
Global Properties of Cytokine-Enhanced HIV-1 Dynamics Model with Adaptive Immunity and Distributed Delays

 , , ,
, , ,
Abstract
:1. Introduction
2. Model Development
3. Biologically Realistic Domain
4. Equilibria
- (I)
- Uninfected equilibrium, , where .
- (II)
- Chronic infection equilibrium with inactive immune response , wherewhere is the basic reproduction number defined as:It follows that exists if , and obviously, represents the contribution of viral infections to , whereas represents the contribution of inflammatory cytokines to .
- (III)
- Chronic infection equilibrium with only CTL immunity , wherewhereThe ratio is the CTL immunity activation number. Then, the equilibrium point exists when . The CTL-mediated immune response is triggered or not depending on the value of the parameter .Let us consider the case when . Then, from Equations (16)–(20), we obtain two equilibria.
- (IV)
- Chronic infection equilibrium with only humoral immunity , whereand satisfies the following equation:whereSince and then , and the equation has two different real roots. The positive root isIt follows that, if , then and . Define the humoral immunity activation number as:Thus, . The humoral immune response is triggered or not based on the parameter . Hence, exists when .
- (V)
- Chronic infection equilibrium with both CTL and humoral immunities, , wherewhere and represent the humoral immunity competitive number and CTL immunity competitive number, respectively, and they are given as follows:
5. Global Stability
6. Comparison Results
- (i)
- Reverse transcriptase inhibitor (RTI), which prevents the virus from infecting the cell [11];
- (ii)
7. Numerical Simulations
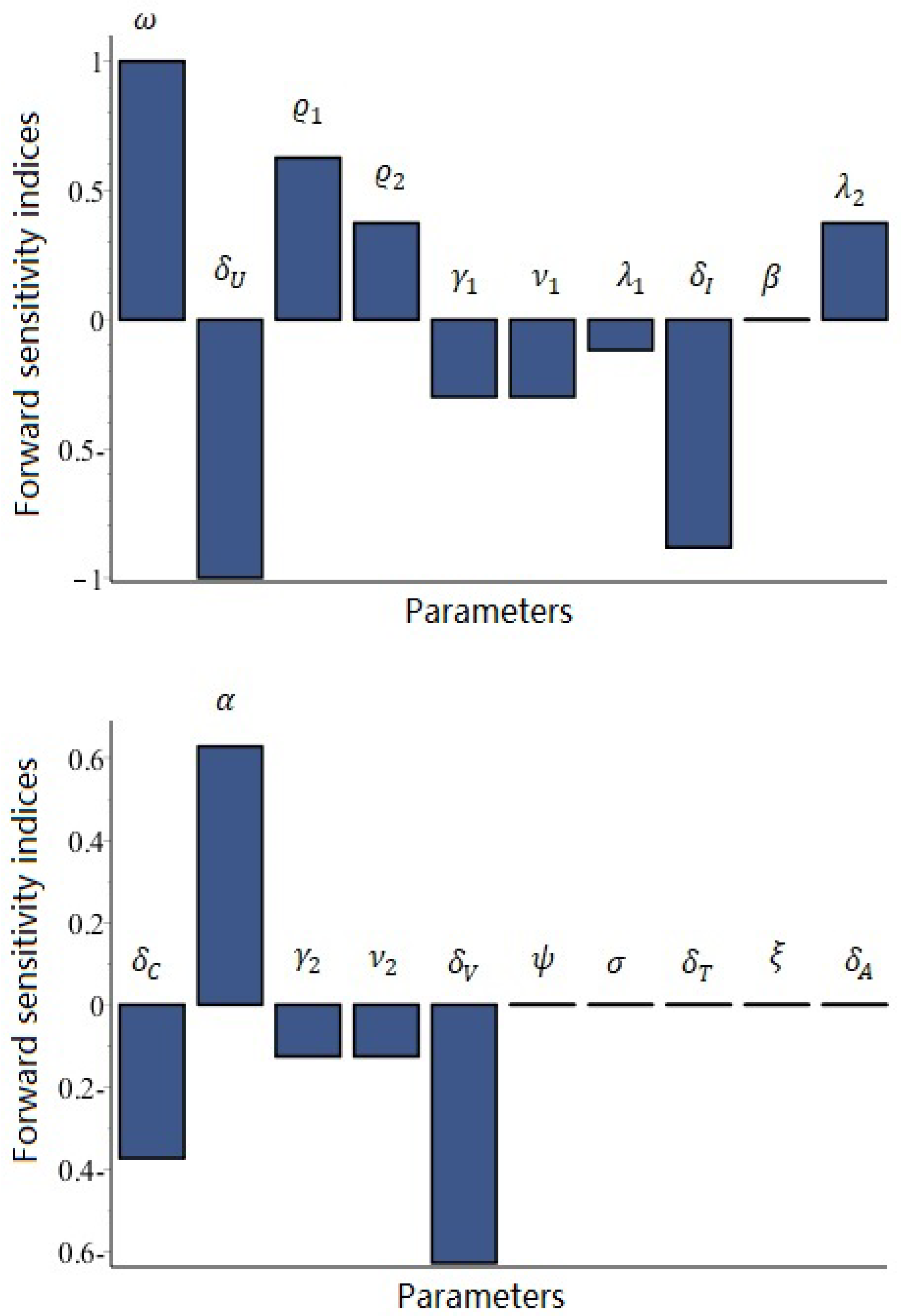
7.1. Sensitivity Analysis of to the Parameters for Model (62)–(67)
- (i)
- The parameters with positive sensitivity indices include , and , withThis implies that any increase or decrease in the values of those parameters directly influences , leading to either an increase or a decrease in its value.
- (ii)
- The parameters with negative sensitivity indices, signifying that an increase in their values leads to a decrease in , include and , as delineated below:
- (iii)
- The parameters and have no impact on the value of .
