
Effect of Maternal Dietary Condensed Tannins from Sainfoin (Onobrychis viciifolia) on Gut Health and Antioxidant-Immune Crosstalk in Suckling Lambs

 ,
,  ,
,  ,
,  , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Animals, Diets, and Experimental Design
2.3. Blood Sample Collection and Fructosamine Analysis
2.4. Coccidian Faecal Egg Count
2.5. Histology and RT-qPCR
2.6. Histopathological Examination and Microscopic Lesion Scoring
2.7. RT-qPCR Analysis
2.8. Statistical Analysis
3. Results
3.1. Lamb Performance and Plasma Fructosamine
3.2. Faecal Oocyst Count
3.3. Histopathological Analysis
3.4. Gene Expression
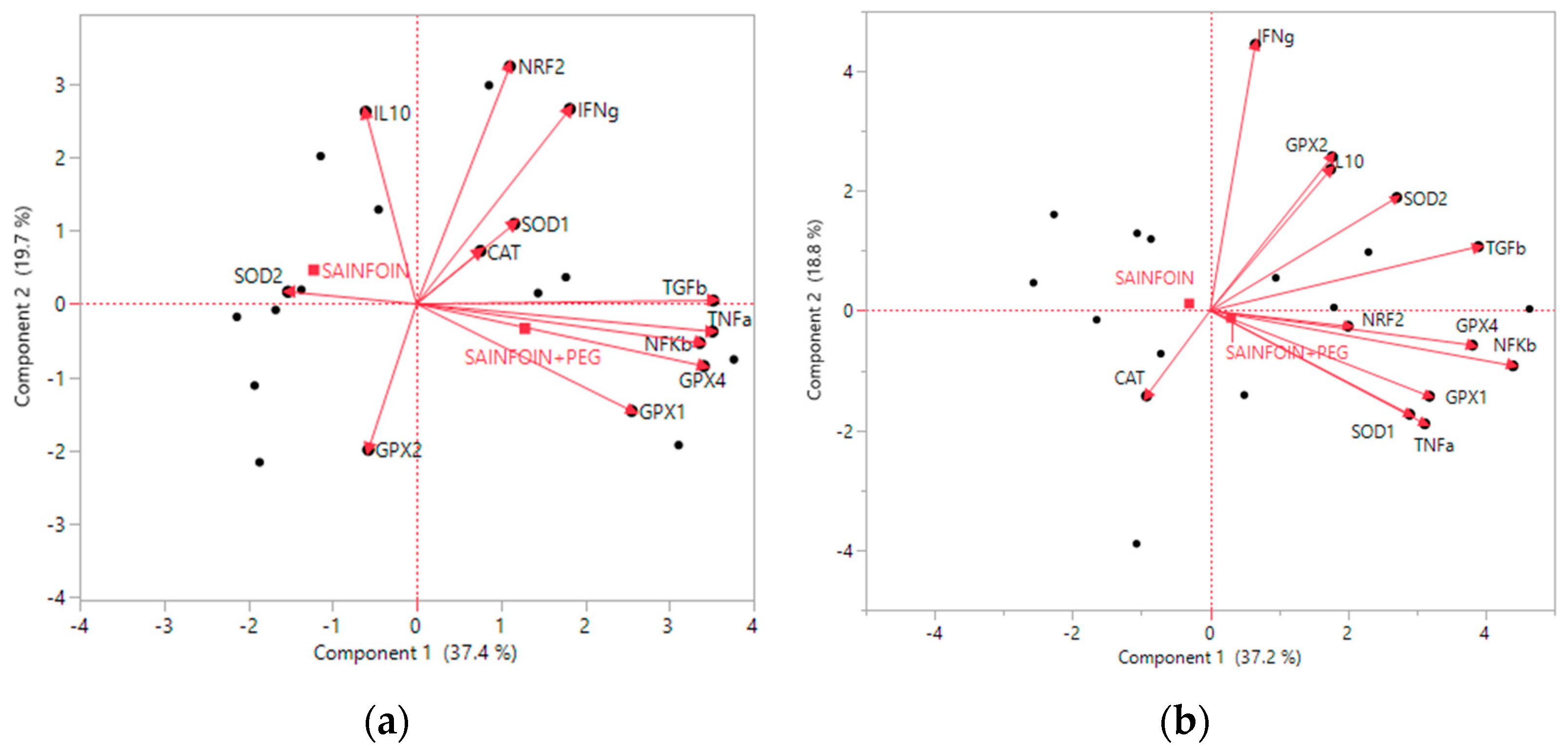
3.5. Principal Components Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sañudo, C.; Sanchez, A.; Alfonso, M. Small ruminant production systems and factors affecting lamb meat quality. Meat Sci. 1998, 49, 29–64. [Google Scholar] [CrossRef]
- Lima., E.; Lovatt., F.; Green, M.; Roden, J.; Davies, P.; Kaler, J. Sustainable lamb production: Evaluation of factors affecting lamb growth using hierarchical, cross classified and multiple memberships models. Prev. Vet. Med. 2020, 174, 104822. [Google Scholar] [CrossRef]
- Hoste, H.; Torres-Acosta, J.F.J.; Sandoval-Castro, C.A.; Mueller-Harvey, I.; Sotiraki, S.; Louvandini, H.; Thamsborg, S.M.; Terrill, T.H. Tannin containing legumes as a model for nutraceuticals against digestive parasites in livestock. Vet. Parasitol. 2015, 212, 5–17. [Google Scholar] [CrossRef] [PubMed]
- Gioxari, A.; Amerikanou, C.; Nestoridi, I.; Gourgari, E.; Pratsinis, H.; Kalogeropoulos, N.; Andrikopoulos, N.K.; Kaliora, A.C. Carob: A Sustainable Opportunity for Metabolic Health. Foods 2022, 11, 2154. [Google Scholar] [CrossRef] [PubMed]
- Keeton, S.T.N.; Navarre, C.B. Coccidiosis in large and small ruminants. Vet. Clin. Food Anim. 2017, 34, 201–208. [Google Scholar] [CrossRef] [PubMed]
- Bangoura, B.; Bardsley, K.D. Ruminant Coccidiosis. Vet. Clin. Food Anim. 2020, 36, 187–203. [Google Scholar] [CrossRef] [PubMed]
- Chartier, C.; Paraud, C. Coccidiosis due to Eimeria in sheep and goats, a review. Small Rumin. Res. 2012, 103, 84–92. [Google Scholar] [CrossRef]
- Saratsis, A.; Voutzourakis, N.; Theodosiou, T.; Stefanakis, A.; Sotiraki, S. The effect of sainfoin (Onobrychis viciifolia) and carob pods (Ceratonia silique) feeding regimes on the control of lamb coccidiosis. Parasitol. Res. 2016, 115, 2233–2242. [Google Scholar] [CrossRef] [PubMed]
- Roila, R.; Branciari, R.; Pecorelli, I.; Cristofani, E.; Carloni, C.; Ranucci, D.; Fioroni, L. Occurrence and Residue Concentration of Coccidiostats in Feed and Food of Animal Origin; Human Exposure Assessment. Foods 2019, 8, 477. [Google Scholar] [CrossRef] [Green Version]
