
Antifungal Effect of Brassica Tissues on the Mycotoxigenic Cereal Pathogen Fusarium graminearum


Abstract
:1. Introduction
2. Results
2.1. Glucosinolate Content of Brassica Leaf Tissue
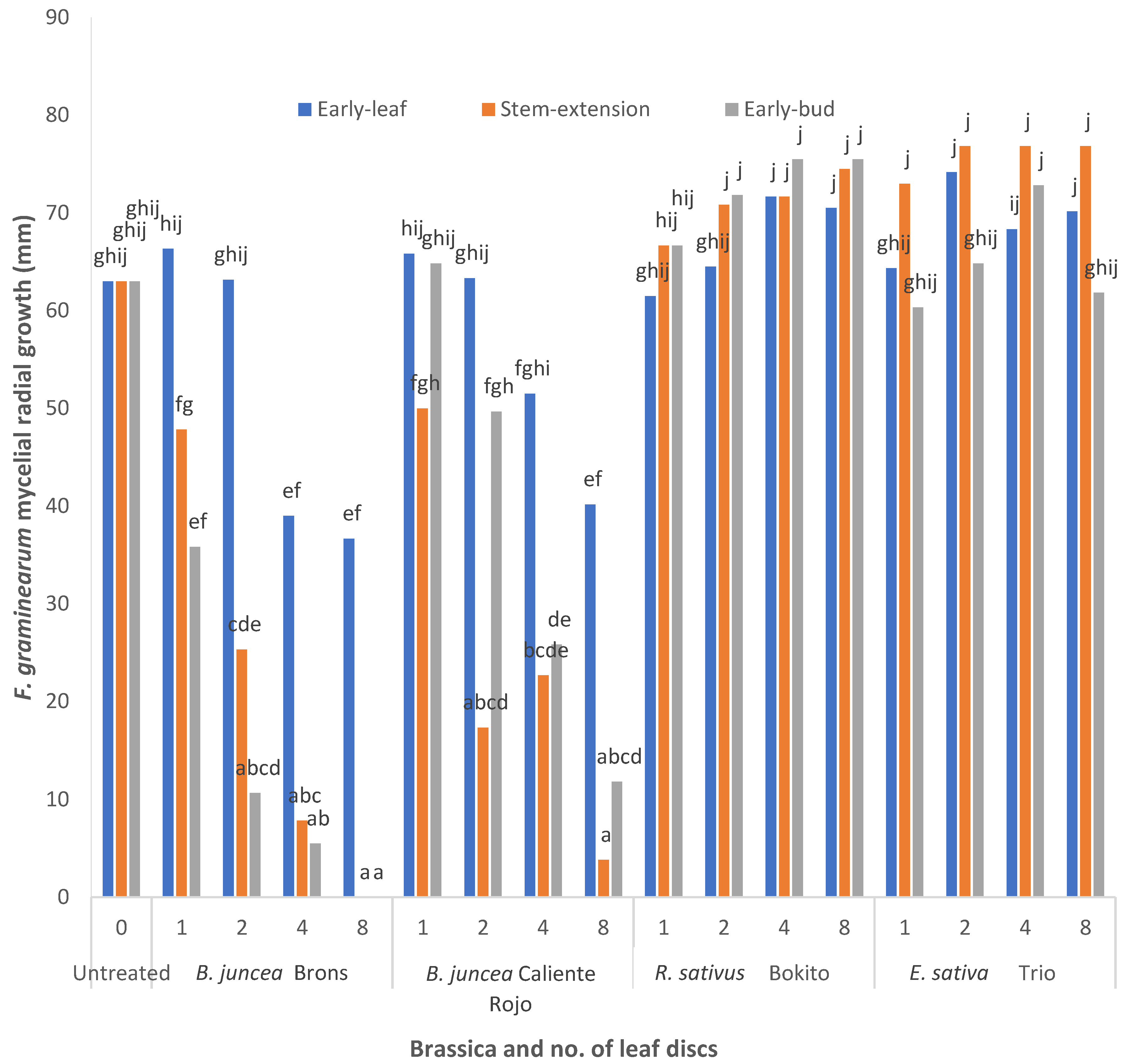
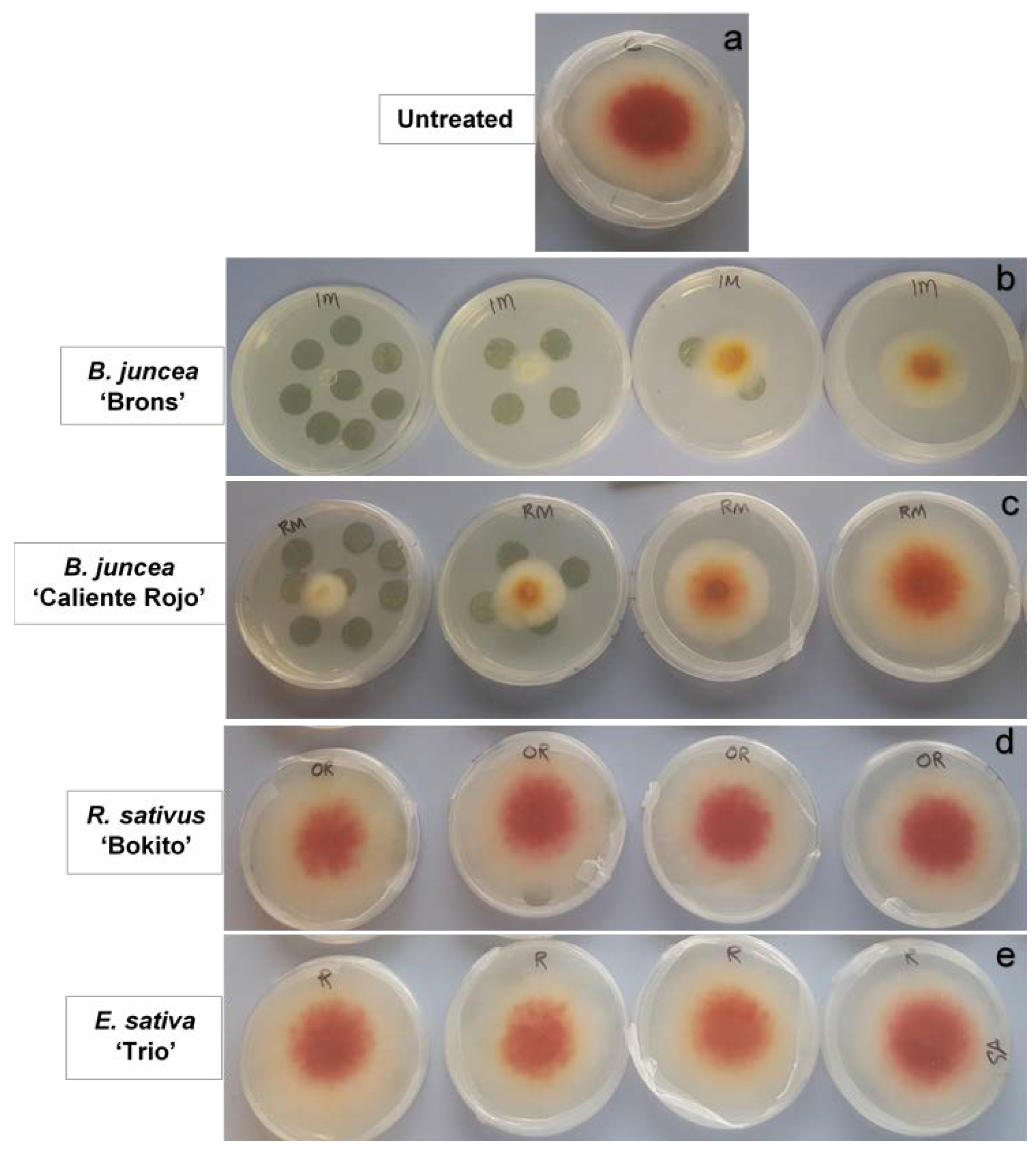
2.2. Effect of Brassica Leaf Discs on Fusarium graminearum
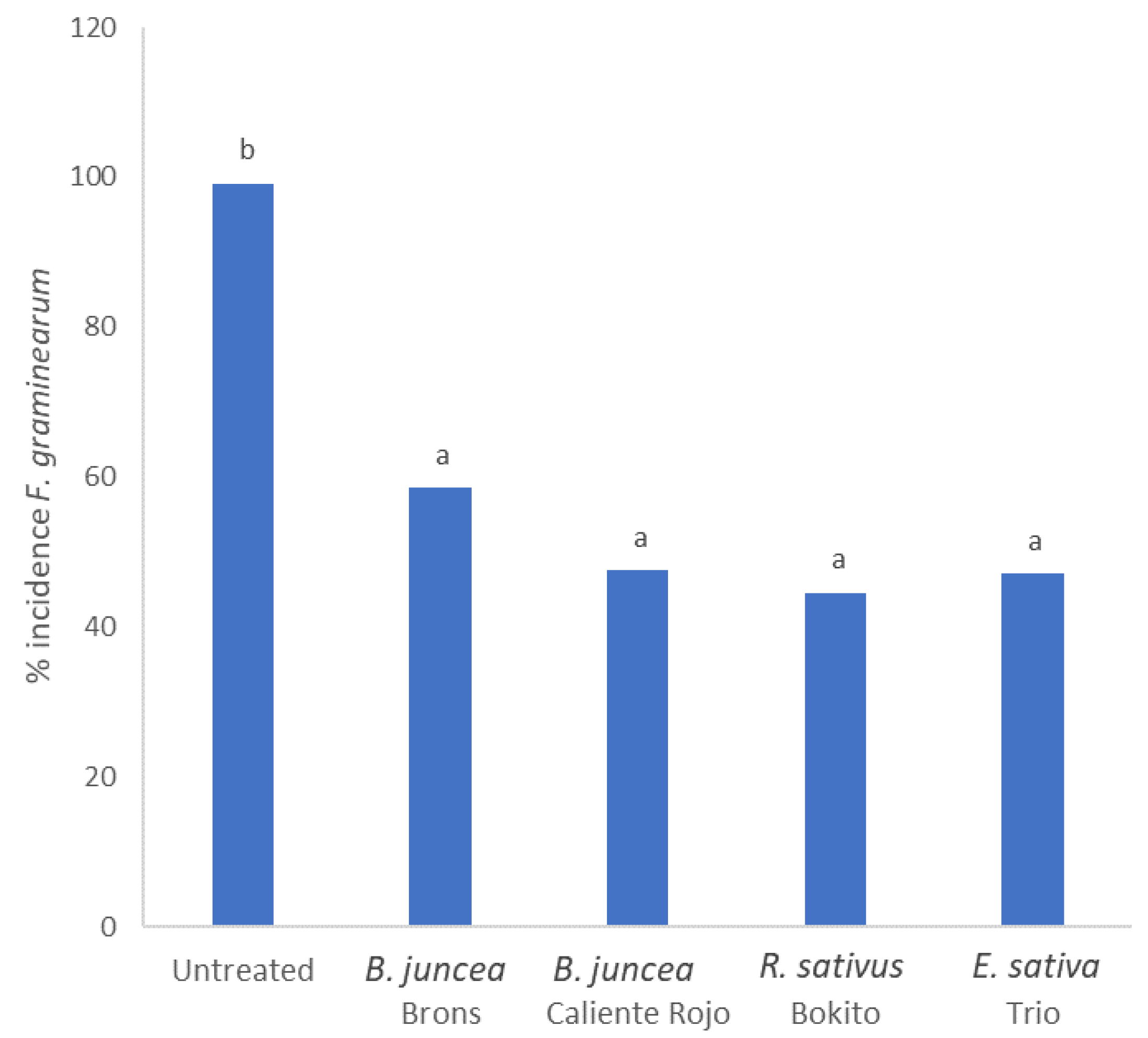
2.3. Biofumigation Effect of Brassicas in Closed Jar Experiment
3. Discussion
4. Materials and Methods
4.1. Brassicas and Fungal Culture
4.2. Leaf Disc Assay
4.3. Closed Jar Experiment
4.3.1. Preparation of Inoculum Bags
4.3.2. Evaluating Effect of Biofumigants on Inoculum
4.4. Glucosinolate Analysis
4.5. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Backhouse, D. Global distribution of Fusarium graminearum, F. asiaticum and F. boothii from wheat in relation to climate. Eur. J. Plant Pathol. 2014, 139, 161–173. [Google Scholar] [CrossRef]
- McMullen, M.; Bergstrom, G.; De Wolf, E.; Dill-Macky, R.; Hershman, D.; Shaner, G.; Van Sanford, D. A unified effort to fight an enemy of wheat and barley: Fusarium head blight. Plant Dis. 2012, 96, 1712–1728. [Google Scholar] [CrossRef] [PubMed]
- Gilchrist, L.; Dubin, H.J. Fusarium head blight. In Bread Wheat Improvement and Production, Plant Production and Protection; Curtis, B.C., Rajaram, S., Macpherson, H.G., Eds.; Series 30; FAO: Rome, Italy, 2002. [Google Scholar]
- Mielniczuk, E.; Skwaryło-Bednarz, B. Fusarium Head Blight, mycotoxins and strategies for their reduction. Agronomy 2020, 10, 509. [Google Scholar] [CrossRef]
- Palazzini, J.; Fumero, V.; Yerkovich, N.; Barros, G.; Cuniberti, M.; Chulze, S. Correlation between Fusarium graminearum and deoxynivalenol during the 2012/13 wheat Fusarium head blight outbreak in Argentina. Cereal Res. Commun. 2015, 43, 627–637. [Google Scholar] [CrossRef]
- Wilson, W.; Dahl, B.; Nganje, W. Economic costs of Fusarium Head Blight, scab and deoxynivalenol. World Mycotoxin J. 2018, 11, 291–302. [Google Scholar] [CrossRef]
