
Statistical Multivariate Methods for the Selection of High-Yielding Rapeseed Lines with Varied Seed Coat Color

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Field Trial
2.3. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chen, W.; Zhang, Y.; Liu, X.; Chen, B.; Tu, J.; Fu, T. Detection of QTL for six yield-related traits in oilseed rape (Brassica napus) using DH and immortalized F2 populations. Theor. Appl. Genet. 2007, 115, 849–858. [Google Scholar] [CrossRef]
- Jiang, H.L.; Hong, J.; Jiang, Y.T.; Yu, S.X.; Zhang, Y.J.; Shi, J.X.; Lin, W.H. Genome-Wide Association Analysis Identifies Candidate Genes Regulating Seed Number Per Silique in Arabidopsis thaliana. Plants 2020, 9, 585. [Google Scholar] [CrossRef]
- Wolko, J.; Dobrzycka, A.; Bocianowski, J.; Bartkowiak-Broda, I. Estimation of heterosis for yield-related traits for single cross and three-way cross hybrids of oilseed rape (Brassica napus L.). Euphytica 2019, 215, 156. [Google Scholar] [CrossRef]
- Wang, H.; Zaman, Q.U.; Huang, W.; Mei, D.; Liu, J.; Wang, W.; Ding, B.; Hao, M.; Fu, L.; Cheng, H.; et al. QTL and Candidate Gene Identification for Silique Length Based on High-Dense Genetic Map in Brassica napus L. Front. Plant Sci. 2019, 10, 1579. [Google Scholar] [CrossRef] [PubMed]
- Udall, J.A.; Quijada, P.A.; Lambert, B.; Osborn, T.C. Quantitative trait analysis of seed yield and other complex traits in hybrid spring rapeseed (Brassica napus L.): 2. Identification of alleles from unadapted germplasm. Theor. Appl. Genet. 2006, 113, 597–609. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.W.; Yang, G.S.; Liu, P.W.; Hong, D.F.; Li, S.P.; He, Q.B. Genetic and correlation analysis of silique-traits in Brassica napus L. by quantitative trait locus mapping. Theor. Appl. Genet. 2011, 122, 21–31. [Google Scholar] [CrossRef]
- Wang, X.; Chen, L.; Wang, A.; Wang, H.; Tian, J.; Zhao, X.; Chao, H.; Zhao, Y.; Zhao, W.; Xiang, J.; et al. Quantitative trait loci analysis and genome-wide comparison for silique related traits in Brassica napus. BMC Plant Biol. 2016, 16, 71. [Google Scholar] [CrossRef]
- Zając, T.; Klimek-Kopyra, A.; Oleksy, A.; Lorenc-Kozik, A.; Ratajczak, K. Analysis of yield and plant traits of oilseed rape (Brassica napus L.) cultivated in temperate region in light of the possibilities of sowing in arid areas. Acta Agrobot. 2016, 69, 1696. [Google Scholar] [CrossRef]
- Rahman, M.; McVetty, P.B.E. A review of Brassica seed color. Can. J. Plant Sci. 2011, 91, 437–446. [Google Scholar] [CrossRef]
- Nesi, N.; Delourme, R.; Bregeon, M.; Falentin, C.; Renard, M. Genetic and molecular approaches to improve nutritional value of Brassica napus L. seed. Comptes Rendus Biol. 2008, 331, 763–771. [Google Scholar] [CrossRef]
