
Screening for Antagonistic Yeasts to Manage Alternaria spp. in Organic Farming

Abstract
:1. Introduction
2. Materials and Methods
2.1. Antagonistic Yeast and Pathogenic Fungi
2.2. Antagonism between Yeast and Alternaria spp.
2.3. Production of Lytic Exoenzymes
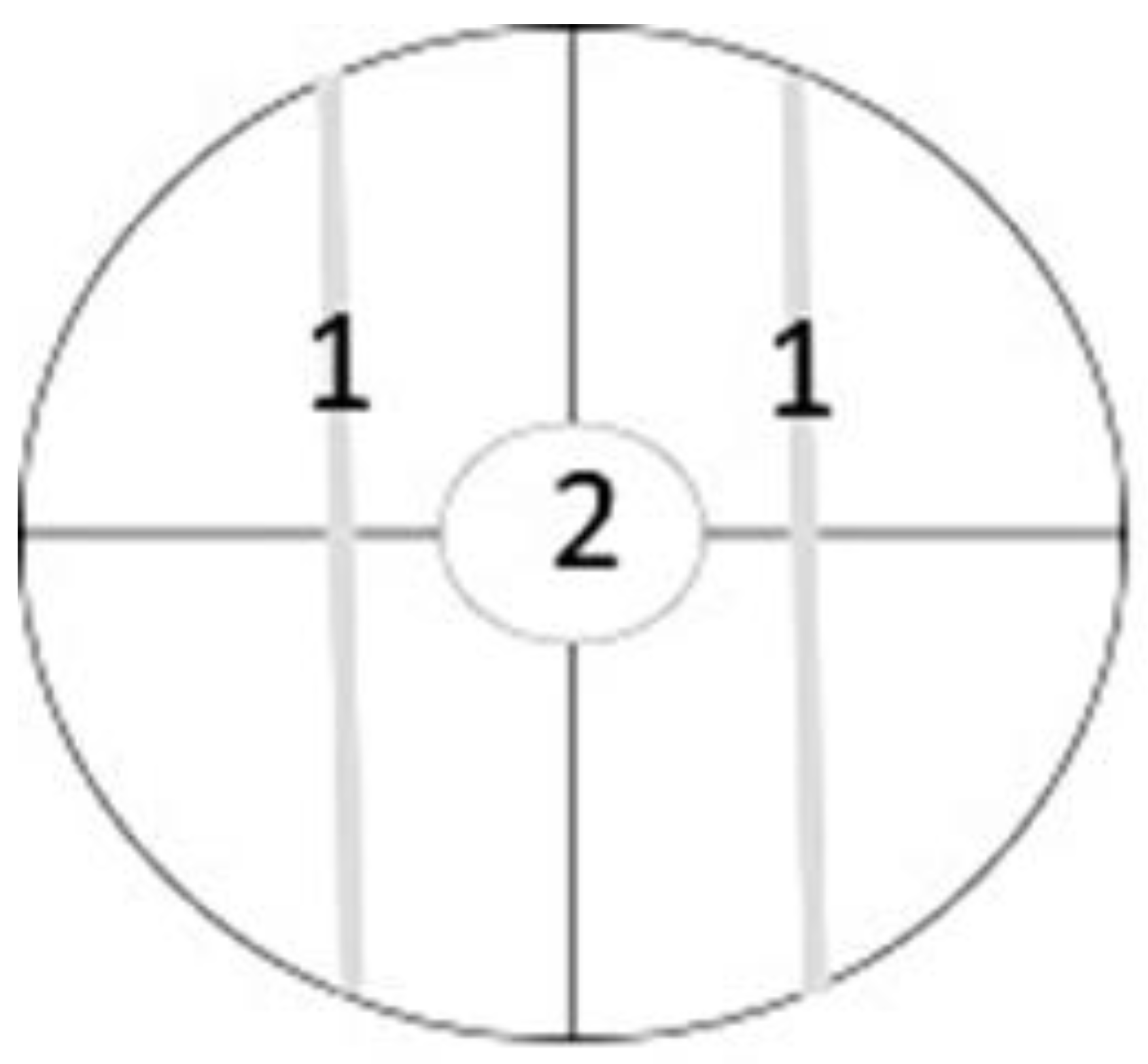
2.4. Alternaria Mycelium Evaluation after Contact with Yeast Strains
2.5. Colonization of Yeasts on the Potato Leaf Surface
2.6. Statistical Analysis
3. Results
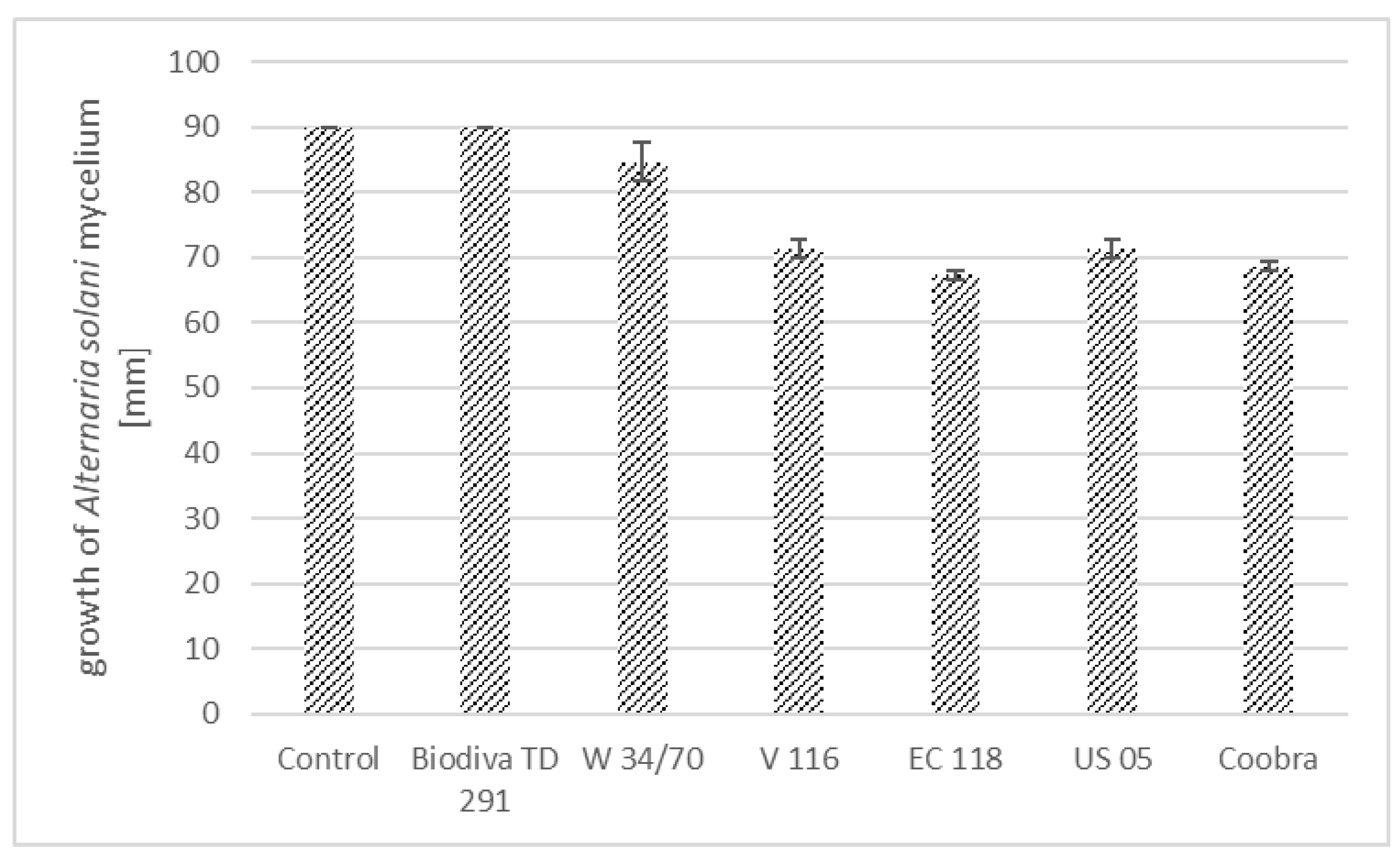
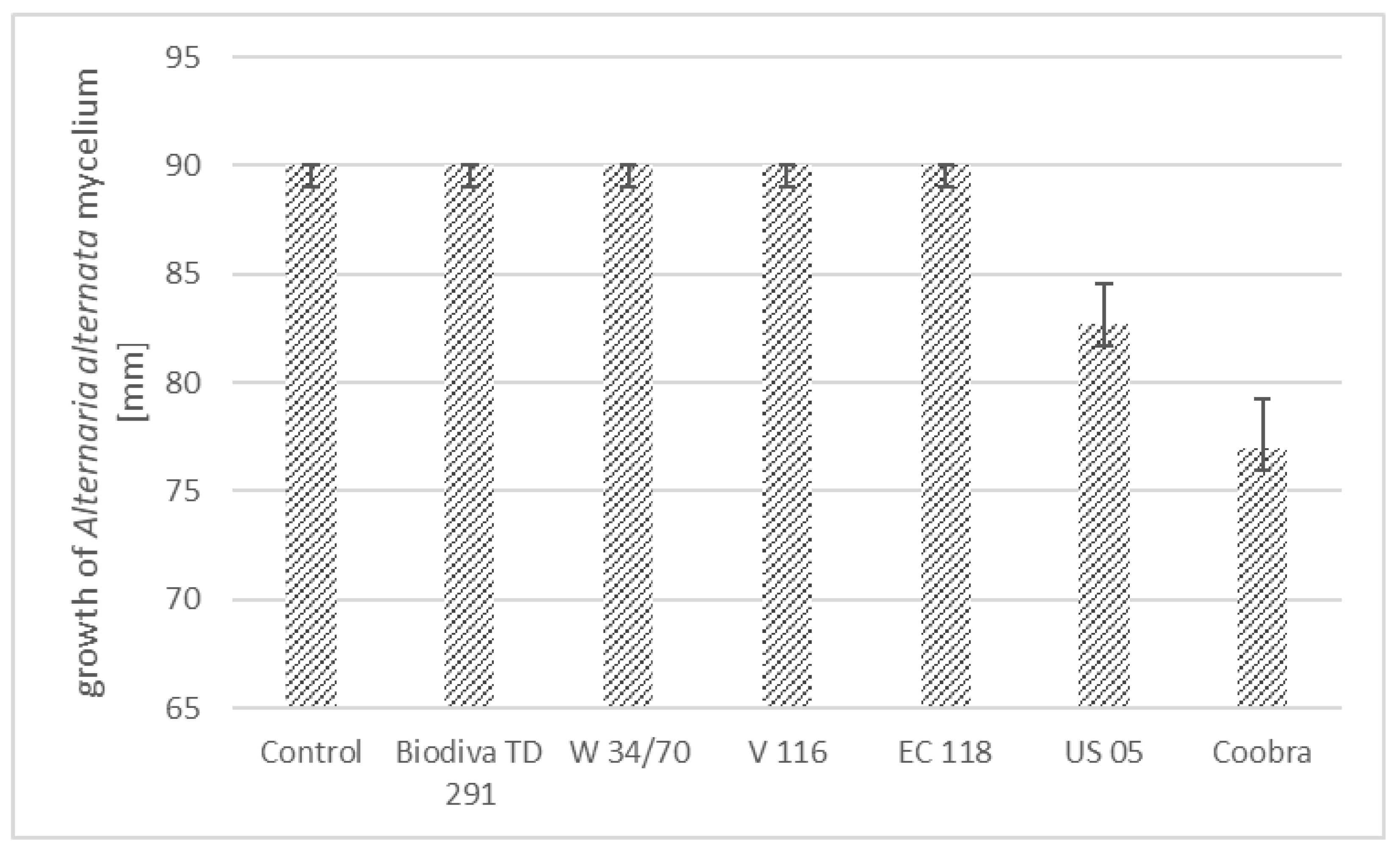
3.1. Growth of Alternaria spp. Inhibited by Antagonistic Yeast Strains
3.2. Reducing Symptoms Caused by Alternaria spp. on Potato Plants in a Greenhouse Experiment
3.3. Production of Lytic Exoenzymes—Results
3.4. Alternaria Mycelium Evaluation after Contact with Yeast Strains—Results
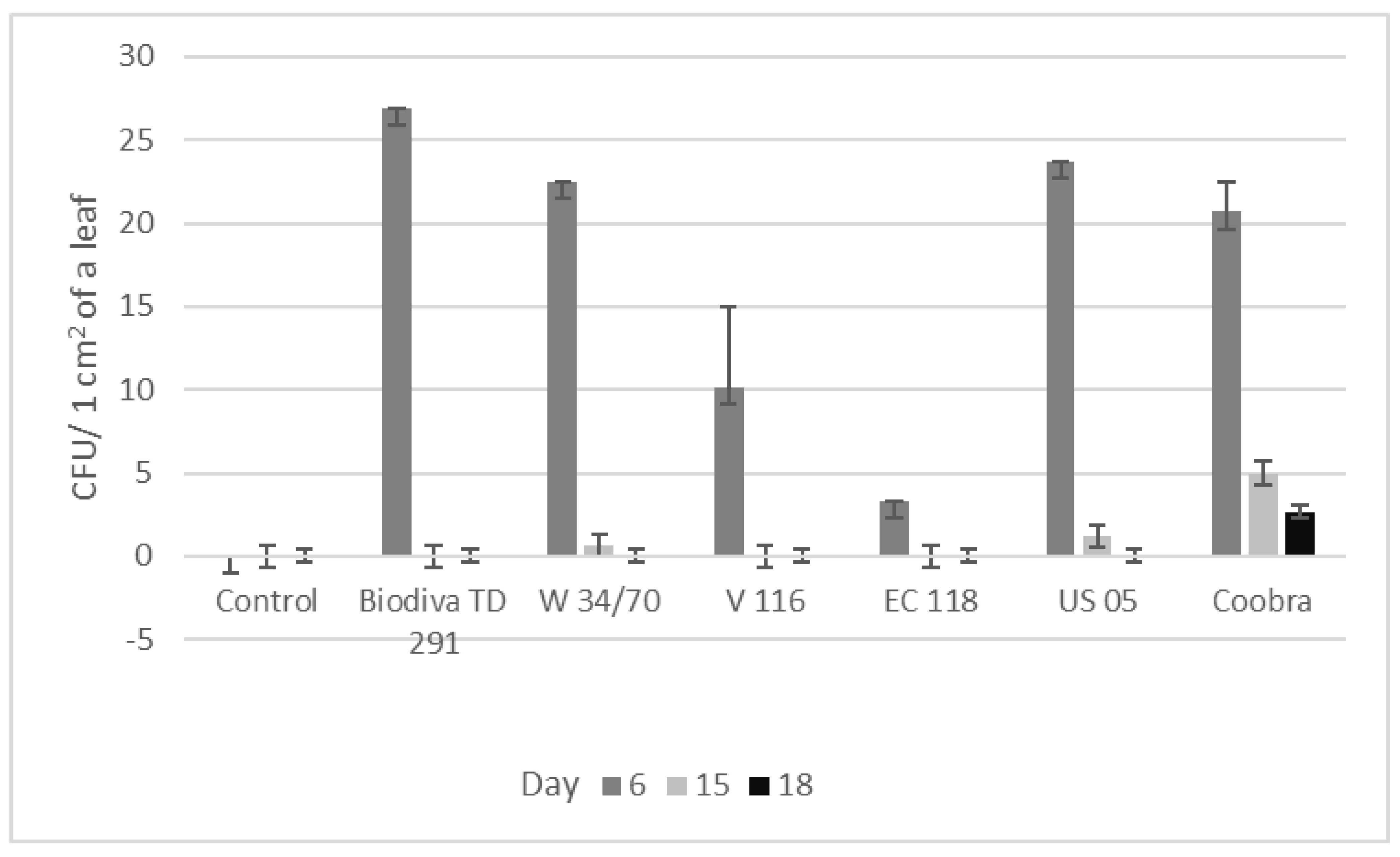
3.5. Population Dynamics of the Antagonistic Yeast Strains on the Potato Leaf Surface
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Budde-Rodriguez, S.; Pasche, J.S.; Mallik, I.; Gudmestad, N.C. Sensitivity of Alternaria spp. from potato to pyrimethanil, cyprodinil, and fludioxonil. Crop Prot. 2022, 152, 105855. [Google Scholar] [CrossRef]
- Jindo, K.; Evenhuis, A.; Kempenaar, C.; Pombo Sudré, C.; Zhan, X.; Goitom Teklu, M.; Kessel, G. Holistic pest management against early blight disease towards sustainable agriculture. Pest Manag. Sci. 2021, 77, 3871–3880. [Google Scholar] [CrossRef]
- Saharan, G.S.; Mehta, N.; Meena, P.D.; Dayal, P. Alternaria Diseases of Crucifers: Biology, Ecology and Disease Management, 1st ed.; Springer: Singapore, 2016; pp. 17–51. [Google Scholar]
- Nowicki, M.; Nowakowska, M.; Niezgoda, A.; Kozik, E. Alternaria black spot of crucifers: Symptoms, the importance of the disease, and perspectives of resistance breeding. Veg. Crops Res. Bull. 2012, 76, 5–19. [Google Scholar] [CrossRef]
- Tsedaley, B. Review on early blight (Alternaria spp.) of potato disease and its management options. J. Biol. Agric. Healthc. 2014, 4, 191–199. [Google Scholar]
- Rashmi, T.; Vishunavat, K. Management of early blight (Alternaria solani) in tomato by integration of fungicides and cultural practices. Int. J. Plant Prot. 2012, 5, 201–206. [Google Scholar]
- Van Bruggen, A.H.C.; Finckh, M.R. Plant diseases and management approaches in organic farming systems. Annu. Rev. Phytopathol. 2016, 54, 25–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, J.; Yadav, A.N. Natural Bioactive Products in Sustainable Agriculture, 1st ed.; Springer Nature: Singapore, 2020; p. 319. [Google Scholar]