- Guo, H.; Ji, J.; Wang, J.S.; Sun, X. Deoxynivalenol: Masked forms, fate during food processing, and potential biological remedies. Compr. Rev. Food Sci. Food Saf. 2020, 19, 895–926. [Google Scholar] [CrossRef] [PubMed]
- Szuets, P.; Mesterhazy, A.; Falkay, G.Y.; Bartok, T. Early telarche symptoms in children and their relations to zearalenon contamination in foodstuffs. Cereal Res. Commun. 1997, 25, 429–436. [Google Scholar] [CrossRef]
- Zhao, F.; Li, R.; Xiao, S.; Diao, H.; Viveiros, M.M.; Song, X.; Ye, X. Postweaning exposure to dietary zearalenone, a mycotoxin, promotes premature onset of puberty and disrupts early pregnancy events in female mice. Toxicol. Sci. 2013, 132, 431–442. [Google Scholar] [CrossRef]
- Edwards, S.G.; Jennings, P. Impact of agronomic factors on Fusarium mycotoxins in harvested wheat. Food Addit. Contam. Part A 2018, 35, 2443–2454. [Google Scholar] [CrossRef]
- Bensassi, F.; Zaied, C.; Abid, S.; Hajlaoui, M.R.; Bacha, H. Occurrence of deoxynivalenol in durum wheat in Tunisia. Food Control. 2010, 21, 281–285. [Google Scholar] [CrossRef]
- Jakšić, S.; Abramović, B.; Jajić, I.; Baloš, M.Ž.; Mihaljev, Ž.; Despotović, V.; Šojić, D. Co-occurrence of fumonisins and deoxynivalenol in wheat and maize harvested in Serbia. Bull. Environ. Contam. Toxicol. 2012, 89, 615–619. [Google Scholar] [CrossRef] [PubMed]
- Becher, R.; Hettwer, U.; Karlovsky, P.; Deising, H.B.; Wirsel, S.G.R. Adaptation of Fusarium graminearum to tebuconazole yielded descendants diverging for levels of fitness, fungicide resistance, virulence, and mycotoxin production. Phytopathology 2010, 100, 444–453. [Google Scholar] [CrossRef] [PubMed]
- Klix, M.B.; Verreet, J.-A.; Beyer, M. Comparison of the declining triazole sensitivity of Gibberella zeae and increased sensitivity achieved by advances in triazole fungicide development. Crop Prot. 2007, 26, 683–690. [Google Scholar] [CrossRef]
