
Application Marker-Assisted Selection (MAS) and Multiplex PCR Reactions in Resistance Breeding of Maize (Zea mays L.)
 , , , ,
, , , ,  ,
,  , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Methodology
2.2.1. Field Experiment
2.2.2. DNA Isolation
2.2.3. Molecular Marker Analysis and PCR Amplification

2.2.4. Electrophoretic separation
2.3. Statistical Analysis
3. Results
3.1. Field Experiment
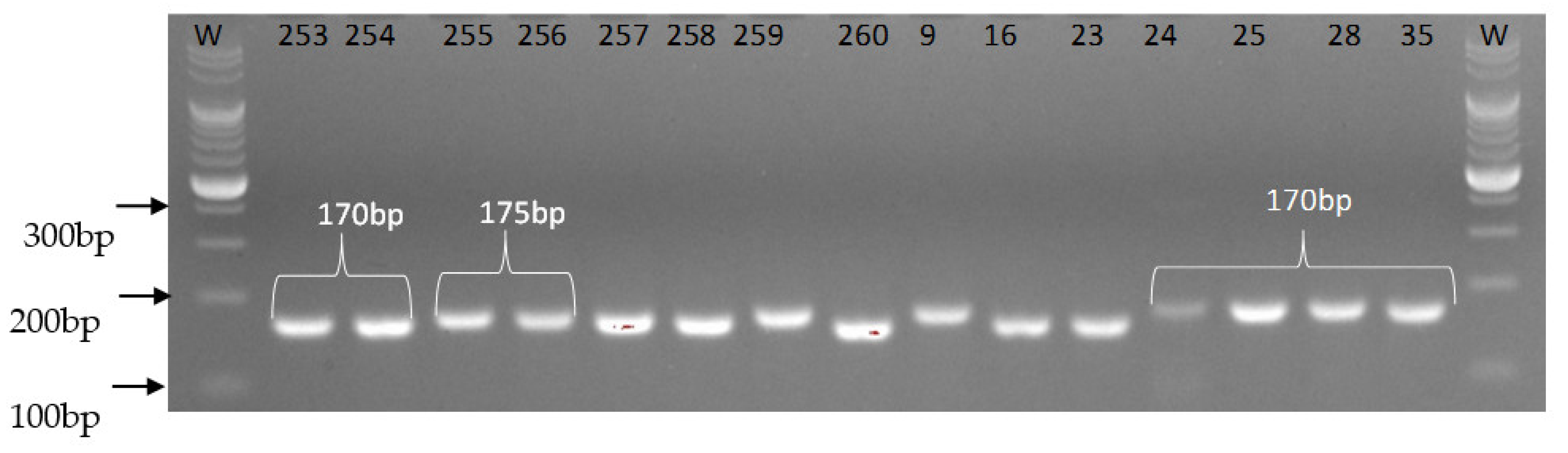
3.2. Identification of Molecular Markers Linked to Fusarium verticillioides Resistance Genes
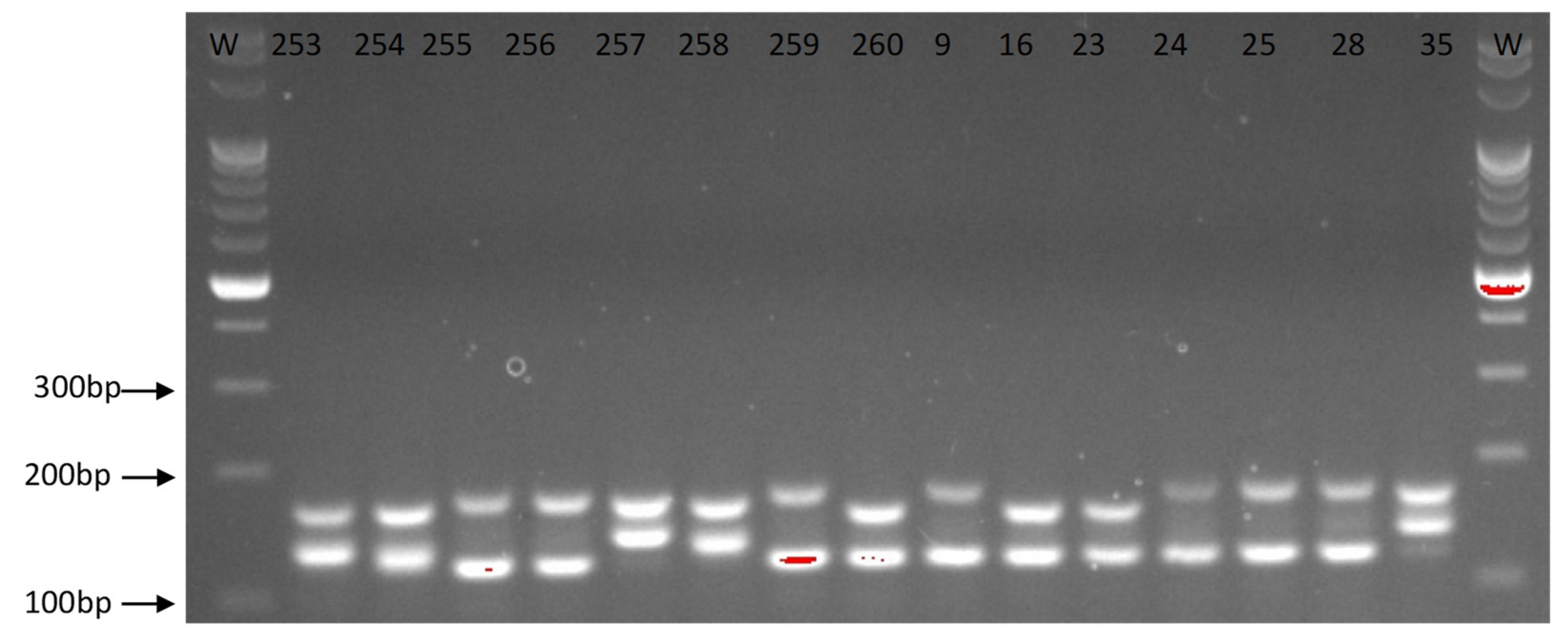
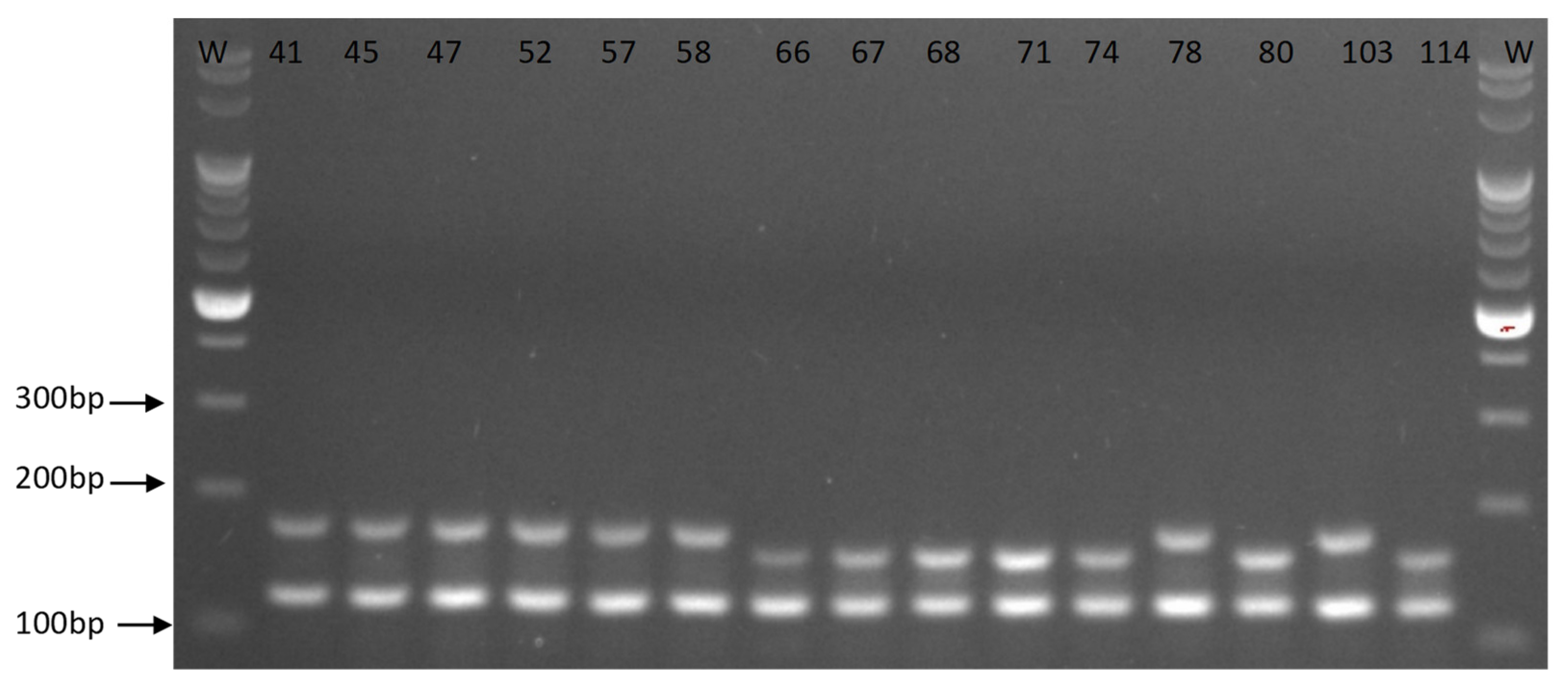
3.3. Multiplex PCR Analyses
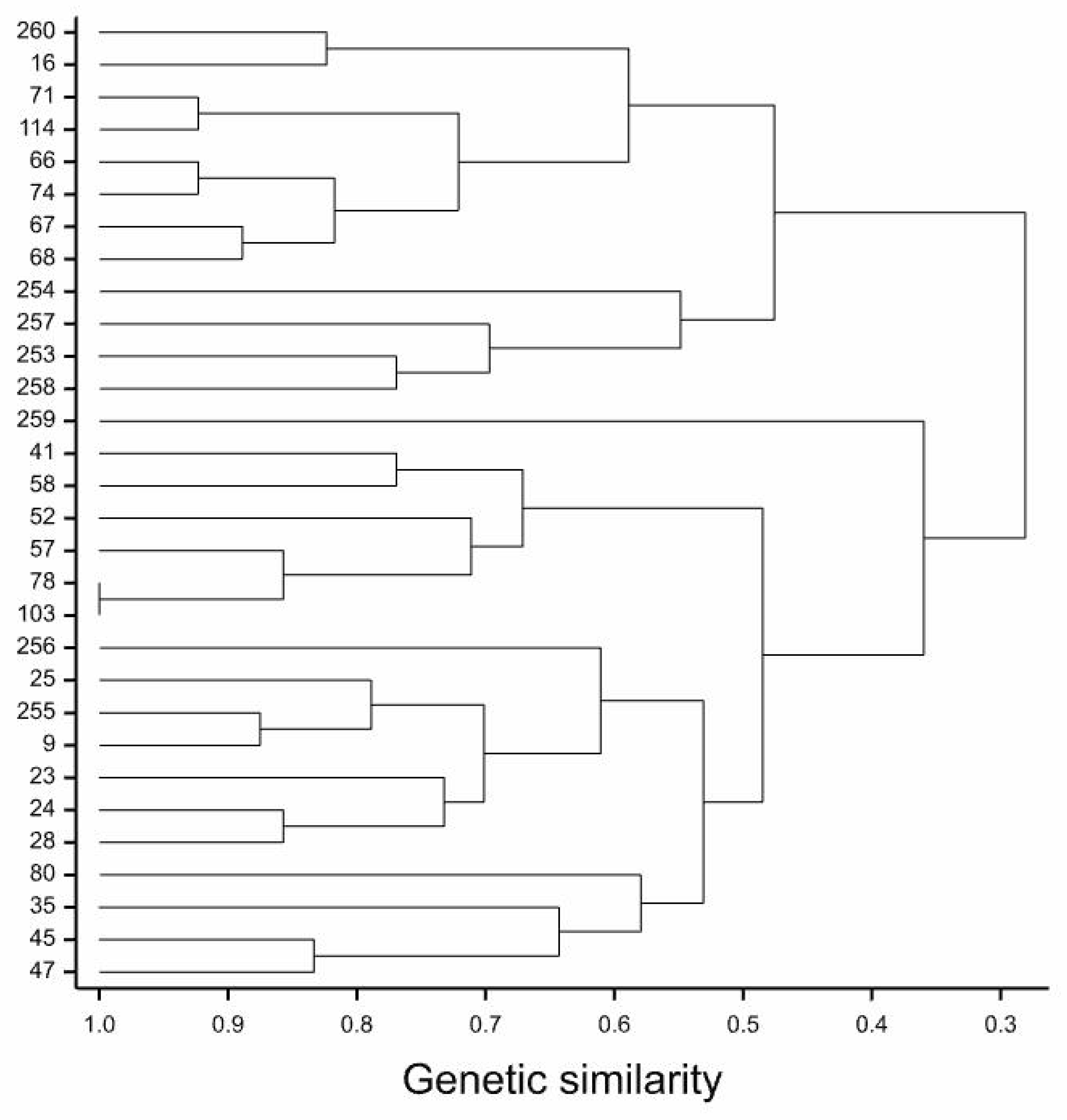
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Yadav, M.K.; Lipsa, D.; Sandeep, R. Evaluation of Host Plant Resistance Traits in Certain Genotypes of Maize (Zea Mays L.) against Maize Spotted Stem Borer (Chilo Partellus Swinhoe, Crambidae). Int. J. Entomol. Res. 2021, 6, 64–69. Available online: www.entomologyjournals.com (accessed on 1 May 2022).
