
The Potential Function of SiLOX4 on Millet Discoloration during Storage in Foxtail Millet

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Experimental Site and Field Experiment
2.3. The Total Carotenoid Content (TCC) Analysis
2.4. Chemicals and Solvents
2.5. HPLC Analysis of Lutein and Zeaxanthin
2.6. Western-Blot Analysis
2.7. Isolation and Sequence Retrieval of LOX Gene Family from Foxtail Millet
2.8. RNA Extraction and Real-Time PCR Analysis
3. Results
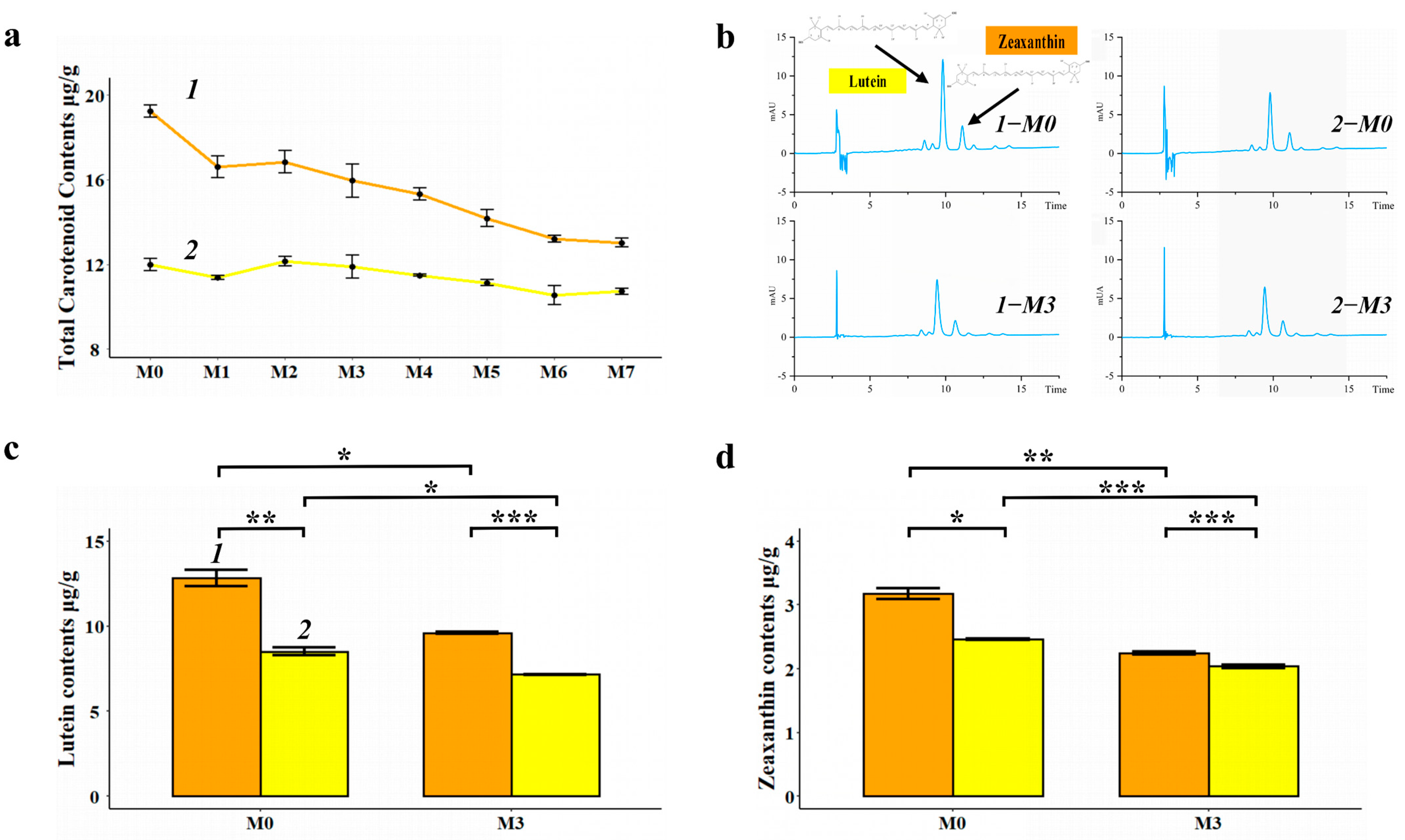
3.1. Changes in the Content and Composition of Carotenoid in the Two Varieties of Foxtail Millet during Storage
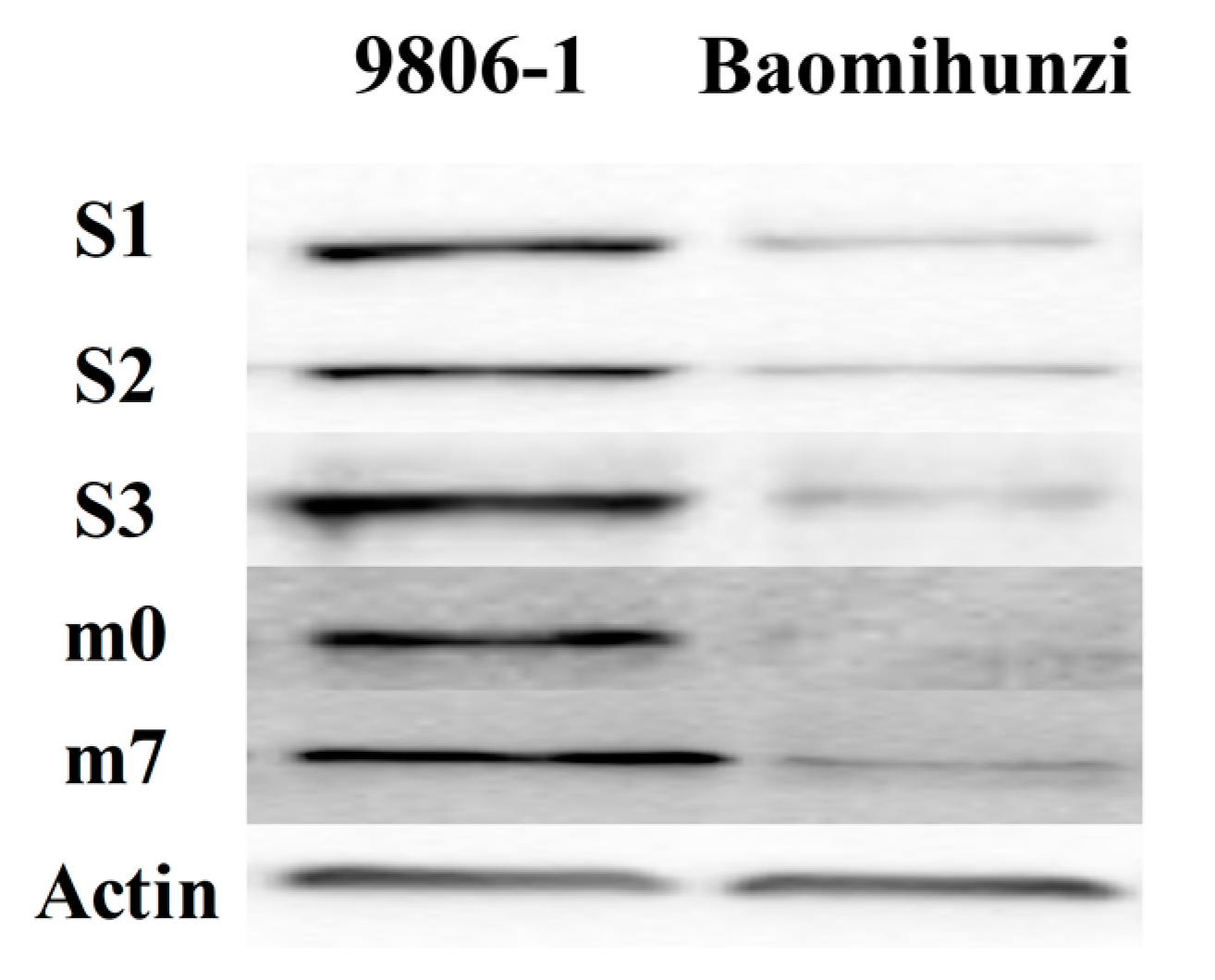
3.2. Expression Patterns of SiLOX Protein in the Two Millet Varieties
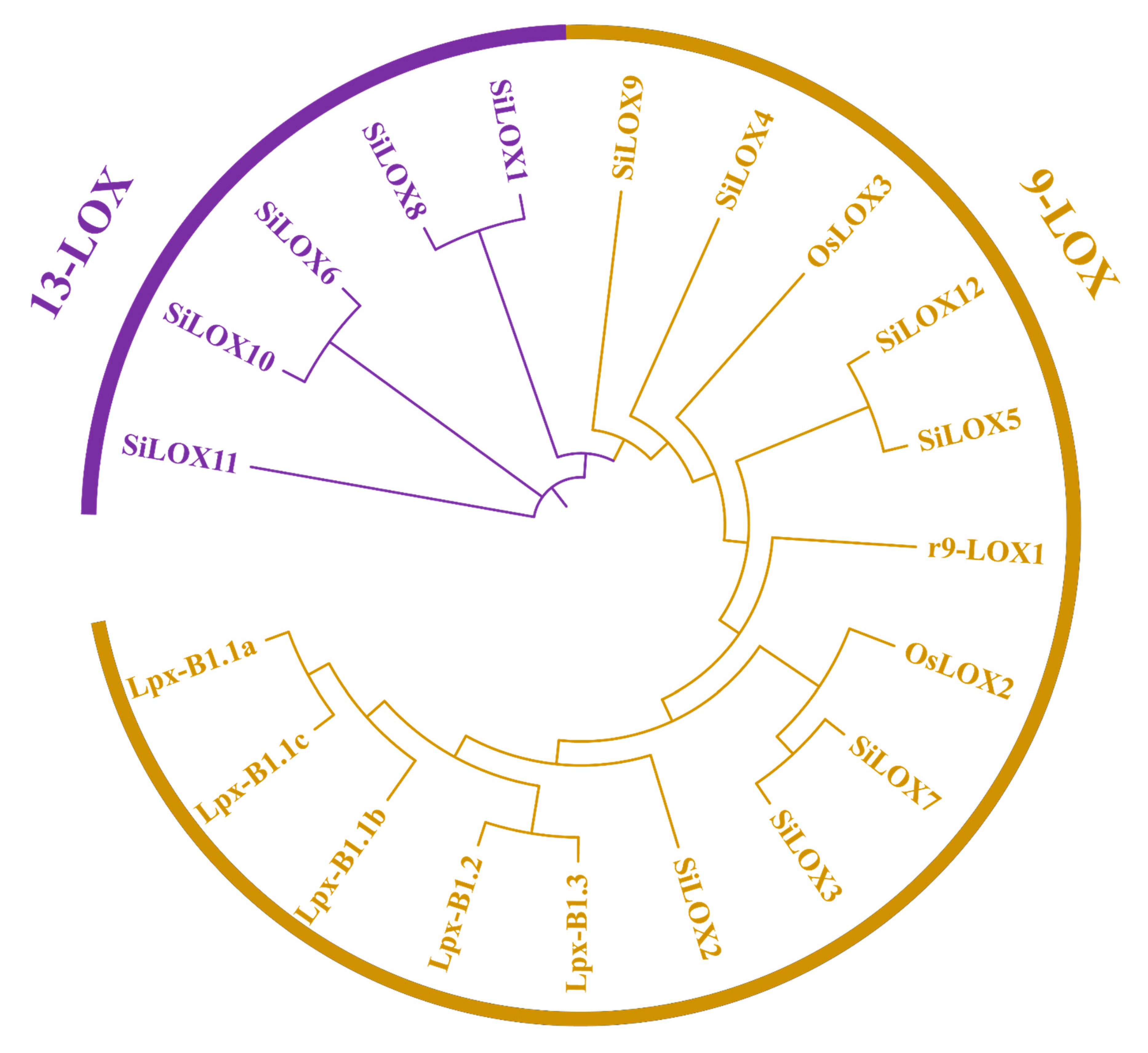
3.3. Phylogenetic Analysis of SiLOX Genes in Foxtail Millet
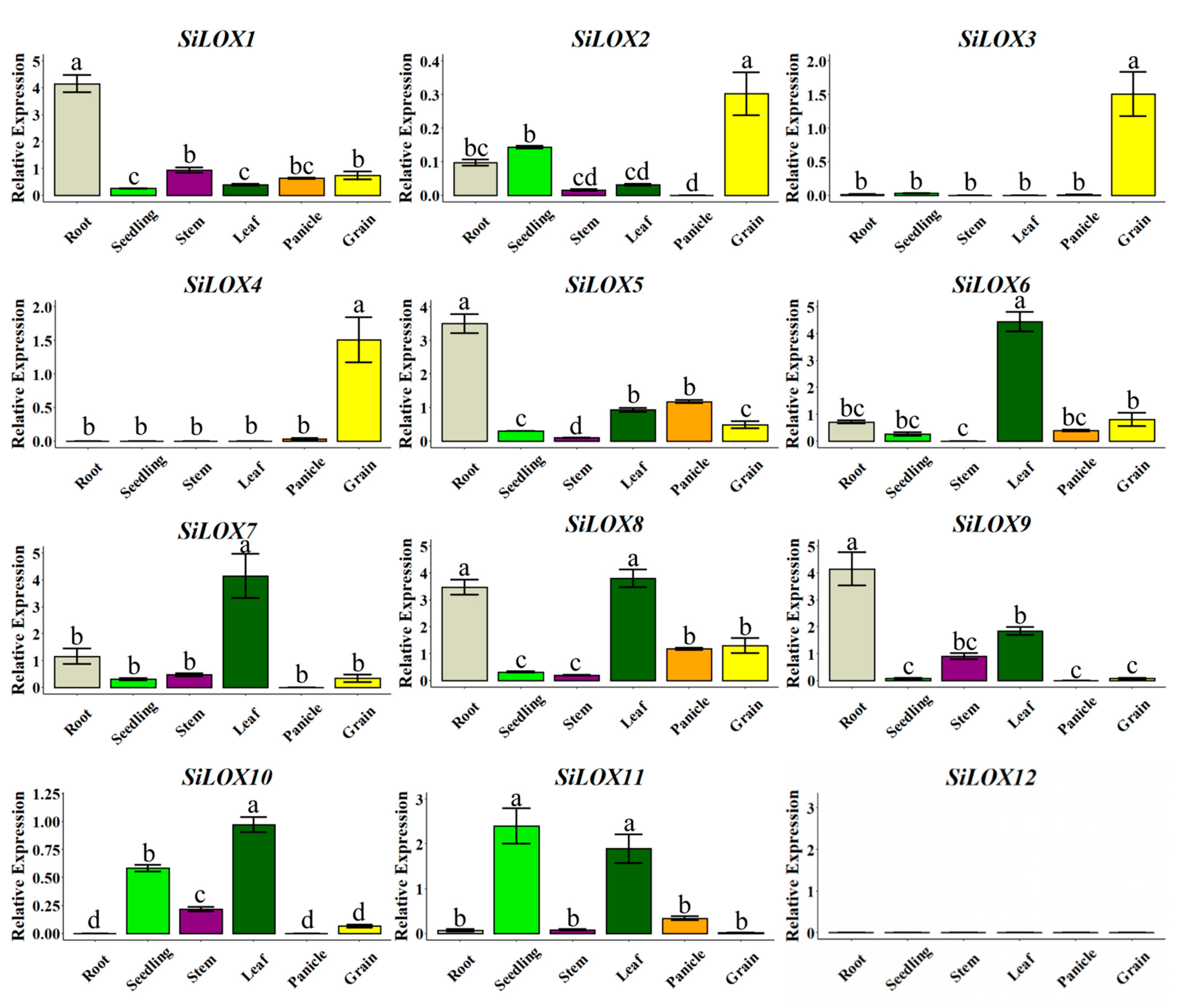
3.4. Tissue-Specific Expression Patterns of SiLOX Genes
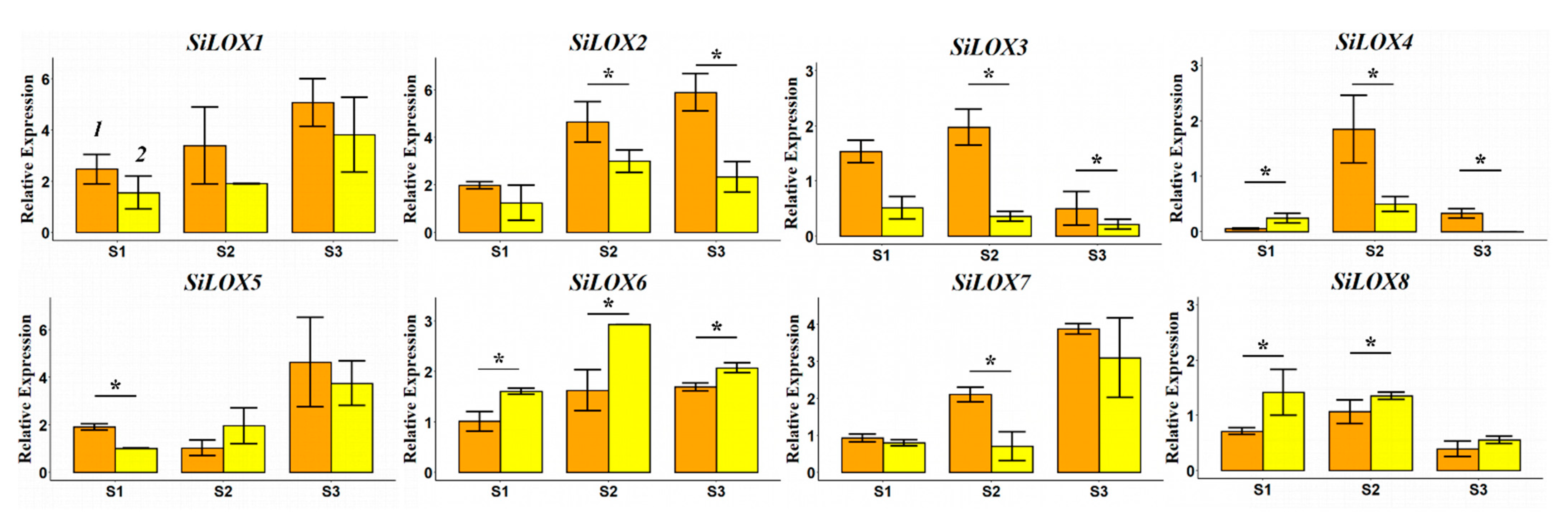
3.5. Expression Patterns of SiLOX Genes during Grain Maturation
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Tanaka, Y.; Sasaki, N.; Ohmiya, A. Biosynthesis of plant pigments: Anthocyanins, betalains and carotenoids. Plant J. 2008, 54, 733–749. [Google Scholar] [CrossRef]
- Howitt, C.A.; Pogson, B.J. Carotenoid accumulation and function in seeds and non-green tissues. Plant Cell Environ. 2006, 29, 435–445. [Google Scholar] [CrossRef] [PubMed]
- Ficco, D.; Mastrangelo, A.M.; Trono, D.; Borrelli, G.M.; Vita, P.D.; Fares, C.; Beleggia, R.; Platani, C.; Papa, R. The colours of durum wheat: A review. Crop Pasture Sci. 2014, 65, 1–15. [Google Scholar] [CrossRef]