- Yerkovich, N.; Cantoro, R.; Palazzini, J.M.; Torres, A.; Chulze, S.N. Fusarium head blight in Argentina: Pathogen aggressiveness, triazole tolerance and biocontrol-cultivar combined strategy to reduce disease and deoxynivalenol in wheat. Crop Prot. 2020, 137, 105300. [Google Scholar] [CrossRef]
- Poulsen, R.; Luong, X.; Hansen, M.; Styrishave, B.; Hayes, T. Tebuconazole disrupts steroidogenesis in Xenopus laevis. Aquat. Toxicol. 2015, 168, 28–37. [Google Scholar] [CrossRef]
- Lv, X.; Pan, L.; Wang, J.; Lu, L.; Yan, W.; Zhu, Y.; Xu, Y.; Guo, M.; Zhuang, S. Effects of triazole fungicides on androgenic disruption and CYP3A4 enzyme activity. Environ. Pollut. 2017, 222, 504–512. [Google Scholar] [CrossRef]
- Kirkegaard, J.A.; Gardner, P.A.; Desmarchelier, J.M.; Angus, J.F. Biofumigation using Brassica species to control pests and diseases in horticulture and agriculture. In 9th Australian Research Assembly on Brassicas; Wratten, M., Mailer, R.J., Eds.; British Society for Plant Pathology, Agricultural Research Institute: Wagga Wagga, NSW, Australia, 1993; pp. 77–82. [Google Scholar]
- Andréasson, E.; Jørgensen, L.B.; Höglund, A.-S.; Rask, L.; Meijer, J. Different myrosinase and idioblast distribution in Arabidopsis and Brassica napus. Plant Physiol. 2001, 127, 1750–1763. [Google Scholar] [CrossRef]
- Thangstad, O.P.; Iversen, T.H.; Slupphaug, G.; Bones, A. Immunocytochemical localization of myrosinase in Brassica napus L. Planta 1990, 180, 245–248. [Google Scholar] [CrossRef]
- Koroleva, O.A.; Gibson, T.M.; Cramer, R.; Stain, C. Glucosinolate-accumulating S-cells in Arabidopsis leaves and flower stalks undergo programmed cell death at early stages of differentiation. Plant J. 2010, 64, 456–469. [Google Scholar] [CrossRef]
- Jørgensen, M.E.; Nour-Eldin, H.H.; Halkier, B.A. Transport of defense compounds from source to sink: Lessons learned from glucosinolates. Trends Plant Sci. 2015, 20, 508–514. [Google Scholar] [CrossRef]
- Bones, A.M.; Rossiter, J.T. The myrosinase-glucosinolate system, its organisation and biochemistry. Physiol. Plant 1996, 97, 194–208. [Google Scholar] [CrossRef]
