


Liver Injury and Metabolic Dysregulation in Largemouth Bass (Micropterus salmoides) after Ammonia Exposure

Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Fish Rearing and Ammonia Challenge Experiment
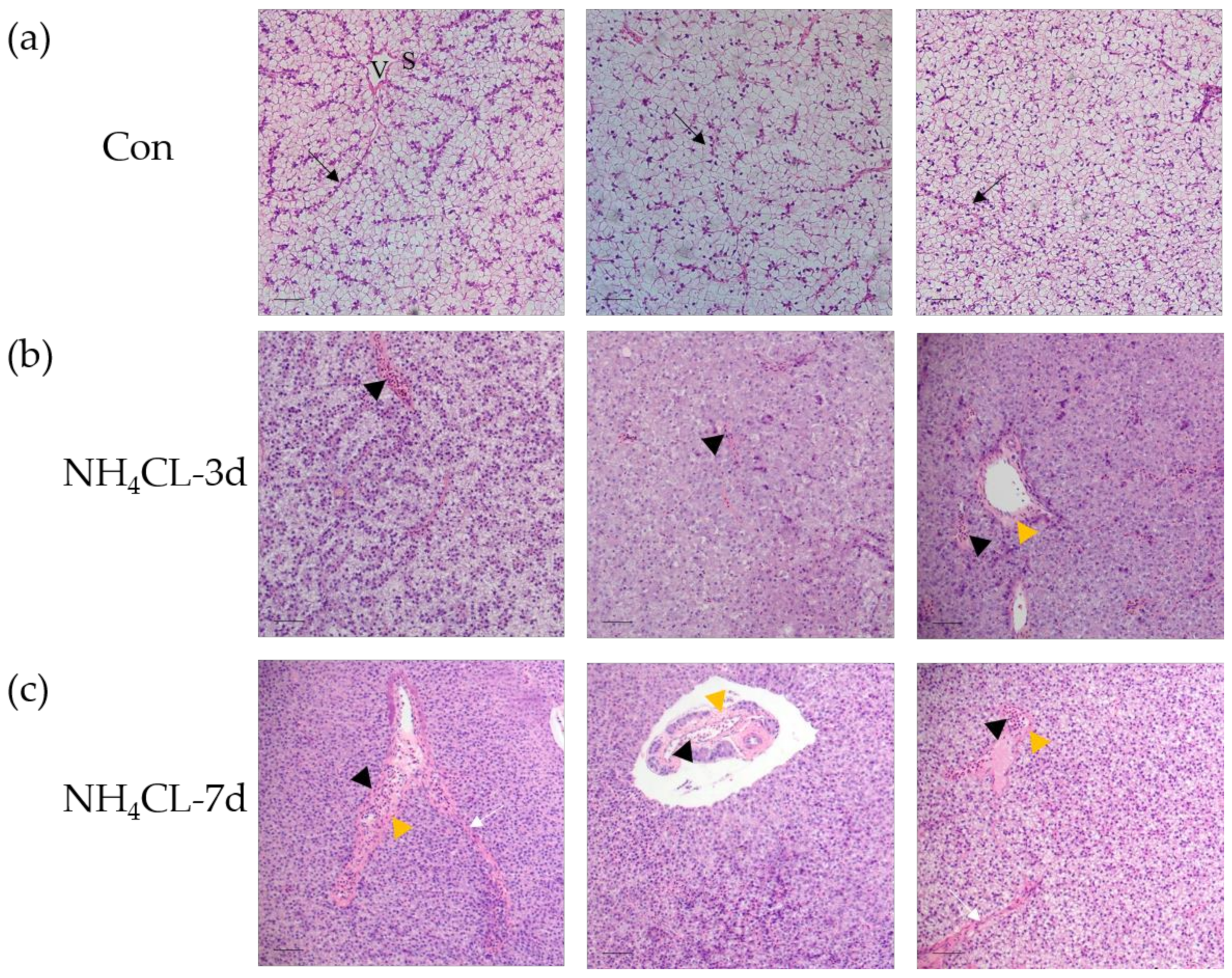
2.3. Histological Analysis
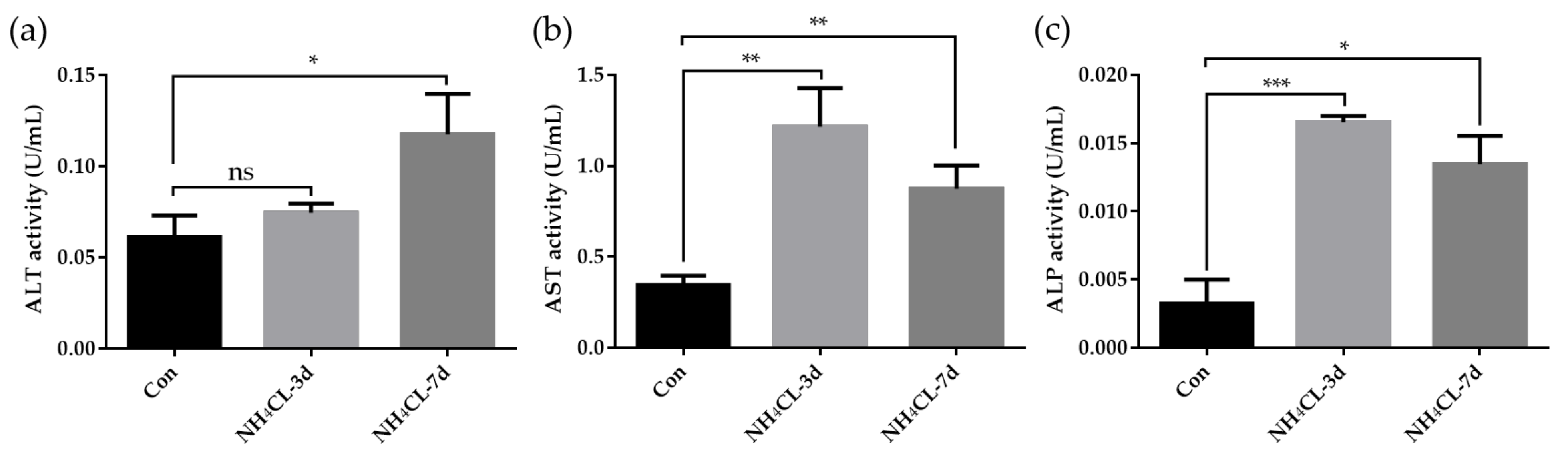
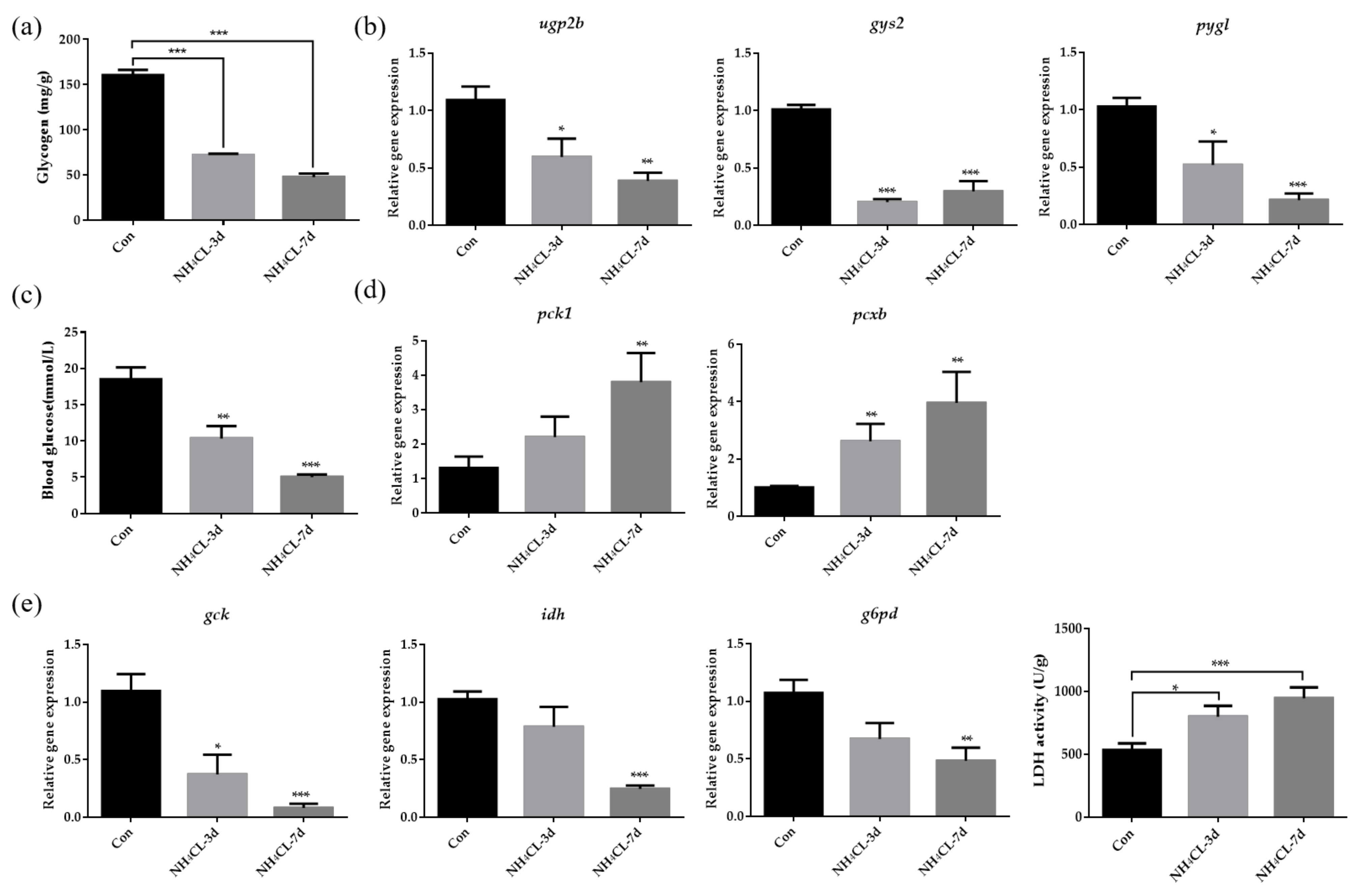
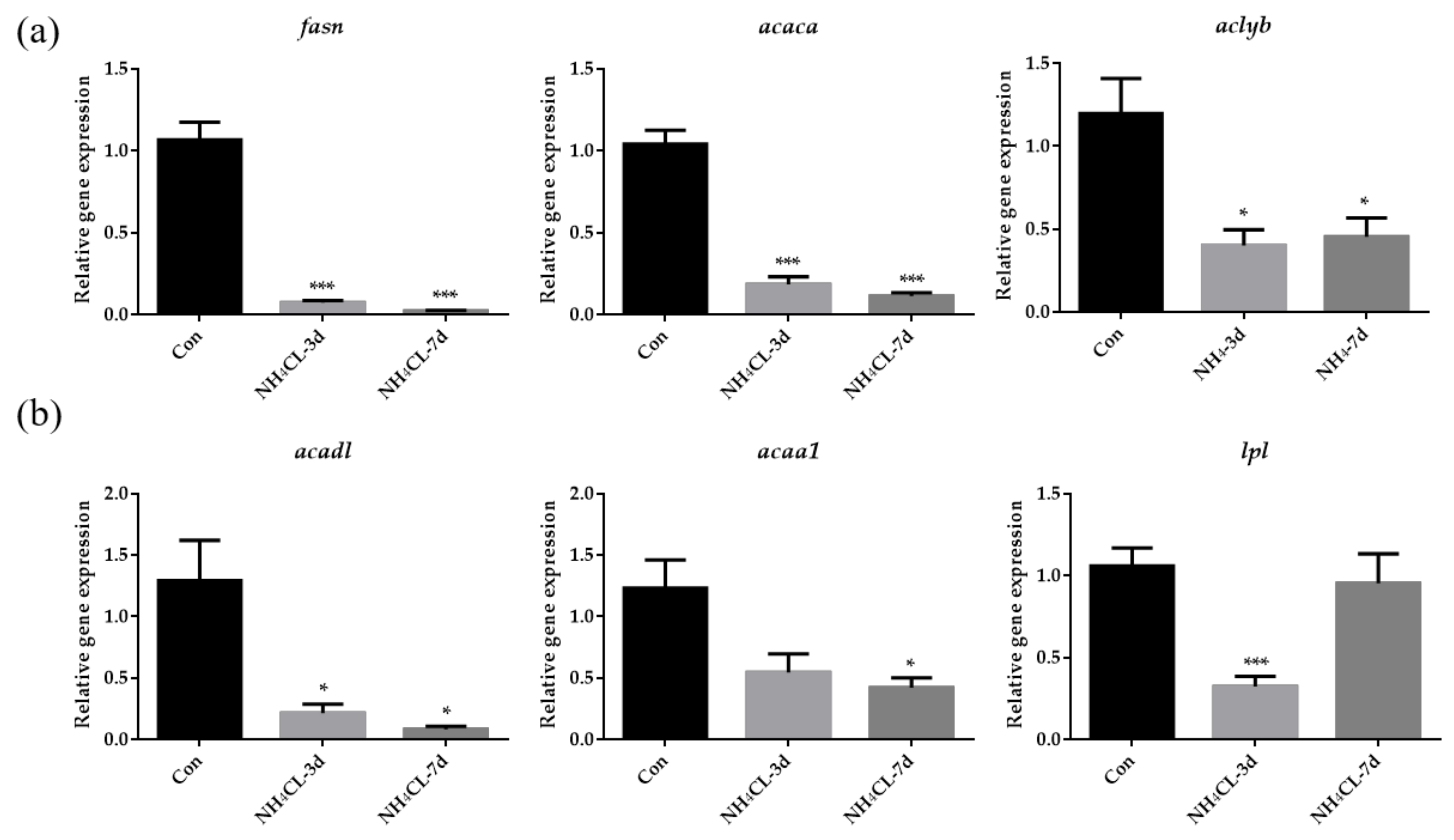
2.4. Assay of Enzyme Activity and Liver Glycogen
2.5. RNA Extraction, Library Preparation, and Transcriptome Sequencing
2.6. RT-qPCR Analysis
2.7. Statistical Analysis
3. Results
3.1. Ammonia Exposure Significantly Affected Liver Health
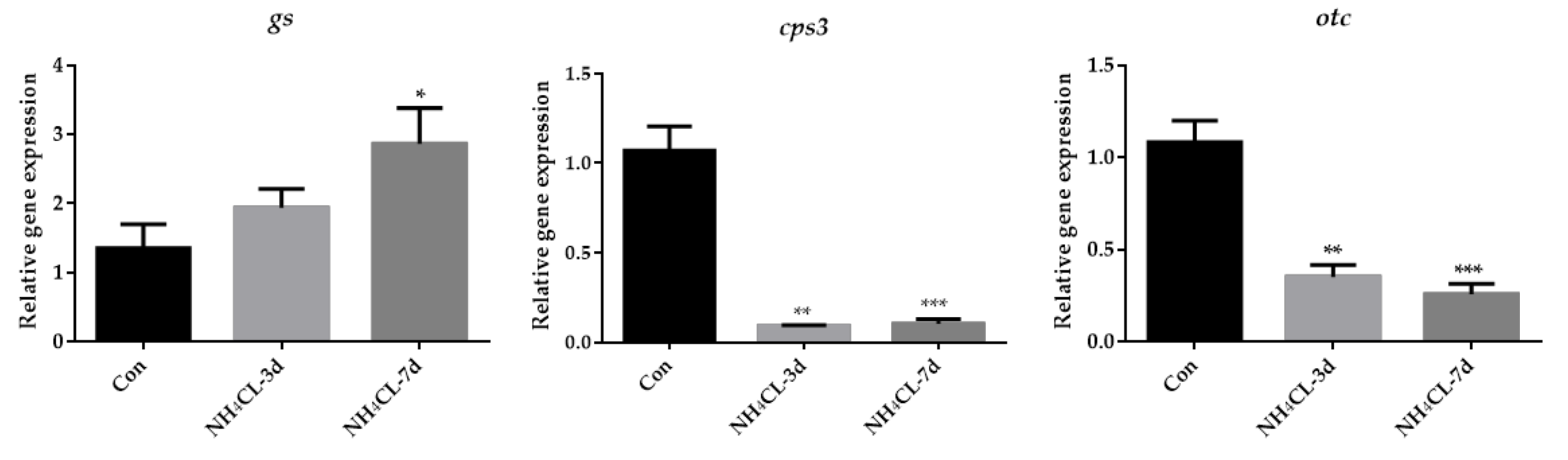
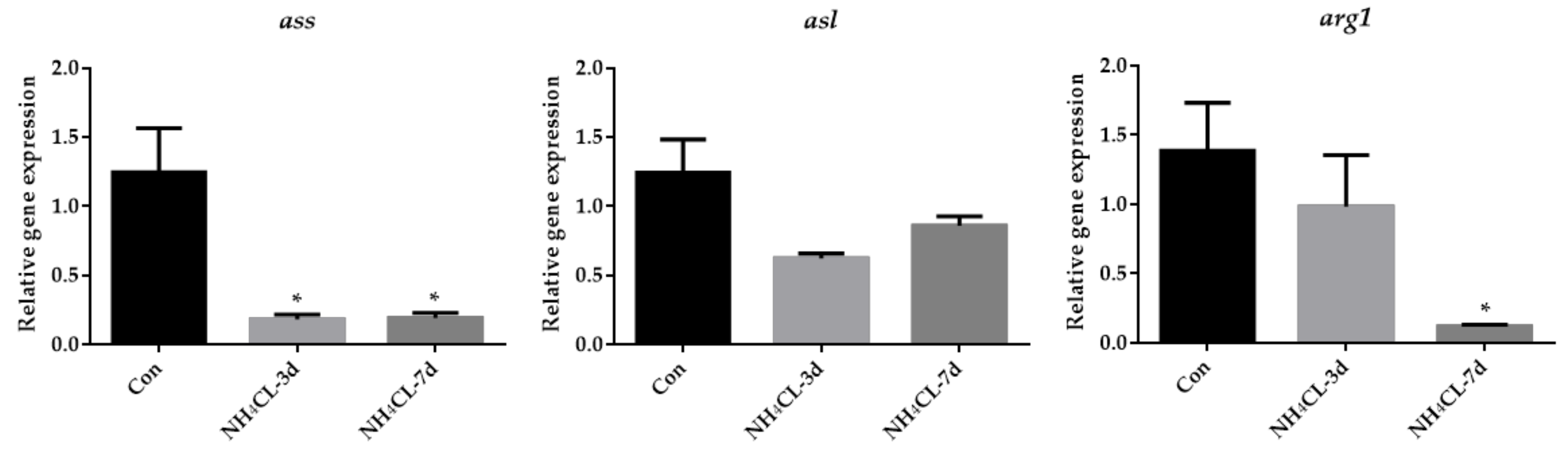
3.2. Ammonia Exposure Induced Hepatic Metabolic Modulation
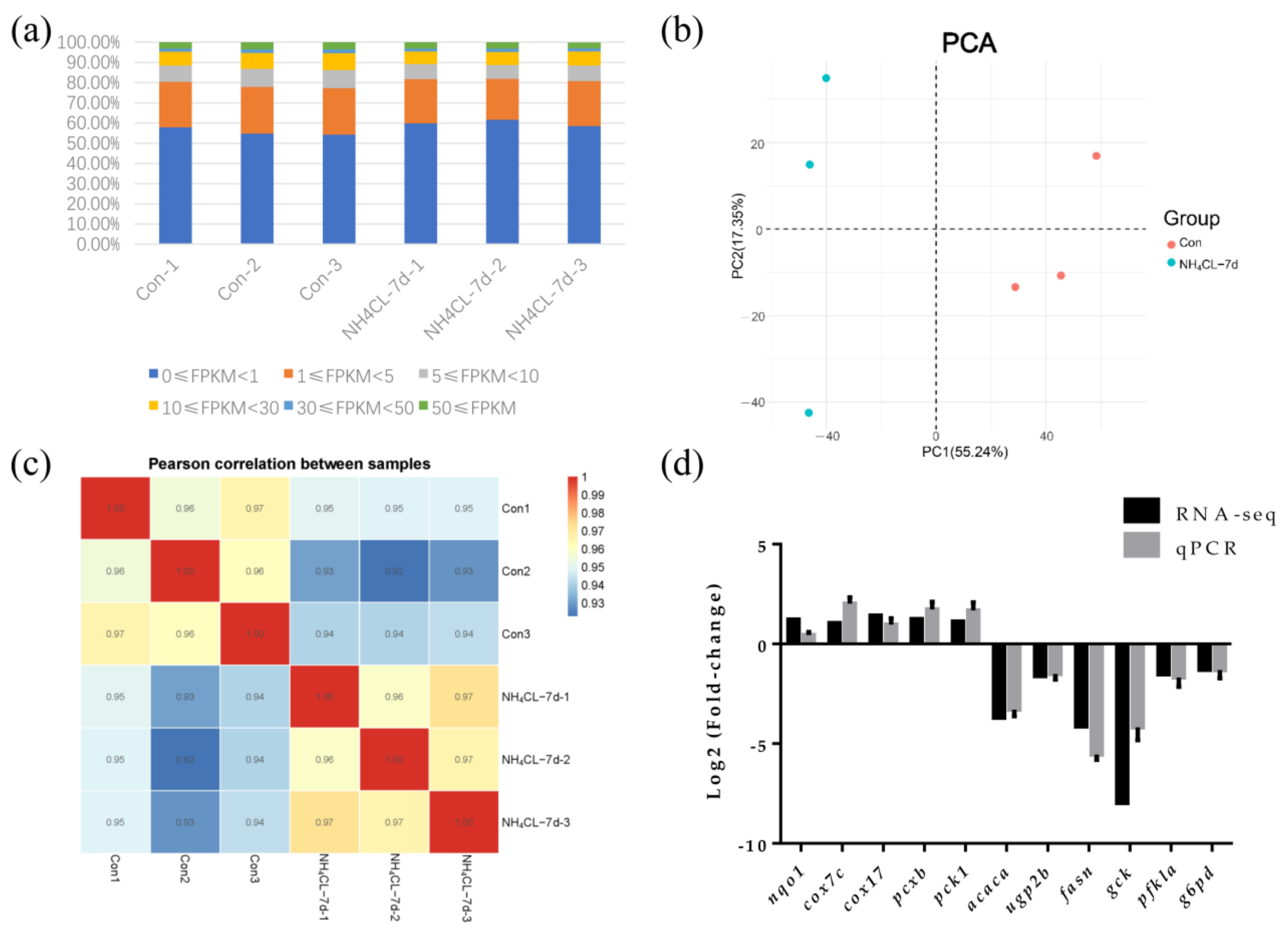
3.3. Transcriptome Sequencing of Largemouth Bass Liver in Both Control Group and Ammonia Exposure Group
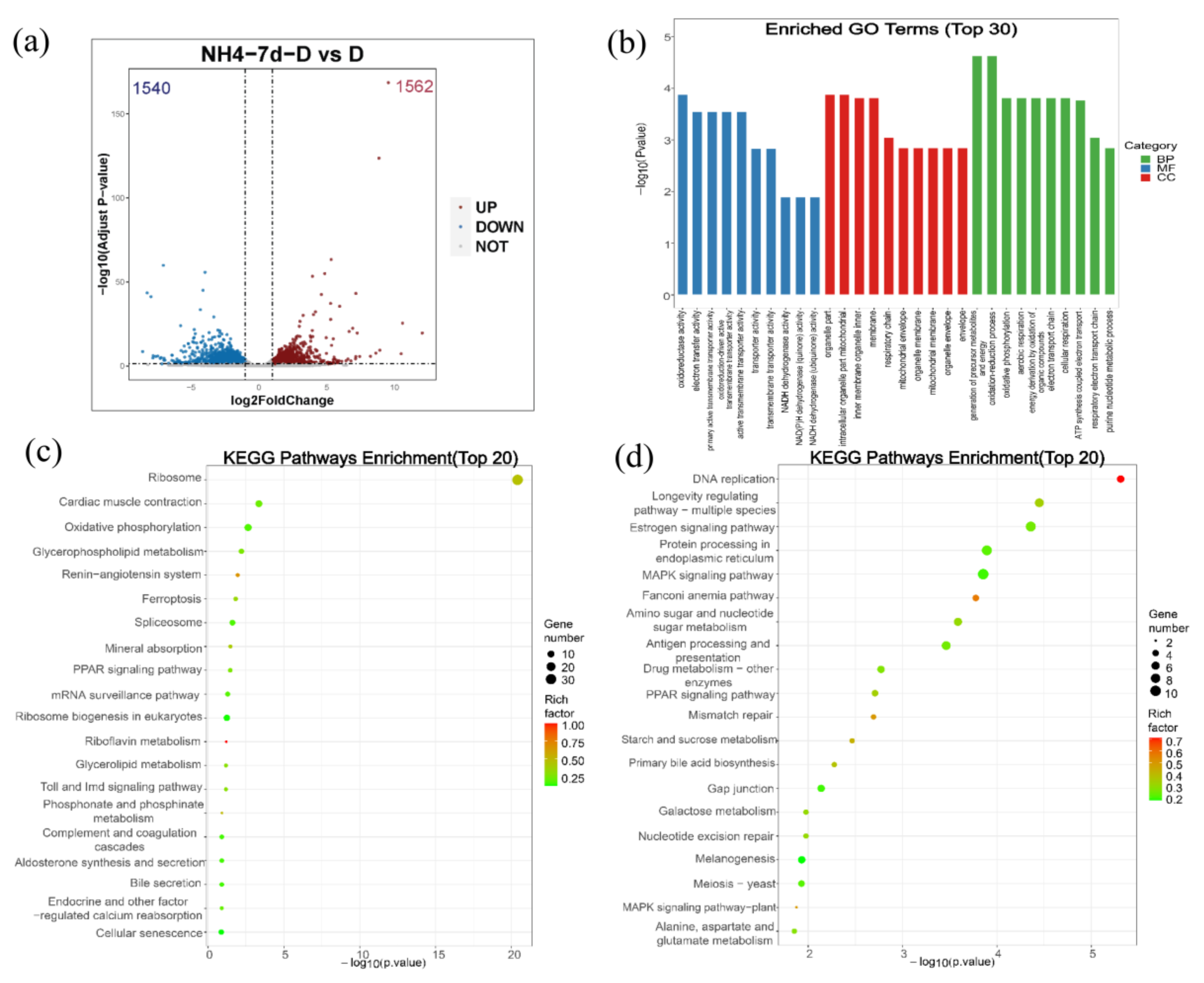
3.4. Differential Gene Expression Analysis of Largemouth Bass Liver in Both Control Group and Ammonia Exposure Group
