
Viewpoint of Chitosan Application in Grapevine for Abiotic Stress/Disease Management towards More Resilient Viticulture Practices
 , ,
, ,  , , , , and
, , , , and {kind=link}
{kind=link}
Abstract
:1. Introduction
Devastating Trunk Diseases in Grapevine
2. Different Objectives for Chitosan Application in Grapevine
2.1. Disease Management
2.2. Induced Secondary Metabolism and Stress Management
3. Postharvest Management
4. The Need for Sustainable Methods to Replace the Chemicals Used in Vineyards
5. Future Prospects: An Opinion with Special Focus to Trunk Diseases Management
6. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Dash, M.; Chiellini, F.; Ottenbrite, R.M.; Chiellini, E. Chitosan-A versatile semi-synthetic polymer in biomedical applications. Prog. Polym. Sci. 2011, 36, 981–1014. [Google Scholar] [CrossRef]
- Vroman, I.; Tighzert, L. Biodegradable Polymers. Materials 2009, 2, 307–344. [Google Scholar] [CrossRef]
- Muzzarelli, R.A.A. Natural Chelating Polymers, Alginic Acid, Chitin, and Chitosan; Pergamon Press: Oxford, UK; New York, NY, USA,, 1973. [Google Scholar]
- Rinaudo, M. Chitin and chitosan: Properties and applications. Prog. Polym. Sci. 2006, 31, 603–632. [Google Scholar] [CrossRef]
- Shukla, S.K.; Mishra, A.K.; Arotiba, O.A.; Mamba, B.B. Chitosan-based nanomaterials: A state-of-the-art review. Int. J. Biol. Macromol. 2013, 59, 46–58. [Google Scholar] [CrossRef]
- Allan, C.R.; Hadwiger, L.A. The fungicidal effect of chitosan on fungi of varying cell wall composition. Exp. Mycol. 1979, 3, 285–287. [Google Scholar] [CrossRef]
- Kaya, M.; Bitim, B.; Mujtaba, M.; Koyuncu, T. Surface morphology of chitin highly related with the isolated body part of butterfly (Argynnis pandora). Int. J. Biol. Macromol. 2015, 81, 443–449. [Google Scholar] [CrossRef]
- Kaya, M.; Mujtaba, M.; Bulut, E.; Akyuz, B.; Zelencova, L.; Sofi, K. Fluctuation in physicochemical properties of chitins extracted from different body parts of honeybee. Carbohydr. Polym. 2015, 132, 9–16. [Google Scholar] [CrossRef]
- Kaya, M.; Sofi, K.; Sargin, I.; Mujtaba, M. Changes in physicochemical properties of chitin at developmental stages (larvae, pupa and adult) of Vespa crabro (wasp). Carbohydr. Polym. 2016, 145, 64–70. [Google Scholar] [CrossRef]
- Rinaudo, M. Main properties and current applications of some polysaccharides as biomaterials. Polym. Int. 2008, 57, 397–430. [Google Scholar] [CrossRef]
- Barber, M.S.; Bertram, R.E.; Ride, J.P. Chitin oligosaccharides elicit lignification in wounded wheat leaves. Physiol. Mol. Plant Pathol. 1989, 34, 3–12. [Google Scholar] [CrossRef]
- Chaouat, C. Design of New Formulation Systems for Depigmenting Active Ingredients, with a View to Their Use by the Cutaneous Route; Université Toulouse III-Paul Sabatier: Toulouse, France, 2013. [Google Scholar]
- Chirkov, S.N.; Surguchova, N.; Atabekov, J.G. Chitosan inhibits systemic infections caused by DNA†containing plant viruses. Arch. Phytopathol. Plant Prot. 1994, 29, 21–24. [Google Scholar] [CrossRef]
