Antibody Phage Display Technology for Sensor-Based Virus Detection: Current Status and Future Prospects


Abstract
:1. Introduction
- -
- Making bacteriophages or choosing them from libraries of available phages;
- -
- Biopanning (affinity-based enrichment of phage libraries);
- -
- Phage-displayed antibodies go through a number of affinity selection procedures before being utilized as specific biosensor receptors [24].
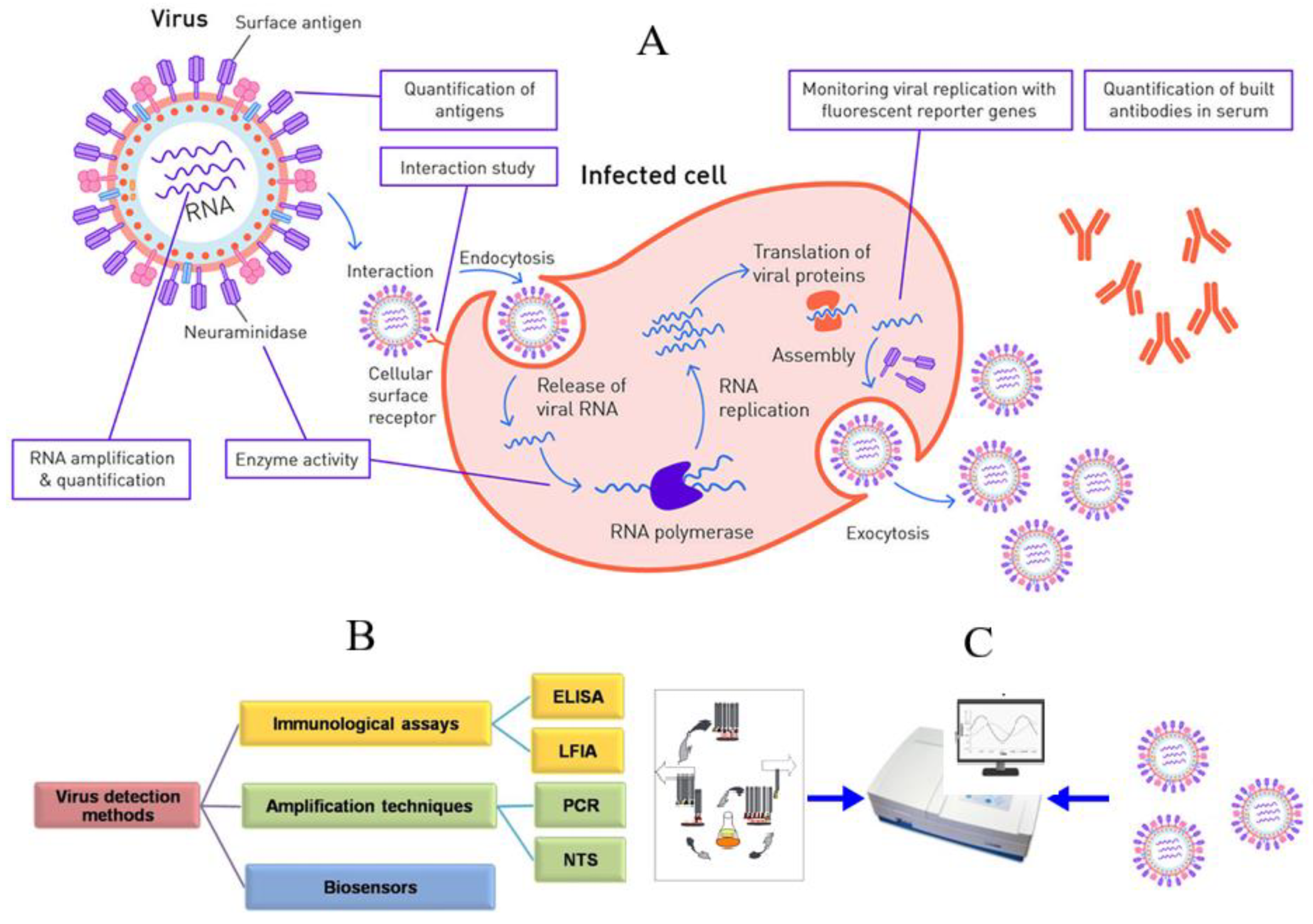
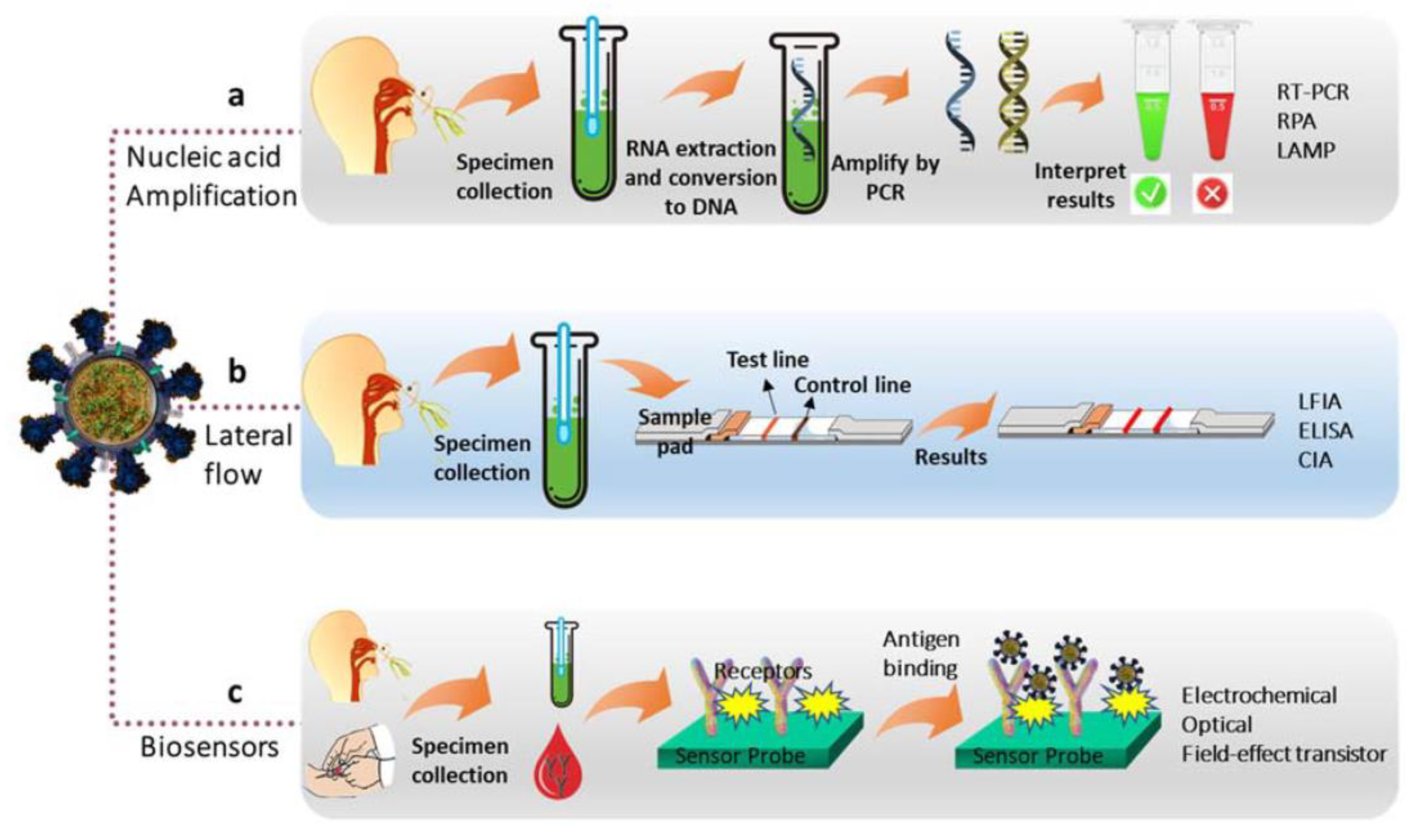
2. Methods of Virus Detection
- -
- Enzymatic biosensors;
- -
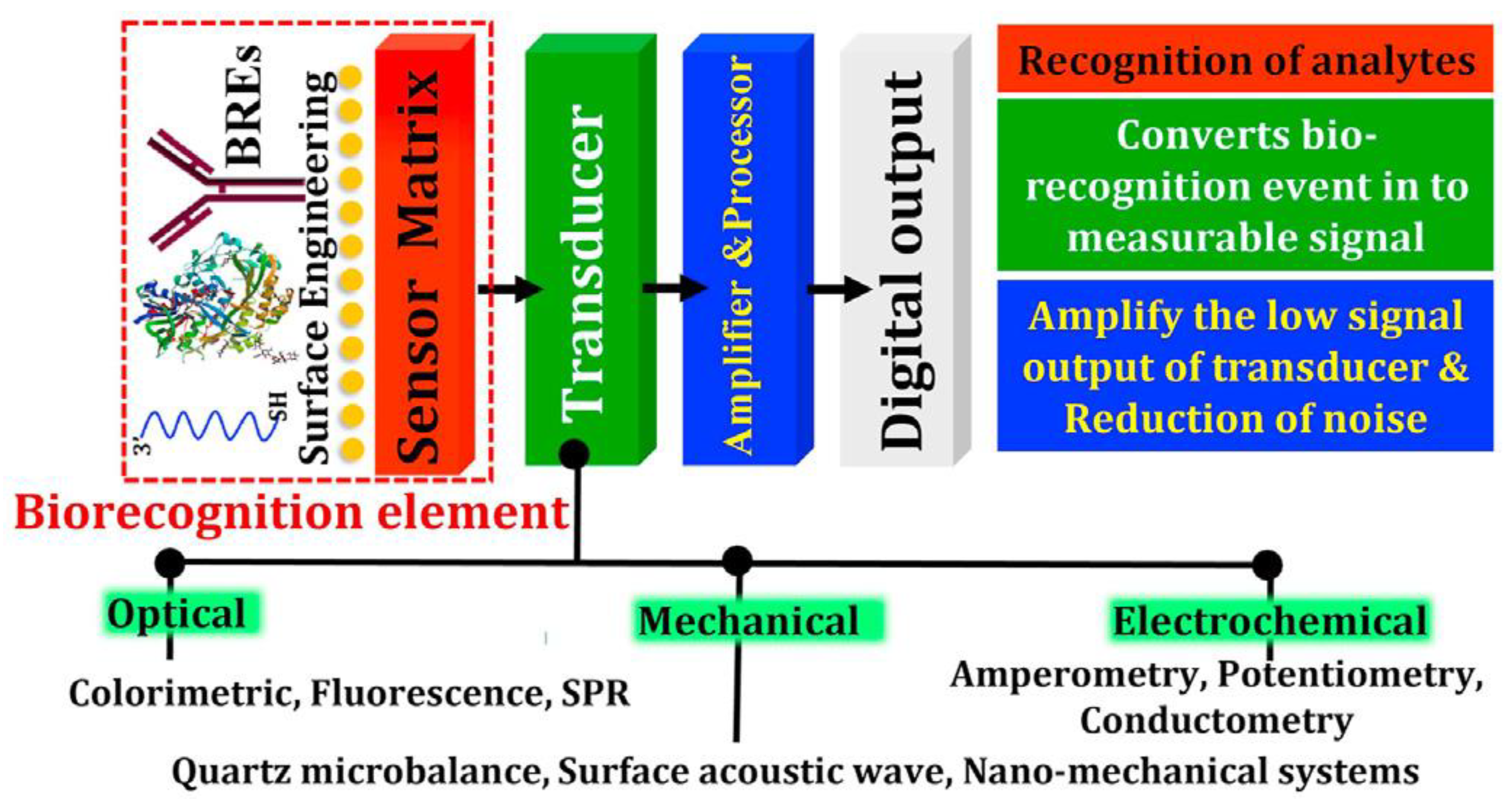
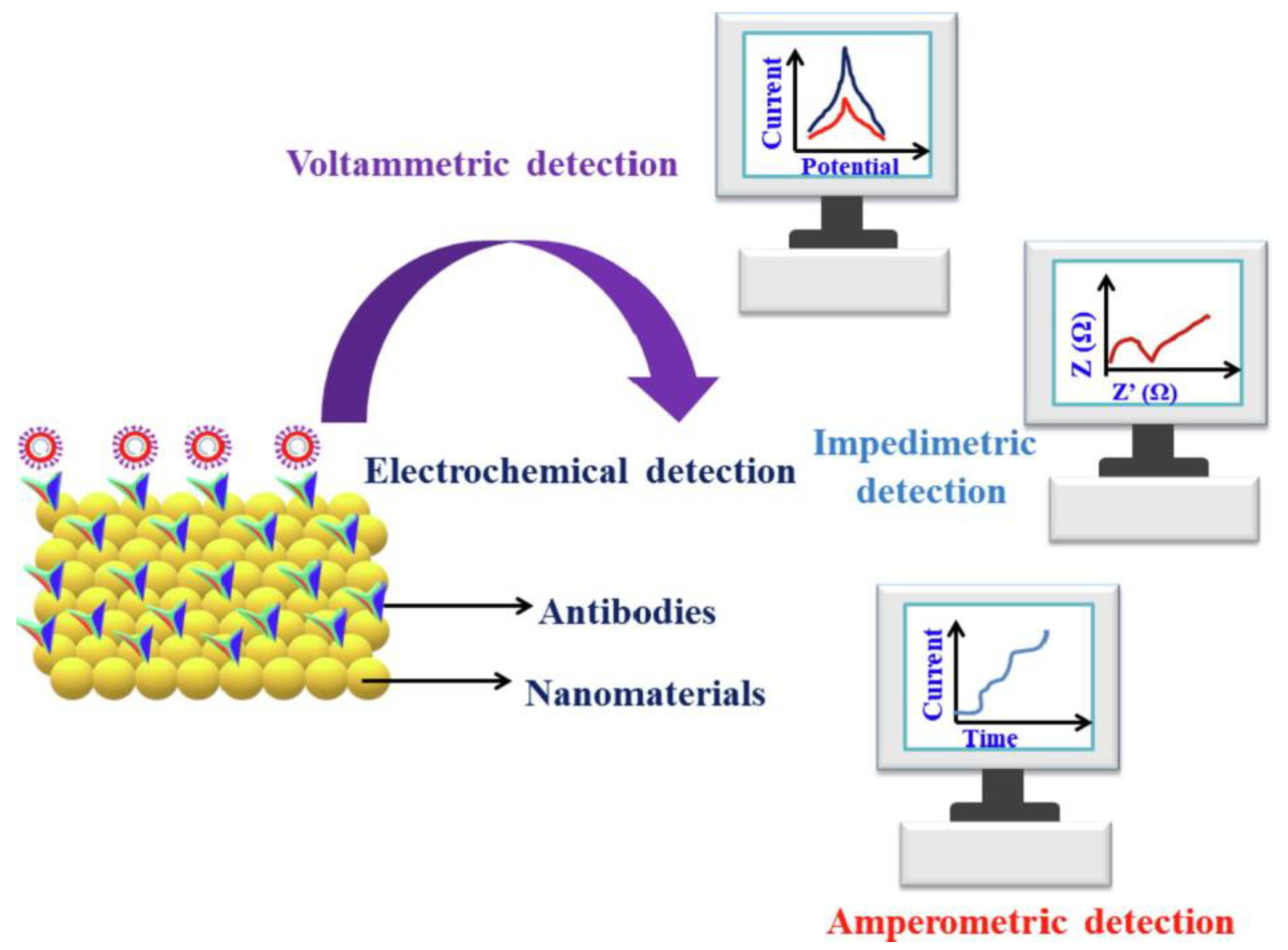
- Affinity biosensors;
- -
- Cell/tissue biosensors.