- Botella-Pavía, P.; Rodríguez-Concepción, M. Carotenoid biotechnology in plants for nutritionally improved foods. Physiol. Plant. 2010, 126, 369–381. [Google Scholar] [CrossRef]
- Sun, T.H.; Yuan, H.; Cao, H.B.; Yazdani, M.; Tadmor, Y.; Li, L. Carotenoid metabolism in plants: The role of plastids. Mol. Plant 2018, 11, 58–74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ceasar, S.A.; Baker, A.; Ignacimuthu, S. Functional characterization of the PHT1 family transporters of foxtail millet with development of a novel Agrobacterium-mediated transformation procedure. Sci. Rep. 2017, 7, 14064. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, L.; Zhang, B.; Wang, X.; Li, H.Y.; Han, Y.H. Foxtail millet: Nutritional and eating quality, and prospects for genetic improvement. Front. Agr. Sci. Eng. 2015, 2, 124–133. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.S.; Wang, L.L.; Zhou, X.R.; Shuang, S.M.; Zhu, Z.H.; Li, N.; Li, Y.; Liu, F.; Liu, S.C.; Lu, P. Determination of protein, fat, starch, and amino acids in foxtail millet [Setaria italica (L.) Beauv.] by fourier transform near-infrared reflectance spectroscopy. Food Sci. Biotechnol. 2013, 22, 1495–1500. [Google Scholar] [CrossRef]
- He, L.; Cheng, L.; Wang, J.J.; Liu, J.; Cheng, J.J.; Yang, Z.R.; Cao, R.; Han, Y.H.; Li, H.Y.; Zhang, B. Carotenoid Cleavage Dioxygenase 1 Catalyzes Lutein Degradation To Influence Carotenoid Accumulation and Color Development in Foxtail Millet Grains. J. Agric. Food Chem. 2022, 70, 9283–9294. [Google Scholar] [CrossRef]