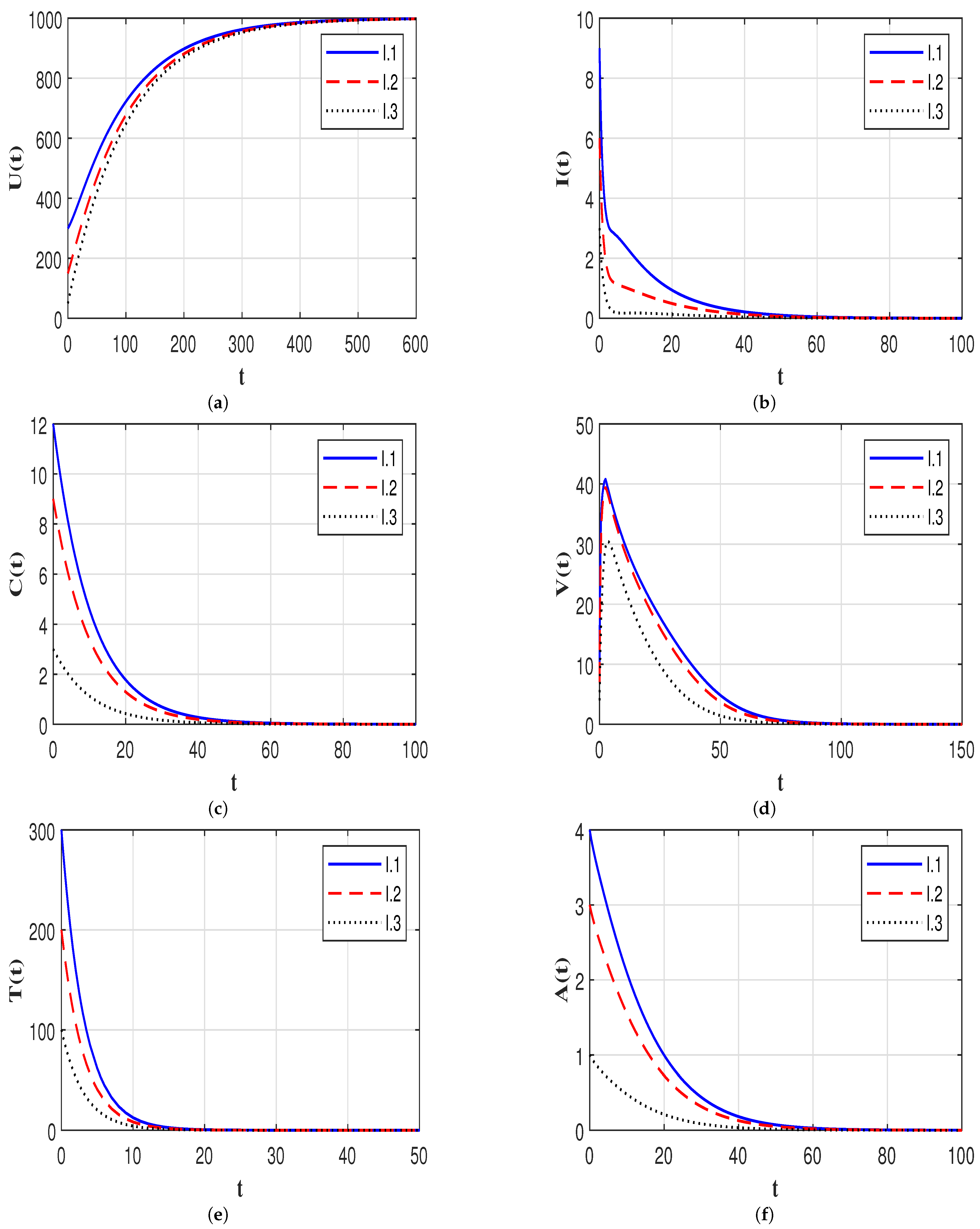
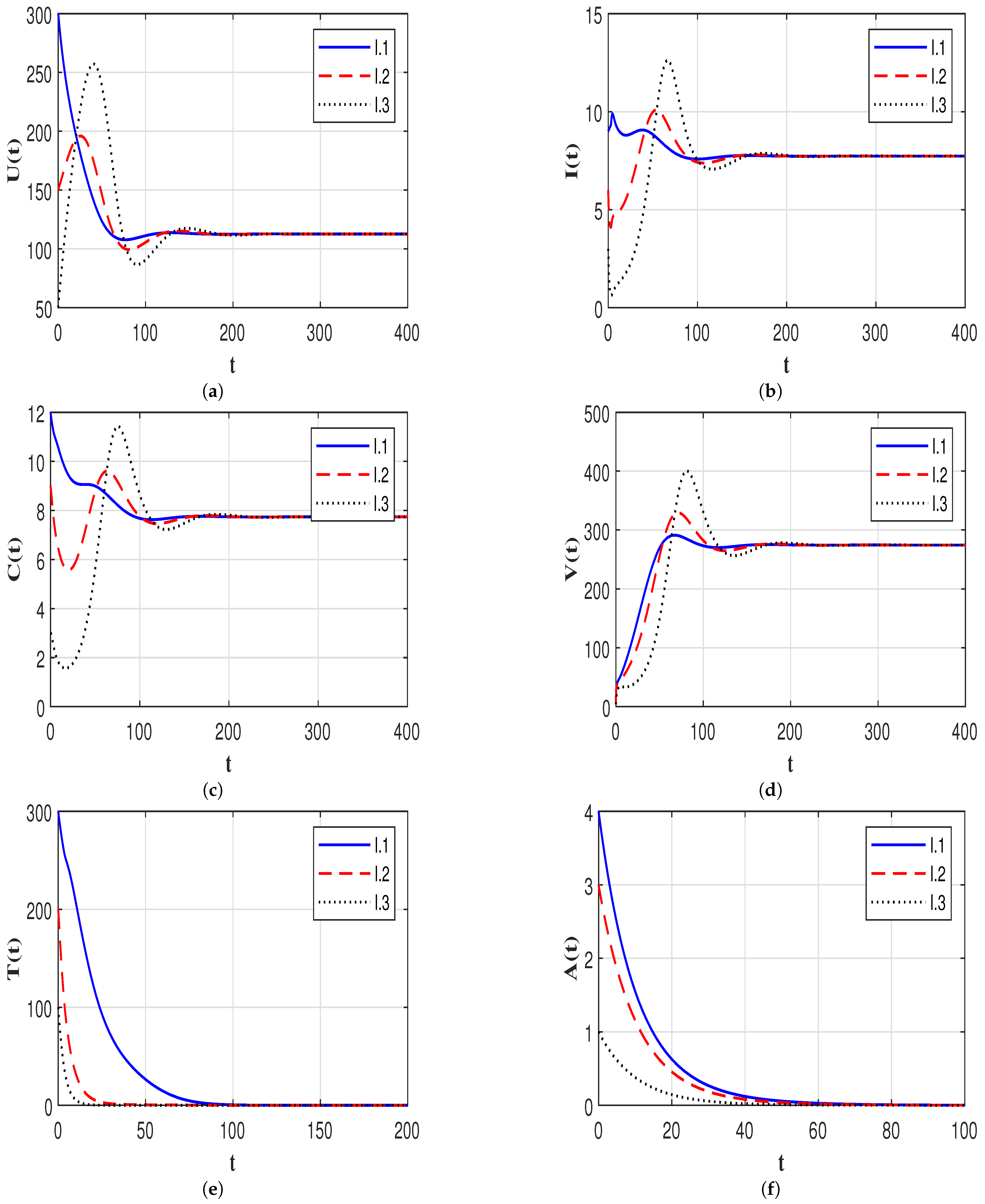
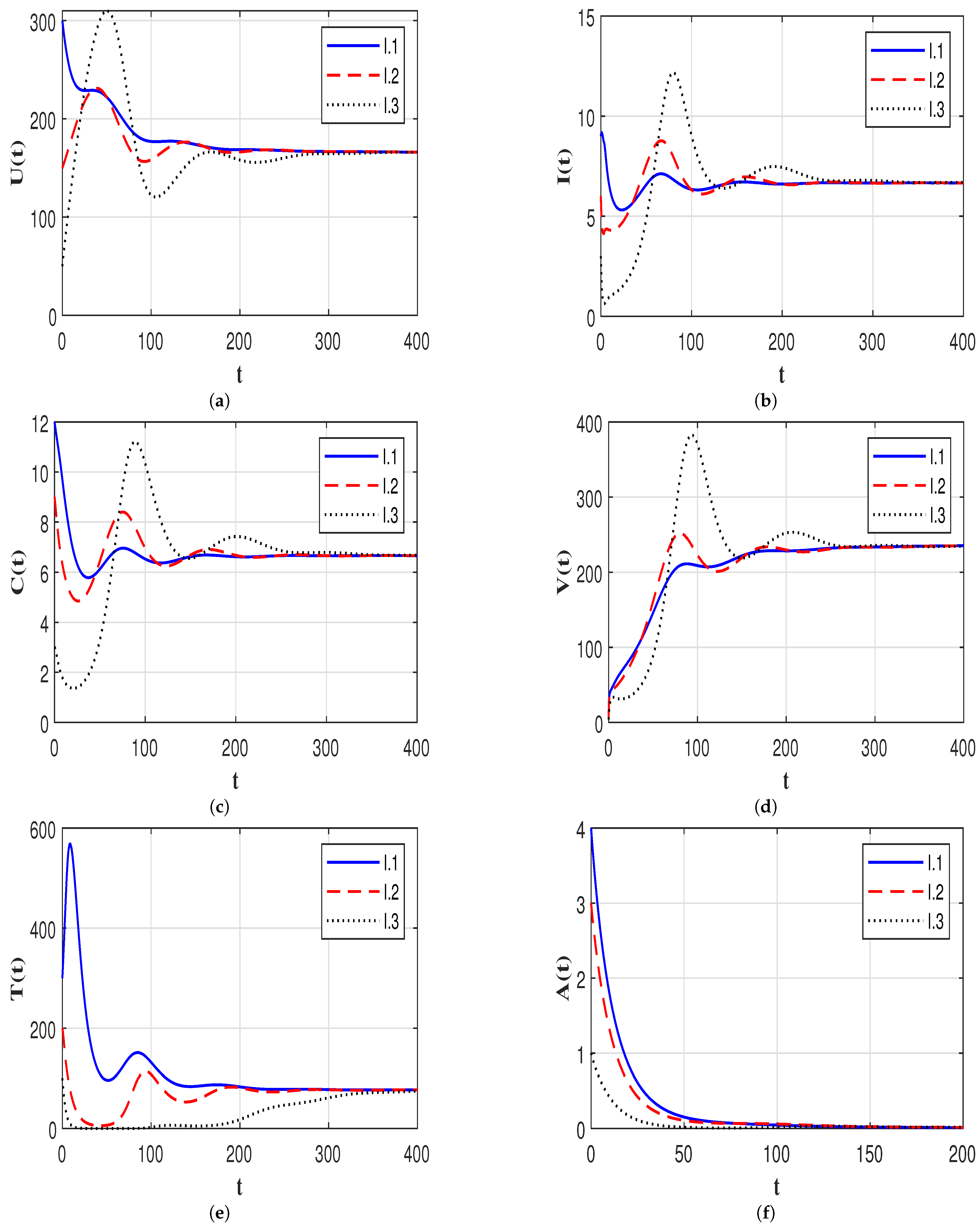
7.2. Stability of the Equilibria
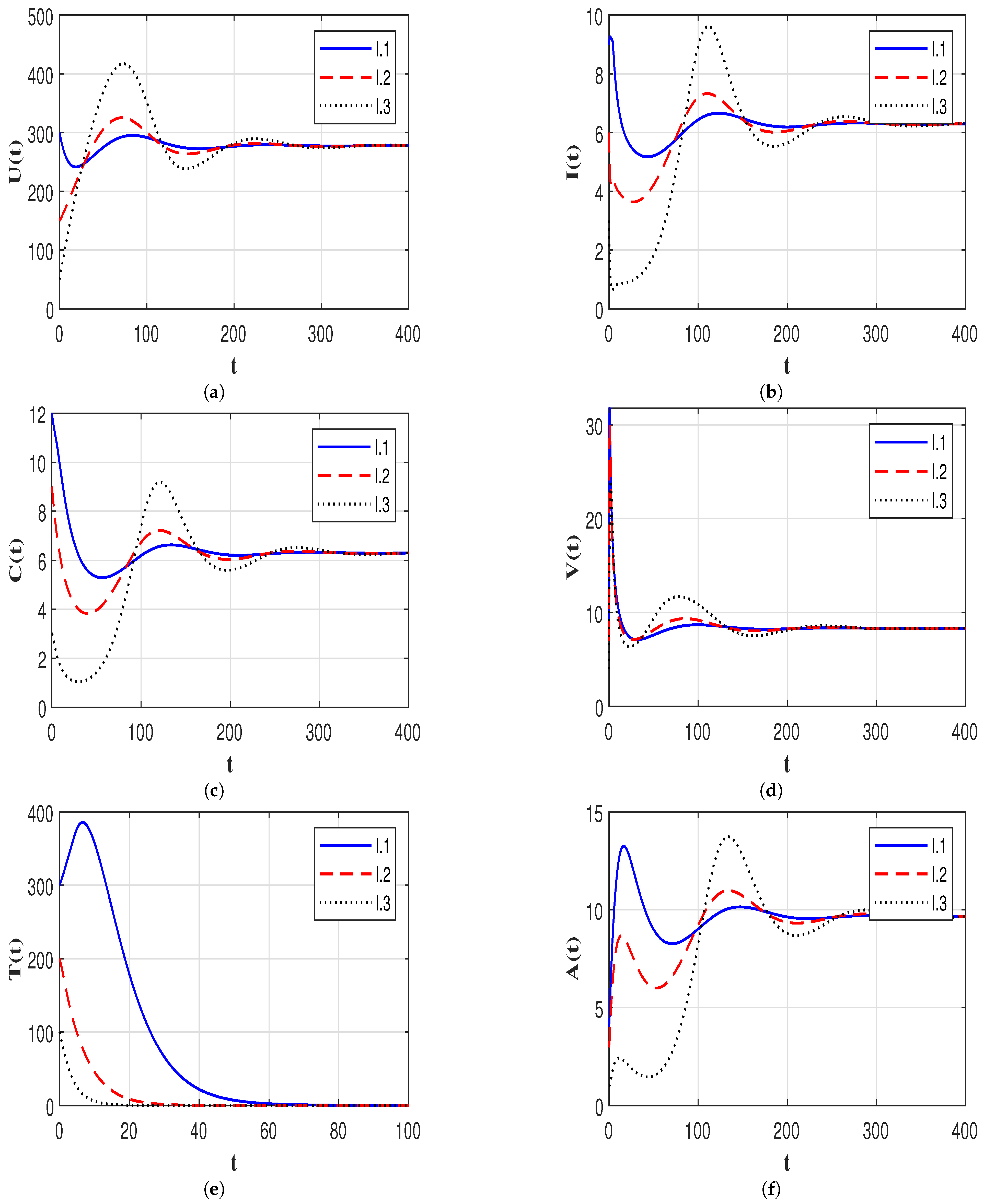
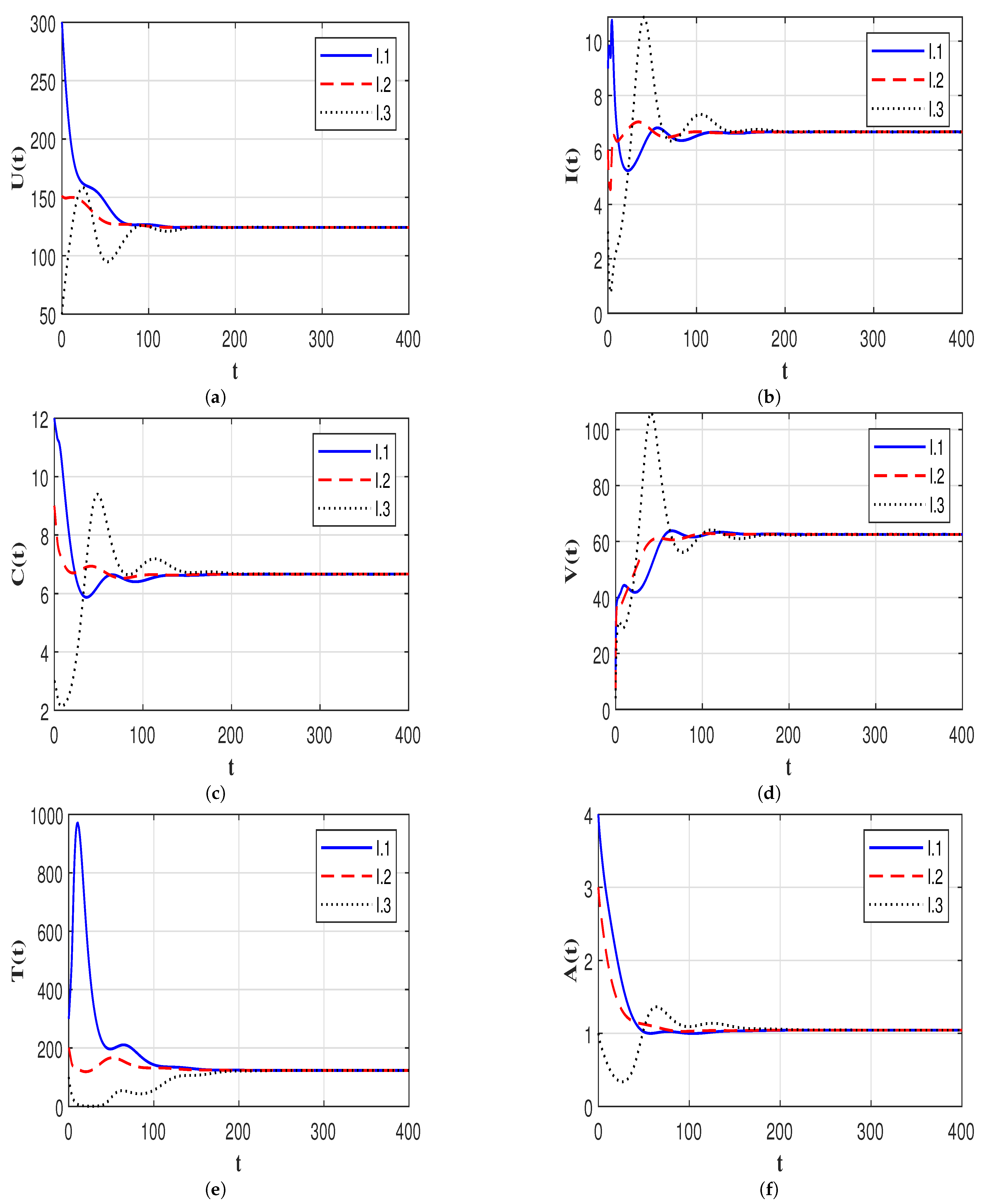
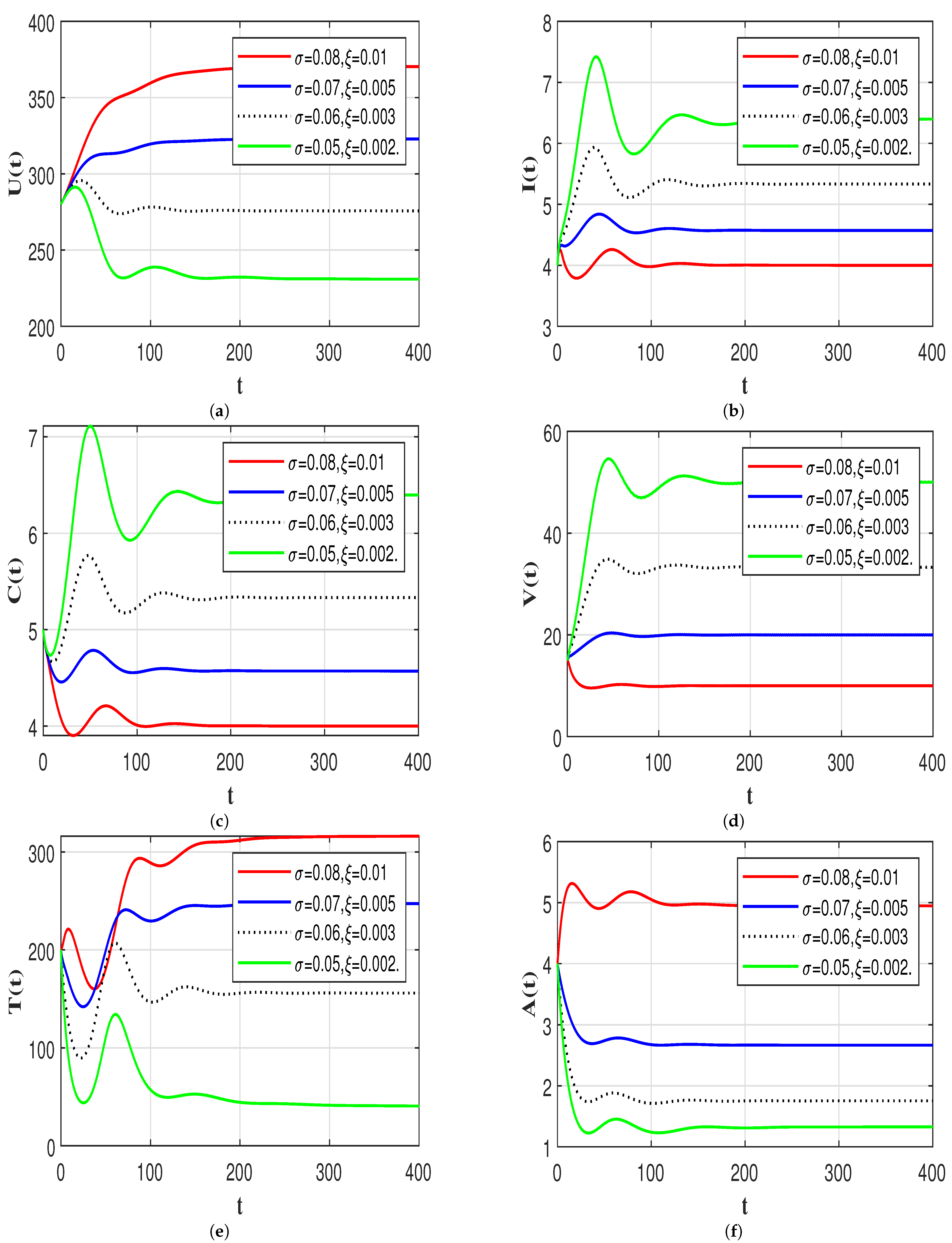
7.3. Effect of Time Delays on the HIV-1 Dynamics
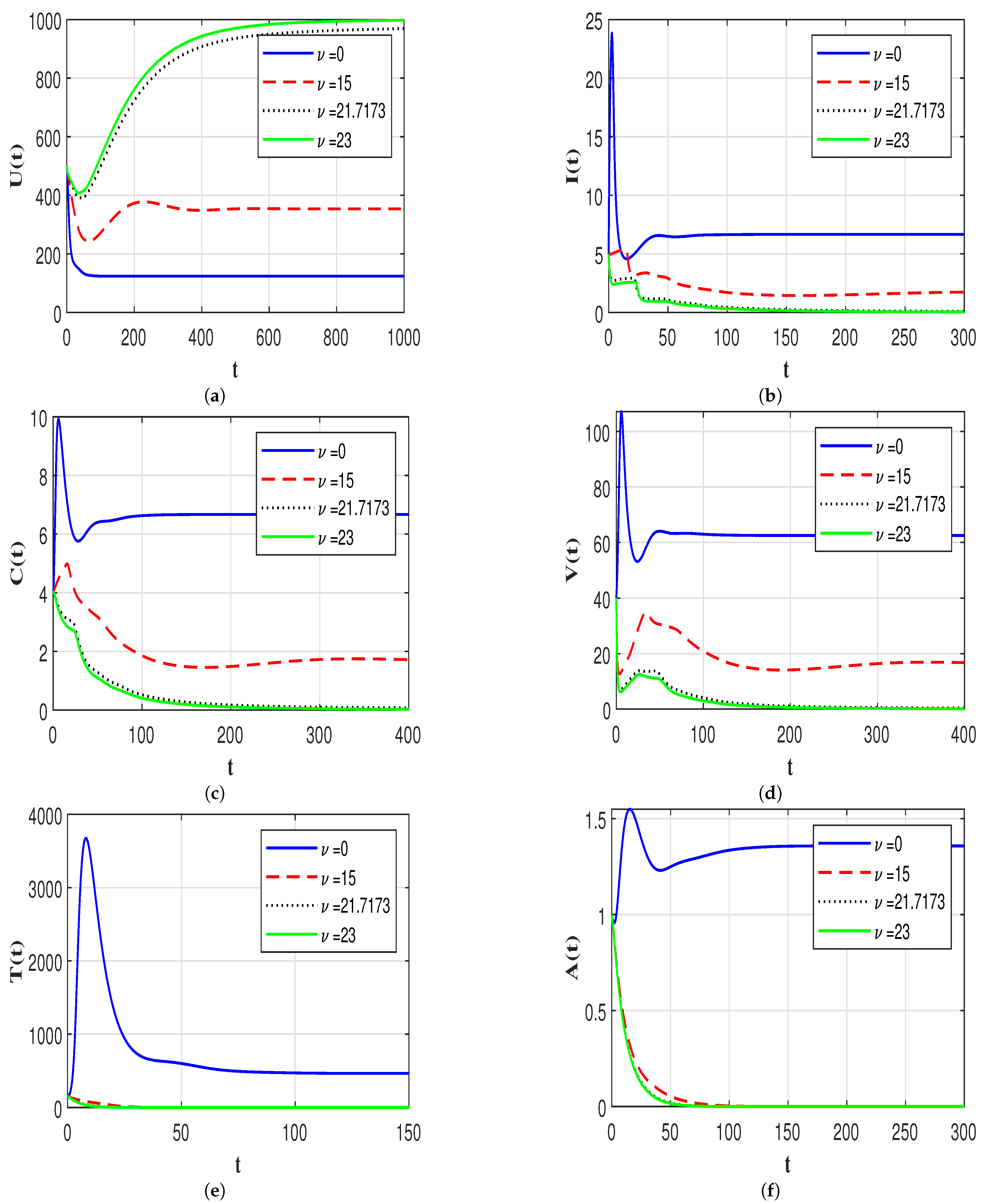
7.4. Effect of Immune Response on the HIV-1 Dynamics
8. Discussion
- The uninfected equilibrium, , usually exists, and it is G.A.S. when . In this state, the number of HIV-1 particles eventually converges to 0. Different control plans can be applied to makeThese plans are, for example:
- The chronic infection equilibrium with inactive immune response, , exists when . Moreover, is G.A.S. when , , and . In this situation, HIV-1 is present, but without any immune response. This can happen when the populations of both HIV-1 and infected cells are insufficient to activate the immune system’s reaction, i.e., and .
- The chronic infection equilibrium with only CTL immunity, , exists when . Further, is G.A.S. when and . In this case, HIV-1 exists in the body under CTL immune response only. This can happen when the number of viruses in the body becomes small and insufficient to activate the humoral immune response, i.e., .
- The chronic infection equilibrium with only humoral immunity, , exists when . Further, is G.A.S. when and . In this case, HIV-1 exists in the body under humoral immune response only. This can happen when the number of infected cells becomes small and insufficient to activate the CTL immune response, i.e., .
- The chronic infection equilibrium with both CTL and humoral immunities, , exists and is G.A.S. when and . In this case, HIV-1 infection is chronic, where both humoral and CTL immune responses are activated.
9. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- WHO. The Global Health Observatory. Available online: https://www.who.int/data/gho/data/themes/hiv-aids (accessed on 1 October 2023).