- Agbodjato, N.A.; Noumavo, P.A.; Adjanohoun, A.; Agbessi, L.; Baba-Moussa, L. Synergistic Effects of Plant Growth Promoting Rhizobacteria and Chitosan on In Vitro Seeds Germination, Greenhouse Growth, and Nutrient Uptake of Maize (Zea mays L.). Biotechnol. Res. Int. 2016, 2016, 20. [Google Scholar] [CrossRef] [PubMed]
- Akyuz, L.; Kaya, M.; Koc, B.; Mujtaba, M.; Ilk, S.; Labidi, J.; Salaberria, A.M.; Cakmak, Y.S.; Yildiz, A. Diatomite as a novel composite ingredient for chitosan film with enhanced physicochemical properties. Int. J. Biol. Macromol. 2017, 105 Pt 2, 1401–1411. [Google Scholar] [CrossRef] [PubMed]
- Kaya, M.; Akyuz, L.; Sargin, I.; Mujtaba, M.; Salaberria, A.M.; Labidi, J.; Cakmak, Y.S.; Koc, B.; Baran, T.; Ceter, T. Incorporation of sporopollenin enhances acid-base durability, hydrophobicity, and mechanical, antifungal and antioxidant properties of chitosan films. J. Ind. Eng. Chem. 2017, 47, 236–245. [Google Scholar] [CrossRef]
- Li, B.; Shi, Y.; Shan, C.; Zhou, Q.; Ibrahim, M.; Wang, Y.; Wu, G.; Li, H.; Xie, G.; Sun, G. Effect of chitosan solution on the inhibition of Acidovorax citrulli causing bacterial fruit blotch of watermelon. J. Sci. Food Agric. 2013, 93, 1010–1015. [Google Scholar] [CrossRef]
- Iriti, M.; Faoro, F. Bioactivity of Grape Chemicals for Human Health. Nat. Prod. Commun. 2009, 4, 1934578X0900400502. [Google Scholar] [CrossRef]
- OIV. World Vitiviniculture Situation; OIV: Dijon, France, 2017. [Google Scholar]
- Ferreira, R.B.; Monteiro, S.S.; Piçarra-Pereira, M.A.; Teixeira, A.R. Engineering grapevine for increased resistance to fungal pathogens without compromising wine stability. Trends Biotechnol. 2004, 22, 168–173. [Google Scholar] [CrossRef]
- Corio-Costet, M.-F.; Dufour, M.-C.; Cigna, J.; Abadie, P.; Chen, W.-J. Diversity and fitness of Plasmopara viticola isolates resistant to QoI fungicides. Eur. J. Plant Pathol. 2011, 129, 315–329. [Google Scholar] [CrossRef]
- Dufour, M.C.; Fontaine, S.; Montarry, J.; Corio-Costet, M.F. Assessment of fungicide resistance and pathogen diversity in Erysiphe necator using quantitative real-time PCR assays. Pest Manag. Sci. 2011, 67, 60–69. [Google Scholar] [CrossRef]
- Bony, S.; Gillet, C.; Bouchez, A.; Margoum, C.; Devaux, A. Genotoxic pressure of vineyard pesticides in fish: Field and mesocosm surveys. Aquat. Toxicol. 2008, 89, 197–203. [Google Scholar] [CrossRef]
- Komarek, M.; Cadkova, E.; Chrastny, V.; Bordas, F.; Bollinger, J.-C. Contamination of vineyard soils with fungicides: A review of environmental and toxicological aspects. Environ. Int. 2010, 36, 138–151. [Google Scholar] [CrossRef] [PubMed]
- Schreck, E.; Gontier, L.; Dumat, C.; Geret, F. Ecological and physiological effects of soil management practices on earthworm communities in French vineyards. Eur. J. Soil Biol. 2012, 52, 8–15. [Google Scholar] [CrossRef]
- Sharma, L.; Bohra, N.; Singh, R.K.; Marques, G. Potential of Entomopathogenic Bacteria and Fungi. In Microbes for Sustainable Insect Pest Management: An Eco-Friendly Approach-Volume 1; Khan, M.A., Ahmad, W., Eds.; Springer International Publishing: Cham, Switzerland, 2019; pp. 115–149. [Google Scholar]
- Jermini, M.; Blaise, P.; Gessler, C. Quantitative effect of leaf damage caused by downy mildew (Plasmopara viticola) on growth and yield quality of grapevine ‘Merlot’ (Vitis vinifera). Vitis: J. Grapevine Res. 2010, 49, 77–85. [Google Scholar]
