
Naringenin and Its Derivatives—Health-Promoting Phytobiotic against Resistant Bacteria and Fungi in Humans

 , and
, and
Abstract
:1. Introduction
2. Content of Naringenin and its Glycosides in Plants and Products of Plant Origin
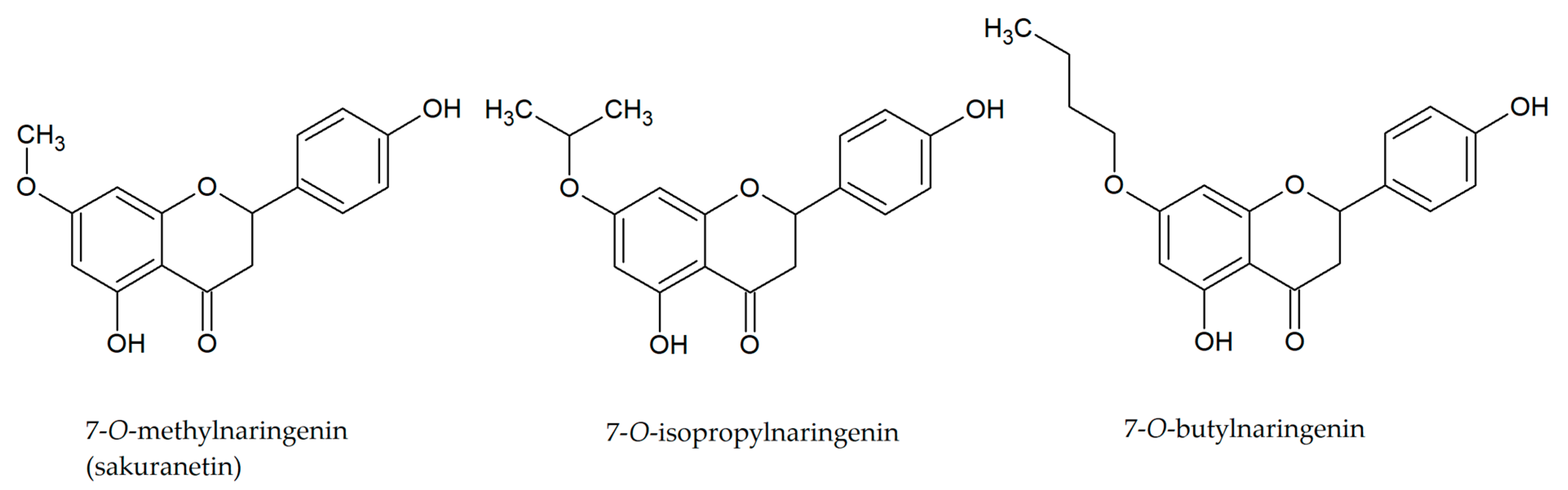
3. Other Derivatives of Naringenin
4. Properties of Naringenin
4.1. Anti-Cancer Activity
4.2. Anti-Inflammatory Activity
4.3. Antioxidant Activity
4.4. Effects on the Nervous System
4.5. Antidiabetic Activity
4.6. Hepatoprotective Properties
4.7. Antimicrobial Activity of Naringenin
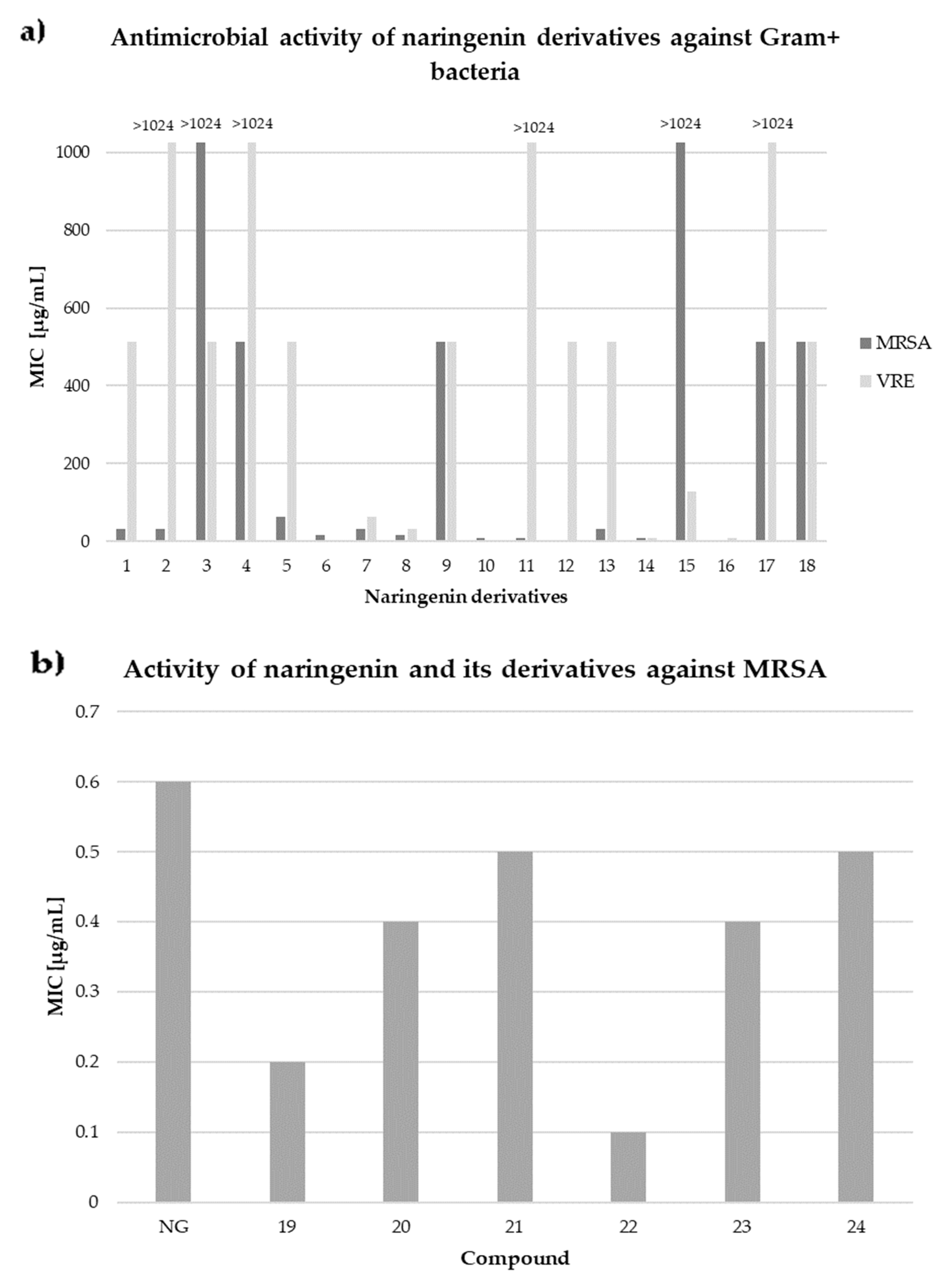
5. Antimicrobial Activity against Gram-Positive Bacteria
6. Antimicrobial Activity against Gram-Negative Bacteria

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Activity of Naringenin | Reference |
|---|---|---|
| P. aeruginosa c.i. | 128 µg/mL | [86] |
| E. coli ATCC 8739 | 1000 µg/mL | [87] |
| E. coli ATCC 11775 | 1000 µg/mL | |
| H. pylori ATCC43504 | 80 µg/mL | [82] |
| H. pylori NCTC11637 | 40 µg/mL | |
| H. pylori NCTC11638 | 40 µg/mL | |
| H. pylori 82516 c.i.g | 40 µg/mL | |
| H. pylori 82548 c.i.g | 40 µg/mL | |
| H. pylori 4 c.i.g | 100 µg/mL | |
| E. coli ATCC 25922 | 1000 µg/mL | [65] |
| E. cloacae DMST 21394 | >512 µg/mL | [88] |
| E. cloacae DMST 21549 | >512 µg/mL | |
| E. cloacae DMST 19719 | >512 µg/mL | |
| E. coli ATCC 25922 | >512 µg/mL | |
| K. pneumoniae ATCC 13883 | 0.5–1 µg/mL | [89] |
| H. pylori ATCC 43504 | 100 µg/mL | [90] |
| H. pylori ATCC 51932 | 100 µg/mL | |
| H. pylori OX.22 c.i. | 100 µg/mL | |
| H. pylori OX. 63 c.i. | 100 µg/mL | |
| H. pylori OX.64 c.i. | 100 µg/mL | |
| H. pylori OX.67 c.i. | 100 µg/mL | |
| H. pylori OX.83 c.i. | 100 µg/mL | |
| Bacteroides galacturonicus DSM 3978 | 250 µg/mL | [91] |
| Escherichia coli DSM 1116 | >250 µg/mL | |
| H. pylori ATCC 43 504 | 100 µg/mL | [92] |
| E. coli ATCC 31030 | 550 µg/mL | [79] |
| P. mirabilis ATCC 25933 | 550 µg/mL | |
| P. aeruginosa ATCC 10145 | 500 µg/mL | |
| S. enterica subsp. enterica serovar Paratyphi B(S-3) | 600 µg/mL | |
| S. enterica subsp. enterica serovar Typhi D(S-58) | 400 µg/mL | |
| S. enterica subsp. enterica serovar Typhimurium ATCC 14028 | 600 µg/mL | |
| S. sonnei D1, L1, S2 ATCC 9290 | 450 µg/mL | |
| S. marcescens ATCC 11105 | 500 µg/mL | |
| S. boydii C2, L1, S2 ATCC 8700 | 100 µg/mL | |
| S. marcescens ATCC 27117 | 400 µg/mL | [93] |
| E. coli ATCC 25922 | >2000 µg/mL | [94] |
| K. pneumoniae ATCC 13883 | 2000 µg/mL | |
| P. mirabilis ATCC 43071 | 2000 µg/mL | |
| P. aeruginosa ATCC 27857 | >2000 µg/mL | |
| S. enterica subsp. enterica serovar Typhimurium ATCC 14028 | >2000 µg/mL | |
| E. coli MTCC 1652 | 12.5 µg/mL | [95] |
| P. aeruginosa MTCC 424 | 23.5 µg/mL | |
| E. coli K-12 MG1655 | 800 µg/mL | [96] |
| S. enterica subsp. enterica serovar Typhimurium LT2 | 1000 µg/mL | |
| P. putida ATCC 795 | 1000 µg/mL | |
| S. enterica subsp. enterica serovar Typhimurium ATCC 14028 | 250 µg/mL | [97] |
| E. coli 916 c.i.g | 125 µg/mL | [98] |
| S. enterica subsp. enterica serovar Typhimurium 450 c.i.g | 125 µg/mL | |
| E. coli ATCC 25922 | 400 µg/mL | [99] |
| Strain | Activity of Naringin | Reference |
| P. aeruginosa ATCC 9027 | 1000 µg/mL | [87] |
| H. pylori ATCC43504 | >100 µg/mL | [82] |
| H. pylori NCTC11637 | >100 µg/mL | |
| H. pylori NCTC11638 | >100 µg/mL | |
| H. pylori 82516 c.i.g | >100 µg/mL | |
| H. pylori 82548 c.i.g | >100 µg/mL | |
| H. pylori 4 c.i.g | >100 µg/mL | |
| H. pylori ATCC 43 504 | >100 µg/mL | [92] |
| E. coli c.i. | 500 µg/mL | [100] |
| P. aeruginosa c.i. | 500 µg/mL | |
| E. coli ATCC 31030 | 900 µg/mL | [93] |
| P. mirabilis ATCC 25933 | 700 µg/mL | |
| P. aeruginosa ATCC 10145 | 600 µg/mL | |
| S. enterica subsp. enterica serovar Typhimurium ATCC 14028 | 800 µg/mL | |
| S. marcescens ATCC 27117 | 600 µg/mL | |
| E. coli ATCC 25922 | >2000 µg/mL | [101] |
| E. coli K-12 MG1655 | >1000 µg/mL | [96] |
| S. enterica subsp. enterica serovar Typhimurium LT2 | >1000 µg/mL | |
| P. putida ATCC 795 | >1000 µg/mL |
7. Antifungal Activity
| Compound | Strain | Activity | Reference |
|---|---|---|---|
 | Candida albicans ATCC 90028 | 5 μM | [107] |
| Candida parapsilosis ATCC 22019 | 5 μM | ||
| Trichosporon beigelii KCTC 7707 | 2,5 μM | ||
| Malassezia furfur KCTC 7743 | 10 μM | ||
| Trichophyton rubrum KCTC 6345 | 5 μM | ||
| Aspergillus flavus KCTC 6905 | 10 μM | ||
| Saccharomyces cerevisiae KCTC 7296 | 5 μM | ||
| Candida albicans ATCC 10231 | 40 μg/mL | [102] | |
| Candida albicans 19–22 | 40 μg/mL | ||