- Muller-Harvey, I.; Bee, G.; Dohme-Meier, F.; Hoste, H.; Karonen, M.; Kölliker, R.; Lüscher, A.; Niderkorn, V.; Pellikaan, W.F.; Salminen, J.P.; et al. Benefits of condensed tannins legumes fed to ruminants: Importance of structure, concentration and diet composition. Crop Sci. 2019, 59, 861–885. [Google Scholar] [CrossRef]
- Álvarez-Rodríguez, J.; Urrutia, O.; Lobón, S.; Ripoll, G.; Bertolín, J.R.; Joy, M. Insights into the role of major bioactive dietary nutrients in lamb meat quality: A review. J. Anim. Sci. Biotechnol. 2022, 13, 20. [Google Scholar] [CrossRef] [PubMed]
- Blomstrand, B.M.; Enemark, H.L.; Steinshamn, H.; Aasen, I.M.; Johanssen, J.R.E.; Athanasiadou, S.; Thamsborg, S.M.; Sørheim, K.M. Administration of spruce bark (Picea abies) extracts in young lambs exhibits anticoccidial effects but reduces milk intake and body weight gain. Acta Vet. Scand. 2022, 64, 10. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Fonseca, A.; Alcala-Canto, Y.; Salem, A.Z.; Alberti-Navarro, A.B. Anticoccidial efficacy of naringenin and a grapefruit peel extract in growing lambs naturally-infected with Eimeria spp. Vet. Parasitol. 2016, 232, 58–65. [Google Scholar] [CrossRef] [PubMed]
- Quijada, J.; Drake, C.; Gaudin, E.; El-Korso, R.; Hoste, H.; Muller-Harvey, I. Condensed tannin changes along the digestive tract in lambs fed with sainfoin pellets or hazelnut skins. J. Agric. Food Chem. 2018, 66, 2136–2142. [Google Scholar] [CrossRef]
- Rivaroli, D.; Prunier, A.; Meteau, K.; do Prado, I.N.; Prache, S. Tannin-rich sainfoin pellet supplementation reduces fat volatile indoles content and delays digestive parasitism in lambs grazing alfalfa. Animal 2019, 13, 1883–1890. [Google Scholar] [CrossRef]
- Niderkorn, V.; Barbier, E.; Macheboeuf, D.; Torrent, A.; Mueller-Harvey, I.; Hoste, H. In vitro rumen fermentation of diets with different types of condensed tannins derived from sainfoin (Onobrychis viciifolia Scop.) pellets and hazelnut (Corylus avellana L.) pericarps. Anim. Feed Sci. Technol. 2020, 259, 114357. [Google Scholar] [CrossRef]
- Petrič, D.; Komáromyová, M.; Batťányi, D.; Kozłowska, M.; Filipiak, W.; Łukomska, A.; Ślusarczyk, S.; Szumacher-Strabel, M.; Cieślak, A.; Várady, M.; et al. Effect of Sainfoin (Onobrychis viciifolia) Pellets on Rumen Microbiome and Histopathology in Lambs Exposed to Gastrointestinal Nematodes. Agriculture 2022, 12, 301. [Google Scholar] [CrossRef]
- Celi, P.; Gabai, G. Oxidant/Antioxidant Balance in Animal Nutrition and Health: The Role of Protein Oxidation. Front. Vet. Sci. 2015, 2, 48. [Google Scholar] [CrossRef] [Green Version]
- Stefanson, A.L.; Bakovic, M. Dietary Regulation of Keap1/Nrf2/ARE Pathway: Focus on Plant-Derived Compounds and Trace Minerals. Nutrients 2014, 6, 3777–3801. [Google Scholar] [CrossRef] [Green Version]