3. Phage Display Technology
- -
- Antibodies can be selected in vitro, and animal immunization can be omitted;
- -
- No need to use laboratory animals or maintain long-term cultures of eukaryotic cells;
- -
- The preparation of individual clone producers of miniantibodies takes 10–14 days, against the several months that hybridoma technology takes;
- -
- Antibodies are relatively easy to prepare, and their cost is low;
- -
- It is possible to make hybrid molecules with marker proteins (e.g., tag peptide) in a short time;
- -
- It is possible to prepare antibodies to autoantigens, weakly immunogenic compounds, and toxins;
- -
- The method can be used in immunotherapy.
- -
- The immobilized antigen is incubated with library clones;
- -
- Particles of the phage that have not yet attached to the antigen are washed away;
- -
- The bound phage particles are eluted;
- -
- The selected clones are infected with bacterial cells (E. coli);
- -
- The affinity clones are amplified and isolated.
- -
- The genes of scFv, Fab, or other fragments are cloned from immune or undamaged humans, mice, rabbits, chickens, pigs, dogs, monkeys, sheep, or cows’ B lymphocyte mRNA;
- -
- These genes, along with the gene coding for the capsid protein (typically p3), are introduced into the phagemid in a single translation frame;
- -
- Phagemid genes are expressed, and virions are put together in infected E. coli cells by using the generated phagemid repertoire.
- -
- Owing to the Fc region of the intact antibody, the tiny size of the antibody fragments is typically accompanied by reduced nonspecific binding;
- -
- The biosensor can immobilize phage antibodies more densely;
- -
- Phage antibodies, as opposed to full-length antibodies, are produced by E. coli cells, which significantly lowers production costs because no specialized equipment is needed for the culture of hybridoma cells [75].
4. Phage Antibodies for Virus Detection/Identification
5. Discussion and Prospects
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- van Rijn, P.; Schirhagl, R. Viruses, artificial viruses and virus-based structures for biomedical applications. Adv. Healthc. Mater. 2016, 5, 1386–1400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zaidi, S.A. An overview of bio-inspired intelligent imprinted polymers for virus determination. Biosensors 2021, 11, 89. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Nolte, R.J.; Cornelissen, J.J. Virus-based nanocarriers for drug delivery. Adv. Drug Deliv. Rev. 2012, 64, 811–825. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Yang, H.; Shin, H.J. Viruses as self-assembled nanocontainers for encapsulation of functional cargoes. Korean J. Chem. Eng. 2013, 30, 1359–1367. [Google Scholar] [CrossRef]
- Manchester, M.; Singh, P. Virus-based nanoparticles (VNPs): Platform technologies for diagnostic imaging. Adv. Drug Deliv. Rev. 2006, 58, 1505–1522. [Google Scholar] [CrossRef]
- Steinmetz, N.F.; Mertens, M.E.; Taurog, R.E.; Johnson, J.E.; Commandeur, U.; Fischer, R.; Manchester, M. Potato virus X as a novel platform for potential biomedical applications. Nano Lett. 2010, 10, 305–312. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.H.; Kim, B.C.; Oh, B.K.; Choi, J.W. Rapid and sensitive determination of HIV-1 virus based on surface enhanced Raman spectroscopy. J. Biomed. Nanotechnol. 2015, 11, 2223–2230. [Google Scholar] [CrossRef]
- Rappuoli, R.; De Gregorio, E.; Del Giudice, G.; Phogat, S.; Pecetta, S.; Pizza, M.; Hanon, E. Vaccinology in the post-COVID-19 era. Proc. Natl. Acad. Sci. USA 2021, 118, e2020368118. [Google Scholar] [CrossRef]
- Daniel, M.C.; Tsvetkova, I.B.; Quinkert, Z.T.; Murali, A.; De, M.; Rotello, V.M.; Kao, C.C.; Dragnea, B. Role of surface charge density in nanoparticle-templated assembly of bromovirus protein cages. ACS Nano 2010, 4, 3853–3860. [Google Scholar] [CrossRef] [Green Version]
- Steinmetz, N.F.; Lin, T.; Lomonossoff, G.P.; Johnson, J.E. Structure-based engineering of an icosahedral virus for nanomedicine and nanotechnology. In Viruses and Nanotechnology. Current Topics in Microbiology and Immunology; Manchester, M., Steinmetz, N.F., Eds.; Springer: Berlin/Heidelberg, Germany, 2009; Volume 327, pp. 23–58. [Google Scholar] [CrossRef]
- Young, M.; Willits, D.; Uchida, M.; Douglas, T. Plant viruses as biotemplates for materials and their use in nanotechnology. Annu. Rev. Phytopathol. 2008, 46, 361–384. [Google Scholar] [CrossRef]
- Nam, K.T.; Kim, D.W.; Yoo, P.J.; Chiang, C.Y.; Meethong, N.; Hammond, P.T.; Chiang, Y.M.; Belcher, A.M. Virus-enabled synthesis and assembly of nanowires for lithium ion battery electrodes. Science 2006, 312, 885–888. [Google Scholar] [CrossRef] [Green Version]
- Tseng, R.J.; Tsai, C.; Ma, L.; Ouyang, J.; Ozkan, C.S.; Yang, Y. Digital memory device based on tobacco mosaic virus conjugated with nanoparticles. Nat. Nanotechnol. 2006, 1, 72–77. [Google Scholar] [CrossRef]
- Steinmetz, N.F.; Bize, A.; Findlay, K.C.; Lomonossoff, G.P.; Manchester, M.; Evans, D.J.; Prangishvili, D. Site-specific and spatially controlled addressability of a new viral nanobuilding block: Sulfolobus islandicus rod-shaped virus 2. Adv. Funct. Mater. 2008, 18, 3478–3486. [Google Scholar] [CrossRef]
- Rong, J.; Niu, Z.; Andrew Lee, L.; Wang, Q. Self-assembly of viral particles. Curr. Opin. Colloid Interface Sci. 2011, 16, 441–450. [Google Scholar] [CrossRef]
- Willats, W.G. Phage display: Practicalities and prospects. Plant Mol. Biol. 2002, 50, 837–854. [Google Scholar] [CrossRef]
- Wang, L.F.; Yu, M. Epitope identification and discovery using phage display libraries: Applications in vaccine development and diagnostics. Curr. Drug Targets 2004, 5, 1–15. [Google Scholar] [CrossRef]
- Kuzmicheva, G.A.; Belyavskaya, V.A. Peptide phage display in biotechnology and biomedicine. Biomed. Khim. 2016, 62, 481–495. [Google Scholar] [CrossRef] [Green Version]
- Smith, G.P. Filamentous fusion phage: Novel expression vectors that display cloned antigens on the virion surface. Science 1985, 228, 1315–1317. [Google Scholar] [CrossRef]
- Williams, D.D.; Benedek, O.; Turnbough, C.L., Jr. Species-specific peptide ligands for the detection of Bacillus anthracis spores. Appl. Environ. Microbiol. 2003, 69, 6288–6293. [Google Scholar] [CrossRef] [Green Version]
- Paoli, G.C.; Chen, C.-Y.; Brewster, J.D. Single-chain Fv antibody with specificity for Listeria monocytogenes. J. Immunol. Methods 2004, 289, 147–155. [Google Scholar] [CrossRef]
- Nanduri, V.; Bhunia, A.K.; Tu, S.-I.; Paoli, G.C.; Brewster, J.D. SPR biosensor for the detection of L. monocytogenes using phage-displayed antibody. Biosens. Bioelectron. 2007, 23, 248–252. [Google Scholar] [CrossRef] [PubMed]
- McCafferty, J.; Griffiths, A.D.; Winter, G.; Chiswell, D.J. Phage antibodies: Filamentous phage displaying antibody variable domains. Nature 1990, 348, 552–554. [Google Scholar] [CrossRef] [PubMed]
- Wang, P.; Yu, G.; Wei, J.; Liao, X.; Zhang, Y.; Ren, Y.; Zhang, C.; Wang, Y.; Zhang, D.; Wang, J.; et al. A single thiolated-phage displayed nanobody-based biosensor for label-free detection of foodborne pathogen. J. Hazard. Mater. 2023, 443, 130157. [Google Scholar] [CrossRef]
- Krejcova, L.; Michalek, P.; Rodrigo, M.M.; Heger, Z.; Krizkova, S.; Vaculovicova, M.; Hynek, D.; Adam, V.; Kizek, R. Nanoscale virus biosensors: State of the art. Nanobiosens. Dis. Diagn. 2015, 4, 47–66. [Google Scholar] [CrossRef] [Green Version]
- Song, M.; Yang, M.; Hao, J. Pathogenic virus detection by optical nanobiosensors. Cell Rep. Phys. Sci. 2021, 2, 100288. [Google Scholar] [CrossRef]
- Aghamirza Moghim Aliabadi, H.; Eivazzadeh-Keihan, R.; Beig Parikhani, A.; Fattahi Mehraban, S.; Maleki, A.; Fereshteh, S.; Bazaz, M.; Zolriasatein, A.; Bozorgnia, B.; Rahmati, S.; et al. COVID-19: A systematic review and update on prevention, diagnosis, and treatment. MedComm 2022, 3, e115. [Google Scholar] [CrossRef]
- WHO Coronavirus (COVID-19) Dashboard. 2023. Available online: https://covid19.who.int/ (accessed on 20 April 2023).
- Ammulu, M.A.; Nelluri, K.D.D.; Vemuri, P.K.; Mallampati, S.C.; Poda, S. COVID 19: A new insight into organ failure and complications caused by novel SARS-CoV-2 virus and discussion on the role of nanotechnology in detection, treatment and prevention of the disease. Curr. Trends Biotechnol. Pharm. 2022, 16, 577–602. [Google Scholar]
- World Health Statistics. 2022. Available online: https://www.who.int/news/item/20-05-2022-world-health-statistics-2022 (accessed on 20 April 2023).
- European Centre for Disease Prevention and Control. Communicable Disease Threats Report, 2–8 January 2023, Week 1. Available online: https://www.ecdc.europa.eu/en/publications-data/communicable-disease-threats-report-2-8-january-2023-week-1 (accessed on 20 April 2023).
- Internet Resource. Available online: https://www.bmglabtech.com/en/blog/virus-detection-methods/ (accessed on 7 April 2023).
- Hsiung, G.D. Diagnostic virology: From animals to automation. Yale J. Biol. Med. 1984, 57, 727–733. [Google Scholar]
- Leland, D.S.; Ginocchio, C.C. Role of cell culture for virus detection in the age of technology. Clin. Microbiol. Rev. 2007, 20, 49–78. [Google Scholar] [CrossRef] [Green Version]
- Coiras, M.T.; Aguilar, J.C.; García, M.L.; Casas, I.; Pérez-Breña, P. Simultaneous detection of fourteen respiratory viruses in clinical specimens by two multiplex reverse transcription nested-PCR assays. J. Med. Virol. 2004, 72, 484–495. [Google Scholar] [CrossRef]
- Syrmis, M.W.; Whiley, D.M.; Thomas, M.; Mackay, I.M.; Williamson, J.; Siebert, D.J.; Nissen, M.D.; Sloots, T.P. A sensitive, specific, and cost-effective multiplex reverse transcriptase-PCR assay for the detection of seven common respiratory viruses in respiratory samples. J. Mol. Diagn. 2004, 6, 125–131. [Google Scholar] [CrossRef] [Green Version]
- Templeton, K.E.; Scheltinga, S.A.; Beersma, M.F.C.; Kroes, A.C.M.; Claas, E.C.J. Rapid and sensitive method using multiplex real-time PCR for diagnosis of infections by influenza a and influenza B viruses, respiratory syncytial virus, and parainfluenza viruses 1, 2, 3, and 4. J. Clin. Microbiol. 2004, 42, 1564–1569. [Google Scholar] [CrossRef] [Green Version]
- Samson, R.; Navale, G.R.; Dharne, M.S. Biosensors: Frontiers in rapid detection of COVID-19. 3 Biotech 2020, 10, 385. [Google Scholar] [CrossRef]
- Zhou, H.; Liu, D.; Ma, L.; Ma, T.; Xu, T.; Ren, L.; Li, L.; Xu, S. A SARS-CoV-2 reference standard quantified by multiple digital PCR platforms for quality assessment of molecular tests. Anal. Chem. 2021, 93, 715–721. [Google Scholar] [CrossRef]
- Mehra, B.; Bhattar, S.; Bhalla, P.; Rawat, D. Rapid tests versus ELISA for screening of HIV infection: Our experience from a voluntary counselling and testing facility of a tertiary care centre in North India. ISRN AIDS 2014, 2014, 296840. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention. Dengue. 2019. Available online: https://www.cdc.gov/dengue/healthcare-providers/diagnosis.html (accessed on 20 April 2023).