 | Candida albicans ATCC 10231 | 16 mg/mL | [108] |
| Candida kruzei ATCC 6258 | 32 mg/mL | ||
 | Aspergillus niger 439 | 100 μg/mL | [109] |
| Fusarium oxysporum (M42) | n.i.e. | ||
| Candida albicans (N/A) | n.i.e. | ||
| Saccharomyces cerevisiae (N/A) | n.i.e. | ||
 | Candida albicans MTCC 183 | 22 μg/mL | [80] |
| Aspergillus niger MTCC9687 | 20 μg/mL | ||
 | Candida albicans MTCC 183 | 18 μg/mL | [80] |
| Aspergillus niger MTCC9687 | 18 μg/mL | ||
 | Candida albicans MTCC 183 | 24 μg/mL | [80] |
| Aspergillus niger MTCC9687 | 20 μg/mL | ||
 | Candida albicans MTCC 183 | 22 μg/mL | [80] |
| Aspergillus niger MTCC9687 | 22 μg/mL | ||
 | Candida albicans MTCC 183 | 22 μg/mL | [80] |
| Aspergillus niger MTCC9687 | 24 μg/mL | ||
 | Candida albicans MTCC 183 | 16 μg/mL | [80] |
| Aspergillus niger MTCC9687 | 16 μg/mL |
8. The Mechanism of Action of Naringenin on Bacteria and Fungi
9. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Shakeel, S.; Rehman, M.U.; Tabassum, N.; Amin, U.; Mir, M. Effect of Naringenin (A Naturally Occurring Flavanone) against Pilocarpine-Induced Status Epilepticus and Oxidative Stress in Mice. Pharmacogn. Mag. 2017, 13, S154–S160. [Google Scholar] [CrossRef] [PubMed]
- Amin, I.; Majid, S.; Farooq, A.; Wani, H.A.; Noor, F.; Khan, R.; Shakeel, S.; Bhat, S.A.; Ahmad, A.; Madkhali, H.; et al. Naringenin (4,5,7-Trihydroxyflavanone) as a Potent Neuroprotective Agent: From Chemistry to Medicine, 1st ed.; Elsevier B.V.: Amsterdam, The Netherlands, 2020; Volume 65, ISBN 9780128179055. [Google Scholar]
- Rashed, K. International Journal of Biomedical and Advance Research Biological Properties of Naringenin: A Review QR Code. Int. J. Biomed. Adv. Res. 2021, 12, 12. [Google Scholar] [CrossRef]
- Chung, T.W.; Li, S.; Lin, C.C.; Tsai, S.W. Antinociceptive and Anti-Inflammatory Effects of the Citrus Flavanone Naringenin. Tzu Chi Med. J. 2019, 31, 81–85. [Google Scholar] [CrossRef]
- Rauf, A.; Shariati, M.A.; Imran, M.; Bashir, K.; Khan, S.A.; Mitra, S.; Bin Emran, T.; Badalova, K.; Uddin, M.S.; Mubarak, M.S.; et al. Comprehensive Review on Naringenin and Naringin Polyphenols as a Potent Anticancer Agent. Environ. Sci. Pollut. Res. 2022, 29, 31025–31041. [Google Scholar] [CrossRef] [PubMed]
- Naeini, F.; Namkhah, Z.; Ostadrahimi, A.; Tutunchi, H.; Hosseinzadeh-Attar, M.J. A Comprehensive Systematic Review of the Effects of Naringenin, a Citrus-Derived Flavonoid, on Risk Factors for Nonalcoholic Fatty Liver Disease. Adv. Nutr. 2021, 12, 413–428. [Google Scholar] [CrossRef]
- Karim, N.; Jia, Z.; Zheng, X.; Cui, S.; Chen, W. A Recent Review of Citrus Flavanone Naringenin on Metabolic Diseases and Its Potential Sources for High Yield-Production. Trends Food Sci. Technol. 2018, 79, 35–54. [Google Scholar] [CrossRef]
- Salehi, B.; Fokou, P.V.T.; Sharifi-Rad, M.; Zucca, P.; Pezzani, R.; Martins, N.; Sharifi-Rad, J. The Therapeutic Potential of Naringenin: A Review of Clinical Trials. Pharmaceuticals 2019, 12, 11. [Google Scholar] [CrossRef] [Green Version]
- Hartogh, D.J.D.; Tsiani, E. Antidiabetic Properties of Naringenin: A Citrus Fruit Polyphenol. Biomolecules 2019, 9, 99. [Google Scholar] [CrossRef] [Green Version]
- McIntosh, C.A.; Mansell, R.L. Three-Dimensional Distribution of Limonin, Limonoate A-Ring Monolactone, and Naringin in the Fruit Tissues of Three Varieties of Citrus Paradisi. J. Agric. Food Chem. 1997, 45, 2876–2883. [Google Scholar] [CrossRef]
- Victor, M.M.; David, J.M.; Sakukuma, M.C.K.; França, E.L.; Nunes, A.V.J. A Simple and Efficient Process for the Extraction of Naringin from Grapefruit Peel Waste. Green Process. Synth. 2018, 7, 524–529. [Google Scholar] [CrossRef]
- Peterson, J.J.; Beecher, G.R.; Bhagwat, S.A.; Dwyer, J.T.; Gebhardt, S.E.; Haytowitz, D.B.; Holden, J.M. Flavanones in Grapefruit, Lemons, and Limes: A Compilation and Review of the Data from the Analytical Literature. J. Food Compos. Anal. 2006, 19, 74–80. [Google Scholar] [CrossRef]
- Ho, P.C.; Saville, D.J.; Coville, P.F.; Wanwimolruk, S. Content of CYP3A4 Inhibitors, Naringin, Naringenin and Bergapten in Grapefruit and Grapefruit Juice Products. Pharm. Acta Helv. 2000, 74, 379–385. [Google Scholar] [CrossRef] [PubMed]
- Sudto, K.; Pornpakakul, S.; Wanichwecharungruang, S. An Efficient Method for the Large Scale Isolation of Naringin from Pomelo (Citrus grandis) Peel. Int. J. Food Sci. Technol. 2009, 44, 1737–1742. [Google Scholar] [CrossRef]
- Lin, L.Y.; Huang, C.Y.; Chen, K.C.; Peng, R.Y. Pomelo Fruit Wastes Are Potentially Valuable Antioxidants, Anti-Inflammatories, Antihypertensives, and Antihyperglycemics. Hortic. Environ. Biotechnol. 2021, 62, 377–395. [Google Scholar] [CrossRef]