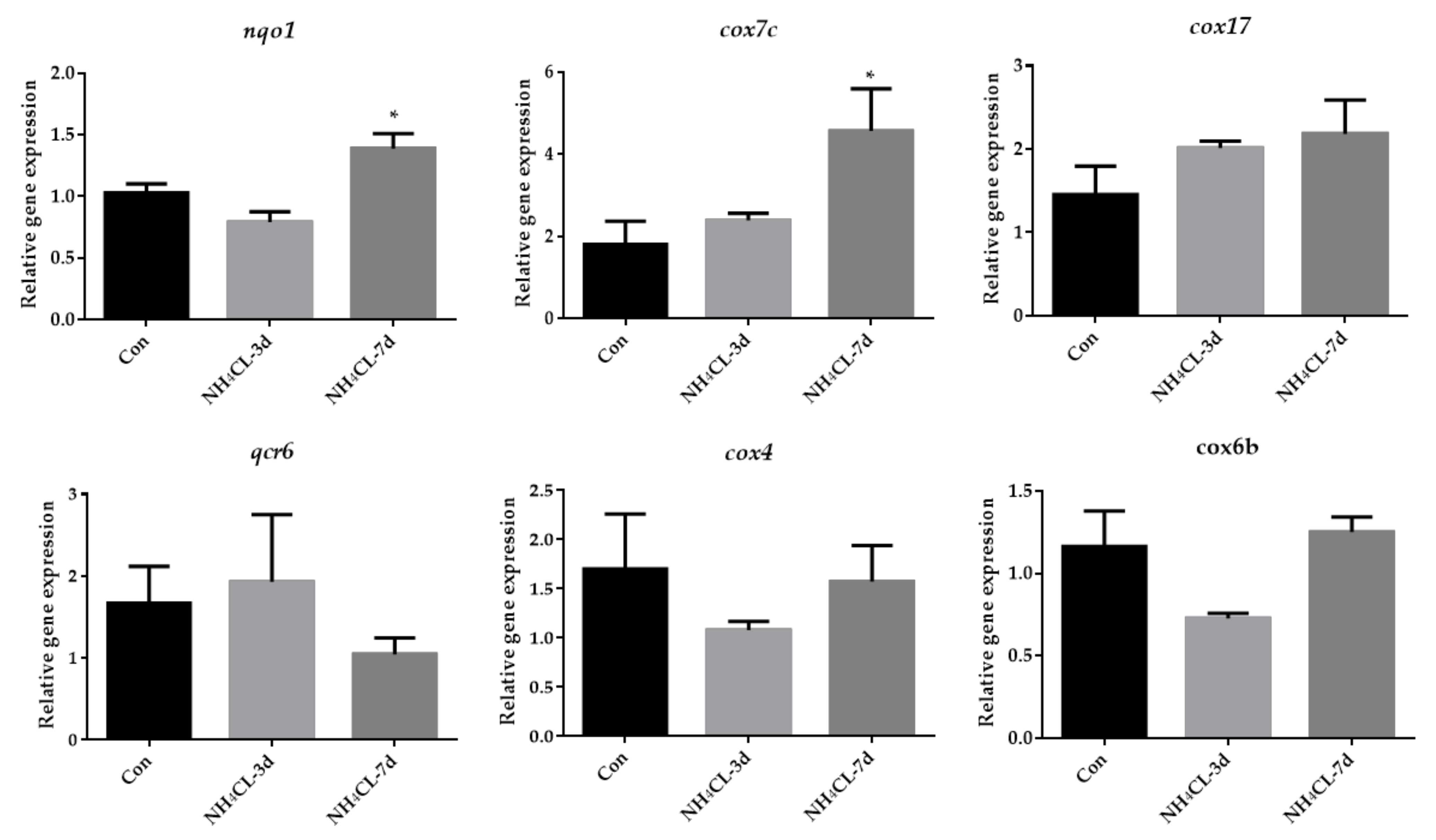
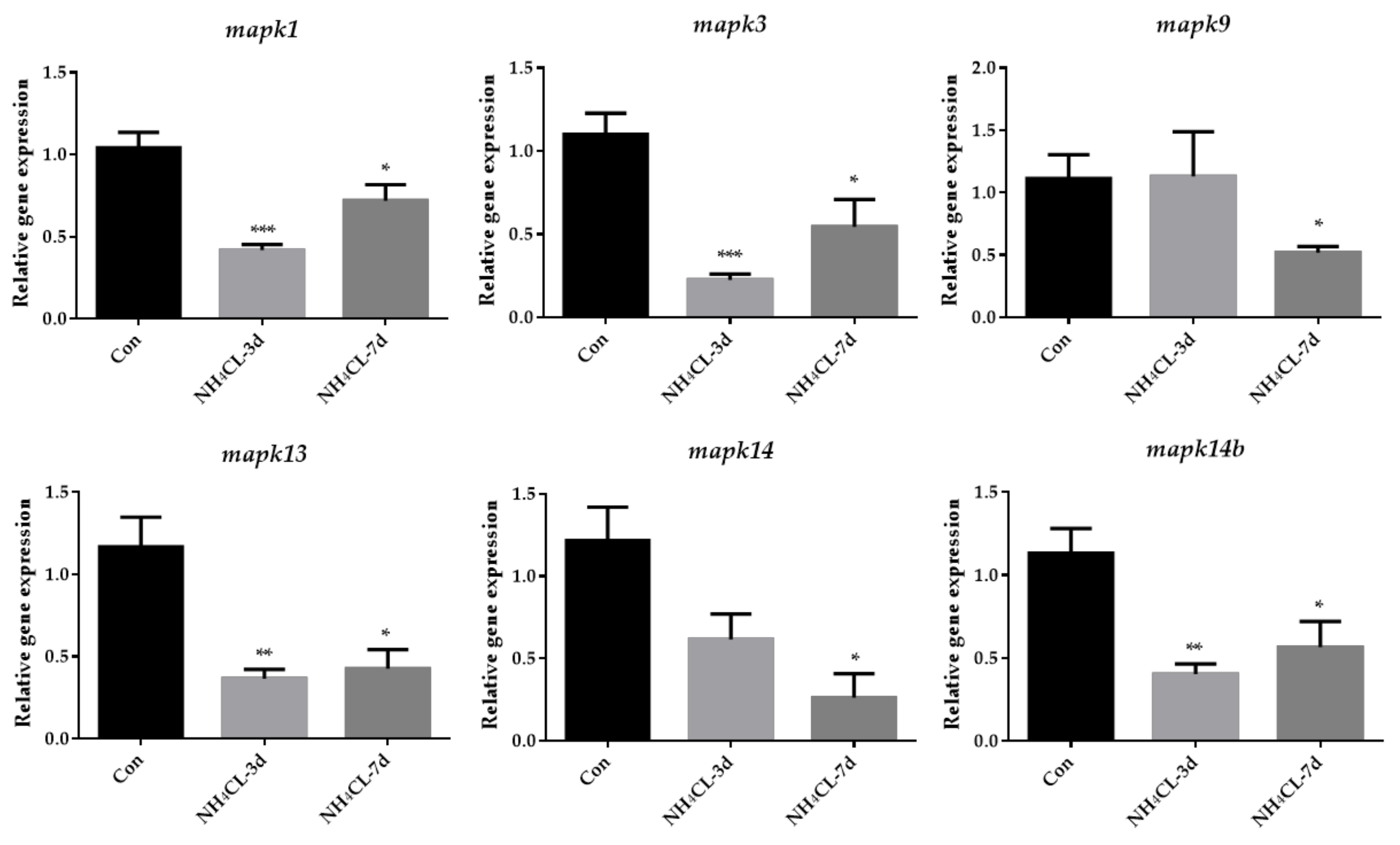
3.5. Ammonia Exposure Increased Oxidative Phosphorylation but Inhibited P38 MAPK Pathway
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Li, L.H.; Qi, H.X. Effect of acute ammonia exposure on the glutathione redox system in FFRC strain common carp (Cyprinus carpio L.). Environ. Sci. Pollut. Res. Int. 2019, 26, 27023–27031. [Google Scholar] [CrossRef] [PubMed]
- Egnew, N.; Renukdas, N.; Ramena, Y.; Yadav, A.K.; Kelly, A.M.; Lochmann, R.T.; Sinha, A.K. Physiological insights into largemouth bass (Micropterus salmoides) survival during long-term exposure to high environmental ammonia. Aquat. Toxicol. 2019, 207, 72–82. [Google Scholar] [CrossRef] [PubMed]
- Divya, M.; Gopi, N.; Iswarya, A.; Govindarajan, M.; Alharbi, N.S.; Kadaikunnan, S.; Khaled, J.M.; Almanaa, T.N.; Vaseeharan, B. β-glucan extracted from eukaryotic single-celled microorganism Saccharomyces cerevisiae: Dietary supplementation and enhanced ammonia stress tolerance on Oreochromis mossambicus. Microb. Pathog. 2020, 139, 103917. [Google Scholar] [CrossRef]