- Gawrysiak-Witulska, M.; Rudzińska, M.; Siger, A.; Bartkowiak-Broda, I. A high drying temperature causes degradation of sterols and tocopherols in yellow-seeded Brassica napus oils. Eur. J. Lipid Sci. Technol. 2015, 117, 483–490. [Google Scholar] [CrossRef]
- Rakow, G.; Relf-Eckstein, J.A.; Raney, J.P. Rapeseed genetic research to improve its agronomic performance and seed quality. Helia 2007, 30, 199–206. [Google Scholar] [CrossRef]
- Piotrowska, A.; Krótka, K.; Krzymański, J. Wartość gospodarcza żółtonasiennych linii rzepaku ozimego (Brassica napus L.). Economical value of yellow-seeded lines of winter oilseed rape (Brassica napus L.). Rośliny Oleiste Oilseed Crops 2000, 21, 359–368. [Google Scholar]
- Daoyu, Z.; Lawes, G.S. Manova and discriminant analysis of phenotypic data as a guide for parent selection in kiwifruit (Actinidia deliciosa) breeding. Euphytica 2000, 114, 151–157. [Google Scholar] [CrossRef]
- Vaylay, R.; van Santen, E. Application of canonical discriminant analysis for the assessment of genetic variation in tall fescue. Crop Sci. 2002, 42, 534–539. [Google Scholar] [CrossRef]
- Cegielska-Taras, T.; Tykarska, T.; Szala, L.; Kuras, M.; Krzymański, J. Direct plant development from microspore-derived embryos of winter oilseed rape Brassica napus L. ssp oleifera (DC.) Metzger. Euphytica 2002, 124, 341–347. [Google Scholar] [CrossRef]
- Michalski, K. Seed color assessment in rapeseed seeds using Color and Near Infrared Reflectance Spectrometers. Rośliny Oleiste Oilseed Crops 2009, 30, 119–132. [Google Scholar]
- Wolko, J.; Dobrzycka, A.; Bocianowski, J.; Szala, L.; Cegielska-Taras, T.; Bartkowiak-Broda, I.; Gacek, K. Genetic variation of traits affecting meal quality in black × yellow seeded doubled haploid population of winter oilseed rape. Agron. Res. 2020, 18, 2259–2270. [Google Scholar] [CrossRef]
- Gacek, K.; Bayer, P.E.; Anderson, R.; Severn-Ellis, A.A.; Wolko, J.; Łopatyńska, A.; Matuszczak, M.; Bocianowski, J.; Edwards, D.; Batley, J. QTL Genetic Mapping Study for Traits Affecting Meal Quality in Winter Oilseed Rape (Brassica napus L.). Genes 2021, 12, 1235. [Google Scholar] [CrossRef] [PubMed]
- Shapiro, S.S.; Wilk, M.B. An analysis of variance test for normality (complete samples). Biometrika 1965, 52, 591–611. [Google Scholar] [CrossRef]
- Rencher, A.C. Interpretation of canonical discriminant functions, canonical variates, and principal components. Am. Stat. 1992, 46, 217–225. [Google Scholar] [CrossRef]
- Seidler-Łożykowska, K.; Bocianowski, J. Evaluation of variability of morphological traits of selected caraway (Carum carvi L.) genotypes. Ind. Crops Prod. 2012, 35, 140–145. [Google Scholar] [CrossRef]