- Wang, S.; Hottz, P.; Schechter, M.; Rong, L. Modeling the slow CD4+T cell decline in HIV-infected individuals. Plos Comput. Biol. 2015, 11, e1004665. [Google Scholar] [CrossRef] [PubMed]
- Nowak, M.A.; Bangham, C.R.M. Population Dynamics of Immune Responses to Persistent Viruses. Science 1996, 272, 74–79. [Google Scholar] [CrossRef]
- Wodarz, D.; May, R.M.; Nowak, M.A. The role of antigen-independent persistence of memory cytotoxic T lymphocytes. Int. Immunol. 2000, 12, 467–477. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Zou, D. Global stability of in host viral models with humoral immunity and intracellular delays. Appl. Math. Model. 2012, 36, 1313–1322. [Google Scholar] [CrossRef]
- Tang, S.; Teng, Z.; Miao, H. Global dynamics of a reaction–diffusion virus infection model with humoral immunity and nonlinear incidence. Comput. Math. Appl. 2019, 78, 786–806. [Google Scholar] [CrossRef]
- Zheng, T.; Luo, Y.; Teng, Z. Spatial dynamics of a viral infection model with immune response and nonlinear incidence. Z. Angew. Math. Phys. 2023, 74, 124. [Google Scholar] [CrossRef]
- Kajiwara, T.; Sasaki, T.; Otani, Y. Global stability for an age-structured multistrain virus dynamics model with humoral immunity. J. Appl. Math. Comput. 2020, 62, 239–279. [Google Scholar] [CrossRef]
- Dhar, M.; Samaddar, S.; Bhattacharya, P. Modeling the effect of non-cytolytic immune response on viral infection dynamics in the presence of humoral immunity. Nonlinear Dyn. 2019, 98, 637–655. [Google Scholar] [CrossRef]
- Murase, A.; Sasaki, T.; Kajiwara, T. Stability analysis of pathogen-immune interaction dynamics. J. Math. Biol. 2005, 51, 247–267. [Google Scholar] [CrossRef]
- Nowak, M.A.; May, R.M. Virus Dynamics; Oxford University Press: New York, NY, USA, 2000. [Google Scholar]