- Damalas, C.A.; Koutroubas, S.D. Current status and recent developments in biopesticide use. Agriculture 2018, 8, 13. [Google Scholar] [CrossRef] [Green Version]
- Pal, K.K.; Gardener, B.M. Biological Control of Plant Pathogens. The Plant Health Instructor. Available online: https://www.apsnet.org/edcenter/disimpactmngmnt/topc/Documents/PHIBiologicalControl.pdf (accessed on 21 April 2022).
- Trivedi, P.; Leach, J.E.; Tringe, S.G.; Sa, T.; Singh, B.K. Plant–microbiome interactions: From community assembly to plant health. Nat. Rev. Microbiol. 2020, 18, 607–621. [Google Scholar] [CrossRef] [PubMed]
- Kowalska, J.; Krzymińska, J.; Tyburski, J. Yeasts as a Potential Biological Agent in Plant Disease Protection and Yield Improvement—A Short Review. Agriculture 2022, 12, 1404. [Google Scholar] [CrossRef]
- Caplice, E.; Fitzgerald, G.F. Food fermentations: Role of microorganisms in food production and preservation. Int. J. Food Microbiol. 1999, 50, 131–149. [Google Scholar] [CrossRef]
- Chambers, P.J.; Pretorius, I.S. Fermenting knowledge: The history of winemaking, science and yeast research. EMBO Rep. 2010, 11, 914–920. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jach, M.E.; Serefko, A.; Sajnaga, E.; Kozak, E.; Poleszak, E.; Malm, A. Dietary supplements based on the yeast biomass. Curr. Top. Nutraceutical Res. 2015, 13, 83–88. [Google Scholar]
- Kurtzman, C.P.; Fell, J.W.; Boekhout, T. (Eds.) The Yeasts: A Taxonomic Study, 5th ed.; Elsevier: Amsterdam, The Netherlands, 2011; p. 2354. [Google Scholar]
- Gasch, A.P.; Werner-Washburne, M. The genomics of yeast responses to environmental stress and starvation. Funct. Integr. Genom. 2002, 2, 181–192. [Google Scholar] [CrossRef] [PubMed]
- Bencheqroun, S.K.; Bajji, M.; Massart, S.; Labhilili, M.; El Jaafari, S.; Jijakli, M.H. In vitro and in situ study of postharvest apple blue mould biocontrol by Aureobasidium pullulans: Evidence for the involvement of competition for nutrients. Postharvest Biol. Technol. 2007, 46, 128–135. [Google Scholar] [CrossRef]
- Spadaro, D.; Droby, S. Development of biocontrol products for postharvest diseases of fruit: The importance of elucidating the mechanisms of action of yeast antagonists. Trends Food Sci. Technol. 2016, 47, 39–49. [Google Scholar] [CrossRef]
- Di Francesco, A.; Ugolini, L.; D’Aquino, S.; Pagnotta, E.; Mari, M. Biocontrol of Monilinia laxa by Aureobasidium pullulans strains: Insights on competition for nutrients and space. Int. J. Food Microbiol. 2017, 248, 32–38. [Google Scholar] [CrossRef] [PubMed]