- Petit, A.N.; Wojnarowiez, G.; Panon, M.L.; Baillieul, F.; Clément, C.; Fontaine, F.; Vaillant-Gaveau, N. Botryticides affect grapevine leaf photosynthesis without inducing defense mechanisms. Planta 2009, 229, 497–506. [Google Scholar] [CrossRef]
- Cesnik, H.B.; Gregorcic, A.; Cus, F. Pesticide residues in grapes from vineyards included in integrated pest management in Slovenia. Food Addit. Contam. Part A Chem. Anal. Control Expo. Risk Assess. 2008, 25, 438–443. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez Alvarez, M.; Noguerol-Pato, R.; Gonzalez-Barreiro, C.; Cancho-Grande, B.; Simal-Gandara, J. Changes of the sensorial attributes of white wines with the application of new anti-mildew fungicides under critical agricultural practices. Food Chem. 2012, 130, 139–146. [Google Scholar] [CrossRef]
- Gramaje, D.; Armengol, J. Fungal Trunk Pathogens in the Grapevine Propagation Process: Potential Inoculum Sources, Detection, Identification, and Management Strategies. Plant Dis. 2011, 95, 1040–1055. [Google Scholar] [CrossRef] [PubMed]
- Rubio, J.J.; Garzón, E. Las enfermedades de madera de vid como amenaza del sector vitícola. Rev. Winetech 2011, 2, 18–21. [Google Scholar]
- Hofstetter, V.; Buyck, B.; Croll, D.; Viret, O.; Couloux, A.; Gindro, K. What if esca disease of grapevine were not a fungal disease? Fungal Divers. 2012, 54, 51–67. [Google Scholar] [CrossRef]
- OIV. Measures Used to Prevent or Limit the Proliferation of Wood Diseases; OIV: Dijon, France, 2006. [Google Scholar]
- OIV. RESOLUTION CST 1/2008; OIV: Dijon, France, 2008. [Google Scholar]
- OIV. Grapevine Trunk Diseases; OIV: Dijon, France, 2016. [Google Scholar]
- Elmer, P.A.G.; Reglinski, T. Biosuppression of Botrytis cinerea in grapes. Plant Pathol. 2006, 55, 155–177. [Google Scholar] [CrossRef]
- Polesani, M.; Bortesi, L.; Ferrarini, A.; Zamboni, A.; Fasoli, M.; Zadra, C.; Lovato, A.; Pezzotti, M.; Delledonne, M.; Polverari, A. General and species-specific transcriptional responses to downy mildew infection in a susceptible (Vitis vinifera) and a resistant (V. riparia) grapevine species. BMC Genom. 2010, 11, 1471–2164. [Google Scholar] [CrossRef]
- Toffolatti, S.L.; Venturini, G.; Maffi, D.; Vercesi, A. Phenotypic and histochemical traits of the interaction between Plasmopara viticolaand resistant or susceptible grapevine varieties. BMC Plant Biol. 2012, 12, 124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delaunois, B.; Cordelier, S.; Conreux, A.; Clément, C.; Jeandet, P. Molecular engineering of resveratrol in plants. Plant Biotechnol. J. 2009, 7, 2–12. [Google Scholar] [CrossRef] [PubMed]
- Vidal, J.R.; Gomez, C.; Cutanda, M.C.; Shrestha, B.R.; Bouquet, A.; Thomas, M.R.; Torregrosa, L. Use of gene transfer technology for functional studies in grapevine. Aust. J. Grape Wine Res. 2010, 16, 138–151. [Google Scholar] [CrossRef]
- Hidangmayum, A.; Dwivedi, P.; Katiyar, D.; Hemantaranjan, A. Application of chitosan on plant responses with special reference to abiotic stress. Physiol. Mol. Biol. Plants 2019, 25, 313–326. [Google Scholar] [CrossRef]