- Mahalanobis, P.C. On the generalized distance in statistics. Proc. Natl. Acad. Sci. India A 1936, 12, 49–55. [Google Scholar] [CrossRef]
- Skomra, U.; Bocianowski, J.; Agacka, M. Agro-morphological differentiation between European hop (Humulus lupulus L.) cultivars in relation to their origin. J. Food Agric. Environ. 2013, 11, 1123–1128. [Google Scholar]
- Bocianowski, J.; Krajewski, P. Comparison of the genetic additive effect estimators based on phenotypic observations and on molecular marker data. Euphytica 2009, 165, 113–122. [Google Scholar] [CrossRef]
- Bocianowski, J. Analytical and numerical comparisons of two methods of estimation of additive × additive interaction of QTL effects. Sci. Agric. 2012, 69, 240–246. [Google Scholar] [CrossRef]
- Bocianowski, J.; Warzecha, T.; Nowosad, K.; Bathelt, R. Genotype by environment interaction using AMMI model and estimation of additive and epistasis gene effects for 1000-kernel weight in spring barley (Hordeum vulgare L.). J. Appl. Genet. 2019, 60, 127–135. [Google Scholar] [CrossRef]
- Liersch, A.; Bocianowski, J.; Nowosad, K.; Mikołajczyk, K.; Spasibionek, S.; Wielebski, F.; Matuszczak, M.; Szała, L.; Cegielska-Taras, T.; Sosnowska, K.; et al. Effect of Genotype × Environment Interaction for Seed Traits in Winter Oilseed Rape (Brassica napus L.). Agriculture 2020, 10, 607. [Google Scholar] [CrossRef]
- Bartkowiak-Broda, I.; Piotrowska, A.; Hernacki, B.; Cegielska-Taras, T.; Michalski, K. Development of germplasm of yellow seeded winter oilseed rape (Brassica napus L. var. oleifera). In Proceedings of the GCIRC Technical Meeting, Manesar, India, 28–29 September 2009. [Google Scholar]
- Myszka, K.; Boros, D.; Piotrowska, A.; Bartkowiak-Broda, I. Porównanie składu chemicznego śrut rzepakowych uzyskanych z rzepaku ozimego (Brassica napus L.) o zróżnicowanej barwie nasion. Comparison of chemical composition of rapeseed meals derived from winter rapeseed (Brassica napus L.) differing in colour of seeds. Rośliny Oleiste Oilseed Crops 2011, 32, 257–268. [Google Scholar]
- Zhu, Y.; Ye, J.; Zhan, J.; Zheng, X.; Zhang, J.; Shi, J.; Wang, X.; Liu, G.; Wang, H. Validation and Characterization of a Seed Number Per Silique Quantitative Trait Locus qSN.A7 in Rapeseed (Brassica napus L.). Front. Plant Sci. 2020, 11, 68. [Google Scholar] [CrossRef]
- Zhao, W.; Zhang, L.; Chao, H.; Wang, H.; Ta, N.; Li, H.; Wang, X.; Li, S.; Xiang, J.; Zhang, L.; et al. Genome-wide identification of silique-related traits based on high-density genetic linkage map in Brassica napus. Mol. Breeding 2019, 39, 86. [Google Scholar] [CrossRef]
- Shi, J.Q.; Li, R.Y.; Qiu, D.; Jiang, C.C.; Long, Y.; Morgan, C.; Bancroft, I.; Zhao, J.Y.; Meng, J.L. Unraveling the complex trait of crop yield with quantitative trait loci mapping in Brassica napus. Genetics 2009, 182, 851–861. [Google Scholar] [CrossRef] [PubMed]
