
Evaluation of Aspergillus flavus Growth and Detection of Aflatoxin B1 Content on Maize Agar Culture Medium Using Vis/NIR Hyperspectral Imaging


Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Preparation
2.2. Image Acquisition and Calibration
2.3. Quantitative Measurement of the AFB1 Concentration
2.4. Data Analysis
3. Results and Discussion
3.1. Critical Growth Characteristics of A. flavus in Culture Media
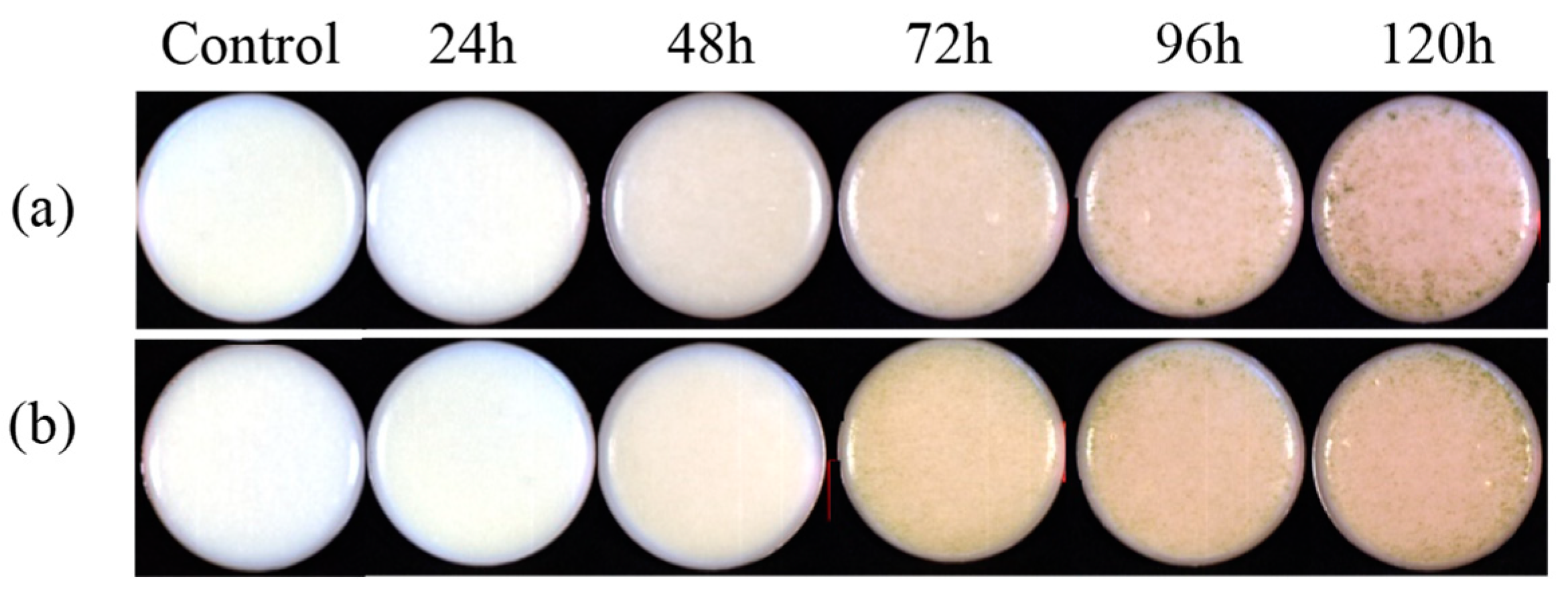
3.1.1. Image Information of A. flavus Growth
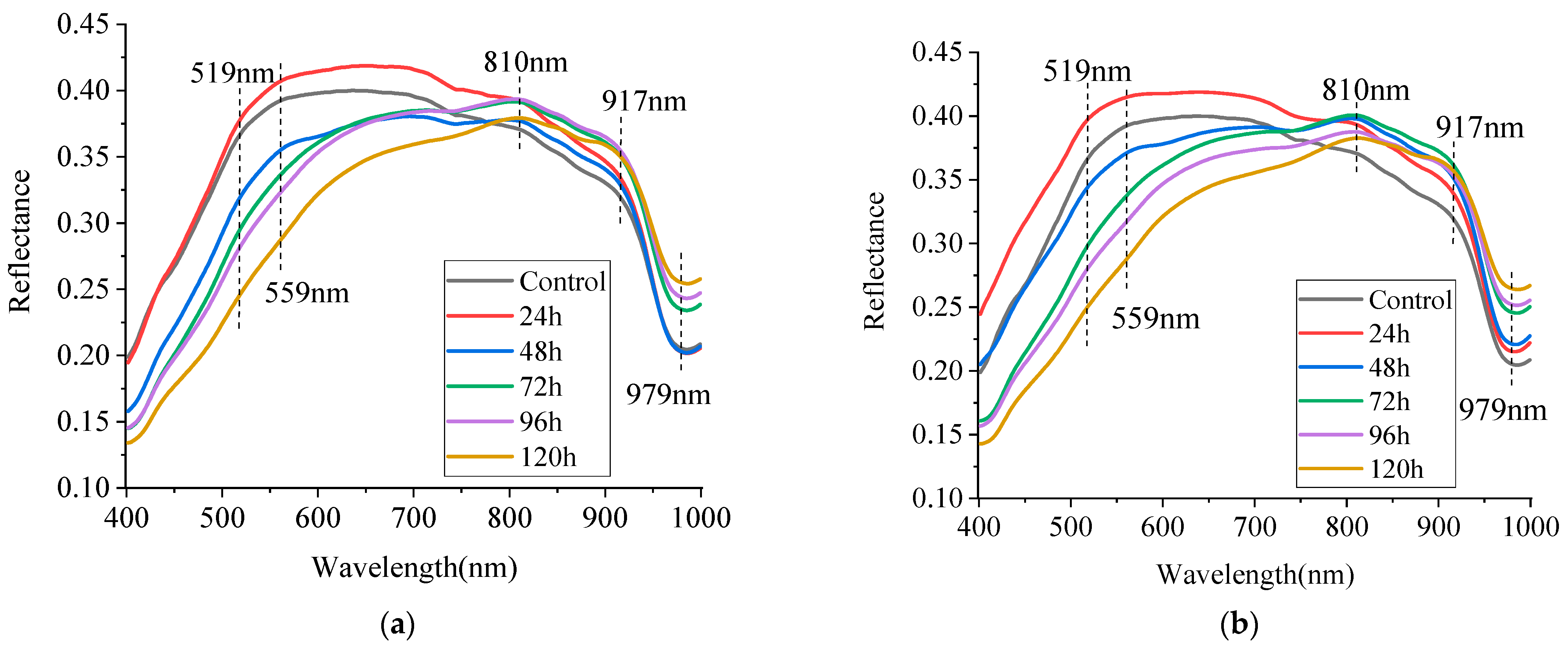
3.1.2. Spectral Characteristics of Fungal Culture in Maize Agar Medium
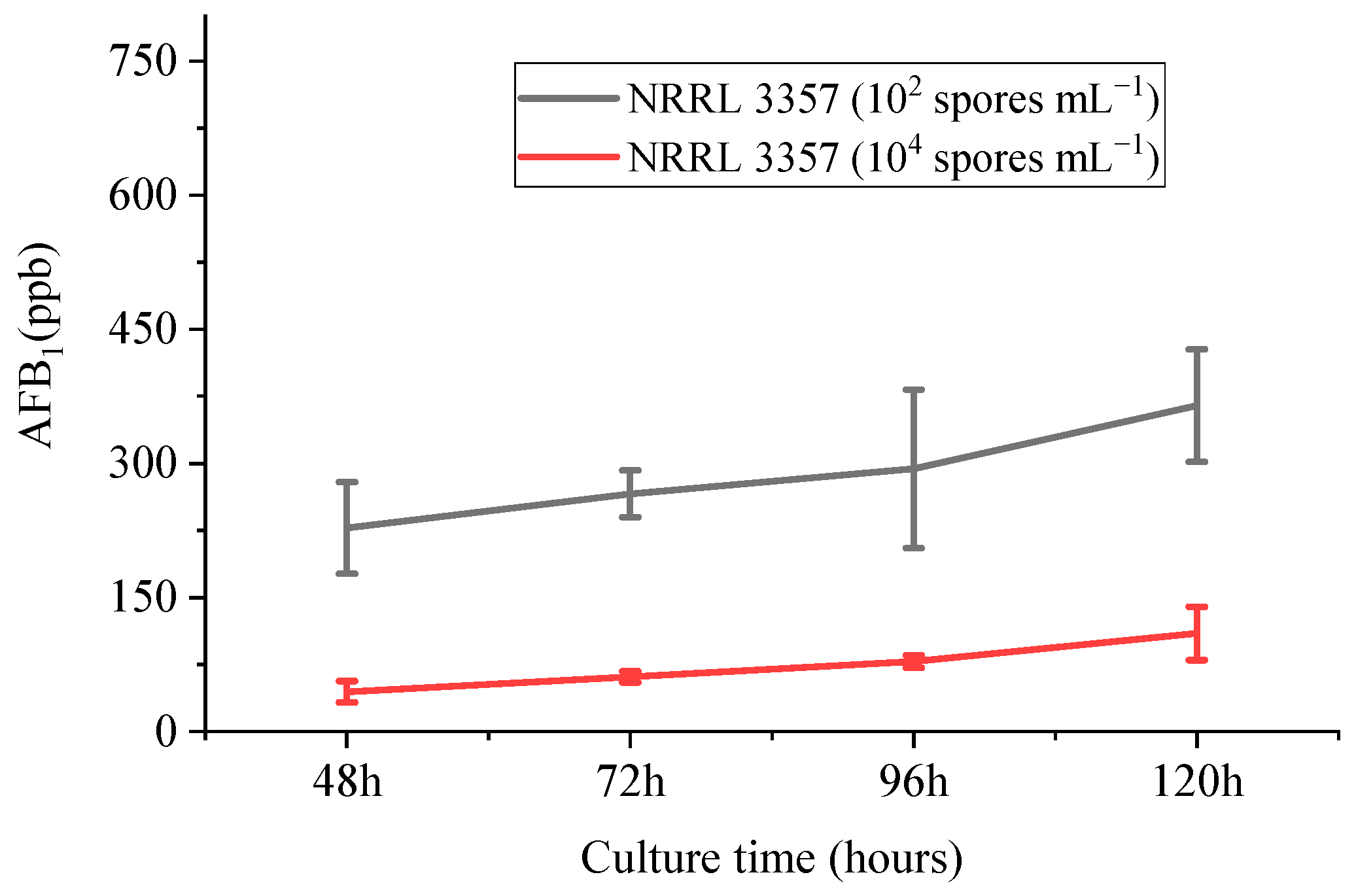
3.1.3. HPLC Analysis of AFB1 Accumulation
3.2. Identification of the A. flavus Growth Period
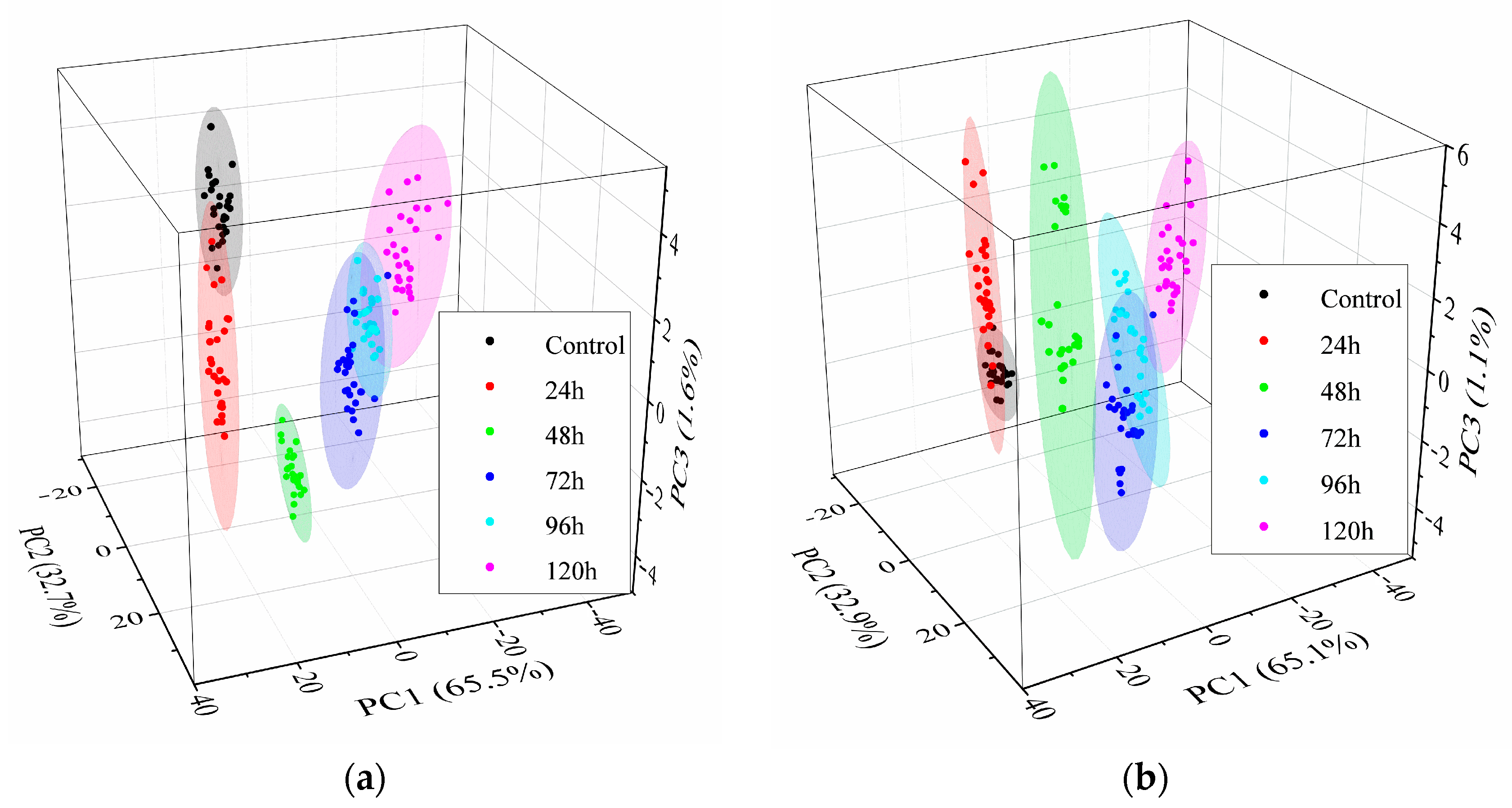
3.2.1. Identification of the A. flavus Growth Period by PCA
3.2.2. Identification of the A. flavus Growth Period based on PCA-SVM
3.3. Detection and Prediction of AFB1 Content in the Culture Medium
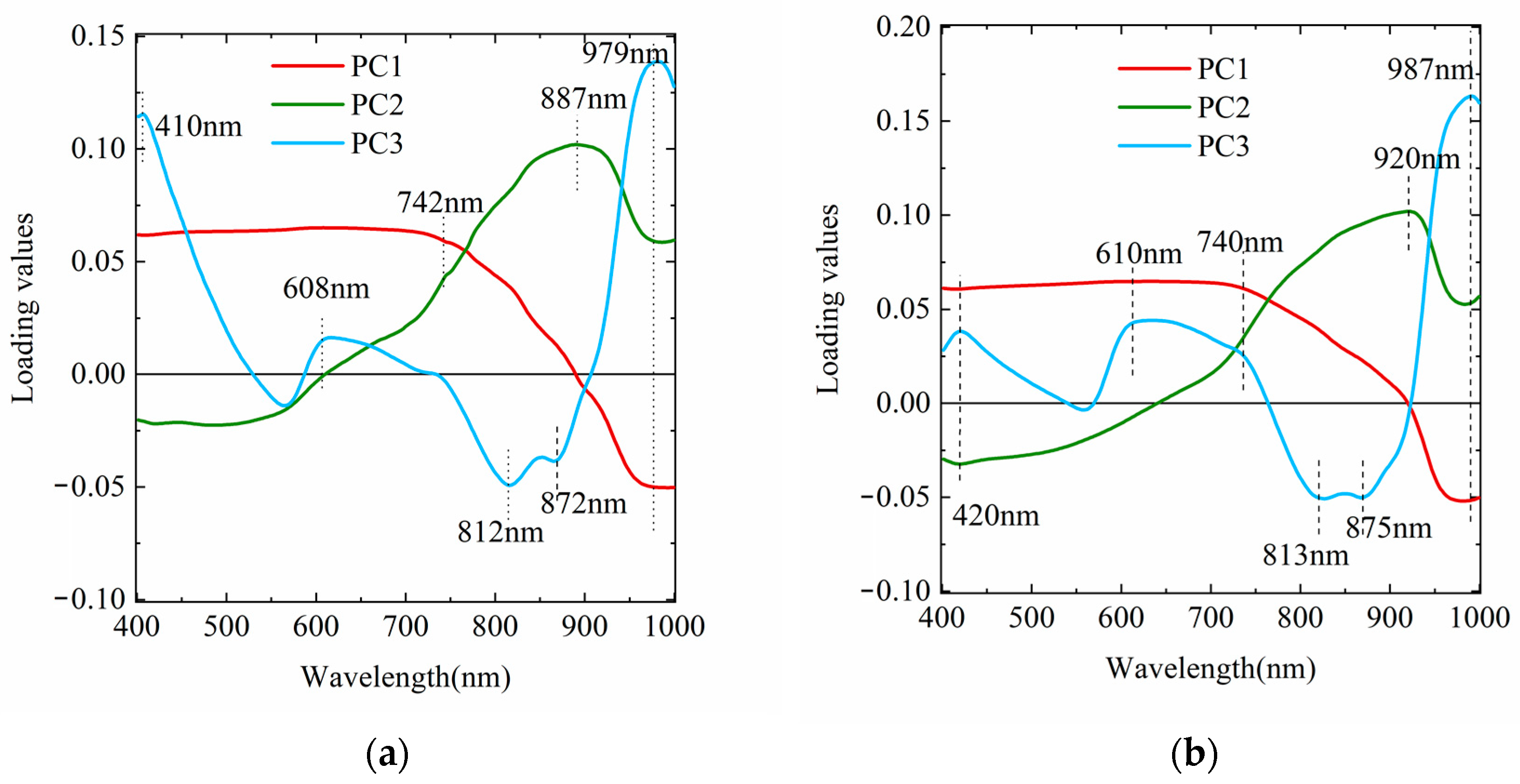
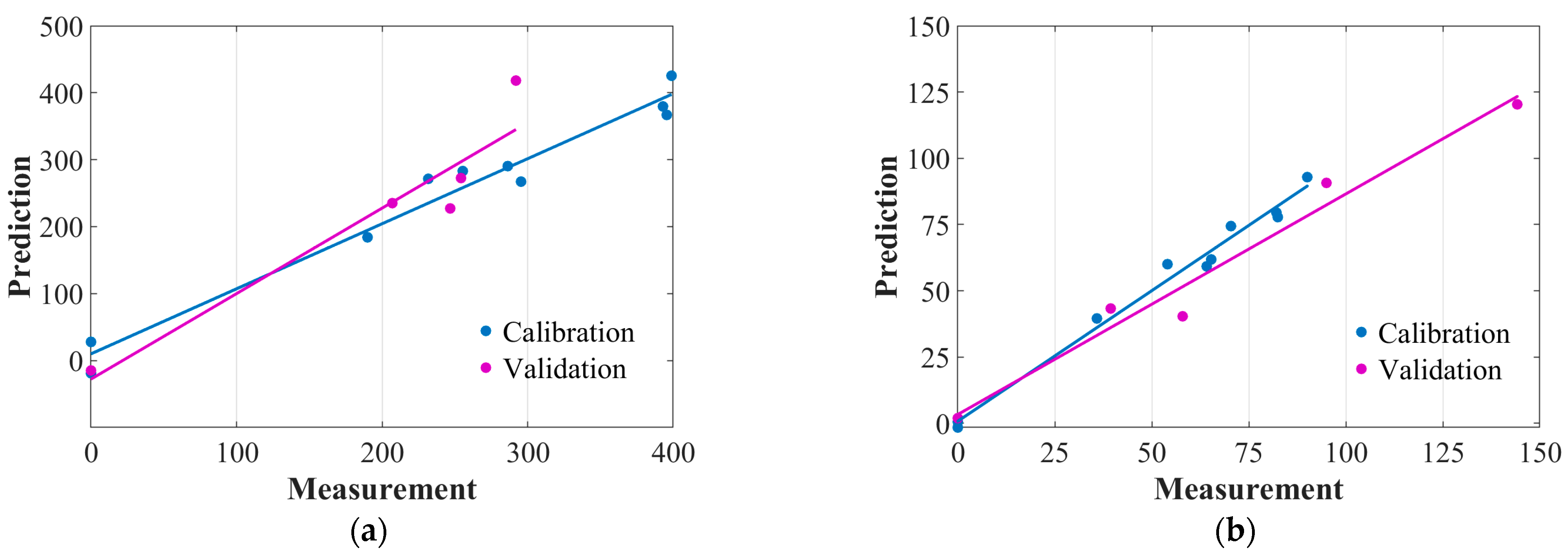
3.3.1. Hyperspectral PLSR Models
3.3.2. Development of Multispectral PLSR Models
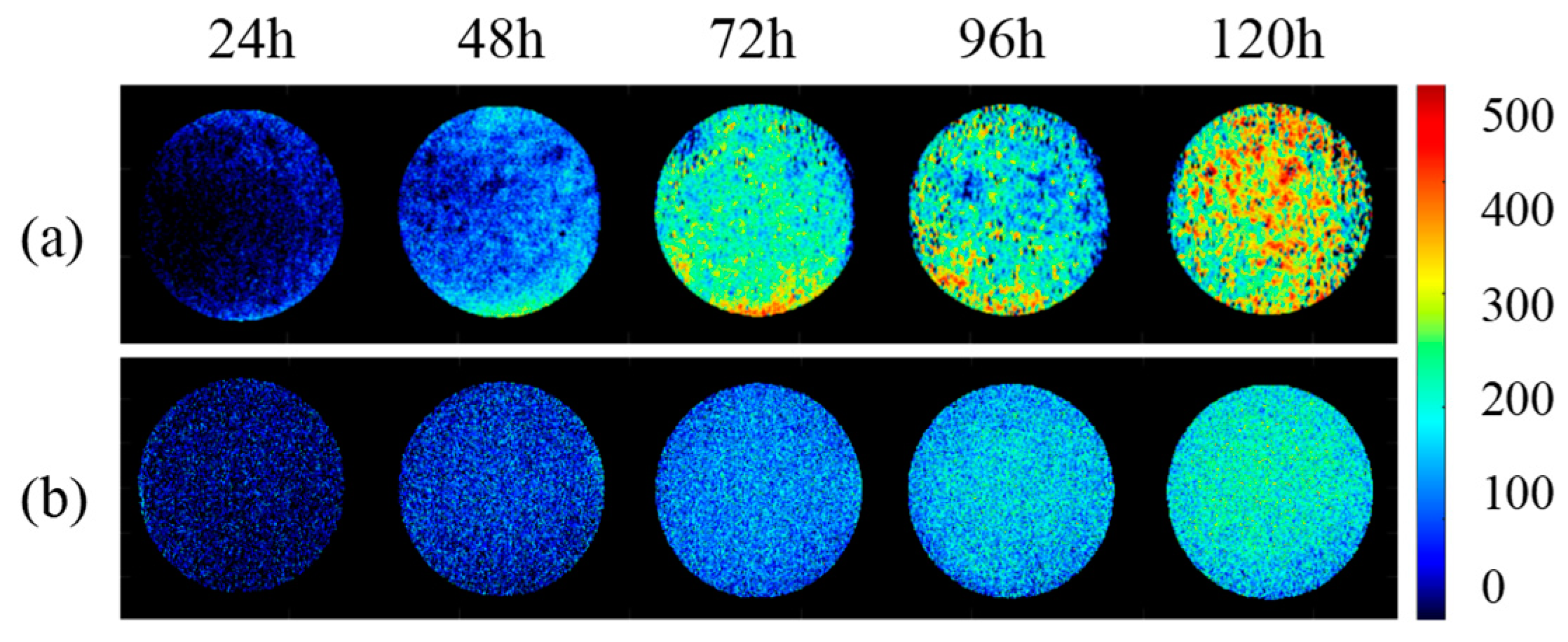
3.3.3. Prediction Image of AFB1 Content
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Su, W.; Yang, C.; Dong, Y.; Johnson, R.; Page, R.; Szinyei, T.; Hirsch, C.D.; Steffenson, B.J. Hyperspectral imaging and improved feature variable selection for automated determination of deoxynivalenol in various genetic lines of barley kernels for resistance screening. Food Chem. 2021, 343, 12857. [Google Scholar] [CrossRef] [PubMed]