- Shen, R.; Yang, S.P.; Zhao, G.H.; Shen, Q.; Diao, X.M. Identification of carotenoids in foxtail millet (Setaria italica) and the effects of cooking methods on carotenoid content. J. Cereal Sci. 2015, 61, 86–93. [Google Scholar] [CrossRef]
- Leenhardt, F.; Lyan, B.; Rock, E.; Boussard, A.; Potus, J.; Chanliaud, E.; Remesy, C. Genetic variability of carotenoid concentration, and lipoxygenase and peroxidase activities among cultivated wheat species and bread wheat varieties. Eur. J. Agron. 2006, 25, 170–176. [Google Scholar] [CrossRef]
- Zhang, Y.; Yu, Z.L.; Lu, Y.X.; Wang, Y.; She, D.; Song, M.; Wu, Y.J. Effect of the absence of lipoxygenase isoenzymes on the storage characteristics of rice grains. J. Stored Prod. Res. 2007, 43, 87–91. [Google Scholar] [CrossRef]
- Sun, Y.N.; Wang, K.; Dong, Y.H.; Li, K.K.; Liu, H.H.; Cui, B.Z.; Fu, H.F.; Chen, X.W.; Wang, Y.Q.; Wang, Y.Y. Effects of radiofrequency blanching on lipoxygenase inactivation, physicochemical properties of sweet corn (Zea mays L.), and its correlation with cell morphology. Food Chem. 2022, 394, 133498. [Google Scholar] [CrossRef] [PubMed]
- Brash, A.R. Lipoxygenases: Occurrence, functions, catalysis, and acquisition of substrate. J. Biol. Chem. 1999, 274, 23679–23682. [Google Scholar] [CrossRef] [Green Version]
- Shen, J.; Tieman, D.; Jones, J.B.; Taylor, M.G.; Schmelz, E.; Huffaker, A.; Bies, D.; Chen, K.; Klee, H.J. A 13-lipoxygenase, TomloxC, is essential for synthesis of C5 flavour volatiles in tomato. J. Exp. Bot. 2014, 65, 419–428. [Google Scholar] [CrossRef] [Green Version]