- Cai, D.; Xiao, Y.; Yang, W.; Ye, W.; Wang, B.; Younas, M.; Wu, J.; Liu, K. Association mapping of six yield-related traits in rapeseed (Brassica napus L.). Theor. Appl. Genet. 2014, 127, 85–96. [Google Scholar] [CrossRef]
- Fu, Y.; Wei, D.; Dong, H.; He, Y.; Cui, Y.; Mei, J.; Wan, H.; Li, J.; Snowdon, R.; Friedt, W.; et al. Comparative quantitative trait loci for silique length and seed weight in Brassica napus. Sci. Rep. 2015, 5, 14407. [Google Scholar] [CrossRef] [PubMed]
- Diepenbrock, W. Yield analysis of winter oilseed rape (Brassica napus L.): A review. Field Crop Res. 2000, 67, 35–49. [Google Scholar] [CrossRef]
- Elliott, R.H.; Mann, L.W.; Olfert, O.O. Effects of seed size and seed weight on seedling establishment, seedling vigour and tolerance of summer turnip rape (Brassica rapa) to flea beetles, Phyllotreta spp. Can. J. Plant Sci. 2007, 87, 385–393. [Google Scholar] [CrossRef]
- Sharafi, Y.; Majidi, M.M.; Jafarzadeh, M.; Mirlohi, A. Multivariate Analysis of Genetic Variation in Winter Rapeseed (Brassica napus L.) Cultivars. J. Agric. Sci. Technol. 2015, 17, 1319–1331. [Google Scholar]
- Łopatyńska, A.; Bocianowski, J.; Cyplik, A.; Wolko, J. Multidimensional Analysis of Diversity in DH Lines and Hybrids of Winter Oilseed Rape (Brassica napus L.). Agronomy 2021, 11, 645. [Google Scholar] [CrossRef]
- Bocianowski, J.; Liersch, A.; Bartkowiak-Broda, I. Badanie zmienności fenotypowej mieszańców F1 CMS ogura rzepaku ozimego i ich form rodzicielskich za pomocą statystycznych metod wielowymiarowych. Investigation of phenotypic distance of F1 CMS ogura winter oilseed rape hybrids and parental lines using multivariate statistical methods. Rośliny Oleiste Oilseed Crops 2009, 30, 161–184. [Google Scholar]
- Parvin, E.; Mahmud, F.; Bhuiyan, S.R.; Haque, M. Multivariate Analysis of Genetic Variation in Rapeseed (Brassica napus L.). Agric. Food Sci. Res. 2019, 6, 1–8. [Google Scholar] [CrossRef]
- Mili, S.M.S.K.; Monalesa, N.; Rahman, M.J.; Afsana, N.; Zeba, N.; Mahmud, F. Genetic diversity analysis in F5 population of Brassica napus L. Int. J. Biosci. 2016, 9, 177–184. [Google Scholar] [CrossRef]
- Singh, S.; Dwivedi, A.K.; Ashutosh, K.O.; Kumar, K. Genetic Divergence Analysis in Indian mustard (Brassica juncea L.). Int. J. Curr. Microbiol. App. Sci. 2018, 7, 2496–2503. [Google Scholar] [CrossRef]
- Bocianowski, J.; Nowosad, K.; Dobrzycka, A.; Wolko, J. Estimation of additive and epistatic gene effects of doubled haploid lines of winter oilseed rape (Brassica napus L.). Euphytica 2017, 213, 122. [Google Scholar] [CrossRef]