- Su, W.; Sun, D. Multispectral Imaging for Plant Food Quality Analysis and Visualization. Compr. Rev. Food Sci. Food Saf. 2018, 17, 220–239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dipaolo, J.A.; Elis, J.; Erwin, H. Teratogenic Response by Hamsters Rats and Mice to Aflatoxin B1. Nature 1967, 215, 638. [Google Scholar] [CrossRef] [PubMed]
- Peraica, M.; Radic, B.; Lucic, A.; Pavlovic, M. Toxic effects of mycotoxins in humans. Bull. World Health Organ. 1999, 77, 754–766. Available online: https://www.webofscience.com/wos/woscc/full-record/WOS:000082975100008 (accessed on 27 September 2021).
- Tao, F.; Yao, H.; Hruska, Z.; Burger, L.W.; Rajasekaran, W.; Bhatnagar, D. Recent development of optical methods in rapid and non-destructive detection of aflatoxin and fungal contamination in agricultural products. TrAC Trends Anal. Chem. 2018, 100, 65–81. [Google Scholar] [CrossRef]
- Xu, L.; Lu, Z.; Cao, L.; Pang, H.; Zhan, Q.; Fu, Y.; Xiong, Y.; Li, Y.; Wang, X.; Wang, J.; et al. In-field detection of multiple pathogenic bacteria in food products using a portable fluorescent biosensing system. Food Control 2017, 75, 21–28. [Google Scholar] [CrossRef]
- Edelstein, L.; Segel, L.A. Growth and Metabolism in Mycelial Fungi. J. Theor. Biol. 1983, 104, 187–210. [Google Scholar] [CrossRef]