- Wittstock, U.; Burow, M. Glucosinolate breakdown in Arabidopsis: Mechanism, regulation and biological significance. Arab. Book 2010, 8, e0134. [Google Scholar] [CrossRef]
- Dahlin, P.; Hallmann, J. New insights on the role of allyl isothiocyanate in controlling the root knot nematode Meloidogyne hapla. Plants 2020, 9, 603. [Google Scholar] [CrossRef] [PubMed]
- Norsworthy, J.K.; Meehan, J.T. Use of isothiocyanates for suppression of Palmer amaranth (Amaranthus palmeri), pitted morningglory (Ipomoea lacunosa), and yellow nutsedge (Cyperus esculentus). Weed Sci. 2005, 53, 884–890. [Google Scholar] [CrossRef]
- Sarwar, M.; Kirkegaard, J.A.; Wong, P.T.W.; Desmarchelier, J.M. Biofumigation potential of brassicas. 3. In vitro toxicity of isothiocyanates to soil-borne fungal pathogens. Plant Soil 1998, 201, 71–89. [Google Scholar]
- Smolinska, U.; Morra, M.J.; Knudsen, G.R.; James, R.L. Isothiocyanates produced by Brassicaceae species as inhibitors of Fusarium oxysporum. Plant Dis. 2003, 87, 407–412. [Google Scholar] [CrossRef]
- Lewis, J.A.; Papavizas, G.C. Effect of volatiles from decomposing plant tissues on pigmentation, growth, and survival of Rhizoctonia solani. Soil Sci. 1974, 118, 156–163. [Google Scholar] [CrossRef]
- Charron, C.S.; Sams, C.E. Inhibition of Pythium ultimum and Rhizoctonia solani by shredded leaves of Brassica species. J. Am. Soc. Hortic. Sci. 1999, 124, 462–467. [Google Scholar] [CrossRef]
- Neubauer, C.; Heitmann, B.; Müller, C. Biofumigation potential of Brassicaceae cultivars to Verticillium dahliae. Eur. J. Plant Pathol. 2014, 140, 341–352. [Google Scholar] [CrossRef]
- Handiseni, M.; Jo, Y.-K.; Lee, K.-M.; Zhou, X.-G. Screening brassicaceous plants as biofumigants for management of Rhizoctonia solani AG1-IA. Plant Dis. 2016, 100, 758–763. [Google Scholar] [CrossRef]
- Leplat, J.; Friberg, H.; Abid, M.; Steinberg, C. Survival of Fusarium graminearum, the causal agent of Fusarium head blight. A review. Agron. Sustainable Dev. 2013, 33, 97–111. [Google Scholar] [CrossRef]