- Ni, H.; Zhang, S.F.; Gao, Q.F.; Hu, Y.; Jiang, Z.D.; Chen, F. Development and Evaluation of Simultaneous Quantification of Naringin, Prunin, Naringenin, and Limonin in Citrus Juice. Food Sci. Biotechnol. 2015, 24, 1239–1247. [Google Scholar] [CrossRef]
- Cerqueira, E.; Silva, L.C.R.; David, J.M.; Borges, R.D.S.Q.; Ferreira, S.L.C.; David, J.P.; Reis, P.S.D.; Bruns, R.E. Determination of Flavanones in Orange Juices Obtained from Different Sources by HPLC/DAD. J. Anal. Methods Chem. 2014, 2014, 13–18. [Google Scholar] [CrossRef]
- Yalim, S.; Özdemir, Y.; Ekiz, H.I. Naringin in Turkish Orange Juices and Its Reduction by Naringinase. J. Food Drug Anal. 2004, 12, 273–276. [Google Scholar] [CrossRef]
- de Lourdes Mata Bilbao, M.; Andrés-Lacueva, C.; Jáuregui, O.; Lamuela-Raventós, R.M. Determination of Flavonoids in a Citrus Fruit Extract by LC-DAD and LC-MS. Food Chem. 2007, 101, 1742–1747. [Google Scholar] [CrossRef]
- Dhuique-Mayer, C.; Caris-Veyrat, C.; Ollitrault, P.; Curk, F.; Amiot, M.J. Varietal and Interspecific Influence on Micronutrient Contents in Citrus from the Mediterranean Area. J. Agric. Food Chem. 2005, 53, 2140–2145. [Google Scholar] [CrossRef]
- Behiry, S.I.; Okla, M.K.; Alamri, S.A.; El-Hefny, M.; Salem, M.Z.; Alaraidh, I.A.; Ali, H.M.; Al-Ghtani, S.M.; Monroy, J.C.; Salem, A.Z. Antifungal and Antibacterial Activities of Musa paradisiaca, L. Peel Extract: HPLC Analysis of Phenolic and Flavonoid Contents. Processes 2019, 11, 215. [Google Scholar] [CrossRef]
- Dębski, H.; Wiczkowski, W.; Horbowicz, M. Effect of Elicitation with Iron Chelate and Sodium Metasilicate on Phenolic Compounds in Legume Sprouts. Molecules 2021, 26, 1345. [Google Scholar] [CrossRef] [PubMed]
- Saeed Alkaltham, M.; Musa Özcan, M.; Uslu, N.; Salamatullah, A.M.; Hayat, K. Effect of Microwave and Oven Roasting Methods on Total Phenol, Antioxidant Activity, Phenolic Compounds, and Fatty Acid Compositions of Coffee Beans. J. Food Process. Preserv. 2020, 44, e14874. [Google Scholar] [CrossRef]
- Paganga, G.; Miller, N.; Rice-Evans, C.A. The Polyphenolic Content of Fruit and Vegetables and Their Antioxidant Activities. What Does a Serving Constitute? Free Radic. Res. 1999, 30, 153–162. [Google Scholar] [CrossRef] [PubMed]
- Bugianesi, R.; Catasta, G.; Spigno, P.; D’Uva, A.; Maiani, G. Naringenin from Cooked Tomato Paste Is Bioavailable in Men. J. Nutr. 2002, 132, 3349–3352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sâmia Andricia, S.; da Silva, M.D.F.A.; Tavares, J.F.; Emídio, V.L. da-Cunha, J.M.B.-F.; Silva, M.S.D. Flavanones from Aerial Parts of Cordia Globosa (Jacq.) Kunth, Boraginaceae. Rev. Bras. Farmacogn. 2010, 20, 675–681. [Google Scholar] [CrossRef] [Green Version]
- Nobakht, M.; Grkovic, T.; Trueman, S.J.; Wallace, H.M.; Katouli, M.; Quinn, R.J.; Brooks, P.R. Chemical Constituents of Kino Extract from Corymbia Torelliana. Molecules 2014, 19, 17862–17871. [Google Scholar] [CrossRef] [Green Version]
- Suksamrarn, A.; Chotipong, A.; Suavansri, T.; Boongird, S.; Timsuksai, P.; Vimuttipong, S.; Chuaynugul, A. Antimycobacterial Activity and Cytotoxicity of Flavonoids from the Flowers of Chromolaena Odorata. Arch. Pharm. Res. 2004, 27, 507–511. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, T.; Lin, F.; Hasegawa, M.; Okada, K.; Nojiri, H.; Yamane, H. Purification and Identification of Naringenin 7-O-Methyltransferase, a Key Enzyme in Biosynthesis of Flavonoid Phytoalexin Sakuranetin in Rice. J. Biol. Chem. 2012, 287, 19315–19325. [Google Scholar] [CrossRef] [Green Version]
- Kozłowska, J.; Potaniec, B.; Zarowska, B.; Anioł, M. Synthesis and Biological Activity of Novel O-Alkyl Derivatives of Naringenin and Their Oximes. Molecules 2017, 22, 1485. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.A.; Moon, S.H.; Lee, J.Y.; Kim, K.T.; Park, Y.S.; Paik, H.D. Antibacterial Activity of a Novel Flavonoid, 7-O-Butyl Naringenin, against Methicillin-Resistant Staphylococcus Aureus (MRSA). Food Sci. Biotechnol. 2013, 22, 1725–1728. [Google Scholar] [CrossRef]
- Moon, S.H.; Lee, J.H.; Kim, K.T.; Park, Y.S.; Nah, S.Y.; Ahn, D.U.; Paik, H.D. Antimicrobial Effect of 7-O-Butylnaringenin, a Novel Flavonoid, and Various Natural Flavonoids against Helicobacter Pylori Strains. Int. J. Environ. Res. Public Health 2013, 10, 5459–5469. [Google Scholar] [CrossRef] [PubMed]
