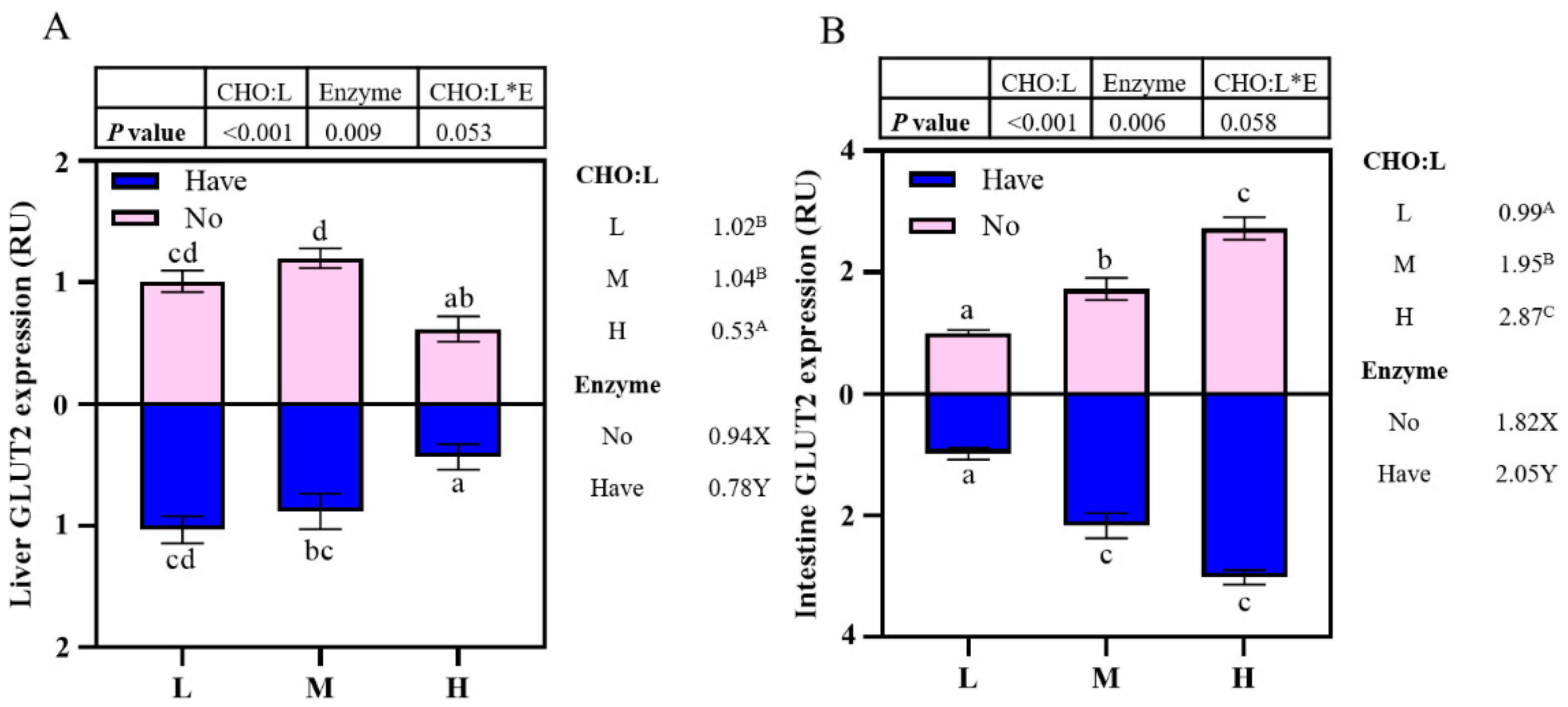
Glucose enters the circulatory system in the digestive tract mainly through the transport of glucose transport carriers across the cell membrane [
60]. The transport of glucose into the cell is the first step in the utilization of glucose by all tissues. Molecular evidence suggested that the main function of glucose transporter 2 (
glut2) in fish is to transport high concentrations of glucose from the intestine to the blood and from the blood to hepatocytes [
61,
62]. As with mammals,
glut2 was as well expressed in the liver and intestine of sea bass [
63] and rainbow trout [
64]. In the present study,
glut2 expression in the liver was significantly decreased with an increase in the dietary CHO:L ratio, while it was upregulated in the intestine. Earlier studies have shown that glucoamylase supplementation on a high-carbohydrate diet significantly promoted
glut2 gene expression in the intestine of pompano (
Trachinotus ovatus) [
16]. Higher glucose concentrations in the intestine have an up-regulatory effect on
glut2 expression. It was shown that the intake of glucose increased with the increase in dietary carbohydrate, and then glucose was rapidly transported by
glut2 in the intestine to maintain glucose homeostasis and sustained energy supply. In our experiments, enzyme supplementation did not cause significant effects, but there was a slight upward trend, which could be related to the different carbohydrate sources used and the different carbohydrase.