- Lin, B.; Blaney, K.M.; Malanoski, A.P.; Ligler, A.G.; Schnur, J.M.; Metzgar, D.; Russell, K.L.; Stenger, D.A. Using a resequencing microarray as a multiple respiratory pathogen detection assay. J. Clin. Microbiol. 2007, 45, 443–452. [Google Scholar] [CrossRef] [Green Version]
- Mehlmann, M.; Bonner, A.B.; Williams, J.V.; Dankbar, D.M.; Moore, C.L.; Kuchta, R.D.; Podsiad, A.B.; Tamerius, J.D.; Dawson, E.D.; Rowlen, K.L. Comparison of the MChip to viral culture, reverse transcription-PCR, and the QuickVue influenza A+B test for rapid diagnosis of influenza. J. Clin. Microbiol. 2007, 45, 1234–1237. [Google Scholar] [CrossRef] [Green Version]
- Huguenin, A.; Moutte, L.; Renois, F.; Leveque, N.; Talmud, D.; Abely, M.; Nguyen, Y.; Carrat, F.; Andreoletti, L. Broad respiratory virus detection in infants hospitalized for bronchiolitis by use of a multiplex RT-PCR DNA microarray system. J. Med. Virol. 2012, 84, 979–985. [Google Scholar] [CrossRef]
- Ince, B.; Sezgintürk, M.K. Lateral flow assays for viruses diagnosis: Up-to-date technology and future prospects. Trends Analyt. Chem. 2022, 157, 116725. [Google Scholar] [CrossRef]
- Rahman, M.M. Progress in electrochemical biosensing of SARS-CoV-2 virus for COVID-19 management. Chemosensors 2022, 10, 287. [Google Scholar] [CrossRef]
- Purohit, B.; Vernekar, P.R.; Shetti, N.P.; Chandra, P. Biosensor nanoengineering: Design, operation, and implementation for biomolecular analysis. Sens. Int. 2020, 1, 100040. [Google Scholar] [CrossRef]
- Ribeiro, B.V.; Cordeiro, T.A.R.; Oliveira e Freitas, G.R.; Ferreira, L.F.; Franco, D.L. Biosensors for the detection of respiratory viruses: A review. Talanta Open 2020, 2, 100007. [Google Scholar] [CrossRef] [PubMed]
- Monteil, S.; Casson, A.J.; Jones, S.T. Electronic and electrochemical viral detection for point-of-care use: A systematic review. PLoS ONE 2021, 16, e0258002. [Google Scholar] [CrossRef] [PubMed]
- Kumar, N.; Shetti, N.P.; Jagannath, S.; Aminabhavi, T.M. Electrochemical sensors for the detection of SARS-CoV-2 virus. Chem. Eng. J. 2022, 430, 132966. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Li, L.; Qiao, Z.; Lei, C.; Fu, Y.; Xie, Q.; Yao, S.; Li, Y.; Ying, Y. Electrochemical conversion of Fe3O4 magnetic nanoparticles to electroactive Prussian blue analogues for self-sacrificial label biosensing of avian influenza virus H5N1. Anal. Chem. 2017, 89, 12145–12151. [Google Scholar] [CrossRef]
- Mahari, S.; Roberts, A.; Shahdeo, D.; Gandhi, S. eCovSens-ultrasensitive novel in-house built printed circuit board based electrochemical device for rapid detection of nCovid-19 antigen, a spike protein domain I of SARS-CoV-2. bioRxiv 2020. [Google Scholar] [CrossRef] [Green Version]
- Białobrzeska, W.; Ficek, M.; Dec, B.; Osella, S.; Trzaskowski, B.; Jaramillo-Botero, A.; Pierpaoli, M.; Rycewicz, M.; Dashkevich, Y.; Łęga, T.; et al. Performance of electrochemical immunoassays for clinical diagnostics of SARS-CoV-2 based on selective nucleocapsid N protein detection: Boron-doped diamond, gold and glassy carbon evaluation. Biosens. Bioelectron. 2022, 209, 114222. [Google Scholar] [CrossRef]
- Lesniewski, A.; Los, M.; Jonsson-Niedziółka, M.; Krajewska, A.; Szot, K.; Los, J.M.; Niedziolka-Jonsson, J. Antibody modified gold nanoparticles for fast and selective, colorimetric T7 bacteriophage detection. Bioconjug. Chem. 2014, 25, 644–648. [Google Scholar] [CrossRef]
- Evtugyn, G.; Porfireva, A.; Stepanova, V.; Kutyreva, M.; Gataulina, A.; Ulakhovich, N.; Evtugyn, V.; Hianik, T. Impedimetric aptasensor for ochratoxin A determination based on Au nanoparticles stabilized with hyper-branched polymer. Sensors 2013, 13, 16129–16145. [Google Scholar] [CrossRef]
- Darwish, N.T.; Sekaran, S.D.; Alias, Y.; Khor, S.M. Immunofluorescence-based biosensor for the determination of dengue virus NS1 in clinical samples. J. Pharm. Biomed. Anal. 2018, 149, 591–602. [Google Scholar] [CrossRef]
- Guliy, O.I.; Kanevskiy, M.V.; Fomin, A.S.; Staroverov, S.A.; Bunin, V.D. Progress in the use of an electro-optical sensor for virus detection. Opt. Commun. 2020, 465, 125605. [Google Scholar] [CrossRef]
- Ballantine, D.S.; White, R.M.; Martin, S.J.; Ricco, A.J.; Zellers, E.T.; Frye, G.C.; Wohltjen, H. Acoustic Wave Sensors: Theory, Design, and Physico-Chemical Applications, 1st ed.; Academic Press: San Diego, CA, USA, 1997. [Google Scholar]
- König, B.; Grätzel, M. A novel immunosensor for herpes viruses. Anal. Chem. 1994, 66, 341–344. [Google Scholar] [CrossRef]
- Bisoffi, M.; Hjelle, B.; Brown, D.C.; Branch, D.W.; Edwards, T.L.; Brozik, S.M.; Bondu-Hawkins, V.S.; Larson, R.S. Detection of viral bioagents using a shear horizontal surface acoustic wave biosensor. Biosens. Bioelectron. 2008, 23, 1397–1403. [Google Scholar] [CrossRef]
- Tamarin, O.; Comeau, S.; Déjous, C.; Moynet, D.; Rebière, D.; Bezian, J.; Pistré, J. Real time device for biosensing: Design of a bacteriophage model using love acoustic waves. Biosens. Bioelectron. 2003, 18, 755–763. [Google Scholar] [CrossRef]
- Guliy, O.; Zaitsev, B.; Teplykh, A.; Balashov, S.; Fomin, A.; Staroverov, S.; Borodina, I. Acoustical slot mode sensor for the rapid coronaviruses detection. Sensors 2021, 21, 1822. [Google Scholar] [CrossRef]
- Tessaro, L.; Aquino, A.; Panzenhagen, P.; Joshi, N.; Conte-Junior, C.A. A systematic review of the advancement on colorimetric nanobiosensors for SARS-CoV-2 detection. J. Pharm. Biomed. Anal. 2023, 222, 115087. [Google Scholar] [CrossRef]
- Tadesse, L.F.; Safir, F.; Ho, C.-S.; Hasbach, X.; Khuri-Yakub, B.P.; Jeffrey, S.S.; Saleh, A.A.E.; Dionne, J. Toward rapid infectious disease diagnosis with advances in surface-enhanced Raman spectroscopy. J. Chem. Phys. 2020, 152, 240902. [Google Scholar] [CrossRef]
- Roth, K.D.R.; Wenzel, E.V.; Ruschig, M.; Steinke, S.; Langreder, N.; Heine, P.A.; Schneider, K.-T.; Ballmann, R.; Fühner, V.; Kuhn, P.; et al. Developing recombinant antibodies by phage display against infectious diseases and toxins for diagnostics and therapy. Front. Cell. Infect. Microbiol. 2021, 11, 697876. [Google Scholar] [CrossRef]
- Rülker, T.; Voß, L.; Thullier, P.; O’ Brien, L.M.; Pelat, T.; Perkins, S.D.; Langermann, C.; Schirrmann, T.; Dübel, S.; Marschall, H.-J.; et al. Isolation and characterisation of a human-like antibody fragment (scFv) that inactivates VEEV in vitro and in vivo. PLoS ONE 2012, 7, e37242. [Google Scholar] [CrossRef] [Green Version]
- Goncalvez, A.P.; Chien, C.-H.; Tubthong, K.; Gorshkova, I.; Roll, C.; Donau, O.; Schuck, P.; Yoksan, S.; Wang, S.-D.; Purcell, R.H.; et al. Humanized monoclonal antibodies derived from chimpanzee Fabs protect against Japanese encephalitis virus in vitro and in vivo. J. Virol. 2008, 82, 7009–7021. [Google Scholar] [CrossRef] [Green Version]
- Fatima, A.; Wang, H.; Kang, K.; Xia, L.; Wang, Y.; Ye, W.; Wang, J.; Wang, X. Development of VHH antibodies against dengue virus type 2 NS1 and comparison with monoclonal antibodies for use in immunological diagnosis. PLoS ONE 2014, 9, e95263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, S.; Moayeri, M.; Leppla, S.H. Anthrax lethal and edema toxins in anthrax pathogenesis. Trends Microbiol. 2014, 22, 317–325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Wyngaardt, W.; Malatji, T.; Mashau, C.; Fehrsen, J.; Jordaan, F.; Miltiadou, D.; du Plessis, D.H. A large semi-synthetic single-chain Fv phage display library based on chicken immunoglobulin genes. BMC Biotechnol. 2004, 4, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tikunova, N.; Dubrovskaya, V.; Morozova, V.; Yun, T.; Khlusevich, Y.; Bormotov, N.; Laman, A.; Brovko, F.; Shvalov, A.; Belanov, E. The neutralizing human recombinant antibodies to pathogenic Orthopoxviruses derived from a phage display immune library. Virus Res. 2012, 163, 141–150. [Google Scholar] [CrossRef] [PubMed]
- Köhler, G.; Milstein, C. Continuous cultures of fused cells secreting antibody of predefined specificity. Nature 1975, 256, 495–497. [Google Scholar] [CrossRef]
- Charlton, K.A.; Moyle, S.; Porter, A.J.; Harris, W.J. Analysis of the diversity of a sheep antibody repertoire as revealed from a bacteriophage display library. J. Immunol. 2000, 164, 6221–6229. [Google Scholar] [CrossRef] [Green Version]
- Bashir, S.; Paeshuyse, J. Construction of antibody phage libraries and their application in veterinary immunovirology. Antibodies 2020, 9, 21. [Google Scholar] [CrossRef]
- Peltomaa, R.; Benito-Peña, E.; Barderas, R.; Moreno-Bondi, M.C. Phage display in the quest for new selective recognition elements for biosensors. ACS Omega 2019, 4, 11569–11580. [Google Scholar] [CrossRef]
- Kuhn, P.; Fühner, V.; Unkauf, T.; Moreira, G.M.S.G.; Frenzel, A.; Miethe, S.; Hust, M. Recombinant antibodies for diagnostics and therapy against pathogens and toxins generated by phage display. Proteom. Clin. Appl. 2016, 10, 922–948. [Google Scholar] [CrossRef]
- Anand, T.; Virmani, N.; Bera, B.C.; Vaid, R.K.; Vashisth, M.; Bardajatya, P.; Kumar, A.; Tripathi, B.N. Phage display technique as a tool for diagnosis and antibody selection for coronaviruses. Curr. Microbiol. 2021, 78, 1124–1134. [Google Scholar] [CrossRef]
- Velappan, N.; Martinez, J.S.; Valero, R.; Chasteen, L.; Ponce, L.; Bondu-Hawkins, V.; Kelly, C.; Pavlik, P.; Hjelle, B.; Bradbury, A.R.M. Selection and characterization of scFv antibodies against the Sin Nombre hantavirus nucleocapsid protein. J. Immunol. Methods 2007, 321, 60–69. [Google Scholar] [CrossRef]
- Unkauf, T.; Miethe, S.; Fühner, V.; Schirrmann, T.; Frenzel, A.; Hust, M. Generation of recombinant antibodies against toxins and viruses by phage display for diagnostics and therapy. In Protein Targeting Compounds. Advances in Experimental Medicine and Biology; Böldicke, T., Ed.; Springer: Cham, Switzerland, 2016; Volume 917, pp. 55–76. [Google Scholar] [CrossRef]
- Avnir, Y.; Tallarico, A.S.; Zhu, Q.; Bennett, A.S.; Connelly, G.; Sheehan, J.; Sui, J.; Fahmy, A.; Huang, C.-Y.; Cadwell, G.; et al. Molecular signatures of hemagglutinin stem-directed heterosubtypic human neutralizing antibodies against influenza A viruses. PLoS Pathog. 2014, 10, e1004103. [Google Scholar] [CrossRef]
- Walter, J.D.; Hutter, C.A.J.; Zimmermann, I.; Wyss, M.; Egloff, P.; Sorgenfrei, M.; Hürlimann, L.M.; Gonda, I.; Meier, G.; Remm, S.; et al. Sybodies targeting the SARS-CoV-2 receptor-binding domain. bioRxiv 2020. [Google Scholar] [CrossRef] [Green Version]