- Butrón, A.; Santiago, R.; Cao, A.; Samayoa, L.F.; Malvar, R.A. QTLs for resistance to fusarium ear rot in a multiparent advanced generation intercross (MAGIC) maize population. Plant Dis. 2019, 103, 897–904. [Google Scholar] [CrossRef]
- Scauflaire, J.; Mahieu, O.; Louvieaux, J.; Foucart, G.; Renard, F.; Munaut, F. Biodiversity of Fusarium species in ears and stalks of maize plants in Belgium. Eur. J. Plant Pathol. 2011, 131, 59–66. [Google Scholar] [CrossRef]
- Logrieco, A.; Mule, G.; Moretti, A.; Bottalico, A. Toxigenic Fusarium species and mycotoxins associated with maize ear rot in Europe. In Mycotoxins in Plant Disease; Logrieco, A., Bailey, J.A., Corazza, L., Cooke, B.M., Eds.; Springer: Berlin/Heidelberg, Germany, 2002; pp. 597–609. ISBN 978-94-010-3939-0. [Google Scholar] [CrossRef]
- Eckard, S.; Wettstein, F.E.; Forrer, H.-R.; Vogelgsang, S. Incidence of Fusarium Species and Mycotoxins in Silage Maize. Toxins 2011, 3, 949–967. [Google Scholar] [CrossRef]
- Garcia-Ceron, D.; Lowe, R.G.T.; McKenna, J.A.; Brain, L.M.; Dawson, C.S.; Clark, B.; Berkowitz, O.; Faou, P.; Whelan, J.; Bleackley, M.R.; et al. Extracellular Vesicles from Fusarium graminearum Contain Protein Effectors Expressed during Infection of Corn. J. Fungi 2021, 7, 977. [Google Scholar] [CrossRef]
- Bush, B.J.; Carson, M.L.; Cubeta, M.A.; Hagler, W.M.; Payne, G.A. Infection and Fumonisin Production by Fusarium verticillioides in Developing Maize Kernels. Genet. Resist. Infect. Am. Phytopathol. Soc. 2003, 94, 88–93. [Google Scholar] [CrossRef]
- Uwe, L.; Miedaner, T.; Bürstmayr, H.; Vögele, R.T. Breeding for Resistance to Fusarium Ear Diseases in Maize and Small-Grain Cereals Using Genomic Tools; Faculty of Agricultural Sciences University of Hohenheim: Stuttgart, Germany, 2020; p. 144. Available online: http://opus.uni-hohenheim.de/volltexte/2021/1865/pdf/Gaikpa_Dissertation_phdKIM_print.pdf (accessed on 1 May 2022).