- Lv, C.; Huang, L.; Yuan, Z. Global stability for an HIV-1 infection model with Beddington-DeAngelis incidence rate and CTL immune response. Commun. Nonlinear Sci. Numer. Simul. 2014, 19, 121–127. [Google Scholar] [CrossRef]
- Jiang, C.; Kong, H.; Zhang, G.; Wang, K. Global properties of a virus dynamics model with self-proliferation of CTLs. Math. Appl. Sci. Eng. 2021, 2, 123–133. [Google Scholar] [CrossRef]
- Ren, J.; Xu, R.; Li, L. Global stability of an HIV infection model with saturated CTL immune response and intracellular delay. Math. Biosci. Eng. 2020, 18, 57–68. [Google Scholar] [CrossRef] [PubMed]
- Wang, A.P.; Li, M.Y. Viral dynamics of HIV-1 with CTL immune response. Discret. Contin. Dyn. Syst. Ser. 2021, 26, 2257–2272. [Google Scholar] [CrossRef]
- Yang, Y.; Xu, R. Mathematical analysis of a delayed HIV infection model with saturated CTL immune response and immune impairment. J. Appl. Math. Comput. 2022, 68, 2365–2380. [Google Scholar] [CrossRef]
- Chen, C.; Zhou, Y. Dynamic analysis of HIV model with a general incidence, CTLs immune response and intracellular delays. Math. Comput. Simul. 2023, 212, 159–181. [Google Scholar] [CrossRef]
- Amarante-Mendes, G.P.; Adjemian, S.; Branco, L.M.; Zanetti, L.; Weinlich, R.; Bortoluci, K.R. Pattern recognition receptors and the host cell death molecular machinery. Front. Immunol. 2018, 9, 2379. [Google Scholar] [CrossRef]
- Jiang, Y.; Zhang, T. Global stability of a cytokine-enhanced viral infection model with nonlinear incidence rate and time delays. Appl. Math. Lett. 2022, 132, 108110. [Google Scholar] [CrossRef]
- Doitsh, G.; Galloway, N.L.K.; Geng, X.; Yang, Z.; Monroe, K.M.; Zepeda, O.; Hunt, P.W.; Hatano, H.; Sowinski, S.; Muñoz-Arias, I.; et al. Pyroptosis drives CD4 T-cell depletion in HIV-1 infection. Nature 2014, 505, 509–514. [Google Scholar] [CrossRef]
- Wang, W.; Zhang, T. Caspase-1-mediated pyroptosis of the predominance for driving CD4+T cells death: A nonlocal spatial mathematical model. Bull. Math. Biol. 2018, 80, 540–582. [Google Scholar] [CrossRef]