- Ashiq, S.; Edwards, S.G.; Fatukasi, O.; Watson, A.; Back, M.A. In vitro activity of isothiocyanates against Fusarium graminearum. Plant Pathol. 2022, 71, 594–601. [Google Scholar] [CrossRef]
- Matteo, R. Non-Food Brassicas for Green Chemistry Purposes through a Biorefinery Approach. Master’s Thesis, University of Bologna, Bologna, Italy, 2017. [Google Scholar]
- Ngala, B.M.; Woods, S.R.; Back, M.A. In vitro assessment of the effects of Brassica juncea and Raphanus sativus leaf and root extracts on the viability of Globodera pallida encysted eggs. Nematology 2015, 17, 543–556. [Google Scholar] [CrossRef]
- Mayton, H.S.; Olivier, C.; Vaughn, S.F.; Loria, R. Correlation of fungicidal activity of Brassica species with allyl isothiocyanate production in macerated leaf tissue. Phytopathology 1996, 86, 267–271. [Google Scholar] [CrossRef]
- Ojaghian, M.R.; Jiang, H.; Xie, G.-L.; Cui, Z.-Q.; Zhang, J.; Li, B. In vitro biofumigation of Brassica tissues against potato stem rot caused by Sclerotinia sclerotiorum. Plant Pathol. J. 2012, 28, 185–190. [Google Scholar] [CrossRef]
- Larkin, R.P.; Griffin, T.S. Control of soilborne potato diseases using Brassica green manures. Crop Prot. 2007, 26, 1067–1077. [Google Scholar] [CrossRef]
- Kirkegaard, J.A.; Wong, P.T.W.; Desmarchelier, J.M. In vitro suppression of fungal root pathogens of cereals by Brassica tissues. Plant Pathol. 1996, 45, 593–603. [Google Scholar] [CrossRef]
- Morra, M.J.; Kirkegaard, J.A. Isothiocyanate release from soil-incorporated Brassica tissues. Soil Biol. Biochem. 2002, 34, 1683–1690. [Google Scholar] [CrossRef]
- Calmes, B.; N’Guyen, G.; Dumur, J.; Brisach, C.A.; Campion, C.; Iacomi, B.; Pigné, S.; Dias, E.; Macherel, D.; Guillemette, T. Glucosinolate-derived isothiocyanates impact mitochondrial function in fungal cells and elicit an oxidative stress response necessary for growth recovery. Front. Plant Sci. 2015, 6, 414. [Google Scholar] [CrossRef]