- Rajput, S.A.; Sun, L.; Zhang, N.-Y.; Khalil, M.M.; Ling, Z.; Chong, L.; Wang, S.; Rajput, I.R.; Bloch, D.M.; Khan, F.A.; et al. Grape Seed Proanthocyanidin Extract Alleviates AflatoxinB1-Induced Immunotoxicity and Oxidative Stress via Modulation of NF-κB and Nrf2 Signaling Pathways in Broilers. Toxins 2019, 11, 23. [Google Scholar] [CrossRef]
- Koudoufio, M.; Feldman, F.; Ahmarani, L.; Delvin, E.; Spahis, S.; Desjardins, Y.; Levy, E. Intestinal protection by proanthocyanidins involves anti-oxidative and anti-inflammatory actions in association with an improvement of insulin sensitivity, lipid and glucose homeostasis. Sci. Rep. 2021, 11, 3878. [Google Scholar] [CrossRef] [PubMed]
- Maynard, C.L.; Elson, C.O.; Hatton, R.D.; Weaver, C.T. Reciprocal interactions of the intestinal microbiota and immune system. Nature 2012, 489, 231–241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chassaing, B.; Kumar, M.; Baker, M.T.; Singh, V.; Vijay-Kumar, M. Mammalian gut immunity. Biomed. J. 2014, 37, 246–258. [Google Scholar] [CrossRef] [PubMed]
- Steele, M.A.; Penner, G.B.; Chaucheyras-Durand, F.; Guan, L.L. Development and physiology of the rumen and the lower gut: Targets for improving gut health. J. Dairy Sci. 2016, 99, 4955–4966. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bush, S.J.; McCulloch, M.E.B.; Muriuki, C.; Salavati, M.; Davis, G.M.; Farquhar, I.L.; Lisowski, Z.M.; Archibald, A.L.; Hume, D.A.; Clark, E.L. Comprehensive Transcriptional Profiling of the Gastrointestinal Tract of Ruminants from Birth to Adulthood Reveals Strong Developmental Stage Specific Gene Expression. G3 2019, 9, 359–373. [Google Scholar] [CrossRef] [Green Version]
- Baila, C.; Joy, M.; Blanco, M.; Casasús, I.; Bertolín, J.R.; Lobón, S. Effects of feeding sainfoin proanthocyanidins to lactating ewes on intake, milk production and plasma metabolites. Animal 2022, 16, 100438. [Google Scholar] [CrossRef]
- Baila, C.; Lobón, S.; Blanco, M.; Casasús, I.; Ripoll, G.; Joy, M. Sainfoin in the Dams’ Diet as a Source of Proanthocyanidins: Effect on the Growth, Carcass and Meat Quality of Their Suckling Lambs. Animals 2022, 12, 408. [Google Scholar] [CrossRef]
- Bowman, D.D. Georgis’ Parasitology for Veterinarians, 11th ed.; Elsevier Inc.: St. Louis, MO, USA, 2020; pp. 299–300. [Google Scholar] [CrossRef]
- Dommels, Y.E.; Butts, C.A.; Zhu, S.; Davy, M.; Martell, S.; Hedderley, D.; Barnett, M.P.; McNabb, W.C.; Roy, N.C. Characterization of intestinal inflammation and identification of related gene expression changes in mdr1a(-/-) mice. Genes Nutr. 2007, 2, 209–223. [Google Scholar] [CrossRef] [Green Version]
- Primer3plus. Available online: www.bioinformatics.nl/primer3plus (accessed on 1 February 2019).
- Primer-Blast. Available online: www.ncbi.nlm.nih.gov/tools/primer-blast/ (accessed on 1 February 2019).