- Castoria, R.; Wright, S.A.I. Host Responses to Biological Control Agents. In Post-Harvest Pathology; Springer: Berlin/Heidelberg, Germany, 2009; Volume 2, pp. 171–181. [Google Scholar]
- Pretscher, J.; Fischkal, T.; Branscheidt, S.; Jäger, L.; Kahl, S.; Schlander, M.; Claus, H. Yeasts from different habitats and their potential as biocontrol agents. Fermentation 2018, 4, 31. [Google Scholar] [CrossRef] [Green Version]
- Mimee, B.; Labbe, C.; Belanger, R.R. Catabolism of flocculosin, an antimicrobial metabolite produced by Pseudozyma flocculosa. Glycobiology 2009, 19, 995–1001. [Google Scholar] [CrossRef] [Green Version]
- Mannazzu, I.; Domizio, P.; Carboni, G.; Zara, S.; Zara, G.; Comitini, F.; Ciani, M. Yeast killer toxins: From ecological significance to application. Crit. Rev. Biotechnol. 2019, 39, 603–617. [Google Scholar] [CrossRef] [PubMed]
- Mari, M.; Bautista-Baños, S.; Sivakumar, D. Decay control in the postharvest system: Role of microbial and plant volatile organic compounds. Postharvest Biol. Technol. 2016, 122, 70–81. [Google Scholar] [CrossRef]
- Perez, M.F.; Contreras, L.; Garnica, N.M.; Fernández-Zenoff, M.V.; Farías, M.E.; Sepulveda, M.; Dib, J.R. Native killer yeasts as biocontrol agents of postharvest fungal diseases in lemons. PLoS ONE 2016, 11, 0165590. [Google Scholar] [CrossRef] [Green Version]
- Belda, I.; Ruiz, J.; Alonso, A.; Marquina, D.; Santos, A. The biology of Pichia membranifaciens killer toxins. Toxins 2017, 9, 112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, S.T.; Chiu, M.C.; Chou, J.Y. Broad-spectrum activity of volatile organic compounds from three yeast-like fungi of the Galactomyces genus against diverse plant pathogens. Mycobiology 2021, 49, 69–77. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Montaño, F.; Alías-Villegas, C.; Bellogín, R.A.; Del Cerro, P.; Espuny, M.R.; Jiménez-Guerrero, I.; López-Baena, F.J.; Ollero, F.J.; Cubo, T. Plant growth promotion in cereal and leguminous agricultural important plants: From microorganism capacities to crop production. Microbiol. Res. 2014, 169, 325–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nafady, N.A.; Hashem, M.; Hassan, E.A.; Ahmed, H.A.; Alamri, S.A. The combined effect of arbuscular mycorrhizae and plant-growth-promoting yeast improves sunflower defense against Macrophomina phaseolina diseases. Biol. Control 2019, 138, 104049. [Google Scholar] [CrossRef]
- Czarnecka, M.; Żarowska, B.; Połomska, X.; Restuccia, C.; Cirvilleri, G. Role of biocontrol yeasts Debaryomyces hansenii and Wickerhamomyces anomalus in plants’ defence mechanisms against Monilinia fructicola in apple fruits. Food Microbiol. 2019, 83, 1–8. [Google Scholar] [CrossRef]