- Wang, T.; Li, Y.; Bi, Y.; Zhang, M.; Zhang, T.; Zheng, X.; Dong, Y.; Huang, Y. Benzyl isothiocyanate fumigation inhibits growth, membrane integrity and mycotoxin production in Alternaria alternata. RSC Adv. 2020, 10, 1829–1837. [Google Scholar] [CrossRef]
- Zhang, M.; Li, Y.; Bi, Y.; Wang, T.; Dong, Y.; Yang, Q.; Zhang, T. 2-Phenylethyl isothiocyanate exerts antifungal activity against Alternaria alternata by affecting membrane integrity and mycotoxin production. Toxins 2020, 12, 124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Björkman, M.; Klingen, I.; Birch, A.N.E.; Bones, A.M.; Bruce, T.J.A.; Johansen, T.J.; Meadow, R.; Mølmann, J.; Seljåsen, R.; Smart, L.E. Phytochemicals of Brassicaceae in plant protection and human health–Influences of climate, environment and agronomic practice. Phytochem. 2011, 72, 538–556. [Google Scholar] [CrossRef] [PubMed]
- Peterson, C.J.; Cosse, A.; Coats, J.R. Insecticidal components in the meal of Crambe abyssinica. J. Agric. Urban Entomol. 2000, 17, 27. [Google Scholar]
- Tsao, R.; Peterson, C.J.; Coats, J.R. Glucosinolate breakdown products as insect fumigants and their effect on carbon dioxide emission of insects. BMC Ecol. 2002, 2, 5. [Google Scholar] [CrossRef]
- Bending, G.D.; Lincoln, S.D. Characterisation of volatile sulphur-containing compounds produced during decomposition of Brassica juncea tissues in soil. Soil Biol. Biochem. 1999, 31, 695–703. [Google Scholar] [CrossRef]
- Motisi, N.; Doré, T.; Lucas, P.; Montfort, F. Dealing with the variability in biofumigation efficacy through an epidemiological framework. Soil Biol. Biochem. 2010, 42, 2044–2057. [Google Scholar] [CrossRef]
- Lazzeri, L.; Leoni, O.; Bernardi, R.; Malaguti, L.; Cinti, S. Plants, techniques and products for optimising biofumigation in full field. Agroindustria 2004, 3, 281–288. [Google Scholar]
- Drakopoulos, D.; Kägi, A.; Gimeno, A.; Six, J.; Jenny, E.; Forrer, H.-R.; Musa, T.; Meca, G.; Vogelgsang, S. Prevention of Fusarium head blight infection and mycotoxins in wheat with cut-and-carry biofumigation and botanicals. Field Crops Res. 2020, 246, 107681. [Google Scholar] [CrossRef]