- Shrestha, K.; Pant, S.; Huang, Y. Genome-wide identification and classification of Lipoxygenase gene family and their roles in sorghum-aphid interaction. Plant Mol. Biol. 2021, 105, 527–541. [Google Scholar] [CrossRef]
- Wilson, R.A.; Gardner, H.W.; Keller, N.P. Cultivar-dependent expression of a maize lipoxygenase responsive to seed infesting fungi. Mol. Plant Microbe Interact. 2001, 14, 980–987. [Google Scholar] [CrossRef] [Green Version]
- Long, Q.; Zhang, W.; Wang, P.; Shen, W.B.; Zhou, T.; Liu, N.N.; Wang, R.; Jiang, L.; Huang, J.X.; Wang, Y.H.; et al. Molecular genetic characterization of rice seed lipoxygenase 3 and assessment of its effects on seed longevity. J. Plant Biol. 2013, 56, 232–242. [Google Scholar] [CrossRef]
- Ma, L.; Zhu, F.G.; Li, Z.W.; Zhang, J.F.; Li, X.; Dong, J.L.; Wang, T. TALEN-Based mutagenesis of lipoxygenase LOX3 enhances the storage tolerance of rice (Oryza sativa) seeds. PLoS ONE. 2015, 10, e0143877. [Google Scholar] [CrossRef]
- Gayen, D.; Ali, N.; Sarkar, S.N.; Datta, S.K.; Datta, K. Down-regulation of lipoxygenase gene reduces degradation of carotenoids of golden rice during storage. Planta 2015, 242, 353–363. [Google Scholar] [CrossRef]
- Carrera, A.; Echenique, V.; Zhang, W.; Helguera, M.; Manthey, F.; Schrager, A.; Picca, A.; Cervigni, G.; Dubcovsky, J. A deletion at the Lpx-B1 locus is associated with low lipoxygenase activity and improved pasta color in durum wheat (Triticum turgidum ssp. durum). J. Cereal Sci. 2007, 45, 67–77. [Google Scholar] [CrossRef] [Green Version]