- Xu, Z.; Cao, J.; Qin, X.; Qiu, W.; Mei, J.; Xie, J. Toxic Effects on Bioaccumulation, Hematological Parameters, Oxidative Stress, Immune Responses and Tissue Structure in Fish Exposed to Ammonia Nitrogen: A Review. Animals 2021, 11, 3304. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; Kuang, Y.; Ouyang, K.; Zhang, C.; Yang, H.; Chen, S.; Tang, R.; Zhang, X.; Li, D.; Li, L. Ammonia in the presence of nano titanium dioxide (nano-TiO(2)) induces greater oxidative damage in the gill and liver of female zebrafish. Ecotoxicol. Environ. Saf. 2022, 236, 113458. [Google Scholar] [CrossRef]
- Mangang, Y.A.; Pandey, P.K.J.A. Hemato-biochemical responses and histopathological alterations in the gill and kidney tissues of Osteobrama belangeri (Valenciennes, 1844) exposed to different sub-lethal unionized ammonia. Aquaculture 2021, 542, 736887. [Google Scholar] [CrossRef]
- Liu, M.J.; Guo, H.Y.; Liu, B.; Zhu, K.C.; Guo, L.; Liu, B.S.; Zhang, N.; Yang, J.W.; Jiang, S.G.; Zhang, D.C. Gill oxidative damage caused by acute ammonia stress was reduced through the HIF-1α/NF-κb signaling pathway in golden pompano (Trachinotus ovatus). Ecotoxicol. Environ. Saf. 2021, 222, 112504. [Google Scholar] [CrossRef]
- Randall, D.J.; Tsui, T.K. Ammonia toxicity in fish. Mar. Pollut. Bull 2002, 45, 17–23. [Google Scholar] [CrossRef]
- Lin, L.Y.; Horng, J.L.; Cheng, C.A.; Chang, C.Y.; Cherng, B.W.; Liu, S.T.; Chou, M.Y. Sublethal ammonia induces alterations of emotions, cognition, and social behaviors in zebrafish (Danio rerio). Ecotoxicol. Environ. Saf. 2022, 244, 114058. [Google Scholar] [CrossRef]
- Gao, X.; Wang, X.; Wang, X.; Fang, Y.; Cao, S.; Huang, B.; Chen, H.; Xing, R.; Liu, B. Toxicity in Takifugu rubripes exposed to acute ammonia: Effects on immune responses, brain neurotransmitter levels, and thyroid endocrine hormones. Ecotoxicol. Environ. Saf. 2022, 244, 114050. [Google Scholar] [CrossRef]
- Mitra, V.; Metcalf, J. Metabolic functions of the liver. Anaesth. Intens. Care 2012, 13, 54–55. [Google Scholar] [CrossRef]