- He, L.; Li, J.; Ren, S.; Sun, S.; Guo, Y.; Qiu, H.; Liao, Y.; Ji, K.; Fan, R.; Zhao, G.; et al. Construction and identification of nanobody phage display library targeting Middle East respiratory syndrome coronavirus. Xi Bao Yu Fen Zi Mian Yi Xue Za Zhi 2017, 33, 1662–1668. (In Chinese) [Google Scholar]
- Ubah, O.; Palliyil, S. Monoclonal antibodies and antibody like fragments derived from immunised phage display libraries. In Recombinant Antibodies for Infectious Diseases. Advances in Experimental Medicine and Biology; Lim, T., Ed.; Springer: Cham, Switzerland, 2017; Volume 1053, pp. 99–117. [Google Scholar] [CrossRef]
- Weir, D.L.; Coggins, S.A.; Vu, B.K.; Coertse, J.; Yan, L.; Smith, I.L.; Laing, E.D.; Markotter, W.; Broder, C.C.; Schaefer, B.C. Isolation and Characterization of Cross-Reactive Human Monoclonal Antibodies That Potently Neutralize Australian Bat Lyssavirus Variants and Other Phylogroup 1 Lyssaviruses. Viruses 2021, 13, 391. [Google Scholar] [CrossRef]
- Gong, X.; Zhu, M.; Li, G.; Lu, X.; Wan, Y. Specific determination of influenza H7N2 virus based on biotinylated single-domain antibody from a phage displayed library. Anal. Biochem. 2016, 500, 66–72. [Google Scholar] [CrossRef] [PubMed]
- Ten Haaf, A.; Kohl, J.; Pscherer, S.; Hamann, H.P.; Eskens, H.U.; Bastian, M.; Gattenlöhner, S.; Tur, M.K. Development of a monoclonal sandwich ELISA for direct detection of bluetongue virus 8 in infected animals. J. Virol. Methods 2017, 243, 172–176. [Google Scholar] [CrossRef]
- Bhatia, S.; Gangil, R.; Gupta, D.S.; Sood, R.; Pradhan, H.K.; Dubey, S.C. Single-chain fragment variable antibody against the capsid protein of bovine immunodeficiency virus and its use in ELISA. J. Virol. Methods 2010, 167, 68–73. [Google Scholar] [CrossRef]
- Li, T.; Huang, M.; Xiao, H.; Zhang, G.; Ding, J.; Wu, P.; Zhang, H.; Sheng, J.; Chen, C. Selection and characterization of specific nanobody against bovine virus diarrhea virus (BVDV) E2 protein. PLoS ONE 2017, 12, e0178469. [Google Scholar] [CrossRef] [Green Version]
- Duan, H.; Ma, Z.; Xu, L.; Zhang, A.; Li, Z.; Xiao, S. A Novel Intracellularly Expressed NS5B-Specific Nanobody Suppresses Bovine Viral Diarrhea Virus Replication. Vet. Microbiol. 2020, 240, 108449. [Google Scholar] [CrossRef]
- Ghannam, A.; Kumari, S.; Muyldermans, S.; Abbady, A.Q. Camelid nanobodies with high affinity for broad bean mottle virus: A possible promising tool to immunomodulate plant resistance against viruses. Plant Mol. Biol. 2015, 87, 355–369. [Google Scholar] [CrossRef] [PubMed]
- Braganza, A.; Wallace, K.; Pell, L.; Parrish, C.R.; Siegel, D.L.; Mason, N.J. Generation and validation of canine single chain variable fragment phage display libraries. Vet. Immunol. Immunopathol. 2011, 139, 27–40. [Google Scholar] [CrossRef] [PubMed]
- Ge, S.; Xu, L.; Li, B.; Zhong, F.; Liu, X.; Zhang, X. Canine parvovirus is diagnosed and neutralized by chicken IgY-scFv generated against the virus capsid protein. Vet. Res. 2020, 51, 110. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.L.; Shriver-Lake, L.C.; Zabetakis, D.; Anderson, G.P.; Goldman, E.R. Selection and characterization of protective anti-chikungunya virus single domain antibodies. Mol. Immunol. 2019, 105, 190–197. [Google Scholar] [CrossRef] [PubMed]
- Ziegler, A.; Torrance, L.; Macintosh, S.M.; Cowan, G.H.; Mayo Cucumber, M.A. Mosaic cucumovirus antibodies from a synthetic phage display library. Virology 1995, 214, 235–238. [Google Scholar] [CrossRef] [Green Version]
- Moreland, N.J.; Tay, M.Y.F.; Lim, E.; Paradkar, P.N.; Doan, D.N.P.; Yau, Y.H.; Geifman Shochat, S.; Vasudevan, S.G. high affinity human antibody fragments to dengue virus nonstructural protein 3. PLoS Negl. Trop. Dis. 2010, 4, e881. [Google Scholar] [CrossRef]
- Shriver-Lake, L.C.; Liu, J.L.; Zabetakis, D.; Sugiharto, V.A.; Lee, C.R.; Defang, G.N.; Wu, S.L.; Anderson, G.P.; Goldman, E.R. Selection and characterization of anti-dengue NS1 single domain antibodies. Sci. Rep. 2018, 8, 18086. [Google Scholar] [CrossRef] [Green Version]
- Tunghirun, C.; Narkthong, V.; Chaicumpa, W.; Chimnaronk, S. Interference of dengue replication by blocking the access of 3′ SL RNA to the viral RNA-dependent RNA polymerase. Antivir. Res. 2020, 182, 104921. [Google Scholar] [CrossRef]
- Wang, Y.; Cui, P.; Zhu, S.; Meng, T.; Hao, F.; Zhu, G.; Zuo, W. Preparation of single-chain antibody against VP3 protein of duck hepatitis virus type 1 by phage display technology. J. Virol. Methods 2018, 257, 73–78. [Google Scholar] [CrossRef]
- Xue, W.; Zhao, Q.; Li, P.; Zhang, R.; Lan, J.; Wang, J.; Yang, X.; Xie, Z.; Jiang, S. Identification and characterization of a novel nanobody against duck hepatitis A virus type 1. Virology 2019, 528, 101–109. [Google Scholar] [CrossRef]
- Shingarova, L.N.; Tikunova, N.V.; Iun, T.E.; Chepurnov, A.A.; Aliev, T.K.; Batanova, T.A.; Boldyreva, E.F.; Nekrasova, O.V.; Toporova, V.A.; Panina, A.A.; et al. Recombinant full-size human antibody to Ebola virus. Bioorg. Khim. 2007, 33, 598–605. [Google Scholar] [CrossRef]
- Goodchild, S.A.; Dooley, H.; Schoepp, R.J.; Flajnik, M.; Lonsdale, S.G. Isolation and characterisation of ebolavirus-specific recombinant antibody fragments from murine and shark immune libraries. Mol. Immunol. 2011, 48, 2027–2037. [Google Scholar] [CrossRef]
- Flego, M.; Frau, A.; Accardi, L.; Mallano, A.; Ascione, A.; Gellini, M.; Fanunza, E.; Vella, S.; Di Bonito, P.; Tramontano, E. Intracellular human antibody fragments recognizing the VP35 protein of Zaire Ebola filovirus inhibit the protein activity. BMC Biotechnol. 2019, 19, 64. [Google Scholar] [CrossRef]
- Froude, J.W.; Herbert, A.S.; Pelat, T.; Miethe, S.; Zak, S.E.; Brannan, J.M.; Bakken, R.R.; Steiner, A.R.; Yin, G.; Hallam, T.J.; et al. Post-exposure protection in mice against sudan virus by a two antibody cocktail. Viruses 2018, 10, 286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herbert, A.S.; Froude, J.W.; Ortiz, R.A.; Kuehne, A.I.; Dorosky, D.E.; Bakken, R.R.; Zak, S.E.; Josleyn, N.M.; Musiychuk, K.; Jones, R.M.; et al. Development of an antibody cocktail for treatment of Sudan virus infection. Proc. Natl. Acad. Sci. USA 2020, 117, 3768–3778. [Google Scholar] [CrossRef]
- Khlusevich, Y.; Matveev, A.; Baykov, I.; Bulychev, L.; Bormotov, N.; Ilyichev, I.; Shevelev, G.; Morozova, V.; Pyshnyi, D.; Tikunova, N. Phage display antibodies against ectromelia virus that neutralize variola virus: Selection and implementation for p35 neutralizing epitope mapping. Antivir. Res. 2018, 152, 18–25. [Google Scholar] [CrossRef]
- Chen, Z.; Ren, X.; Yang, J.; Dong, J.; Xue, Y.; Sun, L.; Zhu, Y.; Jin, Q. An elaborate landscape of the human antibody repertoire against enterovirus 71 infection is revealed by phage display screening and deep sequencing. mAbs 2017, 9, 342–349. [Google Scholar] [CrossRef] [Green Version]
- Thanongsaksrikul, J.; Srimanote, P.; Tongtawe, P.; Glab-Ampai, K.; Malik, A.A.; Supasorn, O.; Chiawwit, P.; Poovorawan, Y.; Chaicumpa, W. Identification and production of mouse scFv to specific epitope of enterovirus-71 virion protein-2 (VP2). Arch. Virol. 2018, 163, 1141–1152. [Google Scholar] [CrossRef]
- Phanthong, S.; Densumite, J.; Seesuay, W.; Thanongsaksrikul, J.; Teimoori, S.; Sookrung, N.; Poovorawan, Y.; Onvimala, N.; Guntapong, R.; Pattanapanyasat, K.; et al. Human Antibodies to VP4 Inhibit Replication of Enteroviruses Across Subgenotypes and Serotypes, and Enhance Host Innate Immunity. Front. Microbiol. 2020, 11, 562768. [Google Scholar] [CrossRef]
- Zhang, D.; Mao, Y.; Cao, Q.; Xiong, L.; Wen, J.; Chen, R.; Zhu, J. Generation and Characterization of a Novel Recombinant Antibody Against LMP1-TES1 of Epstein-Barr Virus Isolated by Phage Display. Viruses 2013, 5, 1131–1142. [Google Scholar] [CrossRef] [Green Version]
- Foord, A.J.; Muller, J.D.; Yu, M.; Wang, L.-F.; Heine, H.G. Production and application of recombinant antibodies to foot-and-mouth disease virus non-structural protein 3ABC. J. Immunol. Methods 2007, 321, 142–151. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Yang, S.; Yin, S.; Shang, Y.; Du, P.; Guo, J.; He, J.; Cai, J.; Liu, X. Characterization of single-domain antibodies against Foot and Mouth Disease Virus (FMDV) serotype O from a camelid and imaging of FMDV in baby hamster kidney-21 cells with single-domain antibody-quantum dots probes. BMC Vet. Res. 2015, 11, 120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harmsen, M.M.; Seago, J.; Perez, E.; Charleston, B.; Eblé, P.L.; Dekker, A. Isolation of single-domain antibody fragments that preferentially detect intact (146s) particles of foot-and-mouth disease virus for use in vaccine quality control. Front. Immunol. 2017, 8, 3389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salem, R.; El-Kholy, A.A.; Ibrahim, M. Eight novel single chain antibody fragments recognising VP2 of foot-and-mouth disease virus serotypes A, O, and SAT 2. Virology 2019, 533, 145–154. [Google Scholar] [CrossRef]
- Orecchia, M.; Nölke, G.; Saldarelli, P.; Dell’Orco, M.; Uhde-Holzem, K.; Sack, M.; Martelli, G.; Fischer, R.; Schillberg, S. Generation and characterization of a recombinant antibody fragment that binds to the coat protein of grapevine leafroll-associated virus 3. Arch. Virol. 2008, 153, 1075–1084. [Google Scholar] [CrossRef] [PubMed]
- Saldarelli, P.; Keller, H.; Dell’Orco, M.; Schots, A.; Elicio, V.; Minafra, A. Isolation of recombinant antibodies (Scfvs) to grapevine virus B. J. Virol. Methods 2005, 124, 191–195. [Google Scholar] [CrossRef]
- Koch, J.; Liang, M.; Queitsch, I.; Kraus, A.A.; Bautz, E.K.F. Human recombinant neutralizing antibodies against hantaan virus G2 protein. Virology 2003, 308, 64–73. [Google Scholar] [CrossRef] [Green Version]
- Pereira, S.S.; Moreira-Dill, L.S.; Morais, M.S.S.; Prado, N.D.R.; Barros, M.L.; Koishi, A.C.; Mazarrotto, G.A.; Gonçalves, G.M.; Zuliani, J.P.; Calderon, L.A.; et al. Novel camelid antibody fragments targeting recombinant nucleoprotein of araucaria hantavirus: A prototype for an early diagnosis of hantavirus pulmonary syndrome. PLoS ONE 2014, 9, e108067. [Google Scholar] [CrossRef]
- Zhu, Z.; Bossart, K.N.; Bishop, K.A.; Crameri, G.; Dimitrov, A.S.; McEachern, J.A.; Feng, Y.; Middleton, D.; Wang, L.F.; Broder, C.C.; et al. Exceptionally potent cross-reactive neutralization of Nipah and Hendra viruses by a human monoclonal antibody. J. Infect. Dis. 2008, 197, 846–853. [Google Scholar] [CrossRef]
- Schofield, D.J.; Satterfield, W.; Emerson, S.U.; Purcell, R.H. Four chimpanzee monoclonal antibodies isolated by phage display neutralize hepatitis a virus. Virology 2002, 292, 127–136. [Google Scholar] [CrossRef] [Green Version]