- Sharif, R.; Mujtaba, M.; Ur Rahman, M.; Shalmani, A.; Ahmad, H.; Anwar, T.; Tianchan, D.; Wang, X. The Multifunctional Role of Chitosan in Horticultural Crops; A Review. Molecules 2018, 23, 872. [Google Scholar] [CrossRef]
- Mohamed, N.L.J.; Farmer, M.J.; Fromentin, J.; Béno, N.; Houot, V.; Milat, M.L.; Blein, J.P. Defense Responses in Grapevine Leaves Against Botrytis cinerea Induced by Application of a Pythium oligandrum Strain or Its Elicitin, Oligandrin, to Roots. Phytopathology 2007, 97, 611–620. [Google Scholar] [CrossRef] [PubMed]
- De Bona, G.S.; Vincenzi, S.; De Marchi, F.; Angelini, E.; Bertazzon, N. Chitosan induces delayed grapevine defense mechanisms and protects grapevine against Botrytis cinerea. J. Plant Dis. Prot. 2021, 128, 715–724. [Google Scholar] [CrossRef]
- Repka, V. Elicitor-Stimulated Induction of Defense Mechanisms and Defense Gene Activation in Grapevine Cell Suspension Cultures. Biol. Plant. 2001, 44, 555–565. [Google Scholar] [CrossRef]
- Ait Barka, E.; Eullaffroy, P.; Clement, C.; Vernet, G. Chitosan improves development, and protects Vitis vinifera L. against Botrytis cinerea. Plant Cell Rep. 2004, 22, 608–614. [Google Scholar] [CrossRef]
- Trotel-Aziz, P.; Couderchet, M.; Vernet, G.; Aziz, A. Chitosan Stimulates Defense Reactions in Grapevine Leaves and Inhibits Development of Botrytis Cinerea. Eur. J. Plant Pathol. 2006, 114, 405–413. [Google Scholar] [CrossRef]
- Król, E. Chitosan activity in an inhibition of in vitro growth of Phomopsis viticola and protection of grapevine canes against the pathogen. Phytopathol. Pol. 2006, 39, 155–162. [Google Scholar]
- Aziz, A.; Trotel-Aziz, P.; Conreux, A.; Dhuicq, L.; Jeandet, P.; Couderchet, M. Chitosan Induces Phytoalexin Synthesis, Chitinase and β-1,3-Glucanase Activities, and Resistance of Grapevine to Fungal Pathogens. In Macromolecules and Secondary Metabolites of Grapevine and Wine; Tec & doc: Gwabegar, Australia, 2007; pp. 83–88. [Google Scholar]
- Munoz, Z.; Moret, A.; Garces, S. Assessment of chitosan for inhibition of Colletotrichum sp. on tomatoes and grapes. Crop Prot. 2009, 28, 36–40. [Google Scholar] [CrossRef]
- Ferri, M.; Tassoni, A.; Franceschetti, M.; Righetti, L.; Naldrett, M.J.; Bagni, N. Chitosan treatment induces changes of protein expression profile and stilbene distribution in Vitis vinifera cell suspensions. Proteomics 2009, 9, 610–624. [Google Scholar] [CrossRef] [PubMed]
- Munoz, Z.; Moret, A. Sensitivity of Botrytis cinerea to chitosan and acibenzolar-S-methyl. Pest. Manag. Sci. 2010, 66, 974–979. [Google Scholar] [CrossRef]
- Reglinski, T.; Elmer, P.A.G.; Taylor, J.T.; Wood, P.N.; Hoyte, S.M. Inhibition of Botrytis cinerea growth and suppression of botrytis bunch rot in grapes using chitosan. Plant Pathol. 2010, 59, 882–890. [Google Scholar] [CrossRef]
- Iriti, M.; Vitalini, S.; DI Tommaso, G.; D’Amico, S.; Borgo, M.; Faoro, F. New chitosan formulation prevents grapevine powdery mildew infection and improves polyphenol content and free radical scavenging activity of grape and wine. Aust. J. Grape Wine Res. 2011, 17, 263–269. [Google Scholar] [CrossRef]