- Zorn, A.M. Liver development. In StemBook; Harvard Stem Cell Institute: Cambridge, MA, USA, 2008. [Google Scholar]
- Trefts, E.; Gannon, M.; Wasserman, D.H. The liver. Curr. Biol. 2017, 27, R1147–R1151. [Google Scholar] [CrossRef]
- Crispe, I.N. The liver as a lymphoid organ. Annu. Rev. Immunol. 2009, 27, 147–163. [Google Scholar] [CrossRef]
- Jin, J.; Wang, Y.; Wu, Z.; Hergazy, A.; Lan, J.; Zhao, L.; Liu, X.; Chen, N.; Lin, L. Transcriptomic analysis of liver from grass carp (Ctenopharyngodon idellus) exposed to high environmental ammonia reveals the activation of antioxidant and apoptosis pathways. Fish Shellfish Immunol. 2017, 63, 444–451. [Google Scholar] [CrossRef]
- Xu, H.; Xu, R.; Wang, X.; Liang, Q.; Zhang, L.; Liu, J.; Wei, J.; Lu, Y.; Yu, D. Co-infections of Aeromonas veronii and Nocardia seriolae in largemouth bass (Micropterus salmoides). Microb. Pathog. 2022, 173, 105815. [Google Scholar] [CrossRef]
- Ministry of Agriculture and Rural Affairs of the People’s Republic of China. China Fishery Statistical Yearbook; China Agriculture Press: Beijing, China, 2022.