- Albuquerque de Oliveira Mendes, L.; Ponciano, C.S.; Depieri Cataneo, A.H.; Wowk, P.F.; Bordignon, J.; Silva, H.; Vieira de Almeida, M. The Anti-Zika Virus and Anti-Tumoral Activity of the Citrus Flavanone Lipophilic Naringenin-Based Compounds. Chem. Biol. Interact. 2020, 331, 109218. [Google Scholar] [CrossRef] [PubMed]
- Kozłowska, J.; Grela, E.; Baczynska, D.; Grabowiecka, A.; Anioł, M. Novel O-Alkyl Derivatives of Naringenin and Their Oximes with Antimicrobial and Anticancer Activity. Molecules 2019, 24, 679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chandrika, B.B.; Steephan, M.; Kumar, T.R.S.; Sabu, A.; Haridas, M. Hesperetin and Naringenin Sensitize HER2 Positive Cancer Cells to Death by Serving as HER2 Tyrosine Kinase Inhibitors. Life Sci. 2016, 160, 47–56. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Aquino, E.; Muriel, P. Beneficial Effects of Naringenin in Liver Diseases: Molecular Mechanisms. World J. Gastroenterol. 2018, 24, 1679–1707. [Google Scholar] [CrossRef]
- Arul, D.; Subramanian, P. Naringenin (Citrus Flavonone) Induces Growth Inhibition, Cell Cycle Arrest and Apoptosis in Human Hepatocellular Carcinoma Cells. Pathol. Oncol. Res. 2013, 19, 763–770. [Google Scholar] [CrossRef]
- Lim, W.; Park, S.; Bazer, F.W.; Song, G. Naringenin-Induced Apoptotic Cell Death in Prostate Cancer Cells Is Mediated via the PI3K/AKT and MAPK Signaling Pathways. J. Cell. Biochem. 2017, 118, 1118–1131. [Google Scholar] [CrossRef]
- Nasr Bouzaiene, N.; Chaabane, F.; Sassi, A.; Chekir-Ghedira, L.; Ghedira, K. Effect of Apigenin-7-Glucoside, Genkwanin and Naringenin on Tyrosinase Activity and Melanin Synthesis in B16F10 Melanoma Cells. Life Sci. 2016, 144, 80–85. [Google Scholar] [CrossRef]
- Stompor, M.; Uram, Ł.; Podgórski, R. In Vitro Effect of 8-Prenylnaringenin and Naringenin on Fibroblasts and Glioblastoma Cells-Cellular Accumulation and Cytotoxicity. Molecules 2017, 22, 1092. [Google Scholar] [CrossRef] [Green Version]
- Lu, W.L.; Yu, C.T.R.; Lien, H.M.; Sheu, G.T.; Cherng, S.H. Cytotoxicity of Naringenin Induces Bax-Mediated Mitochondrial Apoptosis in Human Lung Adenocarcinoma A549 Cells. Environ. Toxicol. 2020, 35, 1386–1394. [Google Scholar] [CrossRef]
- Shi, X.; Luo, X.; Chen, T.; Guo, W.; Liang, C.; Tang, S.; Mo, J. Naringenin Inhibits Migration, Invasion, Induces Apoptosis in Human Lung Cancer Cells and Arrests Tumour Progression in Vitro. J. Cell. Mol. Med. 2021, 25, 2563–2571. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.; Lee, D.H.; Jang, H.; Park, S.Y.; Seol, J.W. Naringenin Exerts Anticancer Effects by Inducing Tumor Cell Death and Inhibiting Angiogenesis in Malignant Melanoma. Int. J. Med. Sci. 2020, 17, 3049–3057. [Google Scholar] [CrossRef] [PubMed]
- Manchope, M.F.; Calixto-Campos, C.; Coelho-Silva, L.; Zarpelon, A.C.; Pinho-Ribeiro, F.A.; Georgetti, S.R.; Baracat, M.M.; Casagrande, R.; Verri, W.A. Naringenin Inhibits Superoxide Anion-Induced Inflammatory Pain: Role of Oxidative Stress, Cytokines, Nrf-2 and the No-CGMP-PKG-KATP Channel Signaling Pathway. PLoS ONE 2016, 11, e0153015. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Wang, N.; Fan, S.; Zheng, X.; Yang, Y.; Zhu, Y.; Lu, Y.; Chen, Q.; Zhou, H.; Zheng, J. The Citrus Flavonoid Naringenin Confers Protection in a Murine Endotoxaemia Model through AMPK-ATF3-Dependent Negative Regulation of the TLR4 Signalling Pathway. Sci. Rep. 2016, 6, 39735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, U.J.; Kim, H.J.; Lee, J.S.; Lee, M.K.; Kim, H.O.; Park, E.J.; Kim, H.K.; Jeong, T.S.; Choi, M.S. Naringin Supplementation Lowers Plasma Lipids and Enhances Erythrocyte Antioxidant Enzyme Activities in Hypercholesterolemic Subjects. Clin. Nutr. 2003, 22, 561–568. [Google Scholar] [CrossRef]
- Rashmi, R.; Bojan Magesh, S.; Mohanram Ramkumar, K.; Suryanarayanan, S.; Venkata SubbaRao, M. Antioxidant Potential of Naringenin Helps to Protect Liver Tissue from Streptozotocin-Induced Damage. Rep. Biochem. Mol. Biol. 2018, 7, 76–84. [Google Scholar]
- Rahman, K. Studies on Free Radicals, Antioxidants, and Co-Factors. Clin. Interv. Aging 2007, 2, 219–236. [Google Scholar]
- Sakihama, Y.; Mano, J.; Sano, S.; Asada, K.; Yamasaki, H. Reduction of Phenoxyl Radicals Mediated by Monodehydroascorbate Reductase. Biochem. Biophys. Res. Commun. 2000, 279, 949–954. [Google Scholar] [CrossRef]
- Pinho-Ribeiro, F.A.; Zarpelon, A.C.; Mizokami, S.S.; Borghi, S.M.; Bordignon, J.; Silva, R.L.; Cunha, T.M.; Alves-Filho, J.C.; Cunha, F.Q.; Casagrande, R.; et al. The Citrus Flavonone Naringenin Reduces Lipopolysaccharide-Induced Inflammatory Pain and Leukocyte Recruitment by Inhibiting NF-ΚB Activation. J. Nutr. Biochem. 2016, 33, 8–14. [Google Scholar] [CrossRef]