- Zhang, B.; Liu, J.; Cheng, L.; Zhang, Y.Y.; Hou, S.Y.; Sun, Z.X.; Li, H.Y.; Han, Y.H. Carotenoid composition and expression of biosynthetic genes in yellow and white foxtail millet [Setaria italica (L.) Beauv]. J. Cereal Sci. 2019, 85, 84–90. [Google Scholar] [CrossRef]
- Quinton, L.A.; Kennedy, J.F. American association of cereal chemists approved methods, 10th ed. CD-ROM. Carbohydr. Polym. 2002, 49, 515–517. [Google Scholar] [CrossRef]
- Kurien, B.T.; Scofield, R.H. Western blotting. Methods 2006, 38, 283–293. [Google Scholar] [CrossRef]
- Noguchi, A.; Nakamura, K.; Sakata, K.; Sato-Fukuda, N.; Ishigaki, T.; Mano, J.; Takabatake, R.; Kitta, K.; Teshima, R.; Kondo, K.; et al. Development and Interlaboratory Validation of a Simple Screening Method for Genetically Modified Maize Using a ∆∆C(q)-Based Multiplex Real-Time PCR Assay. Anal. Chem. 2016, 88, 4285–4293. [Google Scholar] [CrossRef] [PubMed]
- Barton, L.; Newsome, S.D.; Chen, F.H.; Wang, H.; Guilderson, T.P.; Bettinger, R.L. Agricultural origins and the isotopic identity of domestication in northern China. Proc. Natl. Acad. Sci. USA 2009, 106, 5523–5528. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.K.; Tang, X.; Zhang, Y.Z.; Zhao, W. Determination of the volatile composition in brown millet, milled millet and millet bran by gas chromatography/mass spectrometry. Molecules 2012, 17, 2271–2282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boon, C.S.; McClements, D.J.; Weiss, J.; Decker, E.A. Factors influencing the chemical stability of carotenoids in foods. Crit. Rev. Food Sci. Nutr. 2010, 50, 515–532. [Google Scholar] [CrossRef]