- Rahmani, A.; Jinap, S.; Soleimany, F. Qualitative and quantitative analysis of mycotoxins. Compr. Rev. Food Sci. Food Saf. 2009, 8, 202–251. [Google Scholar] [CrossRef]
- Xie, L.; Chen, M.; Ying, Y. Development of Methods for Determination of Aflatoxins. Crit. Rev. Food Sci. Nutr. 2016, 56, 2624–2664. [Google Scholar] [CrossRef]
- Matabaro, E.; Ishimwe, N.; Uwimbabazi, E.; Lee, B.H. Current Immunoassay Methods for the Rapid Detection of Aflatoxin in Milk and Dairy Product. Compr. Rev. Food Sci. Food Saf. 2017, 16, 808–820. [Google Scholar] [CrossRef]
- Xing, F.; Yao, H.; Liu, Y.; Dai, X.; Brown, R.L.; Bhatnagar, D. Recent developments and applications of hyperspectral imaging for rapid detection of mycotoxins and mycotoxigenic fungi in food products. Crit. Rev. Food Sci. Nutr. 2019, 59, 173–180. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Pu, H.; Sun, D.; Liu, D.; Wang, Q.; Xiong, Z. Application of Hyperspectral Imaging for Prediction of Textural Properties of Maize Seeds with Different Storage Periods. Food Anal. Methods 2014, 8, 1535–1545. [Google Scholar] [CrossRef]
- Cotty, P.J. Aflatoxin-producing potential of communities of Aspergillus section Flavi from cotton producing areas in the United States. Mycol Res. 1997, 101, 698–704. [Google Scholar] [CrossRef] [Green Version]