- Pinho-Ribeiro, F.A.; Verri, W.A.; Chiu, I.M. Nociceptor Sensory Neuron–Immune Interactions in Pain and Inflammation. Trends Immunol. 2017, 38, 5–19. [Google Scholar] [CrossRef] [Green Version]
- Straub, I.; Mohr, F.; Stab, J.; Konrad, M.; Philipp, S.E.; Oberwinkler, J.; Schaefer, M. Citrus Fruit and Fabacea Secondary Metabolites Potently and Selectively Block TRPM3. Br. J. Pharmacol. 2013, 168, 1835–1850. [Google Scholar] [CrossRef] [Green Version]
- Li, J.M.; Che, C.T.; Lau, C.B.S.; Leung, P.S.; Cheng, C.H.K. Inhibition of Intestinal and Renal Na+-Glucose Cotransporter by Naringenin. Int. J. Biochem. Cell Biol. 2006, 38, 985–995. [Google Scholar] [CrossRef] [PubMed]
- Andrew Haber, C.; Lam, T.K.T.; Yu, Z.; Gupta, N.; Goh, T.; Bogdanovic, E.; Giacca, A.; George Fantus, I.; Andrew, C.; Gupta, N.; et al. N-Acetylcysteine and Taurine Prevent Hyperglycemia-Induced Insulin Resistance in Vivo: Possible Role of Oxidative Stress. Am. J. Physiol. Endocrinol. Metab. 2003, 285, E744–E753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhattacharya, S.; Christensen, K.B.; Cathrine, L.; Olsen, B.; Christensen, L.P.; Grevsen, K.; Faergeman, N.J.; Kristiansen, K.; Young, J.F.; Oksbjerg, N. Bioactive Components from Flowers of Sambucus nigra L. Increase Glucose Uptake in Primary Porcine Myotube Cultures and Reduce Fat Accumulation in Caenorhabditis elegans. J. Agric. Food Chem. 2013, 61, 11033–11040. [Google Scholar] [CrossRef] [PubMed]
- Claussnitzer, M.; Skurk, T.; Hauner, H.; Daniel, H.; Rist, M.J. Effect of Flavonoids on Basal and Insulin-Stimulated 2-Deoxyglucose Uptake in Adipocytes. Mol. Nutr. Food Res. 2011, 55, S26–S34. [Google Scholar] [CrossRef]
- Yoshida, H.; Watanabe, W.; Oomagari, H.; Tsuruta, E.; Shida, M.; Kurokawa, M. Citrus Flavonoid Naringenin Inhibits TLR2 Expression in Adipocytes. J. Nutr. Biochem. 2013, 24, 1276–1284. [Google Scholar] [CrossRef]
- Bhattacharya, S.; Oksbjerg, N.; Young, J.F.; Jeppesen, P.B. Caffeic Acid, Naringenin and Quercetin Enhance Glucose-Stimulated Insulin Secretion and Glucose Sensitivity in INS-1E Cells. Diabetes, Obes. Metab. 2014, 16, 602–612. [Google Scholar] [CrossRef]
- Ortiz-Andrade, R.R.; Sánchez-Salgado, J.C.; Navarrete-Vázquez, G.; Webster, S.P.; Binnie, M.; García-Jiménez, S.; León-Rivera, I.; Cigarroa-Vázquez, P.; Villalobos-Molina, R.; Estrada-Soto, S. Antidiabetic and Toxicological Evaluations of Naringenin in Normoglycaemic and NIDDM Rat Models and Its Implications on Extra-Pancreatic Glucose Regulation. Diabetes, Obes. Metab. 2008, 10, 1097–1104. [Google Scholar] [CrossRef]
- Sharma, A.K.; Bharti, S.; Ojha, S.; Bhatia, J.; Kumar, N.; Ray, R.; Kumari, S.; Arya, D.S. Up-Regulation of PPARγ, Heat Shock Protein-27 and-72 by Naringin Attenuates Insulin Resistance, β-Cell Dysfunction, Hepatic Steatosis and Kidney Damage in a Rat Model of Type 2 Diabetes. Br. J. Nutr. 2011, 106, 1713–1723. [Google Scholar] [CrossRef] [Green Version]
- Dodds, D.R. Antibiotic Resistance: A Current Epilogue. Biochem. Pharmacol. 2017, 134, 139–146. [Google Scholar] [CrossRef]
- Gross, P.A.; Patel, B. Reducing Antibiotic Overuse: A Call for a National Performance Measure for Not Treating Asymptomatic Bacteriuria. Clin. Infect. Dis. 2007, 45, 1335–1337. [Google Scholar] [CrossRef] [PubMed]
- Davies, J.; Davies, D. Origins and Evolution of Antibiotic Resistance. Microbiol. Mol. Biol. Rev. 2010, 74, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Rebello, C.J.; Beyl, R.A.; Lertora, J.J.L.; Greenway, F.L.; Ravussin, E.; Ribnicky, D.M.; Poulev, A.; Kennedy, B.J.; Castro, H.F.; Campagna, S.R.; et al. Safety and Pharmacokinetics of Naringenin: A Randomized, Controlled, Single-Ascending-Dose Clinical Trial. Diabetes, Obes. Metab. 2020, 22, 91–98. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.H.; Zeng, X.A.; Wang, M.S.; Brennan, C.S.; Gong, D. Modification of Membrane Properties and Fatty Acids Biosynthesis-Related Genes in Escherichia Coli and Staphylococcus aureus: Implications for the Antibacterial Mechanism of Naringenin. Biochim. Biophys. Acta-Biomembr. 2018, 1860, 481–490. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.H.; Wang, M.S.; Zeng, X.A.; Xu, X.M.; Brennan, C.S. Membrane and Genomic DNA Dual-Targeting of Citrus Flavonoid Naringenin against: Staphylococcus Aureus. Integr. Biol. (UK) 2017, 9, 820–829. [Google Scholar] [CrossRef] [PubMed]
- Duda-Madej, A.; Kozłowska, J.; Krzyzek, P.; Anioł, M.; Seniuk, A.; Jermakow, K.; Dworniczek, E. Antimicrobial O -Alkyl Derivatives of Naringenin and Their Oximes against Multidrug-Resistant Bacteria. Molecules 2020, 25, 174–183. [Google Scholar] [CrossRef] [PubMed]