- Wang, W.; Feng, Z. Global dynamics of a diffusive viral infection model with spatial heterogeneity. Nonlinear Anal. Real World Appl. 2023, 72, 103763. [Google Scholar] [CrossRef]
- Wang, W.; Ma, W.; Feng, Z. Complex dynamics of a time periodic nonlocal and time-delayed model of reaction-diffusion equations for modelling CD4+T cells decline. J. Comput. Appl. Math. 2020, 367, 112430. [Google Scholar] [CrossRef]
- Wang, W.; Ren, X.; Ma, W.; Lai, X. New insights into pharmacologic inhibition of pyroptotic cell death by necrosulfonamide: A PDE model. Nonlinear Anal. Real World Appl. 2020, 56, 103173. [Google Scholar] [CrossRef]
- Wang, W.; Ren, X.; Wang, X. Spatial-temporal dynamics of a novel PDE model: Applications to pharmacologic inhibition of pyroptosis by necrosulfonamide. Commun. Nonlinear Sci. Numer. Simul. 2021, 103, 106025. [Google Scholar] [CrossRef]
- Xu, J. Dynamic analysis of a cytokine-enhanced viral infection model with infection age. Math. Biosci. Eng. 2023, 20, 8666–8684. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Xu, X.; Wang, X. Dynamic analysis of a cytokine-enhanced viral infection model with time delays and CTL immune response. Chaos Solitons Fractals 2023, 170, 113357. [Google Scholar] [CrossRef]
- Wodarz, D. Hepatitis C virus dynamics and pathology: The role of CTL and antibody responses. J. Gen. Virol. 2003, 84, 1743–1750. [Google Scholar] [CrossRef]
- Wang, J.; Pang, J.; Kuniya, T.; Enatsu, Y. Global threshold dynamics in a five-dimensional virus model with cell-mediated, humoral immune responses and distributed delays. Appl. Math. Comput. 2014, 241, 298–316. [Google Scholar] [CrossRef]
- Yan, Y.; Wang, W. Global stability of a five-dimensional model with immune responses and delay. Discret. Contin. Dyn. Syst. Ser. B 2012, 17, 401–416. [Google Scholar] [CrossRef]