- Mbuyane, L.L. Investigating Polyol and Acetic Acid Metabolism in Wine-Related Non-Saccharomyces Yeasts. Ph.D. Thesis, Stellenbosch University, Stellenbosch, South Africa, 2017. [Google Scholar]
- Bruner, J.; Marcus, A.; Fox, G. Brewing Efficacy of Non-Conventional Saccharomyces Non-cerevisiae Yeasts. Beverages 2021, 7, 68. [Google Scholar] [CrossRef]
- León-Teutli, A.K.; Hernández-Iturriaga, M.; Miranda-Castilleja, D.E.; Aldrete-Tapia, J.A.; Soto-Muñoz, L.; Martínez-Peniche, R.Á. Selection of native yeasts for the production of sparkling ciders from apple varieties established in Querétaro, Mexico. Rev. Chapingo Ser. Hortic. 2020, 26, 29–47. [Google Scholar] [CrossRef]
- Tyrawa, C.; Preiss, R.; Armstrong, M.; van der Merwe, G. The temperature dependent functionality of Brettanomyces bruxellensis strains in wort fermentations. J. Inst. Brew. 2019, 125, 315–325. [Google Scholar] [CrossRef]
- Wang, X.D.; Bohlscheid, J.C.; Edwards, C.G. Fermentative activity and production of volatile compounds by Saccharomyces grown in synthetic grape juice media deficient in assimilable nitrogen and/or pantothenic acid. J. Appl. Microbiol. 2003, 94, 349–359. [Google Scholar] [CrossRef] [PubMed]
- McFarland, J. The nephelometer: An instrument for estimating the number of bacteria in suspensions used for calculating the opsonic index and for vaccines. J. Am. Med. Assoc. 1907, 49, 1176–1178. [Google Scholar] [CrossRef] [Green Version]
- Qazi, N.; Nayeema, J.; Qaisar, A.; Aadil, A. Status and symptomatology of early blight (Alternaria solani) of potato (Solanum tuberosum L.) in Kashmir valley. Afr. J. Agric. Res. 2013, 8, 5104–5115. [Google Scholar] [CrossRef]
- Gabriel, B.P. Enzymatic activities of some entomophthorous fungi. J. Invertebr. Pathol. 1968, 11, 70–81. [Google Scholar] [CrossRef]
- Strauss, M.L.A.; Jolly, N.P.; Lambrechts, M.G.; Van Rensburg, P. Screening for the production of extracellular hydrolytic enzymes by non-Saccharomyces wine yeasts. J. Appl. Microbiol. 2001, 91, 182–190. [Google Scholar] [CrossRef] [PubMed]
- Al-Maawali, S.; Al-Sadi, A.M.; Ali Khalifa Alsheriqi, S.; Al-Sabahi, J.N.; Velazhahan, R. The potential of antagonistic yeasts and bacteria from tomato phyllosphere and fructoplane in the control of Alternaria fruit rot of tomato. All Life 2021, 14, 34–48. [Google Scholar] [CrossRef]
- Samada, L.H.; Tambunan, U.S.F. Biopesticides as promising alternatives to chemical pesticides: A review of their current and future status. Online J. Biol. Sci. 2020, 20, 66–76. [Google Scholar] [CrossRef]