- Mohammed, N.H.; Mostafa, M.I.; Al-Taher, A.Y. Augmentation Effects of Novel Naringenin Analogues and Ciprofloxacin as Inhibitors for Nora Efflux Pump (EPIs) and Pyruvate Kinase (PK) against MRSA. J. Anim. Vet. Adv. 2015, 14, 386–392. [Google Scholar]
- Denny, B.J.; West, P.W.J.; Mathew, T.C. Antagonistic Interactions between the Flavonoids Hesperetin and Naringenin and β-Lactam Antibiotics against Staphylococcus Aureus. Br. J. Biomed. Sci. 2008, 65, 145–147. [Google Scholar] [CrossRef]
- Song, H.-S.; Kant Bhatia, S.; Gurav, R.; Choi, T.-R.; Joong Kim, H.; Park, L.; Han, Y.-H.; Young Park, J.; Mi Lee, S.; Lee Park, S.; et al. Naringenin as an Antibacterial Reagent Controlling of Biofilm Formation 1 and Fatty Acid Metabolism in MRSA. bioRxiv 2020. [Google Scholar] [CrossRef] [Green Version]
- Yue, J.; Yang, H.; Liu, S.; Song, F.; Guo, J.; Huang, C. Influence of Naringenin on the Biofilm Formation of Streptococcus Mutans. J. Dent. 2018, 76, 24–31. [Google Scholar] [CrossRef]
- Zhang, Y.; Wang, J.F.; Dong, J.; Wei, J.Y.; Wang, Y.N.; Dai, X.H.; Wang, X.; Luo, M.J.; Tan, W.; Deng, X.M.; et al. Inhibition of α-Toxin Production by Subinhibitory Concentrations of Naringenin Controls Staphylococcus Aureus Pneumonia. Fitoterapia 2013, 86, 92–99. [Google Scholar] [CrossRef] [PubMed]
- Yao, W.; Zhang, X.; Xu, F.; Cao, C.; Liu, T.; Xue, Y. The Therapeutic Effects of Naringenin on Bronchial Pneumonia in Children. Pharmacol. Res. Perspect. 2021, 9, e00825. [Google Scholar] [CrossRef] [PubMed]
- Yu, M.; You, D.; Zhuang, J.; Lin, S.; Dong, L.; Weng, S.; Zhang, B.; Cheng, K.; Weng, W.; Wang, H. Controlled Release of Naringin in Metal-Organic Framework-Loaded Mineralized Collagen Coating to Simultaneously Enhance Osseointegration and Antibacterial Activity. ACS Appl. Mater. Interfaces 2017, 9, 19698–19705. [Google Scholar] [CrossRef] [PubMed]
- Rao, K.; Imran, M.; Jabri, T.; Ali, I.; Perveen, S.; Shafiullah; Ahmed, S.; Shah, M.R. Gum Tragacanth Stabilized Green Gold Nanoparticles as Cargos for Naringin Loading: A Morphological Investigation through AFM. Carbohydr. Polym. 2017, 174, 243–252. [Google Scholar] [CrossRef] [PubMed]
- Virolle, C.; Goldlust, K.; Djermoun, S.; Bigot, S.; Lesterlin, C. Plasmid Transfer by Conjugation in Gram-Negative Bacteria: From the Cellular to the Community Level. Genes 2020, 11, 1239. [Google Scholar] [CrossRef]
- Ikigai, H.; Nakae, T.; Hara, Y.; Shimamura, T. Bactericidal Catechins Damage the Lipid Bilayer. Biochim. Biophys. Acta 1993, 1147, 132–136. [Google Scholar] [CrossRef]
- Céliz, G.; Daz, M.; Audisio, M.C. Antibacterial Activity of Naringin Derivatives against Pathogenic Strains. J. Appl. Microbiol. 2011, 111, 731–738. [Google Scholar] [CrossRef] [Green Version]
- Han, S.-S.; Lee, C.-K.; Kim, Y.-S. Antimicrobial Effects of Naringenin Alone and in Combination with Related Flavonoids. Yakhak Hoeji 1992, 36, 407–411. [Google Scholar]
- Murti, Y. Biological Evaluation of Synthesized Naringenin Derivatives as Antimicrobial Agents. Anti-Infective Agents 2020, 19, 192–199. [Google Scholar] [CrossRef]
- Vandeputte, O.M.; Kiendrebeogo, M.; Rasamiravaka, T.; Stévigny, C.; Duez, P.; Rajaonson, S.; Diallo, B.; Mol, A.; Baucher, M.; el Jaziri, M. The Flavanone Naringenin Reduces the Production of Quorum Sensing-Controlled Virulence Factors in Pseudomonas Aeruginosa PAO1. Microbiology 2011, 157, 2120–2132. [Google Scholar] [CrossRef] [Green Version]
- Bae, E.A.; Han, M.J.; Kim, D.H. In Vitro Anti-Helicobacter Pylori Activity of Some Flavonoids and Their Metabolites. Planta Med. 1999, 65, 442–443. [Google Scholar] [CrossRef] [PubMed]
- Vikram, A.; Jesudhasan, P.R.; Jayaprakasha, G.K.; Pillai, S.D.; Jayaraman, A.; Patil, B.S. Citrus Flavonoid Represses Salmonella Pathogenicity Island 1 and Motility in S. Typhimurium LT2. Int. J. Food Microbiol. 2011, 145, 28–36. [Google Scholar] [CrossRef] [PubMed]
- Vikram, A.; Jayaprakasha, G.K.; Jesudhasan, P.R.; Pillai, S.D.; Patil, B.S. Suppression of Bacterial Cell-Cell Signalling, Biofilm Formation and Type III Secretion System by Citrus Flavonoids. J. Appl. Microbiol. 2010, 109, 515–527. [Google Scholar] [CrossRef]
- Marin, A.M.; Souza, E.M.; Pedrosa, F.O.; Souza, L.M.; Sassaki, G.L.; Baura, V.A.; Yates, M.G.; Wassem, R.; Monteiro, R.A. Naringenin Degradation by the Endophytic Diazotroph Herbaspirillum Seropedicae SmR1. Microbiology 2013, 159, 167–175. [Google Scholar] [CrossRef] [PubMed]