- Dubey, P.; Dubey, U.S.; Dubey, B. Modeling the role of acquired immune response and antiretroviral therapy in the dynamics of HIV infection. Math. Comput. Simul. 2018, 144, 120–137. [Google Scholar] [CrossRef]
- Junxian, Y.; Wang, L. Dynamics analysis of a delayed HIV infection model with CTL immune response and antibody immune response. Acta Math. Sci. 2021, 41, 991–1016. [Google Scholar]
- Kuang, Y. Delay Differential Equations with Applications in Population Dynamics; Academic Press: Boston, MA, USA, 1993. [Google Scholar]
- Korobeinikov, A. Global properties of basic virus dynamics models. Bull. Math. Biol. 2004, 66, 879–883. [Google Scholar] [CrossRef]
- Huang, G.; Takeuchi, Y.; Ma, W. Lyapunov functionals for delay differential equations model of viral infections. Siam J. Appl. Math. 2010, 70, 2693–2708. [Google Scholar] [CrossRef]
- Hale, J.K.; Lunel, S.M.V. Introduction to Functional Differential Equations; Springer: New York, NY, USA, 1993. [Google Scholar]
- Khalil, H.K. Nonlinear Systems, 3rd ed.; Prentice Hall: Upper Saddle River, NJ, USA, 2002. [Google Scholar]
- Rathkey, J.K.; Zhao, J.; Liu, Z.; Chen, Y.; Yang, J.; Kondolf, H.C.; Benson, B.L.; Chirieleison, S.M.; Huang, A.Y.; Dubyak, G.R.; et al. Chemical disruption of the pyroptotic pore-forming protein gasdermin D inhibits inflammatory cell death and sepsis. Sci. Immunol. 2018, 3, eaat2738. [Google Scholar] [CrossRef]
- Li, F.; Ma, W. Dynamics analysis of an HTLV-1 infection model with mitotic division of actively infected cells and delayed CTL immune response. Math. Methods Appl. Sci. 2018, 41, 3000–3017. [Google Scholar] [CrossRef]
- Elaiw, A.M.; AlShamrani, N.H. Modeling and analysis of a within-host HTLV-I/HIV co-infection. Bol. Soc. Math. Mex. 2021, 1, 27–38. [Google Scholar] [CrossRef]
- Perelson, A.S.; Kirschner, D.E.; de Boer, R. Dynamics of HIV Infection of CD4+T cells. Math. Biosci. 1993, 114, 81–125. [Google Scholar] [CrossRef]