- Stagnati, L.; Lanubile, A.; Samayoa, L.F.; Bragalanti, M.; Giorni, P.; Busconi, M.; Holland, J.; Mrocco, B.A. A Genome Wide Association Study Reveals Markers and Genes Associated with Resistance to Fusarium Verticillioides Infection of Seedlings in a Maize Diversity Panel. G3 Genes Genomes Genet. 2019, 9, 571–579. [Google Scholar] [CrossRef] [PubMed]
- Salah, N.; Milad, S.I.; El-Rouby, M.M.; Barakat, M.N. Identification of New Molecular Markers Linked to Maize Stalk Rot Disease Resistance (Fusarium Moniliforme) in Maize. Plant Omics J. 2016, 9, 12–18. [Google Scholar] [CrossRef]
- Michel, S.; Wagner, C.; Nosenko, T.; Steiner, B.; Samad-Zamini, M.; Buerstmayr, M.; Mayer, K.; Buerstmayr, H. Merging Genomics and Transcriptomics for Predicting Fusarium Head Blight Resistance in Wheat. Genes 2021, 12, 114. [Google Scholar] [CrossRef] [PubMed]
- Maschietto, V.; Colombi, C.; Pirona, R.; Pea, G.; Strozzi, F.; Marocco, A.; Rossini, L.; Lanubile, A. QTL mapping and candidate genes for resistance to Fusarium ear rot and fumonisin contamination in maize. BMC Plant Biol. 2017, 17, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdel-Rahman, M.M.; Bayoumi, S.R.; Barakat, M.N. Identification of molecular markers linked to Fusarium ear rot genes in maize plants Zea mays L. Biotechnol. Biotechnol. Equip. 2016, 30, 692–699. [Google Scholar] [CrossRef]
- Jones, E.S.; Sullivan, H.; Bhattramakki, D.; Smith, J.S.C. A comparison of simple sequence repeat and single nucleotide polymorphism marker technologies for the genotypic analysis of maize (Zea mays L.). Theor. Appl. Genet. 2007, 115, 361–371. [Google Scholar] [CrossRef] [PubMed]
- Hamblin, M.T.; Fernandez, M.G.S.; Tuinstra, M.R.; Rooney, W.L.; Kresovich, S. Sequence Variation at Candidate Loci in the Starch Metabolism Pathway in Sorghum: Prospects for Linkage Disequilibrium Mapping. Crop Sci. 2007, 47, 125–134. [Google Scholar] [CrossRef]
- Van Inghelandt, D.; Melchinger, A.E.; Lebreton, C.; Stich, B. Population structure and genetic diversity in a commercial maize breeding program assessed with SSR and SNP markers. Theor. Appl. Genet. 2010, 120, 1289–1299. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Zhong, W.; Yang, F.; Zhang, Z. Genetic and Molecular Mechanisms of Quantitative Trait Loci Controlling Maize Inflorescence Architecture. Plant Cell Physiol. 2018, 59, 448–457. [Google Scholar] [CrossRef] [PubMed]
- Arabzai, M.; Gul, H. Application Techniques of Molecular Marker and Achievement of Marker Assisted Selection (MAS ) in Three Major Crops Rice, Wheat and Maize. Int. J. Res. Appl. Sci. Biotechnol. 2021, 8, 82–93. [Google Scholar] [CrossRef]
- Nei, M.; Li, W.H. Mathematical model for studying genetic variation in terms of restriction endonucleases. Proc. Natl. Acad. Sci. USA 1979, 76, 5269–5273. [Google Scholar] [CrossRef]
- Starzycka-Korbas, E.; Weber, Z.; Matuszczak, M.; Bocianowski, J.; Budzianowski, G.; Stefanowicz, M.; Starzycki, M. The diversity of Sclerotinia sclerotiorum (Lib.) de Bary isolates from western Poland. J. Plant Pathol. 2021, 103, 185–195. [Google Scholar] [CrossRef]
- Bocianowski, J.; Kozak, M.; Liersch, A.; Bartkowiak-Broda, I. A heuristic method of searching for interesting markers in terms of quantitative traits. Euphytica 2011, 181, 89–100. [Google Scholar] [CrossRef]