- Garcia, D.; Ramos, A.J.; Sanchis, V.; Marín, S. Modeling kinetics of aflatoxin production by Aspergillus flavus in maize-based medium and maize grain. Int. J. Food Microbiol. 2013, 162, 182–189. [Google Scholar] [CrossRef]
- Williams, P.J.; Geladi, P.; Britz, T.J.; Manley, M. Growth characteristics of three Fusarium species evaluated by near-infrared hyperspectral imaging and multivariate image analysis. Appl. Microbiol. Biotechnol. 2012, 96, 803–813. [Google Scholar] [CrossRef]
- Jin, J.; Lie, T.; Hruska, Z.; Yao, H. Classification of toxigenic and atoxigenic strains of Aspergillus flavus with hyperspectral imaging. Comput. Electron. Agric. 2009, 69, 158–164. [Google Scholar] [CrossRef]
- Chu, X.; Wang, W.; Ni, X.; Zheng, H.; Zhao, X.; Zhang, R.; Li, Y. Growth Identification of Aspergillus flavus and Aspergillus parasiticus by Visible/Near-Infrared Hyperspectral Imaging. Appl. Sci. 2018, 8, 513. [Google Scholar] [CrossRef] [Green Version]
- Lu, Y.; Wang, W.; Huang, M.; Ni, X.; Chu, X.; Li, C. Evaluation and classification of five cereal fungi on culture medium using Visible/Near-Infrared (Vis/NIR) hyperspectral imaging. Infrared Phys. Technol. 2020, 105, 103206. [Google Scholar] [CrossRef]
- Marin, S.; Belli, N.; Lasram, S.; Chebil, S.; Ramos, A.J.; Ghorbel, A.; Sanchis, V. Kinetics of Ochratoxin A Production and Accumulation by Aspergillus carbonarius on Synthetic Grape Medium at Different Temperature Levels. J. Food Sci. 2006, 71, 196–200. [Google Scholar] [CrossRef]