- Ab Rahman, S.F.S.; Singh, E.; Pieterse, C.M.; Schenk, P.M. Emerging microbial biocontrol strategies for plant pathogens. Plant Sci. 2018, 267, 102–111. [Google Scholar] [CrossRef] [Green Version]
- Droby, S.; Wisniewski, M.; Teixidó, N.; Spadaro, D.; Jijakli, M.H. The science, development, and commercialization of postharvest biocontrol products. Postharvest Biol. Technol. 2016, 122, 22–29. [Google Scholar] [CrossRef]
- Raymaekers, K.; Ponet, L.; Holtappels, D.; Berckmans, B.; Cammue, B.P. Screening for novel biocontrol agents applicable in plant disease management—A review. Biol. Control 2020, 144, 104240. [Google Scholar] [CrossRef]
- Menolli, N., Jr.; Marques, V.; Paccola, L.; Andrade-Nóbrega, G. Inhibitory activity of yeasts against soybean pathogenic fungi. Agrotrópica 2020, 32, 75–80. [Google Scholar] [CrossRef]
- Istifadah, N.; Putri, R.A.; Hartati, S. The abilities of bacteria and yeast isolated from vermicompost water extract to inhibit Alternaria solani in vitro and early blight disease on tomato. Cropsaver J. Plant Prot. 2021, 4, 73–79. [Google Scholar] [CrossRef]
- Sabaghian, S.; Braschi, G.; Vannini, L.; Patrignani, F.; Samsulrizal, N.H.; Lanciotti, R. Isolation and identification of wild yeast from Malaysian grapevine and evaluation of their potential antimicrobial activity against grapevine fungal pathogens. Microorganisms 2021, 9, 2582. [Google Scholar] [CrossRef] [PubMed]
- Prendes, L.P.; Merín, M.G.; Fontana, A.R.; Bottini, R.A.; Ramirez, M.L.; de Ambrosini, V.I.M. Isolation, identification and selection of antagonistic yeast against Alternaria alternata infection and tenuazonic acid production in wine grapes from Argentina. Int. J. Food Microbiol. 2018, 266, 14–20. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Xie, F.; Zhao, Y.; Cao, J. Inhibitory effect of postharvest yeast mannan treatment on Alternaria rot of tomato fruit involving the enhancement of hemicellulose polysaccharides and antioxidant metabolism. Sci. Hortic. 2021, 277, 109798. [Google Scholar] [CrossRef]
- Cwalina-Ambroziak, B.; Damszel, M.M.; Glosek-Sobieraj, M. The effect of biological and chemical control agents on the health status of the very early potato cultivar Rosara. J. Plant Prot. Res. 2015, 55, 389–395. [Google Scholar] [CrossRef]
- Jyoti, C.; Jain, D.K. Exploring the potentiality of biocontrol agents against Alternaria solani: A causal agent of early blight of potato and tomato crops. Ann. Biol. 2020, 36, 67–70. [Google Scholar]