- Ng, T.B.; Ling, J.M.L.; Wang, Z.T.; Cai, J.N.; Xu, G.J. Examination of Coumarins, Flavonoids and Polysaccharopeptide for Antibacterial Activity. Gen. Pharmacol. 1996, 27, 1237–1240. [Google Scholar] [CrossRef]
- Rauha, J.P.; Remes, S.; Heinonen, M.; Hopia, A.; Kähkönen, M.; Kujala, T.; Pihlaja, K.; Vuorela, H.; Vuorela, P. Antimicrobial Effects of Finnish Plant Extracts Containing Flavonoids and Other Phenolic Compounds. Int. J. Food Microbiol. 2000, 56, 3–12. [Google Scholar] [CrossRef]
- Eumkeb, G.; Chukrathok, S. Synergistic Activity and Mechanism of Action of Ceftazidime and Apigenin Combination against Ceftazidime-Resistant Enterobacter Cloacae. Phytomedicine 2013, 20, 262–269. [Google Scholar] [CrossRef]
- Negm, W.A.; El-aasr, M.; Kamer, A.A.; Elekhnawy, E. Investigation of the Antibacterial Activity and Efflux Pump Inhibitory Effect of Cycas Thouarsii r.Br. Extract against Klebsiella Pneumoniae Clinical Isolates. Pharmaceuticals 2021, 14, 756. [Google Scholar] [CrossRef]
- Tran Trung, H.; Truong Thi Huynh, H.; Nguyen Thi Thuy, L.; Nguyen Van Minh, H.; Thi Nguyen, M.N.; Luong Thi, M.N. Growth-Inhibiting, Bactericidal, Antibiofilm, and Urease Inhibitory Activities of Hibiscus Rosa sinensis L. Flower Constituents toward Antibiotic Sensitive- And Resistant-Strains of Helicobacter Pylori. ACS Omega 2020, 5, 20080–20089. [Google Scholar] [CrossRef]
- Duda-Chodak. A Polyphenols and the Gut Microbiota. J. Physiol. Pharmacol. 2012, 63, 497–503. [Google Scholar]
- Shin, J.E.; Kim, J.M.; Bae, E.A.; Hyun, Y.J.; Kim, D.H. In Vitro Inhibitory Effect of Flavonoids on Growth, Infection and Vacuolation of Helicobacter Pylori. Planta Med. 2005, 71, 197–201. [Google Scholar] [CrossRef] [PubMed]
- Seong Sun Han, I.J.Y. Studies on Antimicrobial Activities and Safety of Natural Naringin in Korea. Korean J. Mycol. 1988, 16, 33–44. [Google Scholar]
- de Andrade, C.A.; de S. Carvalho, J.L.; Cunico, M.M.; Lordello, A.L.L.; Higaskino, C.E.K.; Almeida, S.C.D.C.; Dias, J.D.F.G.; Kerber, V.A.; Miguel, M.D.; Miguel, O.G. Antioxidant and Antibacterial Activity of Extracts, Fractions and Isolated Substances from the Flowers of Acacia podalyriifolia A. Cunn. Ex G. Don. Brazilian J. Pharm. Sci. 2010, 46, 715–721. [Google Scholar] [CrossRef] [Green Version]
- Mundlia, J.; Ahuja, M.; Kumar, P.; Pillay, V. Improved Antioxidant, Antimicrobial and Anticancer Activity of Naringenin on Conjugation with Pectin. 3 Biotech 2019, 9, 312. [Google Scholar] [CrossRef] [PubMed]
- Mandalari, G.; Bennett, R.N.; Bisignano, G.; Trombetta, D.; Saija, A.; Faulds, C.B.; Gasson, M.J.; Narbad, A. Antimicrobial Activity of Flavonoids Extracted from Bergamot (Citrus bergamia Risso) Peel, a Byproduct of the Essential Oil Industry. J. Appl. Microbiol. 2007, 103, 2056–2064. [Google Scholar] [CrossRef]
- Mandalari, G.; Bisignano, C.; D’Arrigo, M.; Ginestra, G.; Arena, A.; Tomaino, A.; Wickham, M.S.J. Antimicrobial Potential of Polyphenols Extracted from Almond Skins. Lett. Appl. Microbiol. 2010, 51, 83–89. [Google Scholar] [CrossRef]
- Parkar, S.G.; Stevenson, D.E.; Skinner, M.A. The Potential Influence of Fruit Polyphenols on Colonic Microflora and Human Gut Health. Int. J. Food Microbiol. 2008, 124, 295–298. [Google Scholar] [CrossRef]
- Rastogi, N.; Domadia, P.; Shetty, S.; Dasgupta, D. Screening of Natural Phenolic Compounds for Potential to Inhibit Bacterial Cell Division Protein FtsZ. Indian J. Exp. Biol. 2008, 46, 783–787. [Google Scholar]
- Adamczak, A.; Ożarowski, M.; Karpiński, T.M. Antibacterial Activity of Some Flavonoids and Organic Acids Widely Distributed in Plants. J. Clin. Med. 2020, 9, 109. [Google Scholar] [CrossRef] [Green Version]
- Pereira, R.M.S.; Andrades, N.E.D.; Paulino, N.; Sawaya, A.C.H.F.; Eberlin, M.N.; Marcucci, M.C.; Favero, G.M.; Novak, E.M.; Bydlowski, S.P. Synthesis and Characterization of a Metal Complex Containing Naringin and Cu, and Its Antioxidant, Antimicrobial, Antiinflammatory and Tumor Cell Cytotoxicity. Molecules 2007, 12, 1352–1366. [Google Scholar] [CrossRef] [Green Version]
- Soberón, J.R.; Sgariglia, M.A.; Carabajal Torrez, J.A.; Aguilar, F.A.; Pero, E.J.I.; Sampietro, D.A.; Fernández de Luco, J.; Labadie, G.R. Antifungal Activity and Toxicity Studies of Flavanones Isolated from Tessaria Dodoneifolia Aerial Parts. Heliyon 2020, 6, e05174. [Google Scholar] [CrossRef] [PubMed]
- Salazar-Aranda, R.; Granados-Guzmán, G.; Pérez-Meseguer, J.; González, G.M.; De Torres, N.W. Activity of Polyphenolic Compounds against Candida Glabrata. Molecules 2015, 20, 17903. [Google Scholar] [CrossRef] [PubMed]