- Pearson, T.C.; Wicklow, D.T.; Pasikatan, M.C. Reduction of Aflatoxin and Fumonisin Contamination in Yellow Corn by High-Speed Dual-Wavelength Sorting. Cereal Chem. 2004, 81, 490–498. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Heitschmidt, G.W.; Ni, X.; Windham, W.R.; Hawkins, S.; Chu, X. Identification of aflatoxin B1 on maize kernel surfaces using hyperspectral imaging. Food Control 2014, 42, 78–86. [Google Scholar] [CrossRef]
- Jiang, H.; Wang, W.; Zhuang, H.; Yoon, S.C.; Yang, Y.; Zhao, X. Hyperspectral imaging for a rapid detection and visualization of duck meat adulteration in beef. Food Anal. Methods 2019, 12, 2205–2215. [Google Scholar] [CrossRef]
- Gromski, P.S.; Correa, E.; Vaughan, A.A.; Wedge, D.C.; Turner, M.L.; Goodacre, R. A comparison of different chemo-metrics approaches for the robust classification of electronic nose data. Anal. Bioanal. Chem. 2014, 406, 7581–7590. [Google Scholar] [CrossRef] [PubMed]
- Nicolai, B.M.; Theron, K.I.; Lammertyn, J. Kernel PLS regression on wavelet transformed NIR spectra for prediction of sugar content of apple. Chemom. Intell. Lab. Syst. 2007, 85, 243–252. [Google Scholar] [CrossRef]
- Sacre, P.Y.; Deconinck, E.; De Beer, T.; Courselle, P.; Vancauwenberghe, R.; Chiap, P.; Crommen, J.; De Beer, J.O. Comparison and combination of spectroscopic techniques for the detection of counterfeit medicines. J. Pharm. Biomed. Anal. 2010, 53, 445–453. [Google Scholar] [CrossRef] [Green Version]