- Wen, W.-H.; Qin, W.-J.; Gao, H.; Zhao, J.; Jia, L.-T.; Liao, Q.-H.; Meng, Y.-L.; Jin, B.-Q.; Yao, L.-B.; Chen, S.-Y.; et al. An hepatitis B virus surface antigen specific single chain of variable fragment derived from a natural immune antigen binding fragment phage display library is specifically internalized by HepG2.2.15 cells. J. Viral Hepat. 2007, 14, 512–519. [Google Scholar] [CrossRef]
- Barban, V.; Fraysse-Corgier, S.; Paranhos-Baccala, G.; Petit, M.; Manin, C.; Berard, Y.; Prince, A.M.; Mandrand, B.; Meulien, P. Identification of a human epitope in hepatitis C virus (HCV) core protein using a molecularly cloned antibody repertoire from a non-symptomatic, anti-HCV-positive patient. J. Gen. Virol. 2000, 81 Pt 2, 461–469. [Google Scholar] [CrossRef]
- Burioni, R.; Bugli, F.; Mancini, N.; Rosa, D.; Di Campli, C.; Moroncini, G.; Manzin, A.; Abrignani, S.; Varaldo, P.E.; Clementi, M.; et al. Nonneutralizing human antibody fragments against hepatitis C virus E2 glycoprotein modulate neutralization of binding activity of human recombinant Fabs. Virology 2001, 288, 29–35. [Google Scholar] [CrossRef] [Green Version]
- Jittavisutthikul, S.; Seesuay, W.; Thanongsaksrikul, J.; Thueng-In, K.; Srimanote, P.; Werner, R.G.; Chaicumpa, W. Human transbodies to HCV NS3/4A protease inhibit viral replication and restore host innate immunity. Front. Immunol. 2016, 7, 318. [Google Scholar] [CrossRef] [Green Version]
- Glab-Ampai, K.; Chulanetra, M.; Malik, A.A.; Juntadech, T.; Thanongsaksrikul, J.; Srimanote, P.; Thueng-In, K.; Sookrung, N.; Tongtawe, P.; Chaicumpa, W. Human single chain-transbodies that bound to domain-I of non-structural protein 5A (NS5A) of hepatitis C virus. Sci. Rep. 2017, 7, 15042. [Google Scholar] [CrossRef] [Green Version]
- Schofield, D.J.; Glamann, J.; Emerson, S.U.; Purcell, R.H. Identification by phage display and characterization of two neutralizing chimpanzee monoclonal antibodies to the hepatitis E virus capsid protein. J. Virol. 2000, 74, 5548–5555. [Google Scholar] [CrossRef] [Green Version]
- Sanna, P.P.; Williamson, R.A.; De Logu, A.; Bloom, F.E.; Burton, D.R. Directed selection of recombinant human monoclonal antibodies to herpes simplex virus glycoproteins from phage display libraries. Proc. Natl. Acad. Sci. USA 1995, 92, 6439–6443. [Google Scholar] [CrossRef] [Green Version]
- Nejatollahi, F.; Hodgetts, S.J.; Vallely, P.J.; Burnie, J.P. Neutralising human recombinant antibodies to human cytomegalovirus glycoproteins gB and gH. FEMS Immunol. Med. Microbiol. 2002, 34, 237–244. [Google Scholar] [CrossRef]
- Moazen, B.; Ebrahimi, E.; Nejatollahi, F. Single chain antibodies against Gp55 of human cytomegalovirus (HCMV) for prophylaxis and treatment of HCMV Infections. Jundishapur J. Microbiol. 2016, 9, e16241. [Google Scholar] [CrossRef] [Green Version]
- Gustchina, E.; Louis, J.M.; Lam, S.N.; Bewley, C.A.; Clore, G.M. A monoclonal Fab derived from a human nonimmune phage library reveals a new epitope on gp41 and neutralizes diverse human immunodeficiency virus type 1 strains. J. Virol. 2007, 81, 12946–12953. [Google Scholar] [CrossRef] [Green Version]
- da Silva, F.A.; Li, M.; Rato, S.; Maia, S.; Malhó, R.; Warren, K.; Harrich, D.; Craigie, R.; Barbas, C., 3rd; Goncalves, J. Recombinant rabbit single-chain antibodies bind to the catalytic and C-terminal domains of HIV-1 integrase protein and strongly inhibit HIV-1 replication. Biotechnol. Appl. Biochem. 2012, 59, 353–366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mohammadzadeh, S.; Rajabibazl, M.; Fourozandeh, M.; Rasaee, M.J.; Rahbarizadeh, F.; Mohammadi, M. Production of recombinant scFv Against P24 of human immunodeficiency virus type 1 by phage display technology. Monoclon. Antibodies Immunodiagn. Immunother. 2014, 33, 28–33. [Google Scholar] [CrossRef] [PubMed]
- Koch, K.; Kalusche, S.; Torres, J.L.; Stanfield, R.L.; Danquah, W.; Khazanehdari, K.; von Briesen, H.; Geertsma, E.R.; Wilson, I.A.; Wernery, U.; et al. Selection of nanobodies with broad neutralizing potential against primary HIV-1 strains using soluble subtype C gp140 envelope trimers. Sci. Rep. 2017, 7, 8390. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Kumar, R.; Khan, L.; Makhdoomi, M.A.; Thiruvengadam, R.; Mohata, M.; Agarwal, M.; Lodha, R.; Kabra, S.K.; Sinha, S.; et al. CD4-binding site directed cross-neutralizing scFv monoclonals from HIV-1 subtype C infected Indian children. Front. Immunol. 2017, 8, 1568. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; Qiao, Y.; Zhu, Y.; Chong, H.; He, Y. Identification of a novel HIV-1-neutralizing antibody from a CRF07_BC-infected chinese donor. Oncotarget 2017, 8, 63047–63063. [Google Scholar] [CrossRef] [Green Version]
- Williams, J.V.; Chen, Z.; Cseke, G.; Wright, D.W.; Keefer, C.J.; Tollefson, S.J.; Hessell, A.; Podsiad, A.; Shepherd, B.E.; Sanna, P.P.; et al. A recombinant human monoclonal antibody to human metapneumovirus fusion protein that neutralizes virus in vitro and is effective therapeutically in vivo. J. Virol. 2007, 81, 8315–8324. [Google Scholar] [CrossRef] [Green Version]
- Rajput, R.; Sharma, G.; Rawat, V.; Gautam, A.; Kumar, B.; Pattnaik, B.; Pradhan, H.K.; Khanna, M. Diagnostic potential of recombinant scFv antibodies generated against hemagglutinin protein of influenza A virus. Front. Immunol. 2015, 6, 440. [Google Scholar] [CrossRef] [Green Version]
- Wyrzucki, A.; Dreyfus, C.; Kohler, I.; Steck, M.; Wilson, I.A.; Hangartner, L. Alternative recognition of the conserved stem epitope in influenza a virus hemagglutinin by a VH3-30-encoded heterosubtypic antibody. J. Virol. 2014, 88, 7083–7092. [Google Scholar] [CrossRef] [Green Version]
- Vashisht, S.; Verma, S.; Salunke, D.M. Cross-clade antibody reactivity may attenuate the ability of influenza virus to evade the immune response. Mol. Immunol. 2019, 114, 149–161. [Google Scholar] [CrossRef]
- Yodsheewan, R.; Maneewatch, S.; Srimanote, P.; Thueng-In, K.; Songserm, T.; Dong-Din-On, F.; Bangphoomi, K.; Sookrung, N.; Choowongkomon, K.; Chaicumpa, W. Human monoclonal ScFv specific to NS1 protein inhibits replication of influenza viruses across types and subtypes. Antivir. Res. 2013, 100, 226–237. [Google Scholar] [CrossRef]
- Sengupta, D.; Shaikh, A.; Bhatia, S.; Pateriya, A.K.; Khandia, R.; Sood, R.; Prakash, A.; Pattnaik, B.; Pradhan, H.K. Development of single-chain Fv against the nucleoprotein of type A influenza virus and its use in ELISA. J. Virol. Methods 2014, 208, 129–137. [Google Scholar] [CrossRef]
- Wu, J.; Zeng, X.Q.; Zhang, H.B.; Ni, H.Z.; Pei, L.; Zou, L.R.; Liang, L.J.; Zhang, X.; Lin, J.Y.; Ke, C.W. Novel phage display-derived H5N1-specific scFvs with potential use in rapid avian flu diagnosis. J. Microbiol. Biotechnol. 2014, 24, 704–713. [Google Scholar] [CrossRef] [Green Version]
- Zhu, M.; Gong, X.; Hu, Y.; Ou, W.; Wan, Y. Streptavidin-biotin-based directional double nanobody sandwich ELISA for clinical rapid and sensitive detection of influenza H5N1. J. Transl. Med. 2014, 12, 352. [Google Scholar] [CrossRef] [Green Version]
- Velappan, N.; Micheva-Viteva, S.; Adikari, S.H.; Waldo, G.S.; Lillo, A.M.; Bradbury, A.R.M. Selection and verification of antibodies against the cytoplasmic domain of M2 of influenza, a transmembrane protein. mAbs 2020, 12, 1843754. [Google Scholar] [CrossRef]
- Hirano, D.; Ohshima, N.; Kubota-Koketsu, R.; Yamasaki, A.; Kurosawa, G.; Okuno, Y.; Yoshida, S.; Kurosawa, Y. Three types of broadly reacting antibodies against influenza B viruses induced by vaccination with seasonal influenza viruses. J. Immunol. Res. 2018, 8, 7251793. [Google Scholar] [CrossRef] [Green Version]
- Ramage, W.; Gaiotto, T.; Ball, C.; Risley, P.; Carnell, G.W.; Temperton, N.; Cheung, C.Y.; Engelhardt, O.G.; Hufton, S.E. Cross-reactive and lineage-specific single domain antibodies against influenza b hemagglutinin. Antibodies 2019, 8, 14. [Google Scholar] [CrossRef] [Green Version]
- Arakawa, M.; Yamashiro, T.; Uechi, G.; Tadano, M.; Nishizono, A. Construction of human Fab (Gamma1/Kappa) library and identification of human monoclonal Fab possessing neutralizing potency against Japanese encephalitis virus. Microbiol. Immunol. 2007, 51, 617–625. [Google Scholar] [CrossRef] [Green Version]
- Sherwood, L.J.; Osborn, L.E.; Carrion, R., Jr.; Patterson, J.L.; Hayhurst, A. Rapid assembly of sensitive antigen-capture assays for Marburg virus, using in vitro selection of llama single-domain antibodies, at biosafety level 4. J. Infect. Dis. 2007, 196 (Suppl. S2), S213–S219. [Google Scholar] [CrossRef] [Green Version]
- Froude, J.W.; Pelat, T.; Miethe, S.; Zak, S.E.; Wec, A.Z.; Chandran, K.; Brannan, J.M.; Bakken, R.R.; Hust, M.; Thullier, P.; et al. Generation and characterization of protective antibodies to Marburg virus. mAbs 2017, 9, 696–703. [Google Scholar] [CrossRef] [Green Version]
- Amatya, P.; Wagner, N.; Chen, G.; Luthra, P.; Shi, L.; Borek, D.; Pavlenco, A.; Rohrs, H.; Basler, C.F.; Sidhu, S.S.; et al. Inhibition of marburg virus RNA synthesis by a synthetic anti-VP35 antibody. ACS Infect. Dis. 2019, 5, 1385–1396. [Google Scholar] [CrossRef]
- Kim, Y.; Lee, H.; Park, K.; Park, S.; Lim, J.H.; So, M.K.; Woo, H.M.; Ko, H.; Lee, J.M.; Lim, S.H.; et al. Selection and characterization of monoclonal antibodies targeting middle east respiratory syndrome coronavirus through a human synthetic Fab phage display library panning. Antibodies 2019, 8, 42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, C.C.; Woo, P.C.Y.; Lim, T.S. Development of a phage display panning strategy utilizing crude antigens: Isolation of MERS-CoV nucleoprotein human antibodies. Sci. Rep. 2019, 9, 6088. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, W.; Samanta, M.; Crawford, S.E.; Estes, M.K.; Neill, F.H.; Atmar, R.L.; Palzkill, T. Identification of human single-chain antibodies with broad reactivity for noroviruses. Protein Eng. Des. Sel. 2014, 27, 339–349. [Google Scholar] [CrossRef] [PubMed]
- Garaicoechea, L.; Aguilar, A.; Parra, G.I.; Bok, M.; Sosnovtsev, S.V.; Canziani, G.; Green, K.Y.; Bok, K.; Parreño, V. Llama nanoantibodies with therapeutic potential against human norovirus diarrhea. PLoS ONE 2015, 10, e0133665. [Google Scholar] [CrossRef] [PubMed]
- Hurwitz, A.M.; Huang, W.; Kou, B.; Estes, M.K.; Atmar, R.L.; Palzkill, T. Identification and characterization of single-chain antibodies that specifically bind GI noroviruses. PLoS ONE 2017, 12, e0170162. [Google Scholar] [CrossRef] [Green Version]