- Pelegrin-Valls, J.; Serrano-Pérez, B.; Villalba, D.; Martín-Alonso, M.J.; Bertolín, J.R.; Joy, M.; Álvarez-Rodríguez, J. Effect of Dietary Crude Protein on Productive Efficiency, Nutrient Digestibility, Blood Metabolites and Gastrointestinal Immune Markers in Light Lambs. Animals 2020, 10, 328. [Google Scholar] [CrossRef]
- Schmittgen, T.D.; Livak, K.J. Analysing real-time PCR data by the comparative CT method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef]
- Puech, C.; Dedieu, L.; Chantal, I.; Rodrigues, V. Design and evaluation of a unique SYBR Green real-time RT-PCR assay for quantification of five major cytokines in cattle, sheep and goats. BMC Vet. Res. 2015, 11, 65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lesage-Padilla, A.; Forde, N.; Poirée, M.; Healey, G.D.; Giraud-Delville, C.; Reinaud, P.; Eozenou, C.; Carvalho, A.V.; Galio, L.; Raliou, M.; et al. Maternal metabolism affects endometrial expression of oxidative stress and FOXL2 genes in cattle. PLoS ONE 2017, 12, e0189942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saratsis, A.; Regos, I.; Tzanidakis, N.; Voutzourakis, N.; Stefanakis, A.; Treuter, D.; Joachim, A.; Sotiraki, S. In vivo and in vitro efficacy of sainfoin (Onobrychis viciifolia) against Eimeria spp in lambs. Vet. Parasitol. 2012, 188, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Pout, D.D.; Ostler, D.C.; Joyner, L.P.; Norton, C.C. The coccidial population in clinically normal sheep. Vet. Rec. 1966, 78, 455–460. [Google Scholar] [CrossRef] [PubMed]
- Saratsis, A.; Joachim, A.; Alexandros, S.; Sotiraki, S. Lamb coccidiosis dynamics in different dairy production systems. Vet. Parasitol. 2011, 181, 131–138. [Google Scholar] [CrossRef]
- Reeg, K.J.; Gauly, M.; Bauer, C.; Mertens, C.; Erhardt, G.; Zahner, H. Coccidial infections in housed lambs: Oocyst excretion, antibody levels and genetic influences on the infection. Vet. Parasitol. 2005, 127, 209–219. [Google Scholar] [CrossRef] [PubMed]
- Álvarez-Rodríguez, J.; Sanz, A.; Delfa, R.; Revilla, R.; Joy, M. Performance and grazing behaviour of Churra Tensina sheep stocked under different management systems during lactation on Spanish mountain pastures. Livest. Sci. 2007, 107, 152–161. [Google Scholar] [CrossRef]
- Faichney, G.J. Consumption of solid feed by lambs during their transition from pre-ruminant to full ruminant function. Appl. Anim. Behav. Sci. 1992, 34, 85–91. [Google Scholar] [CrossRef]
- Pond, W.G.; Church, D.C.; Pond, K.R. Basic Animal Nutrition and Feeding, 4th ed.; Wiley: New York, NY, USA, 1995; p. 32. [Google Scholar]
- Vi, R.B.; McLeod, K.R.; Klotz, J.L.; Heitmann, R.N. Rumen development, intestinal growth and hepatic metabolism in the pre-and postweaning ruminant. J. Dairy Sci. 2004, 87, 55–65. [Google Scholar] [CrossRef]
- Ovington, K.S.; Alleva, L.M.; Kerr, E.A. Cytokines and immunological control of Eimeria spp. Int. J. Parasitol. 1995, 25, 1331–1351. [Google Scholar] [CrossRef]