- Menesatti, P.; Zanella, A.; D’Andrea, S.; Costa, C.; Paglia, G.; Pallottino, F. Supervised Multivariate Analysis of Hyper-spectral NIR Images to Evaluate the Starch Index of Apples. Food Bioprocess Technol. 2009, 2, 308–314. [Google Scholar] [CrossRef]
- Barlocco, N.; Vadell, A.; Ballesteros, F.; Galietta, G.; Cozzolino, D. Predicting intramuscular fat, moisture and Warner-Bratzler shear force in pork muscle using near infrared reflectance spectroscopy. Anim. Sci. 2006, 82, 111–116. [Google Scholar] [CrossRef]
- Mellon, J.E.; Dowd, M.K.; Cotty, P.J. Time Course Study of Substrate Utilization by Aspergillus flavus in Medium Simulating Corn (Zea mays) Kernels. J. Agric. Food Chem. 2002, 50, 648–652. [Google Scholar] [CrossRef]
- Amare, M.G.; Keller, N.P. Molecular mechanisms of Aspergillus flavus secondary metabolism and development. Fungal Genet. Biol. 2014, 66, 11–18. [Google Scholar] [CrossRef]
- Brown, S.H.; Zarnowski, R.; Sharpee, W.C.; Keller, N.P. Morphological Transitions Governed by Density Dependence and Lipoxygenase Activity in Aspergillus flavus. Appl. Environ. Microb. 2008, 74, 5674–5685. [Google Scholar] [CrossRef] [Green Version]
- Affeldt, K.J.; Brodhagen, M.; Keller, N.P. Aspergillus Oxylipin Signaling and Quorum Sensing Pathways Depend on G Protein-Coupled Receptors. Toxins 2012, 4, 695–717. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Modeling Method | Inoculation Methods | Accuracy | ||
|---|---|---|---|---|