- Inchaya, P.; Sompong, M.; Wongkaew, S.; Athinuwat, D.; Buensanteai, N. Changes in salicylic acid in grapevine treated with chitosan and BTH against Sphaceloma ampelinum, the causal agent of grapevine anthracnose. Afr. J. Microbiol. Res. 2013, 7, 557–563. [Google Scholar]
- Romanazzi, G.; Murolo, S.; Feliziani, E. Effects of an innovative strategy to contain grapevine Bois noir: Field treatment with resistance inducers. Phytopathology 2013, 103, 785–791. [Google Scholar] [CrossRef]
- Xu, A.; Zhan, J.-C.; Huang, W.-D. Combined elicitation of chitosan and ultraviolet C enhanced stilbene production and expression of chitinase and β-1,3-glucanase in Vitis vinifera cell suspension cultures. Plant Cell Tissue Organ Cult. 2016, 124, 105–117. [Google Scholar] [CrossRef]
- Garde-Cerden, T.; Mancini, V.; Carrasco-Quiroz, M.; Servili, A.; Gutierrez-Gamboa, G.; Foglia, R.; Perez-Ãlvarez, E.P.; Romanazzi, G. Chitosan and Laminarin as Alternatives to Copper for Plasmopara viticola Control: Effect on Grape Amino Acid. J. Agric. Food Chem. 2017, 65, 7379–7386. [Google Scholar] [CrossRef] [PubMed]
- Bavaresco, L.; Zamboni, M.; Squeri, C.; Xu, S.; Abramowicz, A.; Lucini, L. Chitosan and grape secondary metabolites: A proteomics and metabolomics approach. BIO Web Conf. 2017, 9, 01004. [Google Scholar] [CrossRef]
- Lucini, L.; Baccolo, G.; Rouphael, Y.; Colla, G.; Bavaresco, L.; Trevisan, M. Chitosan treatment elicited defence mechanisms, pentacyclic triterpenoids and stilbene accumulation in grape (Vitis vinifera L.) bunches. Phytochemistry 2018, 156, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Aziz, A.; Trotel-Aziz, P.; Dhuicq, L.; Jeandet, P.; Couderchet, M.; Vernet, G. Chitosan oligomers and copper sulfate induce grapevine defense reactions and resistance to gray mold and downy mildew. Phytopathology 2006, 96, 1188–1194. [Google Scholar] [CrossRef] [Green Version]
- Meng, X.; Li, B.; Liu, J.; Tian, S. Physiological responses and quality attributes of table grape fruit to chitosan preharvest spray and postharvest coating during storage. Food Chem. 2008, 106, 501–508. [Google Scholar] [CrossRef]
- Gornik, K.; Wgrzesik, M.; Duda, B.R. The effect of chitosan on rooting of grapevine cuttings and on subsequent plant growth under drought and temperature stress. J. Fruit Ornam. Plant Res. 2008, 16, 333–343. [Google Scholar]
- Vitalini, S.F.F.; Di Tommaso, G.; Di Tommaso, D.; Borgo, M.; Piaggesi, A.; Fico, G.; Tomé, F.; Iriti, M. Effects of chitosan and phosphite treatments on total polyphenol content and antioxidant power of different grape berry tissues and wines. In Proceedings of the Workshop Induced Resistance in the Control of Plant Diseases: Effectiveness and Mechanisms of Action of a Sustainable Approach, Ancona, Italy, 18 June 2009. [Google Scholar]
- Santamaria, A.R.; Mulinacci, N.; Valletta, A.; Innocenti, M.; Pasqua, G. Effects of elicitors on the production of resveratrol and viniferins in cell cultures of Vitis vinifera L. cv Italia . J. Agric. Food Chem. 2011, 59, 9094–9101. [Google Scholar] [CrossRef]