- Bottalico, A.; Perrone, G. Toxigenic Fusarium species and mycotoxins associated with head blight in small-grain cereals in Europe. Eur. J. Plant Pathol. 2002, 108, 611–624. [Google Scholar] [CrossRef]
- Desjardins, A.E.; Proctor, R.H. Molecular biology of Fusarium mycotoxins. Int. J. Food Microbiol. 2007, 119, 47–50. [Google Scholar] [CrossRef]
- Pitt, J.I. Toxigenic fungi: Which are important? Med. Mycol. 2000, 38, 17–22. [Google Scholar] [CrossRef] [PubMed]
- Santiago, R.; Cao, A.; Malvar, R.A.; Butrón, A. Genomics of Maize Resistance to Fusarium Ear Rot and Fumonisin Contamination. Toxins 2020, 12, 431. [Google Scholar] [CrossRef] [PubMed]
- Robertson, L.A.; Kleinschmidt, C.E.; White, D.G.; Payne, G.A.; Maragos, C.M.; Holland, J.B. Heritability and correlations of Fusarium ear rot resistance and fumonisin contamination resistance in two maize populations. Crop Sci. 2006, 46, 353–361. [Google Scholar] [CrossRef]
- Ding, J.Q.; Wang, X.M.; Chander, S.; Yan, J.E.; Li, J.S. QTL mapping of resistance to Fusarium ear rot using a RIL population in maize. Mol. Breed. 2008, 22, 395–403. [Google Scholar] [CrossRef]
- Chen, J.; Ding, J.; Li, H.; Li, Z.; Sun, X.; Li, J.; Wang, R.; Dai, X.; Dong, H.; Song, W.; et al. Detection and verification of quantitative trait loci for resistance to Fusarium ear rot in maize. Mol. Breed. 2012, 30, 1649–1656. [Google Scholar] [CrossRef]
- Gaikpa, D.S.; Miedaner, T. Genomics-assisted breeding for ear rot resistances and reduced mycotoxin contamination in maize: Methods, advances and prospects. Theor. Appl. Genet. 2019, 132, 2721–2739. [Google Scholar] [CrossRef]
- Baldwin, T.; Baldwin, S.; Klos, K.; Bregitzer, P.; Marshall, J. Deletion of the benzoxazinoid detoxification gene NAT1 in Fusarium graminearum reduces deoxynivalenol in spring wheat. PLoS ONE 2019, 14, e0214230. [Google Scholar] [CrossRef]
- Cotton, T.E.A.; Pétriacq, P.; Cameron, D.D.; Al Meselmani, M.; Schwarzenbacher, R.; Rolfe, S.A.; Ton, J. Metabolic regulation of the maize rhizobiome by benzoxazinoids. ISME J. 2019, 13, 1647–1658. [Google Scholar] [CrossRef]
- Park, J.S.; Park, J.Y.; Park, K.J.; Lee, J.K. Genetic diversity among waxy corn accessions in Korea revealed by microsatellite markers. Korean J. Breed. Sci. 2008, 40, 250–257. [Google Scholar]
- Sa, K.J.; Hong, T.K.; Lee, J.K. Genetic diversity and association analyses of canadian maize inbred lines with agronomic traits and simple sequence repeat markers. Plant Breed. Biotechnol. 2018, 6, 159–169. [Google Scholar] [CrossRef]
- Ignjatović-Micić, D.; Drinić, S.M.; Nikolić, A.; Lazić-Jančić, V. SSR analysis for genetic structure and diversity determination of maize local populations from former Yugoslavia territories. Russ. J. Genet. 2008, 44, 1317–1324. [Google Scholar] [CrossRef]
- Ashkani, S.; Rafii, M.Y.; Shabanimofrad, M.; Foroughi, M.; Azizia, P.; Akhtar, M.S.; Sahebi, M.; Harun, A.R.; Nasehi, A. Multiplex SSR–PCR approaches for semi-automated genotyping and characterization of loci linked to blast disease resistance genes in rice. Comptes Rendus Biol. 2015, 388, 709–722. [Google Scholar] [CrossRef]
- Bocianowski, J.; Nowosad, K.; Wróbel, B.; Szulc, P. Identification of Associations between SSR Markers and Quantitative Traits of Maize (Zea mays L.). Agronomy 2021, 11, 182. [Google Scholar] [CrossRef]