- Luo, X.; Ding, Y.; Zhang, L.; Yue, Y.; Snyder, J.H.; Ma, C.; Zhu, J. Genomic Prediction of Genotypic Effects with Epistasis and Environment Interactions for Yield-Related Traits of Rapeseed (Brassica napus L.). Front. Genet. 2017, 8, 15. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
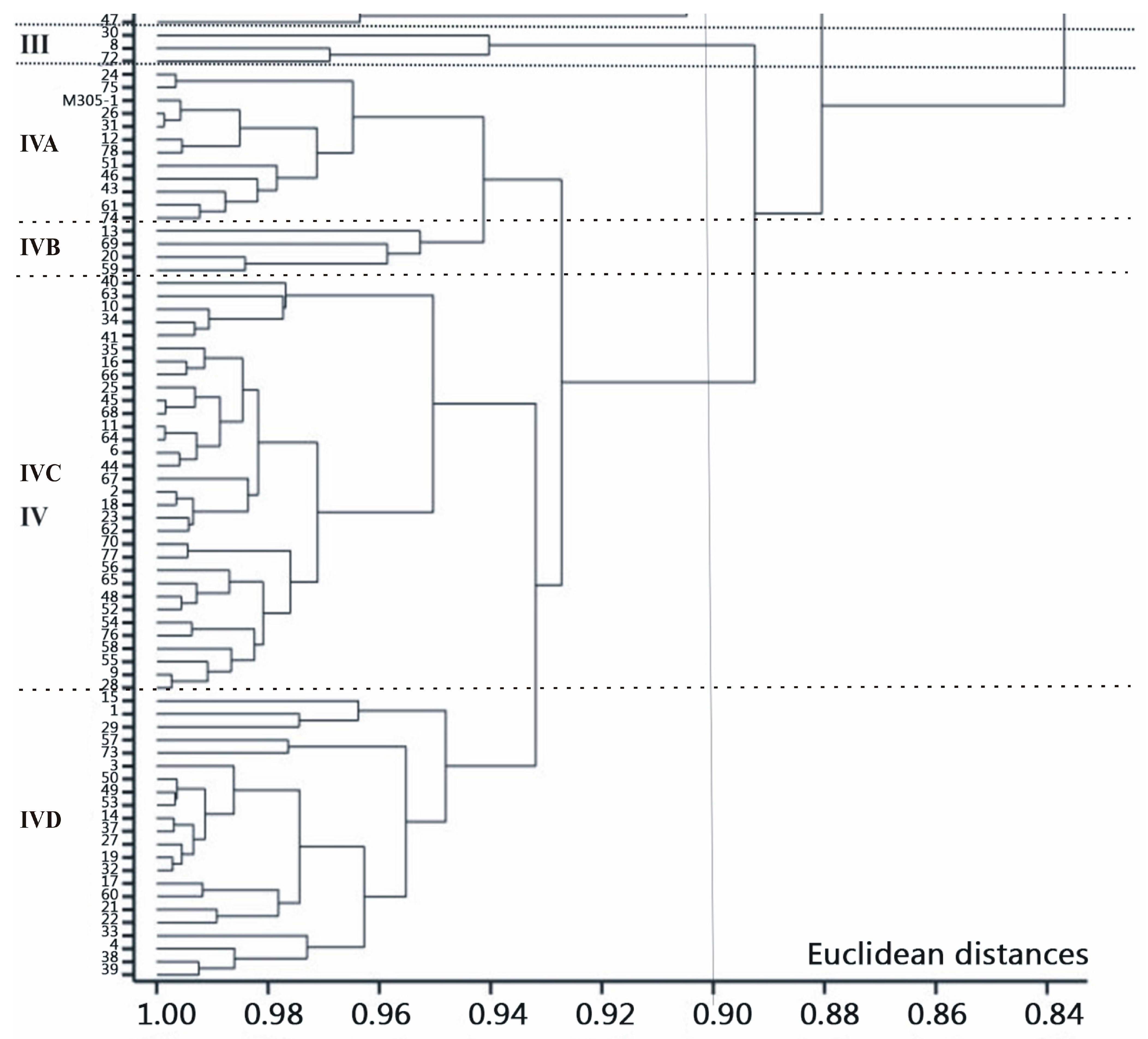{kind=link}
| Silique Length [mm] | Silique Width [mm] | Number of Seeds per Silique | Seed Coat Color [0–5] | Thousand-Seed Weight [g] | |
|---|---|---|---|---|---|
| Mean | 53.66 | 3.65 | 15.44 | 1.52 | 5.01 |
| Min. | 41.98 | 3.12 | 8.50 | 0.10 | 3.72 |
| Max. | 69.63 | 4.26 | 18.88 | 4.75 | 6.57 |
| s.d. | 3.88 | 0.29 | 2.15 | 0.42 | 0.64 |
| LSD0.05 | 7.72 | 0.53 | 3.92 | 0.84 | 1.19 |
| CV [%] | 7.24 | 7.88 | 13.90 | 27.52 | 12.82 |
| Parameter | Estimate | Standard Error | t-Statistic |
|---|---|---|---|
| Constant | −0.229 | 1.043 | −0.22 ns |
| Silique length [mm] | 0.058 | 0.017 | 3.381 ** |
| Silique width [mm] | 0.875 | 0.208 | 4.205 *** |
| Number of seeds per silique | −0.066 | 0.026 | −2.572 * |
| Seed coat color [0–5] | −0.010 | 0.050 | −0.202 ns |
| Percentage variance accounted | 28.90% | ||
| Trait | First Canonical Variate | Second Canonical Variate |
|---|---|---|
| Silique length [mm] | 0.289 ** | 0.64 *** |
| Silique width [mm] | −0.074 | −0.374 *** |
| Number of seeds per silique | −0.129 | −0.101 |
| Seed coat color [0–5] | −0.995 *** | 0.058 |
| Thousand-seed weight [g] | 0.12 | 0.378 *** |
| Percentage of explained multivariate variability | 64.95% | 13.21% |
| Effect | Silique Length [mm] | Silique Width [mm] | Number of Seeds per Silique | Seed Coat Color [0–5] | Thousand-Seed Weight [g] |
|---|---|---|---|---|---|
| parental forms | |||||
| aParents 1 | 4.24 | 0.005 | 0.235 | 2.174 *** | 0.337 |
| doubled haploid lines | |||||
| aDH 2 | 13.83 *** | 0.570 * | 5.190 *** | 2.13 *** | 1.423 ** |
| aaDH 3 | −39.83 *** | −3.087 *** | −10.233 *** | 0.634 ** | −3.586 *** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Łopatyńska, A.; Wolko, J.; Bocianowski, J.; Cyplik, A.; Gacek, K. Statistical Multivariate Methods for the Selection of High-Yielding Rapeseed Lines with Varied Seed Coat Color. Agriculture 2023, 13, 992. https://doi.org/10.3390/agriculture13050992
Łopatyńska A, Wolko J, Bocianowski J, Cyplik A, Gacek K. Statistical Multivariate Methods for the Selection of High-Yielding Rapeseed Lines with Varied Seed Coat Color. Agriculture. 2023; 13(5):992. https://doi.org/10.3390/agriculture13050992
Chicago/Turabian StyleŁopatyńska, Agnieszka, Joanna Wolko, Jan Bocianowski, Adrian Cyplik, and Katarzyna Gacek. 2023. "Statistical Multivariate Methods for the Selection of High-Yielding Rapeseed Lines with Varied Seed Coat Color" Agriculture 13, no. 5: 992. https://doi.org/10.3390/agriculture13050992