| Calibration Set | Validation Set | Cross Validation | ||
| PCA-SVM | NRRL 3357 (102 spores mL−1) | 0.97 | 0.96 | 0.94 |
| NRRL 3357 (104 spores mL−1) | 0.97 | 0.91 | 0.92 | |
| Group | NRRL 3357 (102 spores mL−1) | NRRL 3357 (104 spores mL−1) | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Control | 24 h | 48 h | 72 h | 96 h | 120 h | Control | 24 h | 48 h | 72 h | 96 h | 120 h | |
| Control | 27 | 0 | 0 | 0 | 0 | 0 | 26 | 1 | 0 | 0 | 0 | 0 |
| 24 h | 1 | 26 | 0 | 0 | 0 | 0 | 0 | 27 | 0 | 0 | 0 | 0 |
| 48 h | 0 | 0 | 27 | 0 | 0 | 0 | 0 | 0 | 27 | 0 | 0 | 0 |
| 72 h | 0 | 0 | 0 | 25 | 2 | 0 | 0 | 0 | 0 | 25 | 1 | 1 |
| 96 h | 0 | 0 | 0 | 2 | 25 | 0 | 0 | 0 | 0 | 6 | 21 | 0 |
| 120 h | 0 | 0 | 0 | 0 | 0 | 27 | 0 | 0 | 0 | 0 | 0 | 27 |
| Sensitivity | 1.00 | 0.96 | 1.00 | 0.93 | 0.93 | 1.00 | 0.96 | 1.00 | 1.00 | 0.93 | 0.78 | 1.00 |
| Specificity | 0.99 | 1.00 | 1.00 | 0.99 | 0.99 | 1.00 | 1.00 | 0.99 | 1.00 | 0.96 | 0.99 | 0.99 |
| Accuracy | 0.97 | 0.94 | ||||||||||
| Kappa coefficient | 0.9630 | 0.9333 | ||||||||||
| Inoculation Methods | Preprocessing Methods | LVs | RC | RMSEc | RV | RMSEv | RPD |
|---|---|---|---|---|---|---|---|
| NRRL 3357 (102 spores mL−1) | Raw | 4 | 0.93 | 20.851 | 0.92 | 41.547 | 2.15 |
| SNV | 5 | 0.94 | 35.032 | 0.91 | 56.074 | 2.02 | |
| FD | 3 | 0.91 | 40.609 | 0.87 | 58.475 | 1.99 | |
| NRRL 3357 (104 spores mL−1) | Raw | 4 | 0.94 | 13.177 | 0.93 | 28.957 | 2.26 |
| SNV | 4 | 0.95 | 12.770 | 0.94 | 33.641 | 2.54 | |