- Ferri, M.; Dipalo, S.C.F.; Bagni, N.; Tassoni, A. Chitosan elicits mono-glucosylated stilbene production and release in fed-batch bioreactor cultures of grape cells. Food Chem. 2011, 124, 1473–1479. [Google Scholar] [CrossRef]
- Ruggiero, A.; Vitalini, S.; Burlini, N.; Bernasconi, S.; Iriti, M. Phytosterols in grapes and wine, and effects of agrochemicals on their levels. Food Chem. 2013, 141, 3473–3479. [Google Scholar] [CrossRef]
- Vitalini, S.; Ruggiero, A.; Rapparini, F.; Neri, L.; Tonni, M.; Iriti, M. The application of chitosan and benzothiadiazole in vineyard (Vitis vinifera L. cv Groppello Gentile) changes the aromatic profile and sensory attributes of wine. Food Chem. 2014, 162, 192–205. [Google Scholar] [CrossRef]
- Ferri, M.; Franceschetti, M.; Naldrett, M.J.; Saalbach, G.; Tassoni, A. Effects of chitosan on the protein profile of grape cell culture subcellular fractions. Electrophoresis 2014, 35, 1685–1692. [Google Scholar] [CrossRef] [PubMed]
- Tessarin, P.; Chinnici, F.; Donnini, S.; Liquori, E.; Riponi, C.; RombolÃ, A.D. Influence of canopy-applied chitosan on the composition of organic cv. Sangiovese and Cabernet Sauvignon berries and wines. Food Chem. 2016, 210, 512–519. [Google Scholar] [CrossRef] [PubMed]
- Portu, J.; Lopez, R.; Baroja, E.; Santamaria, P.; Garde-Cerdán, T. Improvement of grape and wine phenolic content by foliar application to grapevine of three different elicitors: Methyl jasmonate, chitosan, and yeast extract. Food Chem. 2016, 201, 213–221. [Google Scholar] [CrossRef] [PubMed]
- El-kenawy, M.A. Effect of Chitosan, Salicylic Acid and Fulvic Acid on Vegetative Growth, Yield and Fruit Quality of Thompson Seedless Grapevines. Egypt. J. Hortic. 2017, 44, 45–59. [Google Scholar] [CrossRef]
- Gutiérrez-Gamboa, G.; Pérez-Álvarez, E.P.; Rubio-Bretón, P.; Garde-Cerdán, T. Changes on grape volatile composition through elicitation with methyl jasmonate, chitosan, and a yeast extract in Tempranillo (Vitis vinifera L.) grapevines. Sci. Hortic. 2019, 244, 257–262. [Google Scholar] [CrossRef]
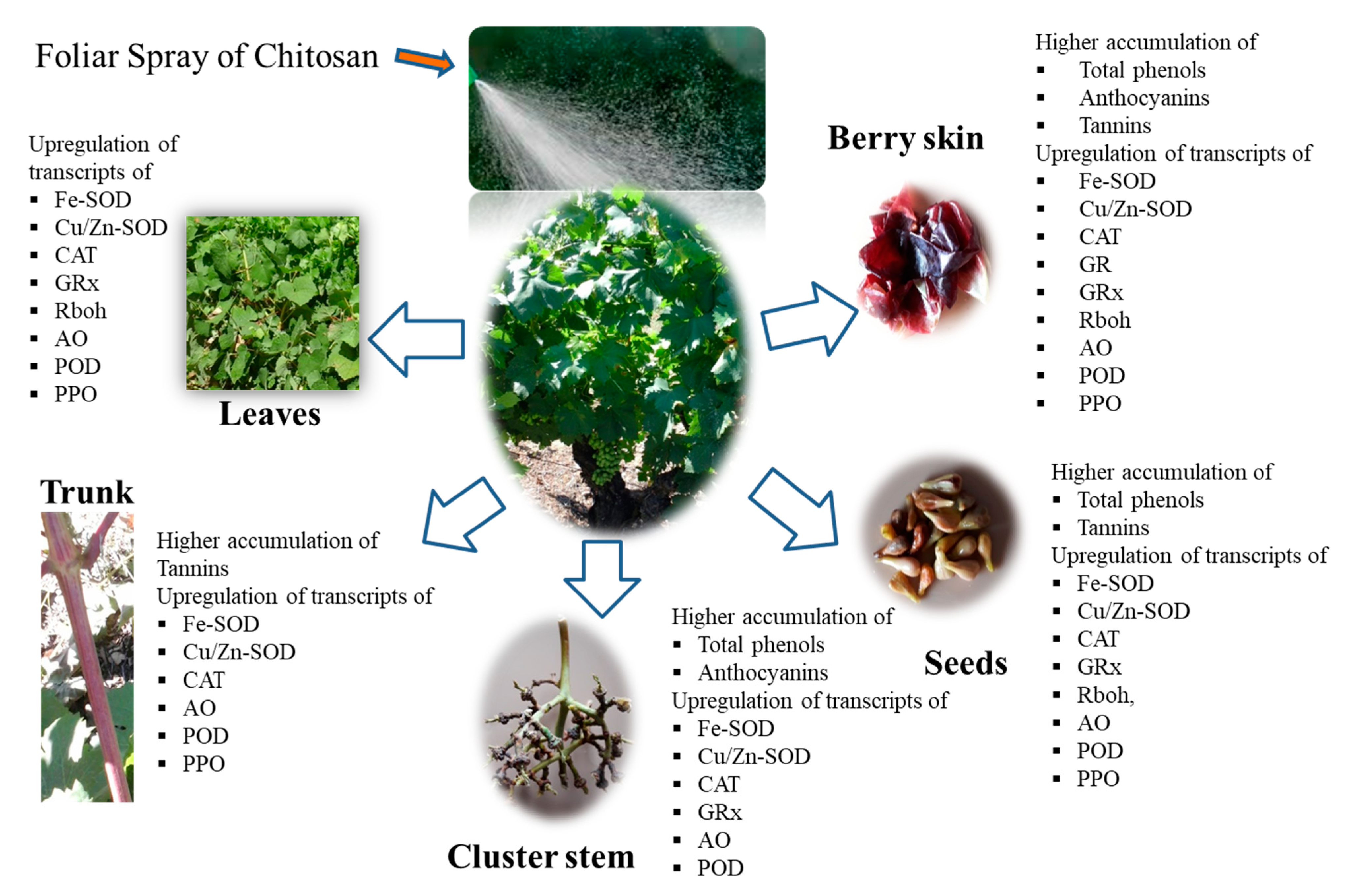
- Singh, R.K.; Soares, B.; Goufo, P.; Castro, I.; Cosme, F.; Pinto-Sintra, A.L.; Inês, A.; Oliveira, A.A.; Falco, V. Chitosan Upregulates the Genes of the ROS Pathway and Enhances the Antioxidant Potential of Grape (Vitis vinifera L. ‘Touriga Franca’ and ‘Tinto Cäo’) Tissues. Antioxidants 2019, 8, 525. [Google Scholar] [CrossRef]
- Singh, R.K.; Martins, V.; Soares, B.; Castro, I.; Falco, V. Chitosan Application in Vineyards (Vitis vinifera L. cv. Tinto Cão) Induces Accumulation of Anthocyanins and Other Phenolics in Berries, Mediated by Modifications in the Transcription of Secondary Metabolism Genes. Int. J. Mol. Sci. 2020, 21, 306. [Google Scholar] [CrossRef]
- Silva, V.; Singh, R.K.; Gomes, N.; Soares, B.G.A.; Silva, A.; Falco, V.; Capita, R.; Alonso-Calleja, C.; Pereira, J.E.; Amaral, J.S.; et al. Comparative Insight upon Chitosan Solution and Chitosan Nanoparticles Application on the Phenolic Content, Antioxidant and Antimicrobial Activities of Individual Grape Components of Sousao Variety. Antioxidants 2020, 9, 178. [Google Scholar] [CrossRef]