- Cyplik, A.; Sobiech, A.; Tomkowiak, A.; Bocianowski, J. Genetic Parameters for Selected Traits of Inbred Lines of Maize (Zea mays L.). Appl. Sci. 2022, 12, 6961. [Google Scholar] [CrossRef]
- Sobiech, A.; Tomkowiak, A.; Nowak, B.; Bocianowski, J.; Wolko, Ł.; Spychała, J. Associative and Physical Mapping of Markers Related to Fusarium in Maize Resistance, Obtained by Next-Generation Sequencing (NGS). Int. J. Mol. Sci. 2022, 23, 6105. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Resistant Genotypes | Susceptible Genotypes | ||
|---|---|---|---|
| No. | Genotype Number | No. | Genotype Number |
| 1 | 9 | 16 | 16 |
| 2 | 25 | 17 | 23 |
| 3 | 28 | 18 | 24 |
| 4 | 35 | 19 | 41 |
| 5 | 45 | 20 | 57 |
| 6 | 47 | 21 | 58 |
| 7 | 52 | 22 | 67 |
| 8 | 66 | 23 | 68 |
| 9 | 71 | 24 | 78 |
| 10 | 74 | 25 | 103 |
| 11 | 80 | 26 | 253 |
| 12 | 114 | 27 | 254 |
| 13 | 255 | 28 | 256 |
| 14 | 257 | 29 | 258 |
| 15 | 260 | 30 | 259 |
| Polymorphism | Marker | Primer Sequences | Product Size (bp) | Melting Temperature (°C) | Reference | |
|---|---|---|---|---|---|---|
| Rewers | Forward | |||||
| bnlg1621 | GGATCTTCGTTGCAGTTCTT | CATCAGTGATCCTCCACCAT | 135–160 | 54 | [13] | |
| SSR | bnlg1063 | GGAGACAACCCCGACGAC | GGTACCAGAGCCACAGATCC | 105–120 | 55 | [13] |
| bnlg1740 | TTTTCTCCTTGAGTTCGTTCG | ACAGGCAGAGCTCTCACACA | 125–160 | 56 | [13] | |
| umc2082 | TAGCTGCCCCTCTTCCGTCT | GTCGTGGCGTAGAGACTAGGGT | 100–130 | 54 | [13] | |
| umc2059 | CTCTTCGATCTTTAAGAGAGAGAGAG | ACACGAGGCACTGGTACTAACG | 170–200 | 54 | [13] | |
| umc2013 | GGAAAAGGAGGAACAGTGTAAGCA | AGCGTGATCAGACGTACAATGCTA | 110–130 | 54 | [13] | |
| SSR85 | GGGACGAGAGTCTGTTGTTGTTG | GTTGATGCATGTGACTCTGGAAAC | 110–125 | 55 | [13] | |
| SSR93 | CGCCGTACAGACTGCTATGA | CACATGCTACGACTGCGATG | 210 | 57 | [10] | |
| SSR105 | GTTCATCCTGATTCCCATCC | CAGCCTTGCTTCTACACCAC | 200 | 58 | [10] | |
| SSR255 | TCGACGAGATACGCGACTAC | CAGTACAAAGCCGATCCAAG | 200 | 55 | [10] | |
| ISSR | AD8 | (AGC)6GC | 410 | 55 | [10] | |
| STS | STS03 | CTTGTATCATCAGCTAGGGCATGT | GTGATCTGAACGCCAACCTC | 300 | 54 | [10] |
| Line No. | Observation of the Degree of Resistance | Number of Molecular Markers Identified | ||
|---|---|---|---|---|
| I Repetition | II Repetition | III Repetition | ||
| 253 | 7 | 7 | 8 | 6 |
| 254 | 4 | 5 | 4 | 5 |
| 255 | 9 | 9 | 9 | 9 |
| 256 | 5 | 7 | 6 | 7 |
| 257 | 5 | 5 | 6 | 6 |
| 258 | 7 | 8 | 7 | 7 |
| 259 | 7 | 7 | 7 | 8 |
| 260 | 9 | 9 | 8 | 9 |
| 9 | 9 | 9 | 8 | 9 |
| 16 | 7 | 6 | 7 | 7 |
| 23 | 6 | 6 | 7 | 8 |
| 24 | 6 | 5 | 4 | 6 |
| 25 | 9 | 8 | 9 | 10 |
| 28 | 9 | 8 | 8 | 8 |
| 35 | 8 | 8 | 7 | 9 |
| 41 | 8 | 7 | 6 | 7 |
| 45 | 7 | 7 | 8 | 8 |
| 47 | 8 | 7 | 8 | 8 |
| 52 | 9 | 8 | 8 | 8 |
| 57 | 5 | 6 | 5 | 7 |
| 58 | 6 | 6 | 5 | 7 |
| 66 | 8 | 7 | 7 | 8 |
| 67 | 4 | 5 | 6 | 6 |
| 68 | 5 | 7 | 4 | 7 |
| 71 | 8 | 8 | 9 | 8 |
| 74 | 8 | 6 | 5 | 8 |
| 78 | 6 | 8 | 6 | 7 |
| 80 | 6 | 6 | 7 | 8 |
| 103 | 8 | 6 | 6 | 7 |
| 114 | 9 | 8 | 9 | 8 |
| Line No. | Molecular Marker | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| SSR85 | bngl1063 | bngl1740 | umc2082 | bngl1621 | umc2059 | umc2013 | SSR93 | SSR105 | SSR255 | AD8 | STS03 | |
| 253 | - | + | + | + | + | + | + | - | - | - | - | - |
| 254 | - | + | + | + | + | + | - | - | - | - | - | - |
| 255 | + | + | + | + | + | + | + | - | + | - | - | + |
| 256 | - | + | + | + | + | + | + | + | - | - | - | - |