| FD | 3 | 0.94 | 14.040 | 0.92 | 37.374 | 2.23 |
| Inoculation Methods | Number | Wavelengths (nm) |
|---|---|---|
| NRRL 3357 (102 spores mL−1) | 7 | 419, 487, 622, 697, 771, 880, 979 |
| NRRL 3357 (104 spores mL−1) | 6 | 417, 475, 559, 619, 796, 873 |
| Inoculation Methods | LVs | RC | RMSEc | RV | RMSEv | RPD |
|---|---|---|---|---|---|---|
| NRRL 3357 (102 spores mL−1) | 4 | 0.98 | 5.426 | 0.95 | 15.235 | 2.42 |
| NRRL 3357 (104 spores mL−1) | 2 | 0.99 | 3.856 | 0.96 | 17.438 | 1.58 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, X.; Jia, B.; Zhang, H.; Ni, X.; Zhuang, H.; Lu, Y.; Wang, W. Evaluation of Aspergillus flavus Growth and Detection of Aflatoxin B1 Content on Maize Agar Culture Medium Using Vis/NIR Hyperspectral Imaging. Agriculture 2023, 13, 237. https://doi.org/10.3390/agriculture13020237
Guo X, Jia B, Zhang H, Ni X, Zhuang H, Lu Y, Wang W. Evaluation of Aspergillus flavus Growth and Detection of Aflatoxin B1 Content on Maize Agar Culture Medium Using Vis/NIR Hyperspectral Imaging. Agriculture. 2023; 13(2):237. https://doi.org/10.3390/agriculture13020237
Chicago/Turabian StyleGuo, Xiaohuan, Beibei Jia, Haicheng Zhang, Xinzhi Ni, Hong Zhuang, Yao Lu, and Wei Wang. 2023. "Evaluation of Aspergillus flavus Growth and Detection of Aflatoxin B1 Content on Maize Agar Culture Medium Using Vis/NIR Hyperspectral Imaging" Agriculture 13, no. 2: 237. https://doi.org/10.3390/agriculture13020237