- Romanazzi, G.; Gabler, F.M.; Smilanick, J.L. Preharvest Chitosan and Postharvest UV Irradiation Treatments Suppress Gray Mold of Table Grapes. Plant Dis. 2006, 90, 445–450. [Google Scholar] [CrossRef]
- Romanazzi, G. Chitosan Treatment for the Control of Postharvest Decay of Table Grapes, Strawberries and Sweet Cherries. In Fresh Produce: New Trends in Postharvest Management of Fresh Produce; Sivakumar, D., Ed.; University of Pretoria: Pretoria, South Africa, 2009; Volume 4, pp. 111–115. ISBN 978-4-903313-40-5. [Google Scholar]
- Romanazzi, G.; Gabler, F.M.; Margosan, D.; Mackey, B.E.; Smilanick, J.L. Effect of chitosan dissolved in different acids on its ability to control postharvest gray mold of table grape. Phytopathology 2009, 99, 1028–1036. [Google Scholar] [CrossRef]
- Meng, X.-H.; Qin, G.-Z.; Tian, S.-P. Influences of preharvest spraying Cryptococcus laurentii combined with postharvest chitosan coating on postharvest diseases and quality of table grapes in storage. LWT-Food Sci. Technol. 2010, 43, 596–601. [Google Scholar] [CrossRef]
- Wang Simeng, R.Y.; Junyu, H.; Linchuan, F.; Houyu, L. Effects of Single and Complex Chitosan Coating on Grape Preservative; College of Agriculture, Guizhou University: Guiyang, China, 2010. [Google Scholar]
- Feliziani, E.; Smilanick, J.L.; Margosan, D.A.; Mansour, M.F.; Romanazzi, G.; Gu, S.; Gohil, H.L.; Ames, Z.R. Preharvest Fungicide, Potassium Sorbate, or Chitosan Use on Quality and Storage Decay of Table Grapes. Plant Dis. 2013, 97, 307–314. [Google Scholar] [CrossRef]
- Melo, N.F.C.B.; MendonçaSoares, B.L.d.; Diniz, K.M.; Leal, C.F.; Canto, D.; Flores, M.A.; Tavares-Filho, J.H.d.C.; Galembeck, A.; Stamford, T.M.; Stamford-Arnaud, T.M.; et al. Effects of fungal chitosan nanoparticles as eco-friendly edible coatings on the quality of postharvest table grapes. Postharvest Biol. Technol. 2018, 139, 56–66. [Google Scholar] [CrossRef]
- Sharma, L.; Oliveira, I.; Raimundo, F.; Torres, L.; Marques, G. Soil Chemical Properties Barely Perturb the Abundance of Entomopathogenic Fusarium oxysporum: A Case Study Using a Generalized Linear Mixed Model for Microbial Pathogen Occurrence Count Data. Pathogens 2018, 7, 89. [Google Scholar] [CrossRef] [PubMed]
- Sharma, L.; Bohra, N.; Rajput, V.D.; Quiroz-Figueroa, F.R.; Singh, R.K.; Marques, G. Advances in Entomopathogen Isolation: A Case of Bacteria and Fungi. Microorganisms 2021, 9, 16. [Google Scholar] [CrossRef]
- Sergazina, M.V.L.; Llompart, M.; Dagnac, T. Occurrence of Fungicides in Vineyard and the Surrounding Environment. Molecules 2021, 26, 6152. [Google Scholar] [CrossRef]
- Sharma, L.; Gonsalves, F.; Oliveira, I.; Torres, L.; Marques, G. Insect-associated fungi from naturally mycosed vine mealybug Planococcus ficus (Signoret) (Hemiptera: Pseudococcidae). Biocontrol. Sci. Technol. 2018, 28, 122–141. [Google Scholar] [CrossRef]
- (EC), E.P.R., Regulation (EC) No 1107/2009 of the European Parliament and of the Council of 21 October 2009 Concerning the Placing of Plant Protection Products on the Market and Repealing Council Directives 79/117/EEC and 91/414/EEC. 2009. Available online: https://www.legislation.gov.uk/eur/2009/1107/contents?view=plain (accessed on 28 August 2022).