- Welch, B.D.; Paduch, M.; Leser, G.P.; Bergman, Z.; Kors, C.A.; Paterson, R.G.; Jardetzky, T.S.; Kossiakoff, A.A.; Lamb, R.A. Probing the functions of the paramyxovirus glycoproteins F and HN with a panel of synthetic antibodies. J. Virol. 2014, 88, 11713–11725. [Google Scholar] [CrossRef] [Green Version]
- Hust, M.; Maiss, E.; Jacobsen, H.-J.; Reinard, T. The production of a genus-specific recombinant antibody (scFv) using a recombinant potyvirus protease. J. Virol. Methods 2002, 106, 225–233. [Google Scholar] [CrossRef]
- Chen, Z.; Chumakov, K.; Dragunsky, E.; Kouiavskaia, D.; Makiya, M.; Neverov, A.; Rezapkin, G.; Sebrell, A.; Purcell, R. Chimpanzee-human monoclonal antibodies for treatment of chronic poliovirus excretors and emergency postexposure prophylaxis. J. Virol. 2011, 85, 4354–4362. [Google Scholar] [CrossRef] [Green Version]
- Chen, G.; Gorelik, L.; Simon, K.J.; Pavlenco, A.; Cheung, A.; Brickelmaier, M.; Chen, L.; Jin, P.; Weinreb, P.H.; Sidhu, S.S. Synthetic antibodies and peptides recognizing progressive multifocal leukoencephalopathy-specific point mutations in Polyomavirus JC capsid viral protein 1. mAbs 2015, 7, 681–692. [Google Scholar] [CrossRef]
- Yang, S.; Shang, Y.; Yin, S.; Wang, D.; Cai, J.; Gong, Z.; Serge, M.; Liu, X. A phage-displayed single domain antibody fused to alkaline phosphatase for detection of porcine circovirus type 2. J. Virol. Methods 2015, 213, 84–92. [Google Scholar] [CrossRef]
- Mu, Y.; Jia, C.; Zheng, X.; Zhu, H.; Zhang, X.; Xu, H.; Liu, B.; Zhao, Q.; Zhou, E.M. A nanobody-horseradish peroxidase fusion protein-based competitive ELISA for rapid detection of antibodies against porcine circovirus type 2. J. Nanobiotechnol. 2021, 19, 34. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Li, L.; Yin, S.; Shang, Y.; Khan, M.U.Z.; He, X.; Yuan, L.; Gao, X.; Liu, X.; Cai, J. Single-domain antibodies as promising experimental tools in imaging and isolation of porcine epidemic diarrhea virus. Appl. Microbiol. Biotechnol. 2018, 102, 8931–8942. [Google Scholar] [CrossRef] [PubMed]
- Bao, F.; Wang, L.; Zhao, X.; Lu, T.; Na, A.M.; Wang, X.; Cao, J.; Du, Y. Preparation and characterization of a single-domain antibody specific for the porcine epidemic diarrhea virus spike protein. AMB Express 2019, 9, 104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, Z.; Wang, T.; Li, Z.; Guo, X.; Tian, Y.; Li, Y.; Xiao, S. A novel biotinylated nanobody-based blocking ELISA for the rapid and sensitive clinical detection of porcine epidemic diarrhea virus. J. Nanobiotechnol. 2019, 17, 96. [Google Scholar] [CrossRef] [Green Version]
- Zhang, F.; Chen, Y.; Ke, Y.; Zhang, L.; Zhang, B.; Yang, L.; Zhu, J. Single chain fragment variable (scFv) antibodies targeting the spike protein of porcine epidemic diarrhea virus provide protection against viral infection in piglets. Viruses 2019, 11, 58. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Wang, Y.; Duan, H.; Zhang, A.; Liang, C.; Gao, J.; Zhang, C.; Huang, B.; Li, Q.; Li, N.; et al. An intracellularly expressed Nsp9-specific nanobody in MARC-145 cells inhibits porcine reproductive and respiratory syndrome virus replication. Vet. Microbiol. 2015, 181, 252–260. [Google Scholar] [CrossRef]
- Liu, H.; Liang, C.; Duan, H.; Zhang, X.; Wang, X.; Xiao, S.; Zhou, E.M. Intracellularly expressed nanobodies against non-structural protein 4 of porcine reproductive and respiratory syndrome virus inhibit virus replication. Biotechnol. Lett. 2016, 38, 1081–1088. [Google Scholar] [CrossRef]
- Sun, L.; Chen, Z.; Yu, L.; Wei, J.; Li, C.; Jin, J.; Shen, X.; Lv, X.; Tang, Q.; Li, D.; et al. Generation and characterization of neutralizing human recombinant antibodies against antigenic site II of rabies virus glycoprotein. Appl. Microbiol. Biotechnol. 2012, 96, 357–366. [Google Scholar] [CrossRef]
- Boruah, B.M.; Liu, D.; Ye, D.; Gu, T.J.; Jiang, C.L.; Qu, M.; Wright, E.; Wang, W.; He, W.; Liu, C.; et al. Single domain antibody multimers confer protection against rabies infection. PLoS ONE 2013, 8, e71383. [Google Scholar] [CrossRef]
- Chen, Z.; Zhang, L.; Tang, A.; Callahan, C.; Pristatsky, P.; Swoyer, R.; Cejas, P.; Nahas, D.; Galli, J.; Cosmi, S.; et al. Discovery and characterization of phage display-derived human monoclonal antibodies against RSV F glycoprotein. PLoS ONE 2016, 11, e0156798. [Google Scholar] [CrossRef] [Green Version]
- Rodrıguez-Dıaz, J.; Monedero, V.; Perez-Martınez, G.; Buesa, J. Single-chain variable fragment (scFv) antibodies against rotavirus NSP4 enterotoxin generated by phage display. J. Virol. Methods 2004, 121, 231–238. [Google Scholar] [CrossRef]
- Monedero, V.; Rodrıguez-Dıaz, J.; Viana, R.; Buesa, J.; Perez-Martınez, G. Selection of single-chain antibodies against the vp8* subunit of rotavirus VP4 outer capsid protein and their expression in Lactobacillus casei. Appl. Environ. Microbiol. 2004, 70, 6936–6939. [Google Scholar] [CrossRef] [Green Version]
- Bracken, C.J.; Lim, S.A.; Solomon, P.; Rettko, N.J.; Nguyen, D.P.; Zha, B.S.; Schaefer, K.; Byrnes, J.R.; Zhou, J.; Lui, I.; et al. QCRG Structural Biology Consortium; Zhou XX, Leung KK, Wells JA. Bi-paratopic and multivalent VH domains block ACE2 binding and neutralize SARS-CoV-2. Nat. Chem. Biol. 2021, 17, 113–121. [Google Scholar] [CrossRef]
- Bertoglio, F.; Fühner, V.; Ruschig, M.; Heine, P.A.; Abasi, L.; Klünemann, T.; Rand, U.; Meier, D.; Langreder, N.; Steinke, S.; et al. A SARS-CoV-2 neutralizing antibody selected from COVID-19 patients by phage display is binding to the ACE2-RBD interface and is tolerant to most known recently emerging RBD mutations. bioRxiv 2020. [Google Scholar] [CrossRef]
- Ku, Z.; Xie, X.; Davidson, E.; Ye, X.; Su, H.; Menachery, V.D.; Li, Y.; Yuan, Z.; Zhang, X.; Muruato, A.E.; et al. Molecular determinants and mechanism for antibody cocktail preventing SARS-CoV-2 escape. Nat. Commun. 2021, 12, 469. [Google Scholar] [CrossRef]
- Lim, S.A.; Gramespacher, J.A.; Pance, K.; Rettko, N.J.; Solomon, P.; Jin, J.; Lui, I.; Elledge, S.K.; Liu, J.; Bracken, C.J.; et al. Bispecific VH/Fab antibodies targeting neutralizing and non-neutralizing Spike epitopes demonstrate enhanced potency against SARS-CoV-2. mAbs 2021, 13, 1893426. [Google Scholar] [CrossRef]
- Ma, H.; Zeng, W.; Meng, X.; Huang, X.; Yang, Y.; Zhao, D.; Zhou, P.; Wang, X.; Zhao, C.; Sun, Y.; et al. Potent Neutralization of SARS-CoV-2 by hetero-bivalent alpaca nanobodies targeting the spike receptor-binding domain. J. Virol. 2021, 95, e02438-20. [Google Scholar] [CrossRef]
- Gai, J.; Ma, L.; Li, G.; Zhu, M.; Qiao, P.; Li, X.; Zhang, H.; Zhang, Y.; Chen, Y.; Ji, W.; et al. A potent neutralizing nanobody against SARS-CoV-2 with inhaled delivery potential. MedComm 2021, 2, 101–113. [Google Scholar] [CrossRef]
- Slezak, T.; Kossiakoff, A.A. Engineered ultra-high affinity synthetic antibodies for SARS-CoV-2 neutralization and detection. J. Mol. Biol. 2021, 433, 166956. [Google Scholar] [CrossRef]
- Zhou, D.; Chan, J.F.; Zhou, B.; Zhou, R.; Li, S.; Shan, S.; Liu, L.; Zhang, A.J.; Chen, S.J.; Chan, C.C.; et al. Robust SARS-CoV-2 infection in nasal turbinates after treatment with systemic neutralizing antibodies. Cell Host Microbe 2021, 29, 551–563.e5. [Google Scholar] [CrossRef]
- Kim, H.-Y.; Lee, J.-H.; Kim, M.J.; Park, S.C.; Choi, M.; Lee, W.; Ku, K.B.; Kim, B.T.; Changkyun Park, E.; Kim, H.G.; et al. Development of a SARS-CoV-2-specific biosensor for antigen detection using scFv-Fc fusion proteins. Biosens. Bioelectron. 2021, 175, 112868. [Google Scholar] [CrossRef] [PubMed]
- Ita, S.; Agostinho, M.R.; Sullivan, K.; Yub Han, S.; Akleh, R.; Johnson, W.E.; Fofana, I.B.F. Analysis of SIVmac Envelope-Specific Antibodies Selected Through Phage Display. AIDS Res. Hum. Retrovir. 2017, 33, 869–879. [Google Scholar] [CrossRef] [PubMed]
- Du, T.; Zhu, G.; Wu, X.; Fang, J.; Zhou, E.-M. Biotinylated single domain antibody-based blocking ELISA for detection of antibodies against swine influenza virus. Int. J. Nanomed. 2019, 14, 9337–9349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, F.; Chen, Y.; Yang, L.; Zhu, J. Construction and characterization of porcine single-chain fragment variable antibodies that neutralize transmissible gastroenteritis virus in vitro. Arch. Virol. 2019, 164, 983–994. [Google Scholar] [CrossRef] [PubMed]
- Schoenenwald, A.K.J.; Gwee, C.P.; Stiasny, K.; Hermann, M.; Vasudevan, S.G.; Skern, T. Development and characterization of specific antiusutu virus chicken-derived single chain variable fragment antibodies. Protein Sci. Publ. Protein Soc. 2020, 29, 2175–2188. [Google Scholar] [CrossRef]
- Shin, Y.W.; Chang, K.-H.; Hong, G.-W.; Yeo, S.-G.; Jee, Y.; Kim, J.-H.; Oh, M.-d.; Cho, D.-H.; Kim, S.-H. Selection of vaccinia virus-neutralizing antibody from a phage display human-antibody library. J. Microbiol. Biotechnol. 2019, 29, 651–657. [Google Scholar] [CrossRef] [Green Version]
- Yuan, L.; Zhang, X.; Xiao, N.; Dai, L.; Chen, W.; Jia, C.; Zhao, R.; Hemmingsen, S.M.; Dai, H. Identification of a WSSV neutralizing scFv antibody by phage display technology and in vitro screening. Dis. Aquat. Organ. 2006, 72, 93–99. [Google Scholar] [CrossRef] [Green Version]
- Duan, T.; Ferguson, M.; Yuan, L.; Xu, F.; Li, G. Human monoclonal Fab antibodies against west nile virus and its neutralizing activity analyzed in vitro and in vivo. J. Antivir. Antiretrovir. 2009, 1, 36–42. [Google Scholar] [CrossRef] [Green Version]
- Rizzo, S.; Imperato, P.; Mora-Cárdenas, E.; Konstantinidou, S.; Marcello, A.; Sblattero, D. Selection and characterization of highly specific recombinant antibodies against west nile virus E protein. J. Biotechnol. 2020, 311, 35–43. [Google Scholar] [CrossRef]
- Long, M.C.; Jager, S.; Mah, D.C.; Jebailey, L.; Mah, M.A.; Masri, S.A.; Nagata, L.P. Construction and characterization of a novel recombinant single-chain variable fragment antibody against Western equine encephalitis virus. Hybridoma 2000, 19, 1–13. [Google Scholar] [CrossRef]
- Burke, C.W.; Froude, J.W.; Miethe, S.; Hülseweh, B.; Hust, M.; Glass, P.J. Human-like neutralizing antibodies protect mice from aerosol exposure with western equine encephalitis virus. Viruses 2018, 10, 147. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.L.; Shriver-Lake, L.C.; Zabetakis, D.; Goldman, E.R.; Anderson, G.P. Selection of single-domain antibodies towards western equine encephalitis virus. Antibodies 2018, 7, 44. [Google Scholar] [CrossRef] [Green Version]