Ultimately, digestive and absorptive capacity plays a crucial role in the growth and development of fish [
65], and endogenous metabolic enzymes are one of the factors that determines the ability of fish to use dietary carbohydrates as an energy source [
66]. In this experiment, intestinal amylase and trypsin increased with the increase in dietary CHO: L ratio, while lipase activity decreased. The results of this study were in agreement with previous research reported on groupers [
19], in juvenile cobia (
Rachycentron canadum) [
67]. Reference [
67] showed that the activity of amylase and protease significantly increased with the increase in starch level from 1.3% to 12.5%. Reference [
20] suggested that the activity of amylase and trypsin in the intestine increased significantly with the varying of CHO: L ratios from 0.65 to 8.51, and a decreasing trend in lipase. The reason for this difference was that even for the same fish, the CHO: L ratio was smaller and the lipid content was lower in our study, and the digestive enzyme activity in the intestine was induced by the food level, which had a significant difference on the glycolipid metabolism of the fish. The digestive enzyme activities of low to medium CHO:L ratio were significantly increased after carbohydrase supplementation. Some studies revealed that the addition of cellulases to duckweed diet could significantly promote the protease, amylase and lipase activities of grass carp [
68]; the exogenous xylanase supplementation in plant-protein-enriched diets reported on Jian carp (
Cyprinus carpio) [
69]; and a commercial enzyme complex of neutral protease, β-glucanase and xylanase supplementation in diets increased amylase and protease activities in tilapia [
70]. Therefore, carbohydrase supplementation at an appropriate carbohydrate–lipid ratio can improve the digestive enzyme activity, improve the digestive ability and promote the growth performance of groupers.
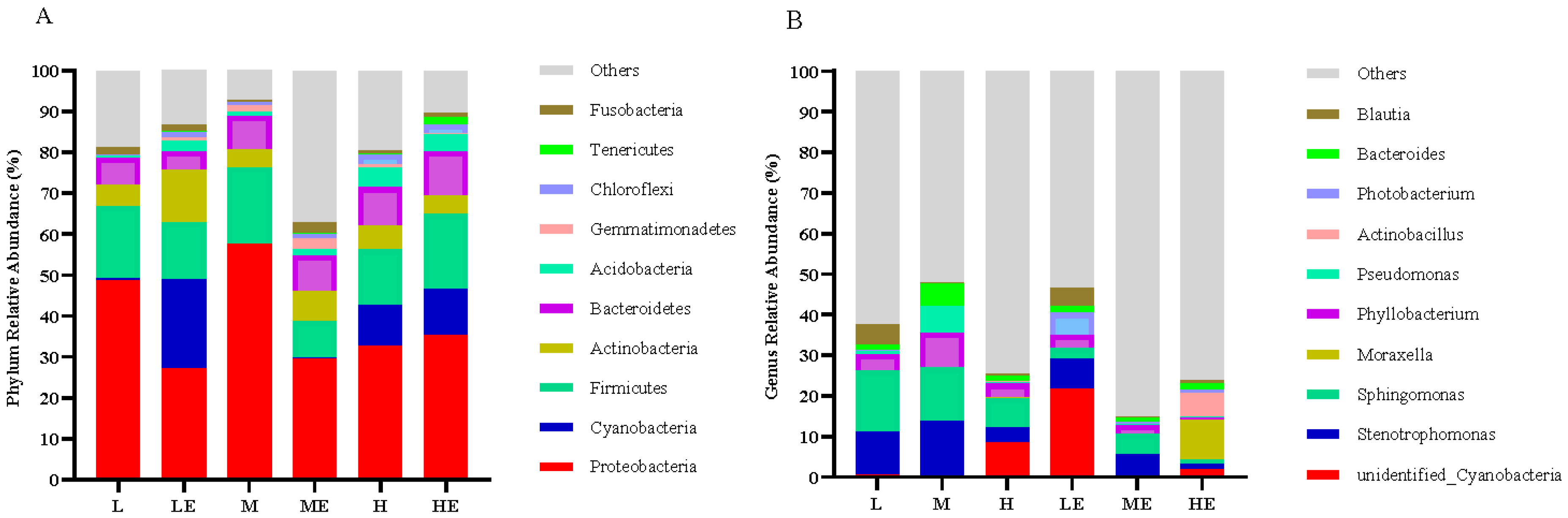
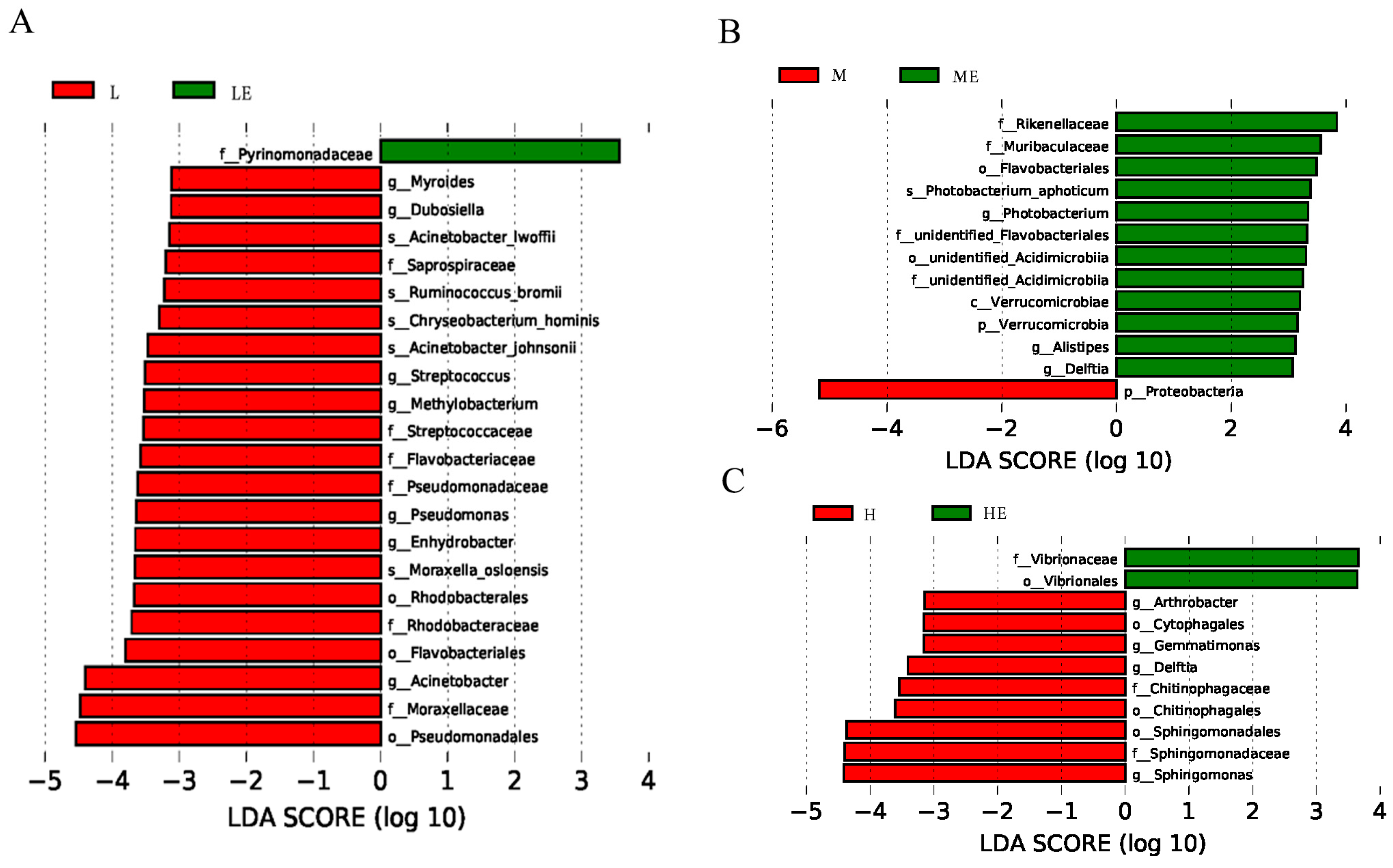
Once fish are exposed to the surrounding aquatic environment and live bait, a variety of bacteria begin to colonize the intestine, and the first colonized bacteria are adapted to the host intestinal environment, so the intestine is the largest number of bacteria in the digestive tract [
71]. Studies on grass carp [
68] have reported that cellulase supplementation promotes the emergence of certain flora, such as
Bacillus and
Sphingomonas, which aid in the digestion of cellulose.