- Ngamwonglumlert, L.; Devahastin, S.; Chiewchan, N.; Raghavan, V. Plant carotenoids evolution during cultivation, postharvest storage, and food processing: A review. Compr. Rev. Food Sci. Food Saf. 2020, 19, 1561–1604. [Google Scholar] [CrossRef]
- Zhang, M.K.; Zhang, M.P.; Mazourek, M.; Tadmor, Y.; Li, L. Regulatory control of carotenoid accumulation in winter squash during storage. Planta 2014, 240, 1063–1074. [Google Scholar] [CrossRef]
- Zhao, J.J.; Zhou, Z.X.; Zhu, M.; Zhao, L.; Gao, A.P.; Chen, Y.Y.; Wang, J.B.; University, H. Changes of carotenoids during dynamic development and ripening of mango fruits. Xi Nan Nong Ye Xue Bao. 2014, 27, 2124–2130. [Google Scholar]
- Dua, P.N.; Day, E.J.; Grogan, C.O. Loss of carotenoids in stored commercial and high-carotenoid yellow corn, Zea mays L. Agron. J. 1965, 57, 501–502. [Google Scholar] [CrossRef]
- Chedea, V.S.; Jisaka, M. Lipoxygenase and carotenoids: A co-oxidation story. Afr. J. Biotechnol. 2013, 12, 2786–2791. [Google Scholar]
- Wu, Z.; Robinson, D.S.; Domoney, C.; Casey, R. High-performance liquid chromatographic analysis of the products of linoleic acid oxidation catalysed by pea (Pisum sativum) seed lipoxygenases. J. Agric. Food Chem. 1995, 43, 337–342. [Google Scholar] [CrossRef]
- Pokorny, J.; Yanishlieva, N.; Gordon, M. Antioxidants in Food Practical Applications; CRC Woodhead Publishing Ltd.: Boca Raton, FL, USA, 2001; p. 288. [Google Scholar]