- Lu, X.; Xiao, H.; Li, S.; Pang, X.; Song, J.; Liu, S.; Cheng, H.; Li, Y.; Wang, X.; Huang, C.; et al. Double lock of a human neutralizing and protective monoclonal antibody targeting the yellow fever virus envelope. Cell Rep. 2019, 26, 438–446.e5. [Google Scholar] [CrossRef] [Green Version]
- Mwale, P.F.; Lee, C.H.; Lin, L.T.; Leu, S.J.; Huang, Y.J.; Chiang, L.C.; Mao, Y.C.; Yang, Y.Y. Expression, purification, and characterization of anti-Zika virus envelope protein: Polyclonal and chicken-derived single chain variable fragment antibodies. Int. J. Mol. Sci. 2020, 21, 492. [Google Scholar] [CrossRef] [Green Version]
- Delfin-Riela, T.; Rossotti, M.; Alvez-Rosado, R.; Leizagoyen, C.; González-Sapienza, G. Highly sensitive detection of Zika virus nonstructural protein 1 in serum samples by a two-site nanobody ELISA. Biomolecules 2020, 10, 1652. [Google Scholar] [CrossRef]
- Liu, J.; Ma, P.; Yu, H.; Wang, M.; Yin, P.; Pang, S.; Jiao, Y.; Dong, T.; Liu, A. Discovery of a phage peptide specifically binding to the SARS-CoV-2 spike S1 protein for the sensitive phage-based enzyme-linked chemiluminescence immunoassay of the SARS-CoV-2 antigen. Anal. Chem. 2022, 94, 11591–11599. [Google Scholar] [CrossRef]
- Li, H.; Zhu, B.; Li, B.; Chen, L.; Ning, X.; Dong, H.; Liang, J.; Yang, X.; Dong, J.; Ueda, H. Isolation of a human SARS-CoV-2 neutralizing antibody from a synthetic phage library and its conversion to fluorescent biosensors. Sci. Rep. 2022, 12, 15496. [Google Scholar] [CrossRef]
- Chen, X.; Kang, S.; Ikbal, M.A.; Zhao, Z.; Pan, Y.; Zuo, J.; Gu, L.; Wang, C. Synthetic nanobody-functionalized nanoparticles for accelerated development of rapid, accessible detection of viral antigens. Biosens. Bioelectron. 2022, 202, 113971. [Google Scholar] [CrossRef]
- Guliy, O.I.; Zaitsev, B.D.; Borodina, I.A.; Shikhabudinov, A.M.; Staroverov, S.A.; Dykman, L.A.; Fomin, A.S. Electro-acoustic sensor for the real-time identification of the bacteriophages. Talanta 2018, 178, 743–750. [Google Scholar] [CrossRef]
- Yang, F.; Liu, L.; Neuenschwander, P.F.; Idell, S.; Vankayalapati, R.; Jain, K.G.; Du, K.; Ji, H.; Yi, G. Phage display-derived peptide for the specific binding of SARS-CoV-2. ACS Omega 2021, 7, 3203–3211. [Google Scholar] [CrossRef]
- Griep, R.A.; Prins, M.; van Twisk, C.; Keller, H.J.; Kerschbaumer, R.J.; Kormelink, R.; Goldbach, R.W.; Schots, A. Application of phage display in selecting Tomato spotted wilt virus-specific single-chain antibodies (scFvs) for sensitive diagnosis in ELISA. Phytopathology 2000, 90, 183–190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, J.; Sakurai, A.; Nomura, N.; Park, E.Y.; Shibasaki, F.; Ueda, H. Isolation of recombinant phage antibodies targeting the hemagglutinin cleavage site of highly pathogenic avian influenza virus. PLoS ONE 2013, 8, e61158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, C.-M.; Chen, I.-C.; Tung, C.-P.; Peng, H.-P.; Jian, J.-W.; Chiu, Y.-K.; Tsou, Y.-L.; Chen, H.-S.; Huang, Y.-J.; Hsiao, W.W.-W.; et al. A panel of anti-influenza virus nucleoprotein antibodies selected from phage-displayed synthetic antibody libraries with rapid diagnostic capability to distinguish diverse influenza virus subtypes. Sci. Rep. 2020, 10, 13318. [Google Scholar] [CrossRef] [PubMed]
- Cabezas, S.; Rojas, G.; Pavon, A.; Bernardo, L.; Castellanos, Y.; Alvarez, M.; Pupo, M.; Guillen, G.; Guzman, M.G. Phage-displayed antibody fragments recognizing dengue 3 and dengue 4 viruses as tools for viral serotyping in sera from infected individuals. Arch. Virol. 2009, 154, 1035–1045. [Google Scholar] [CrossRef]
- Pashaki, A.S.; Safarnejad, M.R.; Safdar, A.H.A.; Safarpour, H.; Tabatabaie, M. Production of a phage-displayed single chain variable fragment antibody against infectious bursal disease virus. Trop. J. Pharm. Res. 2017, 16, 2801–2809. [Google Scholar] [CrossRef] [Green Version]
- Sapats, S.; Gould, G.; Trinidad, L.; Parede, L.H.; David, C.; Ignjatovic, J. An ELISA for detection of infectious bursal disease virus and differentiation of very virulent strains based on single chain recombinant chicken antibodies. Avian Pathol. 2005, 34, 449–455. [Google Scholar] [CrossRef] [Green Version]
- Sapats, S.I.; Trinidad, L.; Gould, G.; Heine, H.G.; van den Berg, T.P.; Eterradossi, N.; Jackwood, D.; Parede, L.; Toquin, D.; Ignjatovic, J. Chicken recombinant antibodies specific for very virulent infectious bursal disease virus. Arch. Virol. 2006, 151, 1551–1566. [Google Scholar] [CrossRef]
- Sharma, G.K.; Mahajan, S.; Matura, R.; Subramaniam, S.; Mohapatra, J.K.; Pattnaik, B. Production and characterization of single-chain antibody (scFv) against 3ABC non-structural protein in Escherichia coli for sero-diagnosis of Foot and Mouth Disease virus. Biologicals 2014, 42, 339–345. [Google Scholar] [CrossRef]
- Chen, S.; Li, S.; Sun, H.; Li, Y.; Ji, S.; Song, K.; Zhang, L.; Luo, Y.; Sun, Y.; Ma, J.; et al. Expression and characterization of a recombinant porcinized antibody against the E2 protein of classical swine fever virus. Appl. Microbiol. Biotechnol. 2018, 102, 961–970. [Google Scholar] [CrossRef]
- Duggan, J.M.; Coates, D.M.; Ulaeto, D.O. Isolation of single-chain antibody fragments against Venezuelan equine encephalomyelitis virus from two different immune sources. Viral Immunol. 2001, 14, 263–273. [Google Scholar] [CrossRef]
- Homola, J. Surface plasmon resonance sensors for detection of chemical and biological species. Chem. Rev. 2008, 108, 462–493. [Google Scholar] [CrossRef]
- Howes, P.D.; Rana, S.; Stevens, M.M. Plasmonic nanomaterials for biodiagnostics. Chem. Soc. Rev. 2014, 43, 3835–3853. [Google Scholar] [CrossRef]
- Walper, S.A.; Aragonés, G.L.; Sapsford, K.E.; Brown, C.W., III; Rowland, C.E.; Breger, J.C.; Medintz, I.L. Detecting biothreat agents: From current diagnostics to developing sensor technologies. ACS Sens. 2018, 3, 1894–2024. [Google Scholar] [CrossRef] [Green Version]
- Torrance, L.; Ziegler, A.; Pittman, H.; Paterson, M.; Toth, R.; Eggleston, I. Oriented immobilisation of engineered single-chain antibodies to develop biosensors for virus detection. J. Virol. Methods 2006, 134, 164–170. [Google Scholar] [CrossRef]
- Kim, J.W.; Cho, A.H.; Shin, H.G.; Jang, S.H.; Cho, S.Y.; Lee, Y.R.; Lee, S. Development and characterization of phage display-derived monoclonal antibodies to the S2 domain of spike proteins of wild-type SARS-CoV-2 and multiple variants. Viruses 2023, 15, 174. [Google Scholar] [CrossRef]
- Soleymani, L.; Li, F. Mechanistic challenges and advantages of biosensor miniaturization into the nanoscale. ACS Sens. 2017, 2, 458–467. [Google Scholar] [CrossRef]
- Domingo, E. Virus as Populations: Composition, Complexity, Dynamics, and Biological Implications, 1st ed.; Academic Press: San Diego, CA, USA, 2015. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
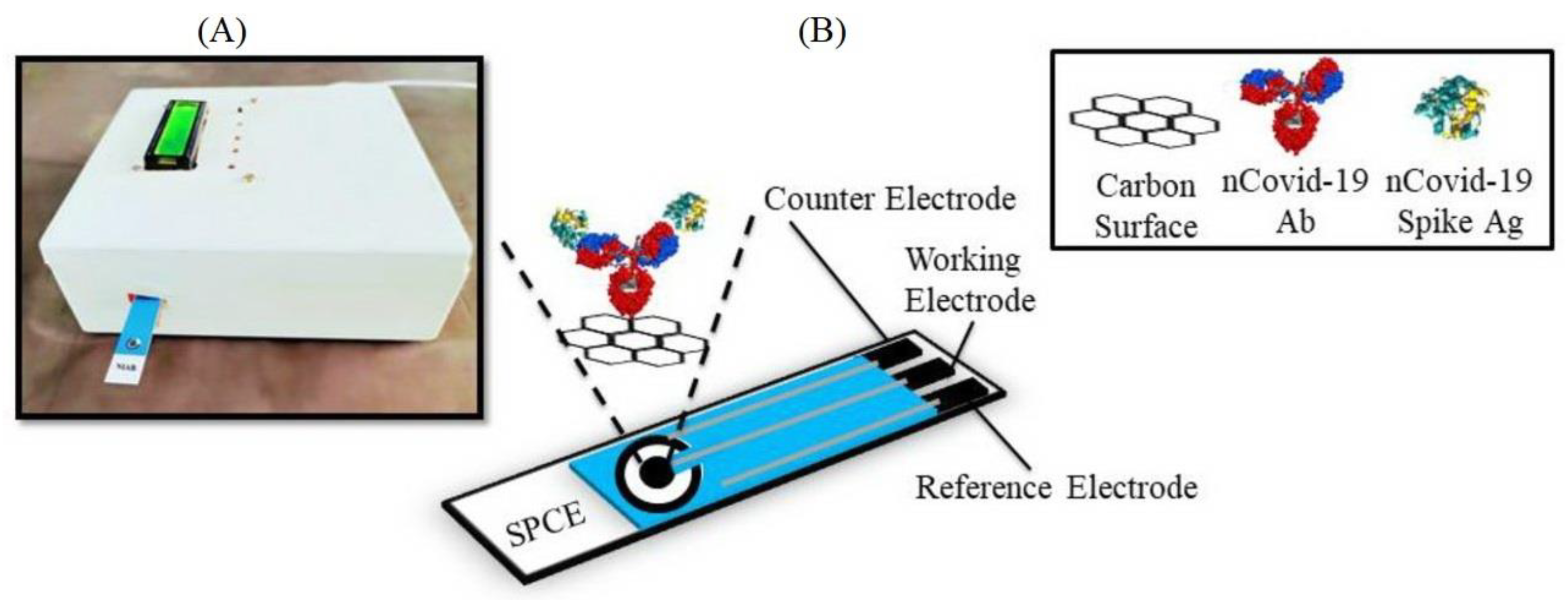{kind=link}
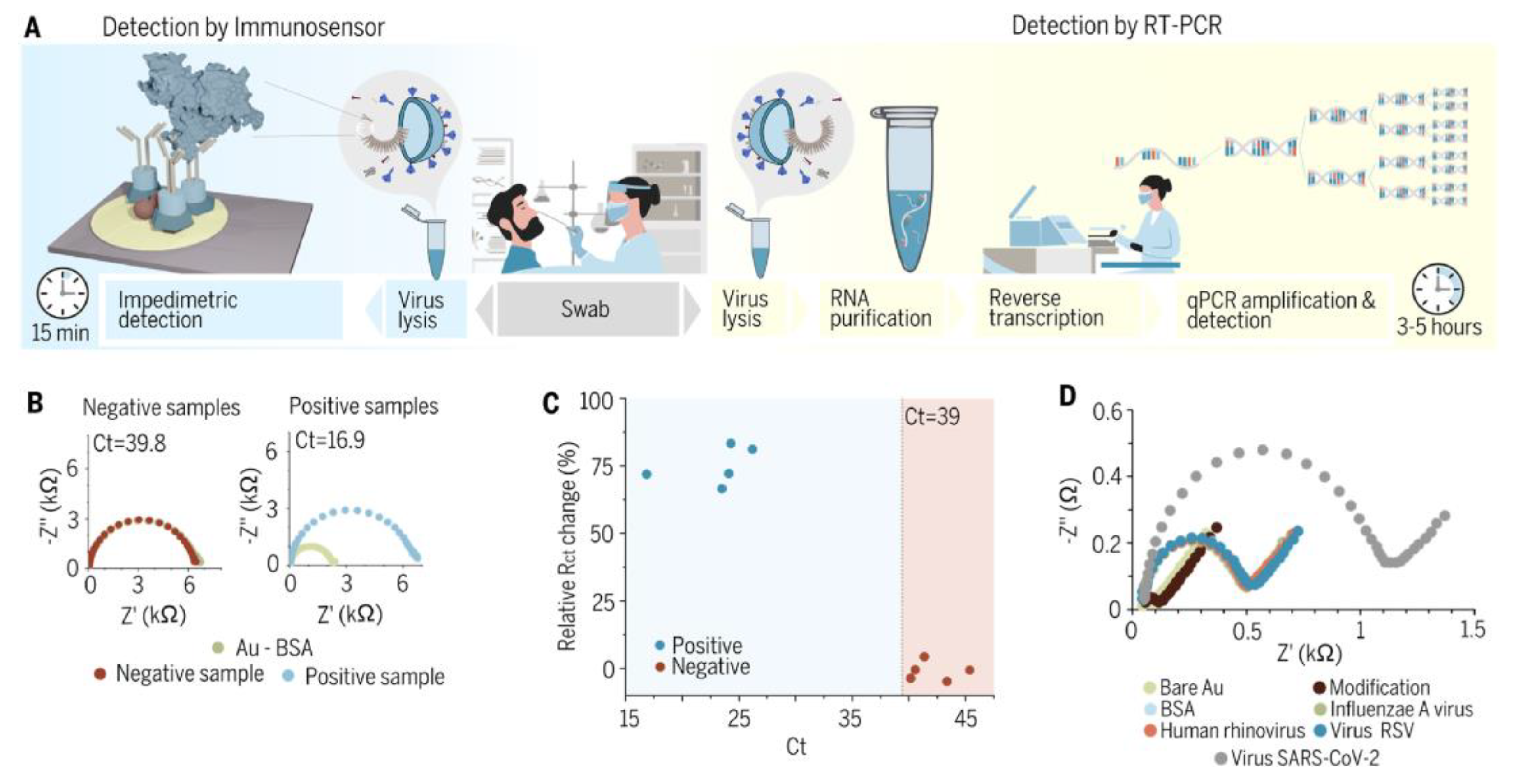{kind=link}
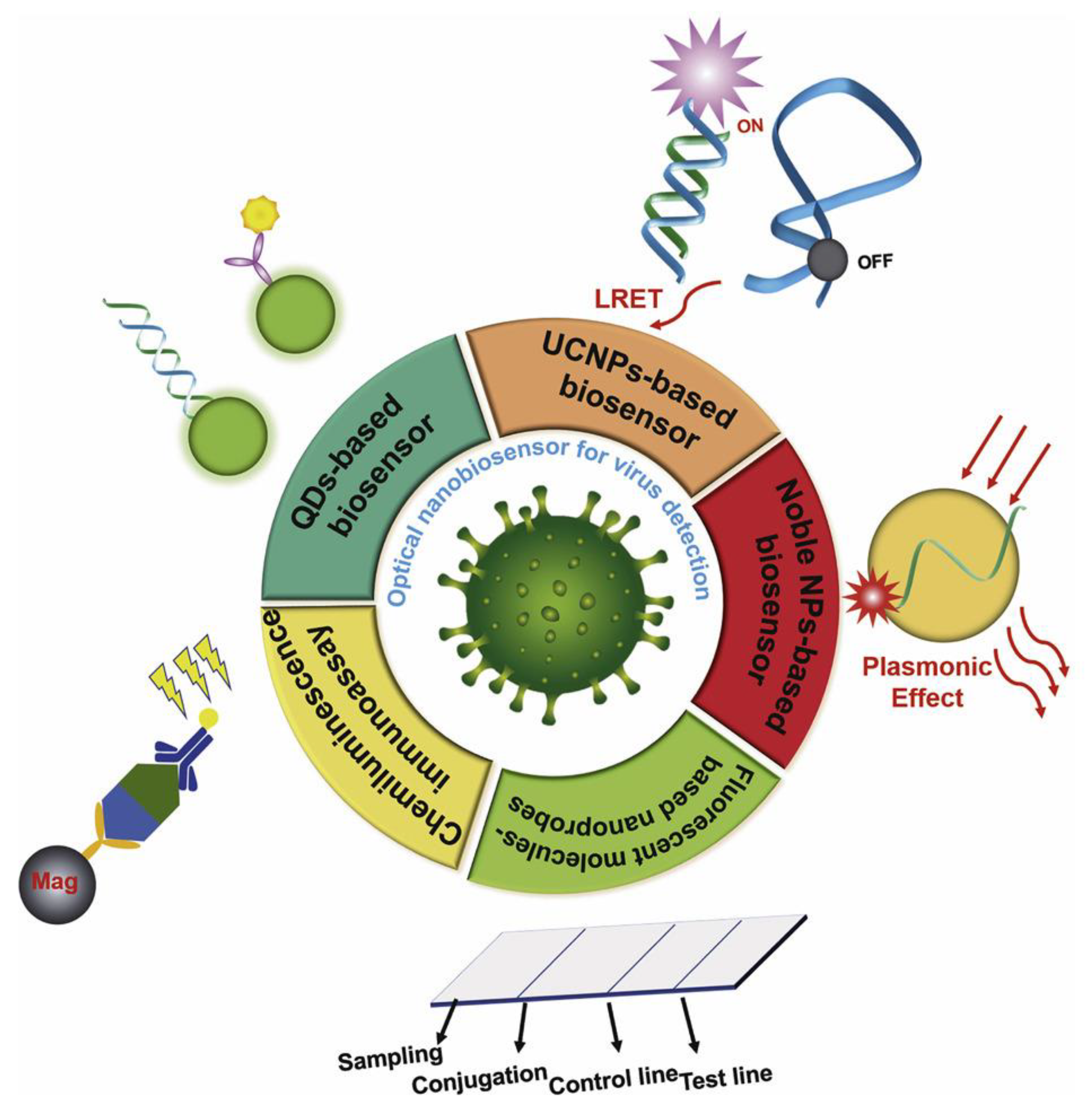{kind=link}
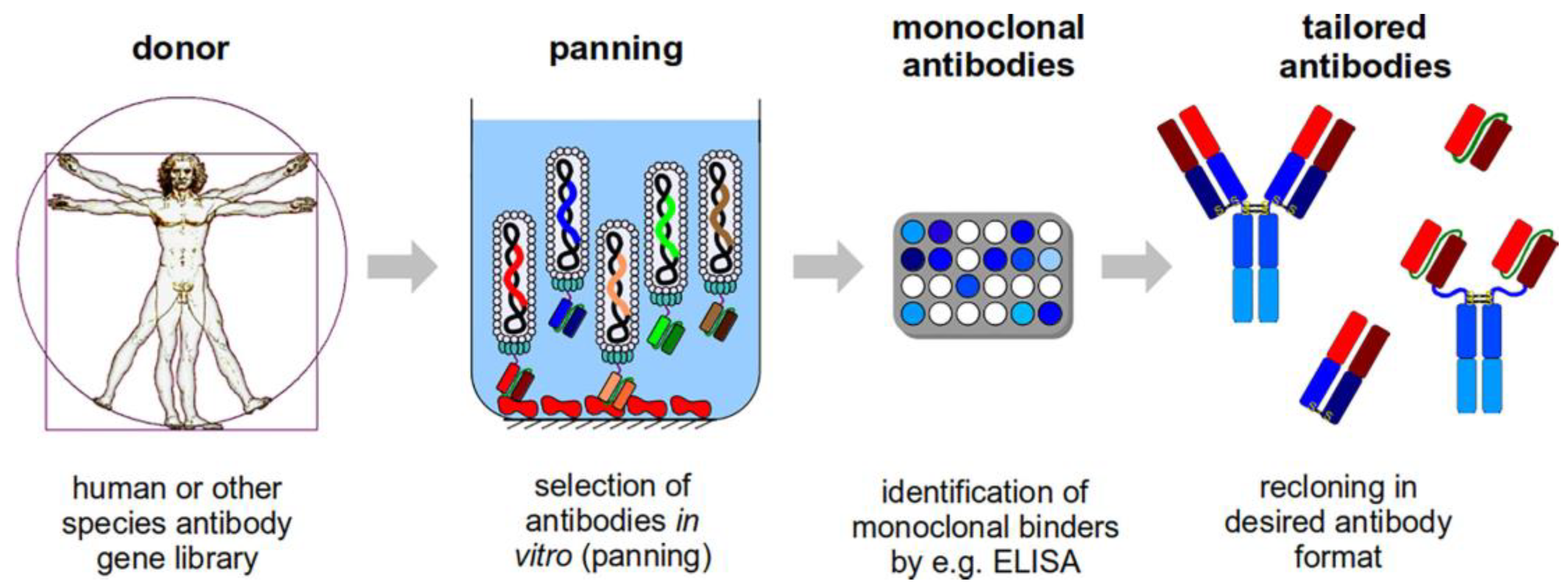{kind=link}
| Technique | Principle | Time | Advantages | Disadvantage |
|---|---|---|---|---|
| PCR | nucleic acid | hours |
|
|
| ELISA | viral protein | hours | highly specific |
|
| Cell culturing | infectivity assay | days to weeks |
|
|
| Electron microscopy | viral particles | hours | rapid method |
|
| Computed tomography | chest scanning | hours | good basis for clinical diagnosis and treatment |
|
| Virus | Phage Ab Target | Type of Phage Library | Format of Phage Abs | Application of Phage Abs | Reference | |
|---|---|---|---|---|---|---|
| Australian bat lyssavirus (ABLV) | glycoprotein G | naïve | Fab, IgG | ELISA, in vitro neutralization | [84] | |
| Avian influenza virus H7N2 (AIV) | complete virus | immune | VHH | ELISA | [85] | |
| Bluetongue virus (BTV) | complete virus | semi-synthetic | scFv, scFv-Fc | ELISA | [86] | |
| Bovine immunodeficiency virus (BIV) | capsid (CA) protein | semi-synthetic | scFv | ELISA, WB | [87] | |
| Bovine viral diarrhea virus (BVDV) | envelope 2 (E2) protein | immune | VHH | ELISA, qRT-PCR, in vitro neutralization | [88] | |
| nonstructural protein 5 (NS5B) | immune | VHH | ELISA | [89] | ||
| Broad bean mottle virus (BBMV) | complete virus | immune | VHH | ELISA, in vitro neutralization | [90] | |
| Canine parvovirus type 2 (CPV2) | complete virus | semi-synthetic | scFv | ELISA | [91] | |
| virus-like particles (VLP) | immune | scFv | ELISA, ICA, virus suppression assay | [92] | ||
| Chikungunya virus (CHIKV) | VLP | immune | VHH | ELISA, MagPlex, in vitro neutralization | [93] | |
| Cucumber mosaic virus (CMV) | complete virus | semi-synthetic | scFv | ELISA, WB | [94] | |
| Dengue virus (DENV) | non-structural protein 3 (NS3) | naïve | Fab | ELISA, in vitro neutralization | [95] | |
| nonstructural protein 1 (NS1) | immune | VHH | MagPlex assay | [96] | ||
| RNA-dependent RNA polymerase (RdRp) in the viral non-structural protein 5 (NS5) | naïve | scFv | ELISA, WB, in vitro inhibition | [97] | ||
| Duck hepatitis A virus (DHAV) | VP3 protein | immune | scFv | ELISA, in vivo neutralization | [98] | |
| VP1 protein | immune | VHH | ELISA, IF, dot-blot | [99] | ||
| Ebola virus | nucleoprotein | synthetic | scFv, IgG | ELISA, WB | [100] | |
| viable Zaire ebolavirus, viral matrix protein VP40, viral nucleoprotein (NP) | immune | scFv, IgNAR V | ELISA, WB | [101] | ||
| multifunctional viral protein (VP35) | semi-synthetic | scFv | ELISA, WB | [102] | ||
| glycoprotein (GP) | immune | scFv, scFv-Fc | ELISA, in vitro neutralization, in vivo protection | [103,104] | ||
| Ectromelia virus (ECTV) | epitope p35 | immune | scFv | ELISA, in vitro neutralization, WB | [105] | |
| Enterovirus 71 (EV71) | inactivated EV71 virions | immune | Fab | ELISA, WB, in vitro neutralization | [106] | |
| virion protein 2 (VP2) | naïve | scFv | ELISA, WB | [107] | ||
| internal capsid protein (VP4) | naïve | scFv | ELISA, WB, IF, in vitro neutralization | [108] | ||
| Epstein–Barr virus (EBV) | latent membrane protein 1 (LMP1) | naïve | Fab | ELISA, WB, IF, FACS, in vitro inhibition | [109] | |
| Foot-and-mouth disease virus (FMDV) | recombinant non-structural protein (NSP) 3ABC | immune | scFv | ELISA, WB | [110] | |
| VLP | immune | VHH | ELISA, IF | [111] | ||
| intact (146S) FMDV | immune | VHH | ELISA | [112] | ||
| VP2 capsid protein | immune | scFv | ELISA | [113] | ||
| Grapevine leafroll-associated virus 3 (GLRaV-3) | coat protein | immune | scFv | ELISA | [114] | |
| Grapevine virus B (GVB) | virus particles | semi-synthetic | scFv | ELISA | [115] | |