- Alshaikh; AlShamrani, N.H.; Elaiw, A.M. Stability of HIV/HTLV co-infection model with effective HIV-specific antibody immune response. Results Phys. 2021, 27, 104448. [Google Scholar] [CrossRef]
- Callaway, D.S.; Perelson, A.S. HIV-1 infection and low steady state viral loads. Bull. Math. Biol. 2002, 64, 29–64. [Google Scholar] [CrossRef]
- Mohri, H.; Bonhoeffer, S.; Monard, S.; Perelson, A.S.; Ho, D. Rapid turnover of T lymphocytes in SIV-infected rhesus macaques. Science 1998, 279, 1223–1227. [Google Scholar] [CrossRef]
- Elaiw, A.M.; AlShamrani, N.H. Stability of a delayed adaptive immunity HIV infection model with sitrnt infected cells and cellular infection. J. Appl. Anal. Comput. 2021, 11, 964–1005. [Google Scholar]
- Elaiw, A.M.; AlShamrani, N.H.; Hobiny, A.D. Stability of an adaptive immunity delayed HIV infection model with active and silent cell-to-cell spread. Math. Biosci. Eng. 2020, 17, 6401–6458. [Google Scholar] [CrossRef]
- Marino, S.; Hogue, I.B.; Ray, C.J.; Kirschner, D.E. A methodology for performing global uncertainty and sensitivity analysis in systems biology. J. Theor. Biol. 2008, 254, 178–196. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Zhang, H.; Yin, G. Distributed dynamic event-triggered secure model predictive control of vehicle platoon against DoS attacks. IEEE Trans. Veh. Technol. 2022, 72, 2863–2877. [Google Scholar] [CrossRef]
- Zhang, H.; Shi, Y.; Wang, J.; Chen, H. A new delay-compensation scheme for networked control systems in controller area networks. IEEE Trans. Ind. Electron. 2018, 65, 7239–7247. [Google Scholar] [CrossRef]
- Elaiw, A.M.; Xia, X. HIV dynamics: Analysis and robust multirate MPC-based treatment schedules. J. Math. Appl. 2009, 359, 285–301. [Google Scholar] [CrossRef]
- Elaiw, A.M.; AlAgha, A.D. Analysis of a delayed and diffusive oncolytic M1 virotherapy model with immune response. Nonlinear Anal. Real World Appl. 2020, 55, 103116. [Google Scholar] [CrossRef]