- Gorai, P.S.; Ghosh, R.; Konra, S.; Mandal, N.C. Biological control of early blight disease of potato caused by Alternaria alternata EBP3 by an endophytic bacterial strain Bacillus velezensis SEB1. Biol. Control 2021, 156, 104551. [Google Scholar] [CrossRef]
- Da Silva, H.A.O.; Teixeira, W.D.; Borges, Á.V.; Silva Junior, A.L.; Alves, K.S.; Rodrigues Junior, O.M.; de Abreu, L.M. Biocontrol of potato early blight and suppression of Alternaria grandis sporulation by Clonostachys spp. Plant Pathol. 2021, 70, 1677–1685. [Google Scholar] [CrossRef]
- Meena, B.R.; Yadav, J.; Chittora, D.; Meena, S.R.; Kumar, S.; Jain, T.; Sharma, K. Biocontrol Efficacy of Thevetia Peruviana Bioformulations Against Early Blight Disease of Potato. Thevetia Peruviana 2022. submitted. [Google Scholar] [CrossRef]
- Tumpa, F.H.; Khokom, M.A.R. Foliar application of chitosan and yeast elicitor fa-cilitate reducing incidence and severity of Alternaria leaf blight of tomato and brinjal. Res. J. Plant Pathol. 2020, 3, 4. [Google Scholar]
- Wang, Y.; Bao, Y.; Shen, D.; Feng, W.; Yu, T.; Zhang, J.; Zheng, X.D. Biocontrol of Alternaria alternata on cherry tomato fruit by use of marine yeast Rhodosporidium paludigenum Fell & Tallman. Int. J. Food Microbiol. 2008, 123, 234–239. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Wang, P.; Xia, J.; Yu, T.; Lou, B.; Wang, J.; Zheng, X.D. Effect of water activity on stress tolerance and biocontrol activity in antagonistic yeast Rhodosporidium paludigenum. Int. J. Food Microbiol. 2010, 143, 103–108. [Google Scholar] [CrossRef] [PubMed]
- Yan, F.; Xu, S.; Chen, Y.; Zheng, X. Effect of rhamnolipids on Rhodotorula glutinis biocontrol of Alternaria alternata infection in cherry tomato fruit. Postharvest Biol. Technol. 2014, 97, 32–35. [Google Scholar] [CrossRef]
- Zhao, Y.; Tu, K.L.; Shao, X.F.; Jing, W.; Yang, J.L.; Su, Z.P. Biological control of the post-harvest pathogens Alternaria solani, Rhizopus stolonifer, and Botrytis cinerea on tomato fruit by Pichia guilliermondii. J. Hortic. Sci. Biotechnol. 2008, 83, 132–136. [Google Scholar] [CrossRef]
- Salazar, O.; Asenjo, J.A. Enzymatic lysis of microbial cells. Biotechnol. Lett. 2007, 29, 985–994. [Google Scholar] [CrossRef]
- Peberdy, J.F. Mycolytic enzymes. In Fungal Protoplasts; CRC Press: Boca Raton, FL, USA, 2020; pp. 31–44. [Google Scholar]
- Bar-Shimon, M.; Yehuda, H.; Cohen, L.; Weiss, B.; Kobeshnikov, A.; Daus, A.; Droby, S. Characterization of extracellular lytic enzymes produced by the yeast biocontrol agent Candida oleophila. Curr. Genet. 2004, 45, 140–148. [Google Scholar] [CrossRef]