- Dean, R.; Van Kan, J.A.L.; Pretorius, Z.A.; Hammond-Kosack, K.E.; Di Pietro, A.; Spanu, P.D.; Rudd, J.J.; Dickman, M.; Kahmann, R.; Ellis, J.; et al. The Top 10 Fungal Pathogens in Molecular Plant Pathology. Mol. Plant Pathol. 2012, 13, 414–430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Padmavati, M.; Sakthivel, N.; Thara, K.V.; Reddy, A.R. Differential Sensitivity of Rice Pathogens to Growth Inhibition by Flavonoids. Phytochemistry 1997, 46, 499–502. [Google Scholar] [CrossRef]
- Katsumata, S.; Hamana, K.; Horie, K.; Toshima, H.; Hasegawa, M. Identification of Sternbin and Naringenin as Detoxified Metabolites from the Rice Flavanone Phytoalexin Sakuranetin by Pyricularia Oryzae. Chem. Biodivers. 2017, 14, e1600240. [Google Scholar] [CrossRef]
- Kim, H.; Lee, D.G. Naringin-Generated ROS Promotes Mitochondria-Mediated Apoptosis in Candida Albicans. IUBMB Life 2021, 73, 953–967. [Google Scholar] [CrossRef]
- Orhan, D.D.; Özçelik, B.; Özgen, S.; Ergun, F. Antibacterial, Antifungal, and Antiviral Activities of Some Flavonoids. Microbiol. Res. 2010, 165, 496–504. [Google Scholar] [CrossRef]
- Son, N.T.; Harada, K.; Cuong, N.M.; Fukuyama, Y. Two New Carboxyethylflavanones from the Heartwood of Dalbergia Tonkinensis and Their Antimicrobial Activities. Nat. Prod. Commun. 2017, 12, 1721–1723. [Google Scholar] [CrossRef] [Green Version]
- Górniak, I.; Bartoszewski, R.; Króliczewski, J. Comprehensive Review of Antimicrobial Activities of Plant Flavonoids. Phytochem. Rev. 2019, 18, 241–272. [Google Scholar] [CrossRef] [Green Version]
- Paczkowski, J.E.; Mukherjee, S.; McCready, A.R.; Cong, J.P.; Aquino, C.J.; Kim, H.; Henke, B.R.; Smith, C.D.; Bassler, B.L. Flavonoids Suppress Pseudomonas Aeruginosa Virulence through Allosteric Inhibition of Quorum-Sensing Receptors. J. Biol. Chem. 2017, 292, 4064–4076. [Google Scholar] [CrossRef] [Green Version]
- Pandey, A.K. Shashank Kumar Perspective on Plant Products as Antimicrobials Agents: A Review. Pharmacologia 2013, 4, 469–480. [Google Scholar] [CrossRef] [Green Version]
- Ammar, N.M.; Hassan, H.A.; Abdallah, H.M.I.; Afifi, S.M.; Elgamal, A.M.; Farrag, A.R.H.; El-Gendy, A.E.N.G.; Farag, M.A.; Elshamy, A.I. Protective Effects of Naringenin from Citrus Sinensis (Var. Valencia) Peels against CCl4-Induced Hepatic and Renal Injuries in Rats Assessed by Metabolomics, Histological and Biochemical Analyses. Nutrients 2022, 14, 841. [Google Scholar] [CrossRef] [PubMed]
- Tsuchiya, H.; Iinuma, M. Reduction of Membrane Fluidity by Antibacterial Sophoraflavanone G Isolated from Sophora Exigua. Phytomedicine 2000, 7, 161–165. [Google Scholar] [CrossRef]
- Jeong, K.W.; Lee, J.Y.; Kang, D., II; Lee, J.U.; Shin, S.Y.; Kim, Y. Screening of Flavonoids as Candidate Antibiotics against Enterococcus Faecalis. J. Nat. Prod. 2009, 72, 719–724. [Google Scholar] [CrossRef]
- Zhang, L.; Kong, Y.; Wu, D.; Zhang, H.; Wu, J.; Chen, J.; Ding, J.; Hu, L.; Jiang, H.; Shen, X. Three Flavonoids Targeting the β-Hydroxyacyl-Acyl Carrier Protein Dehydratase from Helicobacter Pylori: Crystal Structure Characterization with Enzymatic Inhibition Assay. Protein Sci. 2008, 17, 1971–1978. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, Z.P.; Wang, X.D.; Wang, P.F.; Zhou, Y.; Zhang, J.W.; Zhang, L.; Zhou, J.; Zhou, S.S.; Hui, O.; Lin, X.Y.; et al. Design, Synthesis, and Evaluation of Novel Fluoroquinolone-Flavonoid Hybrids as Potent Antibiotics against Drug-Resistant Microorganisms. Eur. J. Med. Chem. 2014, 80, 92–100. [Google Scholar] [CrossRef]
- Oh, E.; Jeon, B. Synergistic Anti-Campylobacter Jejuni Activity of Fluoroquinolone and Macrolide Antibiotics with Phenolic Compounds. Front. Microbiol. 2015, 6, 1129. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Duda-Madej, A.; Stecko, J.; Sobieraj, J.; Szymańska, N.; Kozłowska, J. Naringenin and Its Derivatives—Health-Promoting Phytobiotic against Resistant Bacteria and Fungi in Humans. Antibiotics 2022, 11, 1628. https://doi.org/10.3390/antibiotics11111628
Duda-Madej A, Stecko J, Sobieraj J, Szymańska N, Kozłowska J. Naringenin and Its Derivatives—Health-Promoting Phytobiotic against Resistant Bacteria and Fungi in Humans. Antibiotics. 2022; 11(11):1628. https://doi.org/10.3390/antibiotics11111628
Chicago/Turabian StyleDuda-Madej, Anna, Jakub Stecko, Jakub Sobieraj, Natalia Szymańska, and Joanna Kozłowska. 2022. "Naringenin and Its Derivatives—Health-Promoting Phytobiotic against Resistant Bacteria and Fungi in Humans" Antibiotics 11, no. 11: 1628. https://doi.org/10.3390/antibiotics11111628