- Li, J.K.; Zhang, Y.; Yu, Z.L.; Wang, Y.J.; Yang, Y.; Liu, Z.; Jiang, J.Y.; Song, M.; Wu, Y.J. Superior storage stability in low lipoxygenase maize varieties. J. Stored Prod. Res. 2007, 43, 530–534. [Google Scholar] [CrossRef]
- Mares, D.J.; Cheong, J.; Goonetilleke, S.N.; Mather, D.E. Lipoxygenase in wheat: Genetic control and impact on stability of lutein and lutein esters. Foods 2021, 10, 1149. [Google Scholar] [CrossRef]
- Zhang, Q.X.; Zhao, Y.F.; Zhang, J.L.; Li, X.K.; Ma, F.F.; Duan, M.; Zhang, B.; Li, H.Y. The responses of the lipoxygenase gene fmily to salt and frought stress in foxtail millet (Setaria italica). Life 2021, 11, 1169. [Google Scholar] [CrossRef] [PubMed]
- RoyChowdhury, M.; Li, X.B.; Qi, H.Y.; Li, W.X.; Sun, J.; Huang, C.; Wu, D.X. Functional characterization of 9-/13-LOXs in rice and silencing their expressions to improve grain qualities. Biomed. Res. Int. 2016, 2016, 4275904. [Google Scholar] [CrossRef] [Green Version]
- Verlotta, A.; Simone, V.D.; Mastrangelo, A.M.; Cattivelli, L.; Papa, R.; Trono, D. Insight into durum wheat Lpx-B1: A small gene family coding for the lipoxygenase responsible for carotenoid bleaching in mature grains. BMC Plant Biol. 2010, 10, 263. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer Sequence (5′ to 3′) | Reverse Primer Sequence (5′ to 3′) |
|---|---|---|
| SiACTIN | TGC TCA GTG GAG GCT CAA CA | CAA GAC ACT GTA CTT GCG CTC |
| SiLOX1 | AAA TCA CTG GCT GAG GAC ACA T | CAG GAG CTT GAA GAT CGG GT |