- Guo, J.L.; Kuang, W.M.; Zhong, Y.F.; Zhou, Y.L.; Chen, Y.J.; Lin, S.M. Effects of supplemental dietary bile acids on growth, liver function and immunity of juvenile largemouth bass (Micropterus salmoides) fed high-starch diet. Fish Shellfish Immunol. 2020, 97, 602–607. [Google Scholar] [CrossRef]
- Huang, A.; Sha, L.A.; Xia, C.B.; Hz, C.; Jy, C.; Yi, G.D.; Yo, D.; Dc, A.; Ly, D.; Ll, B.J.A. Comparative pathological description of nocardiosis in largemouth bass (Micropterus salmoides) and other Perciformes. Aquaculture 2020, 534, 736193. [Google Scholar] [CrossRef]
- Suski, C.D.; Kieffer, J.D.; Killen, S.S.; Tufts, B.L. Sub-lethal ammonia toxicity in largemouth bass. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2007, 146, 381–389. [Google Scholar] [CrossRef]
- Zhao, L.; Cui, C.; Liu, Q.; Sun, J.; He, K.; Adam, A.A.; Luo, J.; Li, Z.; Wang, Y.; Yang, S. Combined exposure to hypoxia and ammonia aggravated biological effects on glucose metabolism, oxidative stress, inflammation and apoptosis in largemouth bass (Micropterus salmoides). Aquat. Toxicol. 2020, 224, 105514. [Google Scholar] [CrossRef]
- Gao, J.; Zhu, Y.; Guo, Z.; Xu, G.; Xu, P. Transcriptomic analysis reveals different responses to ammonia stress and subsequent recovery between Coilia nasus larvae and juveniles. Comp Biochem. Physiol. C Toxicol. Pharmacol. 2020, 230, 108710. [Google Scholar] [CrossRef]
- Xue, S.; Lin, J.; Zhou, Q.; Wang, H.; Han, Y.J.A.R. Effect of ammonia stress on transcriptome and endoplasmic reticulum stress pathway for common carp (Cyprinus carpio) hepatopancreas. Aquac. Rep. 2021, 20, 100694. [Google Scholar] [CrossRef]
- Shang, Z.H.; Huang, M.; Wu, M.X.; Mi, D.; You, K.; Zhang, Y.L. Transcriptomic analyses of the acute aerial and ammonia stress response in the gill and liver of large-scale loach (Paramisgurnus dabryanus). Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2021, 250, 109185. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Chen, Z.; Wang, Y.; Zou, J.; Li, S.; Guo, X.; Gao, J.; Wang, Q. Largemouth bass (Micropterus salmoides) exhibited better growth potential after adaptation to dietary cottonseed protein concentrate inclusion but experienced higher inflammatory risk during bacterial infection. Front. Immunol. 2022, 13, 997985. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Kanehisa, M.; Araki, M.; Goto, S.; Hattori, M.; Hirakawa, M.; Itoh, M.; Katayama, T.; Kawashima, S.; Okuda, S.; Tokimatsu, T.; et al. KEGG for linking genomes to life and the environment. Nucleic Acids Res. 2008, 36, D480–D484. [Google Scholar] [CrossRef]
- Yuan, Y.; Jiang, X.; Wang, X.; Chen, N.; Li, S. Toxicological impacts of excessive lithium on largemouth bass (Micropterus salmoides): Body weight, hepatic lipid accumulation, antioxidant defense and inflammation response. Sci. Total. Environ. 2022, 841, 156784. [Google Scholar] [CrossRef]
- Liang, X.; Chen, P.; Wu, X.; Xing, S.; Morais, S.; He, M.; Gu, X.; Xue, M. Effects of High Starch and Supplementation of an Olive Extract on the Growth Performance, Hepatic Antioxidant Capacity and Lipid Metabolism of Largemouth Bass (Micropterus salmoides). Antioxidants 2022, 11, 577. [Google Scholar] [CrossRef]
- Esam, F.; Khalafalla, M.M.; Gewaily, M.S.; Abdo, S.; Hassan, A.M.; Dawood, M.A.O. Acute ammonia exposure combined with heat stress impaired the histological features of gills and liver tissues and the expression responses of immune and antioxidative related genes in Nile tilapia. Ecotoxicol. Environ. Saf. 2022, 231, 113187. [Google Scholar] [CrossRef]
- Rama, S.; Manjabhat, S.N. Protective effect of shrimp carotenoids against ammonia stress in common carp, Cyprinus carpio. Ecotoxicol. Environ. Saf. 2014, 107, 207–213. [Google Scholar] [CrossRef]
- Sharma, U.; Pal, D.; Prasad, R. Alkaline phosphatase: An overview. Indian J. Clin. Biochem. 2014, 29, 269–278. [Google Scholar] [CrossRef]
- Yeong, K.J.; Chang, Y.J. Effects of Ammonia Concentration on Histological and Physiological Status in Black Seabream (Acanthopagrus schlegeli) . Korean. J. Fish. Aquat. Sci. 1996, 29, 828–836. [Google Scholar]
- Hu, Z.; Han, D.; Zhang, T.; Li, D.; Tang, R. Ammonium induces oxidative stress, endoplasmic reticulum stress, and apoptosis of hepatocytes in the liver cell line of grass carp (Ctenopharyngodon idella). Environ. Sci. Pollut. Res. Int. 2022. [Google Scholar] [CrossRef]
- Dalvi, R.S.; Das, T.; Debnath, D.; Yengkokpam, S.; Baruah, K.; Tiwari, L.R.; Pal, A.K. Metabolic and cellular stress responses of catfish, Horabagrus brachysoma (Günther) acclimated to increasing temperatures. J. Therm. Biol. 2017, 65, 32–40. [Google Scholar] [CrossRef]
- Habegger, K.M. Cross Talk Between Insulin and Glucagon Receptor Signaling in the Hepatocyte. Diabetes 2022, 71, 1842–1851. [Google Scholar] [CrossRef]
- Li, D.; Chen, S.; Liu, C.; Wei, B.; Li, X. Liver transcriptome analysis reveals biological pathways and transcription factors in response to high ammonia exposure. Inhal. Toxicol. 2022, 34, 219–229. [Google Scholar] [CrossRef]
- Hertz, L.; Kala, G. Energy metabolism in brain cells: Effects of elevated ammonia concentrations. Metab. Brain Dis. 2007, 22, 199–218. [Google Scholar] [CrossRef]
- Feng, Y.; Xiong, Y.; Qiao, T.; Li, X.; Jia, L.; Han, Y. Lactate dehydrogenase A: A key player in carcinogenesis and potential target in cancer therapy. Cancer Med. 2018, 7, 6124–6136. [Google Scholar] [CrossRef]
- Khare, A.; Chhawani, N.; Kumari, K. Glutathione reductase and catalase as potential biomarkers for synergistic intoxication of pesticides in fish. Biomarkers 2019, 24, 666–676. [Google Scholar] [CrossRef]