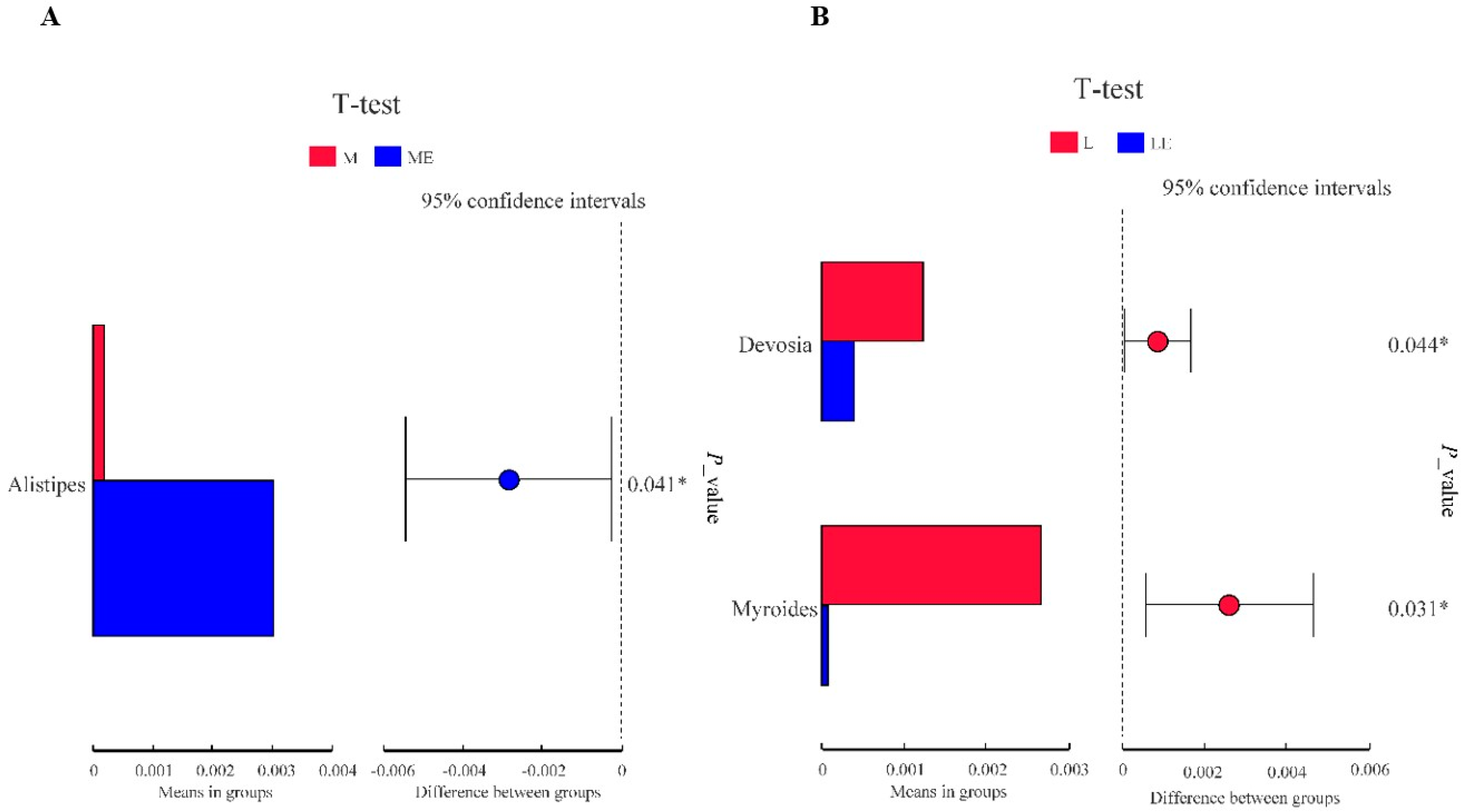
Myroides was related to severe skin and soft tissue infections, including amputation site and urinary tract infections, ventricular ventriculitis and bacteremia [
72].
Alistipes is a producer of acetic acid, a short-chain fatty acid that has been shown to have anti-inflammatory mechanisms in previous studies, and it can be assumed that a decrease in
Alistipes causes a decrease in short-chain fatty acids [
73]. Therefore, to a certain extent, the supplementation of exogenous enzymes promotes the growth of beneficial flora and inhibits the reproduction of harmful flora.
Rikenellaceae is able to use glucose from the environment to produce acetic acid [
74], thus eliminating glucose from the surrounding environment. Studies based on 16S rRNA sequencing have found that among them,
Chryseobacterium,
Cloacibacterium and
Flavobacterium belong to the anerobic class
Flavobacteriales, which are known for their metabolism of lipids and carbohydrates [
75]. This was a result consistent with the emergence of communities that contributed to the digestion of carbohydrates after supplementation with carbohydrase in our experiments.
In summary, our experimental results showed that the best growth performance and feed utilization were obtained at a dietary CHO:L ratio of 1.92; carbohydrase supplementation at dietary CHO:L ratio of 3.91 can significantly improve growth performance, digestive enzymes, glycolytic enzymes, glut2 expression in the intestine, and the appearance of bacteria that contributed to the digestion of carbohydrates. Overall, it is a feasible strategy for carbohydrase supplementation with an appropriately high CHO: L ratio.










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}