- Yahfoufi, N.; Alsadi, N.; Jambi, M.; Matar, C. The Immunomodulatory and Anti-Inflammatory Role of Polyphenols. Nutrients 2018, 10, 1618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jordán, M.J.; Moñino, M.I.; Martíez, C.; Lafuente, A.; Sotomayor, J.A. Introduction of distillate Rosemary leaves into the diets of the Murciano-Granadina goat: Transfer of polyphenolic compounds to goats’ milk and the plasma of suckling goat kids. J. Agric. Food Chem. 2010, 58, 8265–8270. [Google Scholar] [CrossRef] [PubMed]
- Bagchi, D.; Garg, A.; Krohn, R.L.; Bagchi, M.; Tran, M.X.; Balmoori, J. Oxygen free radical scavenging abilities of vitamins C and E, and a grape seed proanthocyanidins extract in vitro. Res. Commun. Mol. Pathol. Pharmacol. 1997, 95, 179–189. [Google Scholar] [PubMed]
- Gessner, D.K.; Fiesel, A.; Most, E.; Dinges, J.; Wen, G.; Ringseis, R.; Eder, K. Supplementation of a grape seed and grape marc meal extract decreases activities of the oxidative stress-responsive transcription factors NF-κB and Nrf2 in the duodenal mucosa of pigs. Acta Vet. Scand. 2013, 55, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Puiggros, F.; Llópiz, N.; Ardévol, A.; Bladé, C.; Arola, L.; Salvadó, M.J. Grape seed procyanidins prevent oxidative injury by modulating the expression of antioxidant enzyme systems. J. Agric. Food Chem. 2005, 53, 6080–6086. [Google Scholar] [CrossRef] [PubMed]
- Mu, C.; Yang, W.; Wang, P.; Zhao, J.; Hao, X.; Zhang, J. Effects of high-concentrate diet supplemented with grape seed proanthocyanidins on growth performance, liver function, meat quality, and antioxidant activity in finishing lambs. Anim. Feed Sci. Technol. 2020, 266, 114518. [Google Scholar] [CrossRef]
- Moghadasian, M.H.; Godin, D.V. Ethanol-induced gastrointestinal damage influence of endogenous antioxidant components and gender. Dig. Dis. Sci. 1996, 41, 791–797. [Google Scholar] [CrossRef] [PubMed]
- Chedea, V.S.; Palade, L.M.; Marin, D.E.; Pelmus, R.S.; Habeanu, M.; Rotar, M.C.; Gras, M.A.; Pistol, G.C.; Taranu, I. Intestinal Absorption and Antioxidant Activity of Grape Pomace Polyphenols. Nutrients 2018, 10, 588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Florian, S.; Wingler, K.; Schmehl, K.; Jacobasch, G.; Kreuzer, O.J.; Meyerhof, W.; Brigelius-Flohé, R. Cellular and subcellular localization of gastrointestinal glutathione peroxidase in normal and malignant human intestinal tissue. Free Radic. Res. 2001, 35, 655–663. [Google Scholar] [CrossRef] [PubMed]
- Filipović, N.; Stojević, Z.; Mašek, T.; Mikulec, Ž.; Prvanović, N. Relationship between fructosamine with serum protein, albumin and glucose concentrations in dairy ewes. Small Rumin. Res. 2011, 96, 46–48. [Google Scholar] [CrossRef]