- Zhang, M.; Hou, C.; Li, M.; Qian, Y.; Xu, W.; Meng, F.; Wang, R. Modulation of lipid metabolism in juvenile yellow catfish (Pelteobagrus fulvidraco) as affected by feeding frequency and environmental ammonia. Fish Physiol. Biochem. 2019, 45, 115–122. [Google Scholar] [CrossRef]
- Chew, S.F.; Ip, Y.K. Excretory nitrogen metabolism and defence against ammonia toxicity in air-breathing fishes. J. Fish Biol. 2014, 84, 603–638. [Google Scholar] [CrossRef] [PubMed]
- Ren, Q.; Li, M.; Yuan, L.; Song, M.; Xing, X.; Shi, G.; Meng, F.; Wang, R. Acute ammonia toxicity in crucian carp Carassius auratus and effects of taurine on hyperammonemia. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2016, 190, 9–14. [Google Scholar] [CrossRef] [PubMed]
- Jow, L.Y.; Chew, S.F.; Lim, C.B.; Anderson, P.M.; Ip, Y.K. The marble goby oxyeleotris marmoratus activates hepatic glutamine synthetase and detoxifies ammonia to glutamine during air exposure. J. Exp. Biol. 1999, 202, 237–245. [Google Scholar] [CrossRef] [PubMed]
- Ip, Y.K.; Lem, C.B.; Chew, S.F.; Wilson, J.M.; Randall, D.J. Partial amino acid catabolism leading to the formation of alanine in Periophthalmodon schlosseri (mudskipper): A strategy that facilitates the use of amino acids as an energy source during locomotory activity on land. J. Exp. Biol. 2001, 204, 1615–1624. [Google Scholar] [CrossRef]
- Frick, N.T.; Wright, P.A. Nitrogen metabolism and excretion in the mangrove killifish Rivulus marmoratus I. The influence of environmental salinity and external ammonia. J. Exp. Biol. 2002, 205, 79–89. [Google Scholar] [CrossRef]
- Chew, S.F.; Wong, M.Y.; Tam, W.L.; Ip, Y.K. The snakehead Channa asiatica accumulates alanine during aerial exposure, but is incapable of sustaining locomotory activities on land through partial amino acid catabolism. J. Exp. Biol. 2003, 206, 693–704. [Google Scholar] [CrossRef]
- Tay, A.S.; Chew, S.F.; Ip, Y.K. The swamp eel Monopterus albus reduces endogenous ammonia production and detoxifies ammonia to glutamine during 144 h of aerial exposure. J. Exp. Biol. 2003, 206, 2473–2486. [Google Scholar] [CrossRef]
- Chen, Y.; Zhang, R.; Ding, S.; Nian, H.; Zeng, X.; Liu, H.; Xing, H.; Li, J.; Bao, J.; Li, X. Transcriptome Revealed Exposure to the Environmental Ammonia Induced Oxidative Stress and Inflammatory Injury in Spleen of Fattening Pigs. Animals 2022, 12, 1204. [Google Scholar] [CrossRef]
- Ma, Q.; Shi, W.; Jahan, K.; Zhao, W.; Xu, Q.; Huo, Z.; Fang, L.; Yang, F.; Yan, X. Transcriptome sequencing reveals improved ammonia nitrogen tolerance in Zebra II strain of the Manila clam Ruditapes philippinarum. Comp. Biochem. Physiol. Part D Genomics. Proteom. 2022, 44, 101016. [Google Scholar] [CrossRef]
- Zhang, T.; Yan, Z.; Zheng, X.; Fan, J.; Wang, S.; Wei, Y.; Yang, L.; Wang, P.; Guo, S. Transcriptome analysis of response mechanism to ammonia stress in Asian clam (Corbicula fluminea). Aquat. Toxicol. 2019, 214, 105235. [Google Scholar] [CrossRef]
- Tang, D.; Wu, Y.; Wu, L.; Bai, Y.; Zhou, Y.; Wang, Z. The effects of ammonia stress exposure on protein degradation, immune response, degradation of nitrogen-containing compounds and energy metabolism of Chinese mitten crab. Mol. Biol. Rep. 2022, 49, 6053–6061. [Google Scholar] [CrossRef]
- Nolfi-Donegan, D.; Braganza, A.; Shiva, S. Mitochondrial electron transport chain: Oxidative phosphorylation, oxidant production, and methods of measurement. Redox. Biol. 2020, 37, 101674. [Google Scholar] [CrossRef]
- Wang, T.; Yang, C.; Zhang, S.; Rong, L.; Yang, X.; Wu, Z.; Sun, W. Metabolic changes and stress damage induced by ammonia exposure in juvenile Eriocheir sinensis. Ecotoxicol. Environ. Saf. 2021, 223, 112608. [Google Scholar] [CrossRef]
- Lawan, A.; Min, K.; Zhang, L.; Canfran-Duque, A.; Jurczak, M.J.; Camporez, J.P.G.; Nie, Y.; Gavin, T.P.; Shulman, G.I.; Fernandez-Hernando, C.; et al. Skeletal Muscle-Specific Deletion of MKP-1 Reveals a p38 MAPK/JNK/Akt Signaling Node That Regulates Obesity-Induced Insulin Resistance. Diabetes 2018, 67, 624–635. [Google Scholar] [CrossRef]
- Roth, R.J.; Le, A.M.; Zhang, L.; Kahn, M.; Samuel, V.T.; Shulman, G.I.; Bennett, A.M. MAPK phosphatase-1 facilitates the loss of oxidative myofibers associated with obesity in mice. J. Clin. Investig. 2009, 119, 3817–3829. [Google Scholar] [CrossRef]
- Wu, J.J.; Roth, R.J.; Anderson, E.J.; Hong, E.G.; Lee, M.K.; Choi, C.S.; Neufer, P.D.; Shulman, G.I.; Kim, J.K.; Bennett, A.M. Mice lacking MAP kinase phosphatase-1 have enhanced MAP kinase activity and resistance to diet-induced obesity. Cell Metab. 2006, 4, 61–73. [Google Scholar] [CrossRef]
- Bengal, E.; Aviram, S.; Hayek, T. p38 MAPK in Glucose Metabolism of Skeletal Muscle: Beneficial or Harmful? Int. J. Mol. Sci. 2020, 21, 6480. [Google Scholar] [CrossRef]
- Bost, F.; Aouadi, M.; Caron, L.; Even, P.; Belmonte, N.; Prot, M.; Dani, C.; Hofman, P.; Pagès, G.; Pouysségur, J.; et al. The extracellular signal-regulated kinase isoform ERK1 is specifically required for in vitro and in vivo adipogenesis. Diabetes 2005, 54, 402–411. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Abbreviation | Primer Sequence | TM (°C) | Amplicon Size (bp) | Source |
|---|---|---|---|---|---|
| 18S ribosomal RNA | 18s | F: GCAAAGCTGAAACTTAAAGGAATTG R: TCCCGTGTTGAGTCAAATTAAGC | 58.1 59.5 | 80 | XR_005447146 |
| NADH-quinone oxidoreductase subunit I-like | nqo1 | F: AAAAATGGCGACGAGGACAC R: TGTCATCACGCATTAAACCAAG | 59.1 57.6 | 110 | XM_038733185 |
| cytochrome c oxidase subunit 7C | cox7c | F: TGAAGGACCAGGGAAGAACCT R: AAGCCACTGCCAAAGAACAAC | 60.4 59.7 | 87 | XM_038704624 |