| SiLOX2 | GGT CCT CGG AAA TGT GTT GG | GCT GAA CTT TAC GCA GGC TTA |
| SiLOX3 | AGG AGT TTG GAC GGG AGA TT | CGAGGTTCCTCTCGATGT |
| SiLOX4 | CCC TGG AGA TGT CCT CAA AG | ATT GCC GTC CAG ATT TCG |
| SiLOX5 | GGA CCT AAG GCA GTA TGG A | AGG ATG AAG AGC TTG TTG TT |
| SiLOX6 | CTG CTG TCC TCG CAC TCC | CTC GCT GTC ATC GTT CCA T |
| SiLOX7 | GAC CGT CTT CCC TCG CAA | GTC GTC GGG GTA GTA GAT GG |
| SiLOX8 | CCC AAC AGC GTC ACC ATC TA | CCC CGC CCG AGT ATA ATG AG |
| SiLOX9 | GCG GGT GAT GAT GAC ATA AGT A | TCT TTG TGG CTA TGA TGA ACG |
| SiLOX10 | TAC CAC TAC GGC GGC TAC TT | TCT GCG TTG GGA GCA TGT C |
| SiLOX11 | AAC CTC CTG TCG TCG CAC TC | GGG GTC CTT GTT CCT ACT ATC G |
| SiLOX12 | TAGAAGCCTACACCGATGATAC | CTTCCAGGTTGTGCTGAATAT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ma, Q.; Wang, J.; Cheng, L.; Li, Y.; Zhang, Q.; Li, H.; Han, Y.; Zhen, X.; Zhang, B. The Potential Function of SiLOX4 on Millet Discoloration during Storage in Foxtail Millet. Agriculture 2022, 12, 1283. https://doi.org/10.3390/agriculture12081283
Ma Q, Wang J, Cheng L, Li Y, Zhang Q, Li H, Han Y, Zhen X, Zhang B. The Potential Function of SiLOX4 on Millet Discoloration during Storage in Foxtail Millet. Agriculture. 2022; 12(8):1283. https://doi.org/10.3390/agriculture12081283
Chicago/Turabian StyleMa, Qi, Junjie Wang, Lu Cheng, Yaqiong Li, Qianxiang Zhang, Hongying Li, Yuanhuai Han, Xiaoxi Zhen, and Bin Zhang. 2022. "The Potential Function of SiLOX4 on Millet Discoloration during Storage in Foxtail Millet" Agriculture 12, no. 8: 1283. https://doi.org/10.3390/agriculture12081283