- Champeil, A.; Fourbet, J.-F.; Doré, T.; Rossignol, L. Influence of cropping system on Fusarium head blight and mycotoxin levels in winter wheat. Crop Prot. 2004, 23, 531–537. [Google Scholar] [CrossRef]
- Vogelgsang, S.; Beyer, M.; Pasquali, M.; Jenny, E.; Musa, T.; Bucheli, T.D.; Wettstein, F.E.; Forrer, H.-R. An eight-year survey of wheat shows distinctive effects of cropping factors on different Fusarium species and associated mycotoxins. Eur. J. Agron. 2019, 105, 62–77. [Google Scholar] [CrossRef]
- Ngala, B.M.; Haydock, P.P.J.; Woods, S.; Back, M.A. Biofumigation with Brassica juncea, Raphanus sativus and Eruca sativa for the management of field populations of the potato cyst nematode Globodera pallida. Pest Manag. Sci. 2015, 71, 759–769. [Google Scholar] [CrossRef] [PubMed]
- Waalwijk, C.; van der Heide, R.; de Vries, I.; van der Lee, T.; Schoen, C.; Costrel-de Corainville, G.; Häuser-Hahn, I.; Kastelein, P.; Köhl, J.; Lonnet, P. Quantitative detection of Fusarium species in wheat using TaqMan. Eur. J. Plant Pathol. 2004, 110, 481–494. [Google Scholar] [CrossRef]
- Edwards, S.G.; Seddon, B. Selective media for the specific isolation and enumeration of Botrytis cinerea conidia. Lett. Appl. Microbiol. 2001, 32, 63–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hofgaard, I.S.; Seehusen, T.; Aamot, H.U.; Riley, H.; Razzaghian, J.; Le, V.H.; Hjelkrem, A.-G.R.; Dill-Macky, R.; Brodal, G. Inoculum potential of Fusarium spp. relates to tillage and straw management in Norwegian fields of spring oats. Front. Microbiol. 2016, 7, 556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leslie, J.F.; Summerell, B.A. The Fusarium Laboratory Manual; Blackwell Publishing: Ames, IA, USA, 2006. [Google Scholar]
- ISO 9167 Rapeseed and Rapeseed Meals—Determination of Glucosinolates Content—Method Using HPLC. Available online: https://www.iso.org/standard/72207.html (accessed on 21 March 2022).