- Bae, J.; Kuroda, K.; Ueda, M. Proximity effect among cellulose-degrading enzymes displayed on the Saccharomyces cerevisiae cell surface. Appl. Environ. Microbiol. 2015, 81, 59–66. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Chi, Z.M. Screening and identification of a cellulase producing marine yeast and optimization of medium and cultivation conditions for cellulase production. J. Ocean Univ. China 2007, 37, 101–108. [Google Scholar]
- Saravanakumar, D.; Spadaro, D.; Garibaldi, A.; Gullino, M.L. Detection of enzymatic activity and partial sequence of a chitinase gene in Metschnikowia pulcherrima strain MACH1 used as post-harvest biocontrol agent. Eur. J. Plant Pathol. 2009, 123, 183–193. [Google Scholar] [CrossRef]
- Hesham, A.; Sulaiman, A.; Mona, A.; Al-Dayel, M.A.; Salah, H.A. Screening and genetic identification of acidic and neutral protease-producing yeasts strains by 26S rRNA gene sequencing. Genet. Cytol. 2017, 51, 221–229. [Google Scholar] [CrossRef]
- Barzkar, N. Marine microbial alkaline protease: An efficient and essential tool for various industrial applications. Int. J. Biol. Macromol. 2020, 161, 1216–1229. [Google Scholar] [CrossRef] [PubMed]
- Duarte, A.W.F.; Dos Santos, J.A.; Vianna, M.V.; Vieira, J.M.F.; Mallagutti, V.H.; Inforsato, F.J.; Durães Sette, L. Cold-adapted enzymes produced by fungi from terrestrial and marine Antarctic environments. Crit. Rev. Biotechnol. 2018, 38, 600–619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, P.H.; Chen, R.Y.; Chou, J.Y. Screening and evaluation of yeast antagonists for biological control of Botrytis cinerea on strawberry fruits. Mycobiology 2018, 46, 33–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kapoor, D.; Karnwal, A. Yeast as a plant growth promoter and biocontrol agent. In Fungi Bio-Prospects in Sustainable Agriculture, Environment and Nano-Technology; Academic Press: Cambridge, MA, USA, 2021; pp. 429–457. [Google Scholar]
- Shariq, M.; Sohail, M. Application of Candida tropicalis MK-160 for the production of xylanase and ethanol. J. King Saud Univ.—Sci. 2019, 31, 1189–1194. [Google Scholar] [CrossRef]
- Van Driesche, R.; Bellows, T.S., Jr. Biological Control, 1st ed.; Springer Science & Business Media: Norwell, MA, USA, 2012; p. 539. [Google Scholar]
- Cheng, Y.T.; Zhang, L.; He, S.Y. Plant-microbe interactions facing environmental challenge. Cell Host Microbe 2019, 26, 183–192. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, S.D.; Castillo, J.A. Influence of light on plant–phyllosphere interaction. Front. Plant Sci. 2018, 9, 1482. [Google Scholar] [CrossRef]