| 257 | - | + | + | + | + | + | + | - | - | - | - | - |
| 258 | - | + | + | + | + | + | + | - | - | + | - | - |
| 259 | - | - | + | + | + | + | + | - | + | - | + | + |
| 260 | + | - | + | + | + | + | + | - | + | + | - | + |
| 9 | + | + | + | + | + | + | + | - | + | - | - | + |
| 16 | + | - | + | + | + | + | - | - | + | - | - | + |
| 23 | + | - | + | + | + | + | + | - | + | - | - | + |
| 24 | + | - | + | + | + | + | + | - | - | - | - | - |
| 25 | + | + | + | + | + | + | + | + | + | - | - | + |
| 28 | + | - | + | + | + | + | + | + | - | - | - | + |
| 35 | + | + | + | + | + | + | + | + | - | - | - | + |
| 41 | + | - | + | + | + | + | + | - | - | + | - | - |
| 45 | + | - | + | + | + | + | + | - | + | - | - | + |
| 47 | + | - | + | + | + | + | + | - | + | - | - | + |
| 52 | + | + | + | + | + | + | + | - | - | + | - | - |
| 57 | + | - | + | + | + | + | + | - | - | + | - | - |
| 58 | + | + | + | + | + | + | + | - | - | - | - | - |
| 66 | + | + | + | + | + | + | + | - | - | + | - | - |
| 67 | + | - | + | + | + | + | + | - | - | - | - | - |
| 68 | + | - | + | + | + | + | + | - | - | + | - | - |
| 71 | + | + | + | + | + | + | + | - | - | + | - | - |
| 74 | + | + | + | + | + | + | + | - | - | + | - | - |
| 78 | + | - | + | + | + | + | + | + | - | - | - | - |
| 80 | - | + | + | + | + | + | + | - | + | - | - | + |
| 103 | + | - | + | + | + | + | + | + | - | - | - | - |
| 114 | + | + | + | + | + | + | + | - | - | + | - | - |
| Marker | Effect | Percentage Variance Accounted |
|---|---|---|
| SSR85 | 0.921 *** | 9.9 |
| bngl1063 | 0.565 *** | 7.4 |
| bngl1740 125 bp | −0.093 * | 1.5 |
| bngl1740 155 bp | −0.432 *** | 6.7 |
| bngl1740 160 bp | 0.518 *** | 2.8 |
| umc2082 100 bp | 0.503 *** | 3.4 |
| umc2082 115 bp | 0.067 * | 1.2 |
| umc2082 120 bp | 0.164 *** | 4.7 |
| bngl1621 170 bp | −0.521 *** | 0.7 |
| bngl1621 175 bp | 0.521 *** | 0.7 |
| umc2059 110 bp | 0.694 ** | 1.8 |
| umc2059 120 bp | −2.7 ** | 11.2 |
| umc2059 125 bp | 0.417 * | 7.5 |
| umc2059 130 bp | −0.476 ** | 5.5 |
| umc2013 120 bp | −0.224 * | 2.8 |
| umc2013 125 bp | −0.093 * | 1.5 |
| SSR93 | 0.486 ** | 3.6 |
| SSR105 | 1.033 *** | 11.3 |
| SSR255 | 0.483 *** | 5.7 |
| AD8 | 0.06 | |
| STS03 | 1.25 *** | 19.9 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sobiech, A.; Tomkowiak, A.; Bocianowski, J.; Nowak, B.; Weigt, D.; Kurasiak-Popowska, D.; Kwiatek, M.; Mikołajczyk, S.; Niemann, J.; Szewczyk, K. Application Marker-Assisted Selection (MAS) and Multiplex PCR Reactions in Resistance Breeding of Maize (Zea mays L.). Agriculture 2022, 12, 1412. https://doi.org/10.3390/agriculture12091412
Sobiech A, Tomkowiak A, Bocianowski J, Nowak B, Weigt D, Kurasiak-Popowska D, Kwiatek M, Mikołajczyk S, Niemann J, Szewczyk K. Application Marker-Assisted Selection (MAS) and Multiplex PCR Reactions in Resistance Breeding of Maize (Zea mays L.). Agriculture. 2022; 12(9):1412. https://doi.org/10.3390/agriculture12091412
Chicago/Turabian StyleSobiech, Aleksandra, Agnieszka Tomkowiak, Jan Bocianowski, Bartosz Nowak, Dorota Weigt, Danuta Kurasiak-Popowska, Michał Kwiatek, Sylwia Mikołajczyk, Janetta Niemann, and Katarzyna Szewczyk. 2022. "Application Marker-Assisted Selection (MAS) and Multiplex PCR Reactions in Resistance Breeding of Maize (Zea mays L.)" Agriculture 12, no. 9: 1412. https://doi.org/10.3390/agriculture12091412