| Hantaviruses | virus particles | immune | Fab | ELISA, IF, WB | [116] | |
| nucleoprotein | immune | VHH | ELISA, WB | [117] | ||
| Hendra virus (NiV) and Nipah virus (HeV) | attachment envelope glycoprotein G | naïve | Fab, IgG | ELISA, immunoprecipitation, WB, in vitro neutralization, IF, in vivo neutralization | [118] | |
| Hepatitis A (HAV) | HAV capsid | immune | Fab, IgG | ELISA, in vitro neutralization | [119] | |
| Hepatitis B virus (HBV) | hepatitis B virus surface antigen (HBsAg) | immune | Fab, scFv | ELISA, IF | [120] | |
| Hepatitis C virus (HCV) | core protein | immune | Fab | ELISA | [121] | |
| E2 glycoprotein | immune | Fab | ELISA, in vitro neutralization | [122] | ||
| non-structural protein-3/4A (NS3/4A) | naïve | scFv | ELISA, IF, in vitro neutralization | [123] | ||
| non-structural protein-5A (NS5A) | naïve | scFv | ELISA, WB, IF, in vitro neutralization | [124] | ||
| Hepatitis E virus (HEV) | ORF2 protein | immune | Fab | ELISA, WB, in vitro neutralization | [125] | |
| Herpes simplex virus (HSV-1, HSV-2) | glycoproteins gD and gB | immune | Fab | ELISA, immuno precipitation, WB, in vitro neutralization | [126] | |
| Human cytomegalovirus (HCMV) | gycoprotein B (gB) and H (gH) | immune | scFv | ELISA, in vitro neutralization | [127] | |
| glycoprotein 55 (gp55) | immune | scFv | ELISA, in vitro neutralization | [128] | ||
| Human immunodeficiency viruses (HIV) | transmembrane glycoprotein gp41 | synthetic | Fab | WB, in vitro neutralization | [129] | |
| integrase (IN) protein | immune | scFv | ELISA, WB, IF, in vitro neutralization | [130] | ||
| p24 | immune | scFv | ELISA | [131] | ||
| envelope glycoprotein gp140 | immune | VHH | ELISA, in vitro neutralization | [132] | ||
| CD4bs region of subtype C | immune | scFv | ELISA, in vitro neutralization | [133] | ||
| Envelope glycoprotein gp120 | immune | Fab | ELISA, in vitro neutralization, IF | [134] | ||
| Human metapneumovirus (HMPV) | F protein | immune | Fab | ELISA, IF, in vitro neutralization, in vivo protection | [135] | |
| Influenza A | H1N1 | hemagglutinin protein (HA) | immune | scFv | ELISA | [136] |
| H2N2 | hemagglutinin protein HA (stem region) | immune | Fab | ELISA, in vitro neutralization | [137] | |
| H3N2 | HA protein and its variants | semi-synthetic | scFv | ELISA | [138] | |
| non-structural protein-1 (NS1) | naïve | scFv | ELISA, WB, in vitro neutralization, IF | [139] | ||
| nucleoprotein (NP) | immune | scFv | ELISA, WB, in vitro inhibition | [140] | ||
| hemagglutinin protein (HA) | semi-synthetic | scFv | ELISA | [141] | ||
| complete inactivated virus | immune | VHH | ELISA | [142] | ||
| Influenza A | M2 protein (cytoplasmatic domain) | naïve | scFv, scFv-Fc | WB, IHC | [143] | |
| Influenza B | whole virus | immune | Fab | ELISA, WB, IF, in vitro neutralization | [144] | |
| hemagglutinin protein (HA) | immune | VHH | ELISA | [145] | ||
| Japanese encephalitis virus (JEV) | purified virion | immune | Fab | ELISA, immunoprecipitation, in vitro neutralization | [146] | |
| domains I, II, III of envelope protein | immune | Fab, IgG | ELISA, WB, immunoprecipitation, in vitro neutralization, in vivo protection | [67] | ||
| Marburg virus | nucleoprotein | immune | sdAbs | ELISA | [147] | |
| transmembrane glycoprotein (GP) | immune | scFv, scFv-Fc | WB, in vitro neutralization, in vivo protection | [148] | ||
| VP35 protein | synthetic | Fab | ELISA | [149] | ||
| Middle East respiratory syndrome-related coronavirus (MERS-CoV) | S2 subunit of the MERS-CoV spike protein (MERS-S2P) | synthetic | Fab, IgG | ELISA, IF, in vitro neutralization, ACCEL ELISA™ | [150] | |
| nucleoprotein (NP) | naïve | scFv | ELISA | [151] | ||
| Norovirus | VLP | semi-synthetic | scFv | ELISA, WB | [152] | |
| VLP with major capsid protein VP1 and a minor structural protein VP2 | immune | VHH | ELISA, WB, in vitro inhibition, IF | [153] | ||
| P-domain of the GI.1 (VLP) major capsid protein | semi-synthetic | scFv | ELISA, dot-blot, surface plasmon resonance (SPR) | [154] | ||
| Paramyxovirus | glycoproteins F (fusion protein) and HN (attachment protein) | synthetic | Fab, sAb | ELISA, in vitro neutralization, immunoprecipitation | [155] | |
| Plum pox virus (PPV) | NIa protease | semi-synthetic | scFv | WB, dot-blot | [156] | |
| Poliovirus | capsid proteins VP1 and VP3 | immune | Fab, IgG | ELISA, in vitro neutralization, in vivo protection | [157] | |
| Polyomavirus | major capsid viral protein 1 (VP1) | synthetic | Fab, IgG | ELISA | [158] | |
| Porcine circovirus type-2 (PCV2) | complete virus | immune | sdAbs | Western blot, ELISA, and SPR | [159] | |
| cap protein | immune | VHH | ELISA | [160] | ||
| Porcine epidemic diarrhea virus (PEDV) | membrane protein of PEDV | immune | sdAb fragments (sdAb-Mc19/29/30/37) | ELISA | [161] | |
| S1 domain of spike protein | immune | VHH | ELISA, in vitro neutralization | [162] | ||
| nucleocapsid (N) protein | immune | VHH | ELISA | [163] | ||
| S1 region of the spike protein | immune | scFv | ELISA, IF, in vivo protection | [164] | ||
| Porcine reproductive and respiratory syndrome virus (PRRSV) | non-structural protein 9 (Nsp9) | immune | VHH | ELISA, IF, immunoprecipitation | [165] | |
| non-structural protein 4 (Nsp4) | immune | VHH | ELISA, immunoprecipitation | [166] | ||
| Rabies lyssavirus | glycoprotein (antigenic site II) | immune | Fab, IgG | ELISA, IF, in vitro neutralization | [167] | |
| glycoprotein G | naïve | VHH | ELISA, in vitro neutralization, in vivo protection | [168] | ||
| Respiratory syncytial virus (RSV) | glycoprotein F | synthetic | Fab, IgG | ELISA, in vitro neutralization | [169] | |
| Rotavirus | non-structural protein Nsp4 | semi-synthetic | scFv | ELISA, WB | [170] | |
| VP8* fraction of rotaviral VP4 outer capsid | semi-synthetic | scFv | ELISA, WB, in vitro inhibition | [171] | ||
| Severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) | S protein (RBD) | synthetic | VH | ELISA, in vitro neutralization | [172] | |
| immune | scFv, scFv-Fc, IgG | ELISA, in vitro inhibition, in vitro neutralization, in vivo protection | [173] | |||
| naïve | scFv | ELISA, in vitro neutralization, in vitro inhibition, in vivo protection | [174] | |||
| synthetic | Fab, biospecific Fab+VH | in vitro neutralization | [175] | |||
| immune | VHH, VHH-Fc | ELISA, in vitro neutralization | [176] | |||
| RBD | immune | VHH | ELISA, in vitro inhibition, in vitro neutralization | [177] | ||
| semi-synthetic | Fab | ELISA, in vitro neutralization | [178] | |||
| immune | Fab, IgG | ELISA, in vitro neutralization assay, in vivo protection | [179] | |||
| nucleocapsid protein (NP) | immune | scFv, scFv-Fc | ELISA, WB, dot-blot, lateral flow strip assay | [180] | ||
| Sin Nombre orthohantavirus (SNV) | Sin Nombre Virus nucleocapsid protein (SNV-N) | naïve | scFv | ELISA, WB, dot-blot | [78] | |
| Simian immunodeficiency virus (SIV) | trimeric (gp140) and monomeric (gp120) forms of the SIVmac239 envelope glycoprotein | immune | scFv, scFv-Fc | ELISA, WB, in vitro neutralization | [181] | |
| Swine influenza virus (SIV) | SIV nucleoprotein (SIV-NP) | immune | VHH | ELISA, WB | [182] | |
| Transmissible gastroenteritis virus (TGEV) | whole virus | immune | scFv | ELISA, in vitro neutralization, IF, WB | [183] | |
| Usutu virus (USUV) | domain III (DIII) of the USUV E protein | immune | scFv | ELISA, WB, in vitro neutralization | [184] | |
| Vaccinia virus (VACV) | virus particles | immune | scFv, IgG | inhibition ELISA, in vitro neutralization | [185] | |
| Venezuelan equine encephalitis viruses (VEEV) | viral E1 envelope protein | immune | scFv, scFv-Fc | ELISA, WB, in vitro neutralization, in vivo protection | [66] | |
| White spot syndrome virus (WSSV) | virus particles | immune | scFv | ELISA, in vitro neutralization | [186] | |
| West Nile virus (WNV) | domain IIII of WNV envelope (E) protein | immune | Fab | ELISA, IF, in vitro neutralization, in vivo protection | [187] | |
| envelope (E)protein | naïve | scFv, scFv-Fc | ELISA, WB, in vitro neutralization | [188] | ||
| Western equine encephalitis viruses (WEEV) | virus | immune | scFv | Enzyme-linked immunosorbent assays (ELISAs) | [189] | |
| virus particles | immune | scFv, scFv-Fc | in vitro neutralization, in vivo protection | [190] | ||
| E2/E3E2 envelope proteins | immune | VHH | ELISA (MagPlex assay) | [191] | ||
| Yellow fever virus (YFV) | domain II of envelope protein | immune | scFv, IgG | in vitro neutralization, in vivo protection | [192] | |
| Zika virus (ZIKV) | envelope (E) protein | immune | scFv | ELISA, WB, FACS, in vitro inhibition | [193] | |
| nonstructural protein 1 (NS1) | immune | VHH | ELISA | [194] | ||
| Type of Sensory System | Phage Antibody Format | Virus | Target | Detection Limit | Analysis Time | Reference |
|---|---|---|---|---|---|---|
| Biosensor based on the lateral flow immunoassay (LFIA) | scFv–Fc | SARS-CoV-2 | nucleocapsid protein (NP) | 2 ng of antigenic protein 2.5 × 104 pfu cultured virus | 20 min | [180] |
| Sandwiched phage-based enzyme-linked chemiluminescence immunoassay (ELCLIA) | n.i. | SARS-CoV-2 | S1 domain of the spike glycoprotein | 78 pg/mL of antigenic protein | n.i. | [195] |
| Quench-based fluorescent immunosensor (Quenchbodies) | Fab, Ig | SARS-CoV-2 | S1 domain of the spike glycoprotein | 0.1 nM of S protein trimer 105 copies/mL of virus | 2 min | [196] |
| Nanobody-functionalized nanoparticles for rapid, electronic detection (Nano2RED) | n.i. | SARS-CoV-2 | spike protein receptor binding domain (RBD) | 40 pg/mL 1.3 pM | 5–20 min | [197] |
| Ebola virus | secreted glycoprotein (sGP) | 10 pg/mL 0.13 pM | ||||
| Electro-acoustic sensor | Ig | bacteriophage FAl-SR65 | whole virus particles | 106 phages/mL | 5 min | [198] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guliy, O.I.; Evstigneeva, S.S.; Khanadeev, V.A.; Dykman, L.A. Antibody Phage Display Technology for Sensor-Based Virus Detection: Current Status and Future Prospects. Biosensors 2023, 13, 640. https://doi.org/10.3390/bios13060640
Guliy OI, Evstigneeva SS, Khanadeev VA, Dykman LA. Antibody Phage Display Technology for Sensor-Based Virus Detection: Current Status and Future Prospects. Biosensors. 2023; 13(6):640. https://doi.org/10.3390/bios13060640
Chicago/Turabian StyleGuliy, Olga I., Stella S. Evstigneeva, Vitaly A. Khanadeev, and Lev A. Dykman. 2023. "Antibody Phage Display Technology for Sensor-Based Virus Detection: Current Status and Future Prospects" Biosensors 13, no. 6: 640. https://doi.org/10.3390/bios13060640