- Ibeagha-Awemu, E.M.; Bissonnette, N. Do, D.N.; Dudemaine, P.L.; Wang, M.; Facciuolo, A.; Griebel, P. Regionally Distinct Immune and Metabolic Transcriptional Responses in the Bovine Small Intestine and Draining Lymph Nodes during a Subclinical Mycobacterium avium subsp. paratuberculosis Infection. Front. Immunol. 2021, 12, 760931. [Google Scholar] [CrossRef] [PubMed]
- Hansen, H.F.; Nielsen, D.S.; Kverka, M.; Zakostelska, Z.; Klimesova, K.; Hudcovic, T.; Tlaskalova-Hogenova, H.; Hansen, A.K. Patterns of Early Gut Colonization Shape Future Immune Responses of the Host. PLoS ONE 2012, 7, e34043. [Google Scholar] [CrossRef] [Green Version]
- Lüpertz, R.; Chovolou, Y.; Kampkötter, A.; Wätjen, W.; Kahl, R. Catalase overexpression impairs TNF-alpha induced NF-kappaB activation and sensitizes MCF-7 cells against TNF-alpha. J. Cell. Biochem. 2008, 103, 1497–1511. [Google Scholar] [CrossRef] [PubMed]
- Lei, X.G.; Zhu, J.H.; Cheng, W.H.; Bao, Y.; Ho, Y.S.; Reddi, A.R.; Holmgren, A.; Arnér, E.S. Paradoxical Roles of Antioxidant Enzymes: Basic Mechanisms and Health Implications. Physiol. Rev. 2016, 96, 307–364. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Gene | Tissue-Function Ratio | Forward and Reverse Primer (5′–3′) 1 | bp 2 | Access. No. | E (%) 3 | nM 4 | Source |
|---|---|---|---|---|---|---|---|
| GAPDH | Reference genes | F: ATCTCGCTCCTGGAAGATG R: TCGGAGTGAACGGATTCG | 200 | NM_001190390.1 | 1.90 | 600 300 | [34] |
| ACTB | Reference genes | F: CTGGACTTCGAGCAGGAGAT R: GATGTCGACGTCACACTTC | 194 | NM_001009784 | 1.94 | 600 | [34] |
| IL10 | Anti-inflammatory | F: TTAAGGGTTACCTGGGTTGC R: TTCACGTGCTCCTTGATGTC | 109 | NM_001009327.1 | 1.96 | 200 | [32] |
| TGFB | Anti-inflammatory | F: TTGACGTCACTGGAGTTGTG R: CGTTGATGTCCACTTGAAGC | 120 | NM_001009400.2 | 2.04 | 200 | [32] |
| TNFA | Pro-inflammatory | F: CAAATAACAAGCCGGTAGCC R: TGGTTGTCTTTCAGCTCCAC | 118 | NM_001024860.1 | 1.96 | 200 | [32] |
| IFNG | Pro-inflammatory | F: AAGTTCTTGAACGGCAGCTC R: TTGGCGACAGGTCATTCATC | 130 | NM_001009803.1 | 1.91 | 500 | [30,31] |
| NRF2 | Redox system | F: GAGCCCAGTCTTCAATGCTC R: TCAGCCAGCTTGTCATTTTG | 171 | XM_015093345.2 | 1.97 | 200 | [30,31] |
| NFKB | Transcription factor | F: CTACACCTTGCCTGTGAGCA R: AAGGACACCAACAGCTCCAC | 173 | NM_001166184.1 | 1.93 | 300 | [30,31] |
| SOD1 | Antioxidant enzyme | F: CACCATCCACTTCGAGGCAA R: GCACTGGTACAGCCTTGTGT | 126 | NM_174615.2 | 2.06 | 200 | [35] |
| SOD2 | Antioxidant enzyme | F: GGATCCCCTGCAAGGAACAA R: TGGCCTTCAGATAATCGGGC | 110 | NM_201527.2 | 2.03 | 200 | [35] |
| CAT | Antioxidant enzyme | F: TTCCGTCCTTTATCCACAGC R: CCATTGGCATTAACCAGCTT | 199 | XM_004016396.5 | 2.04 | 300 | [30,31] |
| GPX1 | Antioxidant enzyme | F: GGCATCAGGAAAACGCCAAG R: GGGGACCACGTGATGAACTT | 217 | XM_004018462.5 | 1.98 | 300 | [30,31] |
| GPX2 | Antioxidant enzyme | F: ATTGAGAATGTGGCCTCGCT R: CCAGGGCGGACATACTTGAG | 179 | XM_004010720.5 | 2.07 | 300 | [30,31] |