- Bellomo, N.; Burini, D.; Outada, N. Multiscale models of Covid-19 with mutations and variants. Netw. Heterog. Media 2022, 17, 293–310. [Google Scholar] [CrossRef]
- Gibelli, L.; Elaiw, A.M.; Alghamdi, M.A.; Althiabi, A.M. Heterogeneous population dynamics of active particles: Progression, mutations, and selection dynamics. Math. Model. Methods Appl. Sci. 2017, 27, 617–640. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Equilibrium Point | Global Stability Conditions | |
|---|---|---|
| None | ||
| , and | ||
| and | ||
| and | ||
| and | and |
| Parameter | Value | Source | Parameter | Value | Source | Parameter | Value | Source |
|---|---|---|---|---|---|---|---|---|
| 10 | [39,40,41] | [27] | [42] | |||||
| [40,43,44] | [27] | [27] | ||||||
| [27] | 13 | [27] | 0.1 | [42] | ||||
| [27] | [32] | 0.1 | [45] | |||||
| [46] | Assumed |
| Parameter | Sensitivity Index | Parameter | Sensitivity Index | Parameter | Sensitivity Index |
|---|---|---|---|---|---|
| 1 | |||||
| 0 | 0 | ||||
| 0 | |||||
| 0 | |||||
| 0 | |||||
| 0 | |||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dahy, E.; Elaiw, A.M.; Raezah, A.A.; Zidan, H.Z.; Abdellatif, A.E.A. Global Properties of Cytokine-Enhanced HIV-1 Dynamics Model with Adaptive Immunity and Distributed Delays. Computation 2023, 11, 217. https://doi.org/10.3390/computation11110217
Dahy E, Elaiw AM, Raezah AA, Zidan HZ, Abdellatif AEA. Global Properties of Cytokine-Enhanced HIV-1 Dynamics Model with Adaptive Immunity and Distributed Delays. Computation. 2023; 11(11):217. https://doi.org/10.3390/computation11110217
Chicago/Turabian StyleDahy, Elsayed, Ahmed M. Elaiw, Aeshah A. Raezah, Hamdy Z. Zidan, and Abd Elsattar A. Abdellatif. 2023. "Global Properties of Cytokine-Enhanced HIV-1 Dynamics Model with Adaptive Immunity and Distributed Delays" Computation 11, no. 11: 217. https://doi.org/10.3390/computation11110217