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Glucosinolate | Brassica juncea Brons | Brassica juncea Caliente Rojo | Raphanus sativus Bokito | Eruca sativa Trio | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| I a | II b | III c | I | II | III | I | II | III | I | II | III | |
| Glucoberin | 2.47 (0.6) d | 3.15 (0.49) | 2.01 (0.41) | 1.29 (0.2) | 1.81 (0.3) | 1.33 (0.37) | 2.28 (0.4) | 1.16 (0.18) | 2.15 (0.6) | 2.54 (0.6) | 2.55 (0.33) | 2.37 (0.75) |
| Progoitrin | 0.38 (0.1) | 0.34 (0.02) | 0.30 (0.10) | 0.25 (0.04) | 0.23 (0.11) | 0.26 (0.09) | 0.35 (0.08) | 0.37 (0.12) | 0.46 (0.08) | 0.30 (0.1) | 0.00 | 0.00 |
| Sinigrin | 21.44 (1.7) | 24.02 (2.94) | 48.52 (3.03) | 22.72 (1.88) | 59.54 (2.32) | 50.9 (3.8) | - | - | - | - | - | - |
| Gluconapin | 0.00 | 0.00 | 0.35 (0.09) | 0.27 (0.07) | 0.35 (0.03) | 0.25 (0.09) | - | - | - | - | - | - |
| Glucobrassicin | 0.14 (0.04) | 0.03 (0.01) | 0.11 (0.02) | 0.11 (0.02) | 0.14 (0.02) | 0.23 (0.12) | 1.62 (0.26) | 3.87 (1.9) | 2.62 (0.6) | 0.22 (0.03) | 0.54 (0.08) | 0.58 (0.08) |
| Gluconasturtiin | 0.11 (0.01) | 0.39 (0.05) | 0.58 (0.08) | 0.24 (0.08) | 0.97 (0.19) | 0.76 (0.12) | 0.32 (0.13) | 1.13 (0.2) | 0.96 (0.2) | 0.77 (0.12) | 0.42 (0.15) | 0.24 (0.10) |
| Neoglucobrassicin | 0.00 | 0.00 | 0.00 | 0.05 (0.01) | 0.04 (0.01) | 0.01 (0.007) | 0.00 | 0.00 | 0.00 | 0.00 | 0.06 (0.02) | 0.00 |
| Glucoraphanin | - | - | - | - | - | - | 2.90 (0.5) | 23.56 (3.2) | 7.29 (2.1) | 2.42 (0.51) | 2.84 (0.64) | 2.75 (0.6) |
| Glucoraphenin | - | - | - | - | - | - | 0.18 (0.05) | 1.34 (0.4) | 0.44 (0.07) | - | - | - |
| 4 hydroxy glucobrassicin | - | - | - | 0.05 (0.01) | 0.37 (0.1) | 0.4 (0.15) | 0.00 | 0.04 (0.01) | 0.03 (0.01) | - | - | - |
| Glucoraphasatin | - | - | - | - | - | - | 1.02 (0.03) | 8.10 (1.1) | 11.09 (3.1) | - | - | - |
| Glucoalyssin | - | - | - | - | - | - | - | - | - | 0.86 (0.18) | 0.51 (0.2) | 0.45 (0.14) |
| Glucoerucin | - | - | - | - | - | - | - | - | - | 0.57 (0.14) | 1.20 (0.01) | 1.50 (0.28) |
| 4-mercaptobutyl | - | - | - | - | - | - | - | - | - | 3.58 (0.72) | 5.70 (0.44) | 3.21 (0.58) |
| unknown | - | - | - | - | - | - | - | - | - | 0.61 (0.2) | 0.83 (0.23) | 0.69 (0.19) |
| unknown | - | - | - | - | - | - | - | - | - | 1.04 (0.12) | 2.59 (0.66) | 2.83 (0.74) |
| Total glucosinolates | 24.54 (2.26) | 27.94 (4.18) | 51.87 (3.34) | 24.98 (4.59) | 63.45 (3.46) | 54.15 (2.94) | 8.67 (2.33) | 39.56 (1.25) | 25.05 (3.69) | 12.90 (1.20) | 17.23 (2.47) | 14.63 (2.57) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ashiq, S.; Edwards, S.; Watson, A.; Blundell, E.; Back, M. Antifungal Effect of Brassica Tissues on the Mycotoxigenic Cereal Pathogen Fusarium graminearum. Antibiotics 2022, 11, 1249. https://doi.org/10.3390/antibiotics11091249
Ashiq S, Edwards S, Watson A, Blundell E, Back M. Antifungal Effect of Brassica Tissues on the Mycotoxigenic Cereal Pathogen Fusarium graminearum. Antibiotics. 2022; 11(9):1249. https://doi.org/10.3390/antibiotics11091249
Chicago/Turabian StyleAshiq, Samina, Simon Edwards, Andrew Watson, Emma Blundell, and Matthew Back. 2022. "Antifungal Effect of Brassica Tissues on the Mycotoxigenic Cereal Pathogen Fusarium graminearum" Antibiotics 11, no. 9: 1249. https://doi.org/10.3390/antibiotics11091249