| cytochrome c oxidase copper chaperone | cox17 | F: GGCCTCCAAACAAATTAAGCC R: GTCTCTGGACAAGCACAGCA | 58.0 60.2 | 127 | XM_038721185 |
| cytochrome b-c1 complex subunit 6 | qcr6 | F: CATGCACGAGACCATTGTGTAGC R: ACAGAAAGGGTGGCATGTTGT | 62.3 60.4 | 197 | XM_038718343 |
| cytochrome c oxidase subunit 4 isoform 1 | cox4 | F: GGAAGTCTGTCGTTGGAGGG R: GGGACAGGGCCATACACATAC | 60.0 60.2 | 91 | XM_038701371 |
| cytochrome c oxidase subunit 6B1 | cox6b | F: CTGTCCCATGAGCTGGGTTG R: TGGCGATTTCAGACCTTCCC | 60.7 60.0 | 78 | XM_038711198 |
| fatty acid synthase | fasn | F: CGGGTTGACCTGGGAAGAAT R: ACTAATCGCTTCCTGCGGAC | 59.7 60.2 | 109 | XM_038735140 |
| acetyl-CoA carboxylase alpha | acaca | F: TCCCCATCATCACTGGACAC R: AGGCTGCAAATACGGTGGAG | 59.1 60.4 | 114 | XM_038709737 |
| ATP citrate lyase b | aclyb | F: AGGTCTTCCAGCAGCCAAAG R: GTTCCTCCCGAGAGCATACG | 60.2 60.0 | 125 | XM_038735486 |
| acyl-CoA dehydrogenase long chain | acadl | F: ACAAACCGCAAAGCGAAGAC R: GGTAGCCGCACATCCTCAAA | 60.0 60.4 | 152 | XM_038721438 |
| acetyl-CoA acyltransferase 1 | acaa1 | F: ATCATTTCGGGCCACTTGTCT R: GTCAGGCGTCGTGTCCTTT | 60.0 60.3 | 177 | XM_038734444 |
| lipoprotein lipase | lpl | F: CATGGCTGGACGGTAACAGG R: GTCAGCCAGTCCACAACGAT | 60.7 60.3 | 107 | XM_038715978 |
| lipase, hepatic a | lipca | F: CAGTATGGGCTCCTCGGTTTT R: TTGTCACTGCACCTGTAGGC | 60.1 60.3 | 122 | XM_038701258 |
| glucokinase (hexokinase 4) | gck | F: CCCGAGGGATCAGTGTGTG R: CTTGGTCTCCACCTTCCAGC | 59.8 60.3 | 132 | XM_038703172 |
| phosphofructokinase | pfkla | F: AGCTGTGACCAGAATGGGC R: CTATCACAGTCCCACCCAGC | 60.0 59.8 | 149 | XM_038720351 |
| isocitrate dehydrogenase (NADP(+)) 1 | idh1 | F: AGCACAGGACATCATTCCGC R: CTTGATCTTCTGAGACATCCTGATT | 60.8 58.4 | 87 | XM_038721368 |
| dihydrolipoamide S-succinyltransferase | dlst | F: TGGCTACACCAAAGGGTCTG R: AGCCAGCTCATTCTTACGGG | 59.6 59.8 | 119 | XM_038728954 |
| glucose-6-phosphate 1-dehydrogenase | g6pd | F: ACATTCTCTCTCTGCCCGGA R: CGGTTCCAGCCTTTTGTGCT | 60.3 61.5 | 127 | XM_038722146 |
| phosphoenolpyruvate carboxykinase 1 | pck1 | F: GGCTGGAAGGTTGAGTGTGT R: CACGTAGGTTGCCTTGGTTG | 60.2 59.4 | 70 | XM_038696646 |
| pyruvate carboxylase b | pcxb | F: TGGACAAATGCACAGGCAGA R: GAGAGGAAACCGTAGCCTGG | 60.2 59.8 | 150 | XM_038731352 |
| phosphorylase, glycogen, liver | pygl | F: GATCATCGGGGGAAAGGCTG R: CTTGTTTCCCACCACAGGGT | 60.5 60.1 | 109 | XM_038728721 |
| UDP-glucose pyrophosphorylase 2b | ugp2b | F: GGTTCAGGAGTACGCATCCC R: ATGATGACGGTGCCCTTGAG | 60.1 60.1 | 120 | XM_038737118 |
| glycogen synthase 2 | gys2 | F: GGCCCTACTTCGAACACAACT R: CGGCCAAAATGAACCTGGCA | 60.3 61.8 | 118 | XM_038738888 |
| glucan (1,4-alpha-), branching enzyme 1b | gbe1b | F: GACTTCAGGAGGAGGTATGAGC R: GCTGCACACCAAAGGTCAAG | 59.6 60.0 | 109 | XM_038694127 |
| glutamate-ammonia ligase (glutamine synthase) a | gs | F: TTCCGCACCGGAGAATGAG R: TTCCTACTATAACACACCTGGAGA | 59.8 58.1 | 119 | XM_038718011 |
| N-acetylglutamate synthase | nags | F: CAAGGCTTTTCTCCGTGAAG R: TCACCACCACAGGCTTCATA | 57.0 58.6 | 206 | XM_038692344 |
| ornithine transcarbamylase | otc | F: GAGTTGCACGGACACAGCTA R: AGGTGAGGAAGTCAGCCAGA | 60.3 60.2 | 170 | XM_038720771 |
| argininosuccinate synthetase | ass | F: CAGGATAAATGCGGTCAGGT R: GCAAAAAGGTAGGCAATGGA | 57.7 56.6 | 160 | XM_038733331 |
| argininosuccinate lyase | asl | F: CGTGAAAGCTCTGGAAAAGG R: GCAGGAGCACCAATTAGCTC | 57.0 59.0 | 177 | XM_038709368 |
| arginase-1-like | arg1 | F: GCTGGGTGTGAAGGTGTTTT R: TAGGTGAGTCCTCCCACCAC | 58.9 60.3 | 186 | XM_038735677 |
| mitogen-activated protein kinase 1 | mapk1 | F: CTATGGGATGGTGTGCTCTG R: GATCTCTCTCAGGGTGCGTT | 57.8 59.2 | 112 | XM_038702794 |
| mitogen-activated protein kinase 3 | mapk3 | F: CAACCACATACTGAGCGTCCT R: TTGGGCTTTTCAGGTAGGGC | 60.1 60.3 | 105 | XM_038696984 |
| mitogen-activated protein kinase 9 | mapk9 | F: TCGTGCCTGAAAGACGGAG R: ACCAGCACGGTCTTCTCCC | 59.7 61.6 | 90 | XM_038699205 |
| mitogen-activated protein kinase 13 | mapk13 | F: CTGCTTGAGAAGATGCTGGTT R: AGGCTGTCGAAATATGGGTG | 58.6 57.7 | 83 | [29] |
| mitogen-activated protein kinase 14 | mapk14 | F: TTCGATGGAGACGAGATGG R: GAGATGAATGACCGCAGGC | 56.4 58.7 | 161 | [29] |
| mitogen-activated protein kinase 14b | mapk14b | F: TCCCTGGCACAGACCACATTG R: CTTGGGCATGTGGGGAAGTG | 62.6 61.3 | 134 | XM_038696748 |
| Sample | Raw_Reads | Clean_Reads | Clean_Bases | Error (%) | Q20 (%) | Q30 (%) | GC (%) |
|---|---|---|---|---|---|---|---|
| Con-1 | 46,376,164 | 45,499,224 | 6.78 G | 0.03 | 97.82 | 93.8 | 47.88 |
| Con-2 | 51,233,194 | 50,215,662 | 7.49 G | 0.03 | 97.59 | 93.26 | 48.08 |
| Con-3 | 43,772,118 | 42,896,248 | 6.38 G | 0.03 | 97.8 | 93.77 | 48.01 |
| NH4Cl-7d-1 | 55,241,382 | 54,140,060 | 8.03 G | 0.03 | 97.65 | 93.39 | 46.92 |
| NH4Cl-7d-2 | 50,965,374 | 50,045,184 | 7.47 G | 0.03 | 97.59 | 93.22 | 47.87 |
| NH4Cl-7d-3 | 50,297,386 | 49,430,098 | 7.39 G | 0.03 | 97.76 | 93.58 | 46.8 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zou, J.; Hu, P.; Wang, M.; Chen, Z.; Wang, H.; Guo, X.; Gao, J.; Wang, Q. Liver Injury and Metabolic Dysregulation in Largemouth Bass (Micropterus salmoides) after Ammonia Exposure. Metabolites 2023, 13, 274. https://doi.org/10.3390/metabo13020274
Zou J, Hu P, Wang M, Chen Z, Wang H, Guo X, Gao J, Wang Q. Liver Injury and Metabolic Dysregulation in Largemouth Bass (Micropterus salmoides) after Ammonia Exposure. Metabolites. 2023; 13(2):274. https://doi.org/10.3390/metabo13020274
Chicago/Turabian StyleZou, Jiahong, Peng Hu, Mengya Wang, Zhenwei Chen, Huan Wang, Xiaolong Guo, Jian Gao, and Qingchao Wang. 2023. "Liver Injury and Metabolic Dysregulation in Largemouth Bass (Micropterus salmoides) after Ammonia Exposure" Metabolites 13, no. 2: 274. https://doi.org/10.3390/metabo13020274