| GPX4 | Antioxidant enzyme | F: GGGAGTAATGCGGAGATCAA R: CCACACAGCCGTTCTTATCA | 199 | XM_027970172.1 | 2.04 | 300 | [30,31] |
| Faecal Oocyst Count | SAINFOIN | SAINFOIN + PEG | p-Value |
|---|---|---|---|
| Log-transformed OPG 1 (Log-values ± SE 2 between parenthesis) | 19 (1.28 ± 0.65) | 219 (2.34 ± 0.65) | 0.28 |
| Lambs excreting oocyst (%) | |||
| >0 oocysts/g faeces | 50 | 83.3 | 0.21 |
| >10 oocysts/g faeces | 33.3 | 83.3 | 0.07 |
| >500 oocysts/g faeces | 16.7 | 33.3 | 0.50 |
| Treatment 2 | Tissue 2 | p-Value 3 | |||||
|---|---|---|---|---|---|---|---|
| Item (µm) 1 | SAINFOIN | SAINFOIN + PEG | Jejunal | Ileal | SE | Treatment | Tissue |
| Total inflammatory | 4.3 | 4.3 | 5.1 | 3.5 | 0.23 | 1.0 | <0.0001 |
| Total tissue destruction | 2.1 | 2.0 | 3.3 | 0.8 | 0.23 | 0.61 | <0.0001 |
| Total Tissue repair | 0.01 | 0.04 | 0.04 | 0.01 | 0.03 | 0.32 | 0.32 |
| ∑ HIS | 6.4 | 6.3 | 8.4 | 4.3 | 0.42 | 0.83 | <0.0001 |
| Cluster | Gene Members | Most Representative Variable | Proportion of Explained Cluster Variation | Proportion of Total Variation |
|---|---|---|---|---|
| 1 | TNFA, TGFB, GPX4, NFKB, GPX1, and SOD2 | TNFA | 0.69 | 0.35 |
| 2 | NRF2, IFNG, IL10, and SOD1 | NRF2 | 0.55 | 0.18 |
| 3 | CAT and GPX2 | CAT | 0.75 | 0.12 |
| Cluster | Gene Members | Most Representative Variable | Proportion of Explained Cluster Variation | Proportion of Total Variation |
|---|---|---|---|---|
| 1 | NFKB, GPX4, GPX1, TGFB, TNFA, and SOD1 | NFKB | 0.65 | 0.32 |
| 2 | IFNG, GPX2, and SOD2 | IFNG | 0.64 | 0.16 |
| 3 | CAT and IL10 | CAT | 0.71 | 0.12 |
| 4 | NRF2 | NRF2 | 1 | 0.08 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pelegrin-Valls, J.; Álvarez-Rodríguez, J.; Martín-Alonso, M.J.; Ramírez, G.A.; Baila, C.; Lobon, S.; Joy, M.; Serrano-Pérez, B. Effect of Maternal Dietary Condensed Tannins from Sainfoin (Onobrychis viciifolia) on Gut Health and Antioxidant-Immune Crosstalk in Suckling Lambs. Agriculture 2022, 12, 1694. https://doi.org/10.3390/agriculture12101694
Pelegrin-Valls J, Álvarez-Rodríguez J, Martín-Alonso MJ, Ramírez GA, Baila C, Lobon S, Joy M, Serrano-Pérez B. Effect of Maternal Dietary Condensed Tannins from Sainfoin (Onobrychis viciifolia) on Gut Health and Antioxidant-Immune Crosstalk in Suckling Lambs. Agriculture. 2022; 12(10):1694. https://doi.org/10.3390/agriculture12101694
Chicago/Turabian StylePelegrin-Valls, Jonathan, Javier Álvarez-Rodríguez, María José Martín-Alonso, Gustavo Adolfo Ramírez, Claudia Baila, Sandra Lobon, Margalida Joy, and Beatriz Serrano-Pérez. 2022. "Effect of Maternal Dietary Condensed Tannins from Sainfoin (Onobrychis viciifolia) on Gut Health and Antioxidant-Immune Crosstalk in Suckling Lambs" Agriculture 12, no. 10: 1694. https://doi.org/10.3390/agriculture12101694