- Sharma, L.; Oliveira, I.; Gonsalves, F.; Raimundo, F.; Singh, R.K.; Torres, L.; Marques, G. Effect of Soil Chemical Properties on the Occurrence and Distribution of Entomopathogenic Fungi in Portuguese Grapevine Fields. Pathogens 2021, 10, 137. [Google Scholar] [CrossRef]
- Silva-Castro, I.; Martín-García, J.; Diez, J.J.; Flores-Pacheco, J.A.; Martín-Gil, J.; Martín-Ramos, P. Potential control of forest diseases by solutions of chitosan oligomers, propolis and nanosilver. Eur. J. Plant Pathol. 2018, 150, 401–411. [Google Scholar] [CrossRef]
- Saharan, V.; Sharma, G.; Yadav, M.; Choudhary, M.K.; Sharma, S.S.; Pal, A.; Raliya, R.; Biswas, P. Synthesis and in vitro antifungal efficacy of Cu–chitosan nanoparticles against pathogenic fungi of tomato. Int. J. Biol. Macromol. 2015, 75, 346–353. [Google Scholar] [CrossRef]
- Saharan, V.; Mehrotra, A.; Khatik, R.; Rawal, P.; Sharma, S.S.; Pal, A. Synthesis of chitosan based nanoparticles and their in vitro evaluation against phytopathogenic fungi. Int. J. Biol. Macromol. 2013, 62, 677–683. [Google Scholar] [CrossRef]
- Matei, P.M.; Martín-Ramos, P.; Sánchez-Báscones, M.; Hernández-Navarro, S.; Correa-Guimaraes, A.; Navas-Gracia, L.M.; Rufino, C.A.; Ramos-Sánchez, M.C.; Martín-Gil, J. Synthesis of Chitosan Oligomers/Propolis/Silver Nanoparticles Composite Systems and Study of Their Activity against Diplodia seriata. Int. J. Polym. Sci. 2015, 2015, 864729. [Google Scholar] [CrossRef]
- Sathiyabama, M.; Parthasarathy, R. Biological preparation of chitosan nanoparticles and its in vitro antifungal efficacy against some phytopathogenic fungi. Carbohydr. Polym. 2016, 151, 321–325. [Google Scholar] [CrossRef] [PubMed]
- Jain, A.; Sarsaiya, S.; Wu, Q.; Lu, Y.; Shi, J. A review of plant leaf fungal diseases and its environment speciation. Bioengineered 2019, 10, 409–424. [Google Scholar] [CrossRef]
- Xing, K.; Shen, X.; Zhu, X.; Ju, X.; Miao, X.; Tian, J.; Feng, Z.; Peng, X.; Jiang, J.; Qin, S. Synthesis and in vitro antifungal efficacy of oleoyl-chitosan nanoparticles against plant pathogenic fungi. Int. J. Biol. Macromol. 2016, 82, 830–836. [Google Scholar] [CrossRef] [PubMed]
- Kumaraswamy, R.V.; Kumari, S.; Choudhary, R.C.; Pal, A.; Raliya, R.; Biswas, P.; Saharan, V. Engineered chitosan based nanomaterials: Bioactivities, mechanisms and perspectives in plant protection and growth. Int. J. Biol. Macromol. 2018, 113, 494–506. [Google Scholar] [CrossRef] [PubMed]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Singh, R.K.; Ruiz-May, E.; Rajput, V.D.; Minkina, T.; Gómez-Peraza, R.L.; Verma, K.K.; Shekhawat, M.S.; Pinto, C.; Falco, V.; Quiroz-Figueroa, F.R. Viewpoint of Chitosan Application in Grapevine for Abiotic Stress/Disease Management towards More Resilient Viticulture Practices. Agriculture 2022, 12, 1369. https://doi.org/10.3390/agriculture12091369
Singh RK, Ruiz-May E, Rajput VD, Minkina T, Gómez-Peraza RL, Verma KK, Shekhawat MS, Pinto C, Falco V, Quiroz-Figueroa FR. Viewpoint of Chitosan Application in Grapevine for Abiotic Stress/Disease Management towards More Resilient Viticulture Practices. Agriculture. 2022; 12(9):1369. https://doi.org/10.3390/agriculture12091369
Chicago/Turabian StyleSingh, Rupesh Kumar, Eliel Ruiz-May, Vishnu D. Rajput, Tatiana Minkina, Rosa Luz Gómez-Peraza, Krishan K. Verma, Mahipal S. Shekhawat, Catia Pinto, Virgilio Falco, and Francisco Roberto Quiroz-Figueroa. 2022. "Viewpoint of Chitosan Application in Grapevine for Abiotic Stress/Disease Management towards More Resilient Viticulture Practices" Agriculture 12, no. 9: 1369. https://doi.org/10.3390/agriculture12091369