- González-Estrada, R.R.; de Jesus Ascencio-Valle, F.; Ragazzo-Sánchez, J.A.; Santoyo, M.C. Use of a marine yeast as a biocontrol agent of the novel pathogen Penicillium citrinum on Persian lime. Emir. J. Food Agric. 2017, 29, 114–122. [Google Scholar] [CrossRef] [Green Version]
- Zheng, F.; Zhang, W.; Sui, Y.; Ding, R.; Yi, W.; Hu, Y.; Zhu, C. Sugar protectants improve the thermotolerance and biocontrol efficacy of the biocontrol yeast, Candida oleophila. Front. Microbiol. 2019, 10, 187. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain Code in Tests | Comercially Strain | Supplier | Species |
|---|---|---|---|
| 01 | Biodiva TD 291 | A | Torulaspora delbrueckii |
| 02 | W 34/70 | B | Saccharomyces pastorianus |
| 03 | V 116 | C | Saccharomyces cerevisiae |
| 04 | EC 118 | C | Saccharomyces bayanus |
| 05 | US 05 | B | Saccharomyces cerevisiae |
| 06 | Coobra | D | Saccharomyces cerevisiae |
| Strain | Control | Biodiva TD 291 | W 34/70 | V 116 | EC 118 | US 05 | Coobra |
|---|---|---|---|---|---|---|---|
| Effect | 0 | 0 | + | + | ++ | + | ++ |
| Strain | Control | Biodiva TD 291 | W 34/70 | V 116 | EC 118 | US 05 | Coobra |
|---|---|---|---|---|---|---|---|
| Effect | 0 | 0 | + | + | ++ | + | ++ |
| Strain | Control | Biodiva TD 291 | W 34/70 | V 116 | EC 118 | US 05 | Coobra |
|---|---|---|---|---|---|---|---|
| A. alternata | 60.00 ± 3.27 a | 54.67 ± 3.77 a | 60.00 ± 3.27 a | 41.33 ± 1.89 b | 46.67 ± 1.89 b | 17.33 ± 1.25 c | 14.67 ± 1.89 c |
| A. solani | 61.33 ± 4.99 a | 49.33 ± 7.54 a | 62.67 ± 1.89 a | 25.33 ± 4.99 b | 26.67 ± 5.56 | 26.67 ± 3.77 b | 9.33 ± 2.87 c |
| Exoenzyme | Yeast Strain | |||||
|---|---|---|---|---|---|---|
| Biodiva TD 291 | W 34/70 | V 116 | EC 118 | US 05 | Coobra | |
| Amylase | + | - | - | - | + | + |
| Xylanase | - | - | - | - | - | - |
| Lipase | - | - | - | - | + | - |
| Pectinase | - | - | - | + | - | + |
| Protease | + | - | - | - | - | + |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kowalska, J.; Krzymińska, J.; Matysiak, K.; Jakubowska, M. Screening for Antagonistic Yeasts to Manage Alternaria spp. in Organic Farming. Agriculture 2022, 12, 1693. https://doi.org/10.3390/agriculture12101693
Kowalska J, Krzymińska J, Matysiak K, Jakubowska M. Screening for Antagonistic Yeasts to Manage Alternaria spp. in Organic Farming. Agriculture. 2022; 12(10):1693. https://doi.org/10.3390/agriculture12101693
Chicago/Turabian StyleKowalska, Jolanta, Joanna Krzymińska, Kinga Matysiak, and Magdalena Jakubowska. 2022. "Screening for Antagonistic Yeasts to Manage Alternaria spp. in Organic Farming" Agriculture 12, no. 10: 1693. https://doi.org/10.3390/agriculture12101693