d-Cysteine-Induced Rapid Root Abscission in the Water Fern Azolla Pinnata: Implications for the Linkage between d-Amino Acid and Reactive Sulfur Species (RSS) in Plant Environmental Responses


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
3. Results
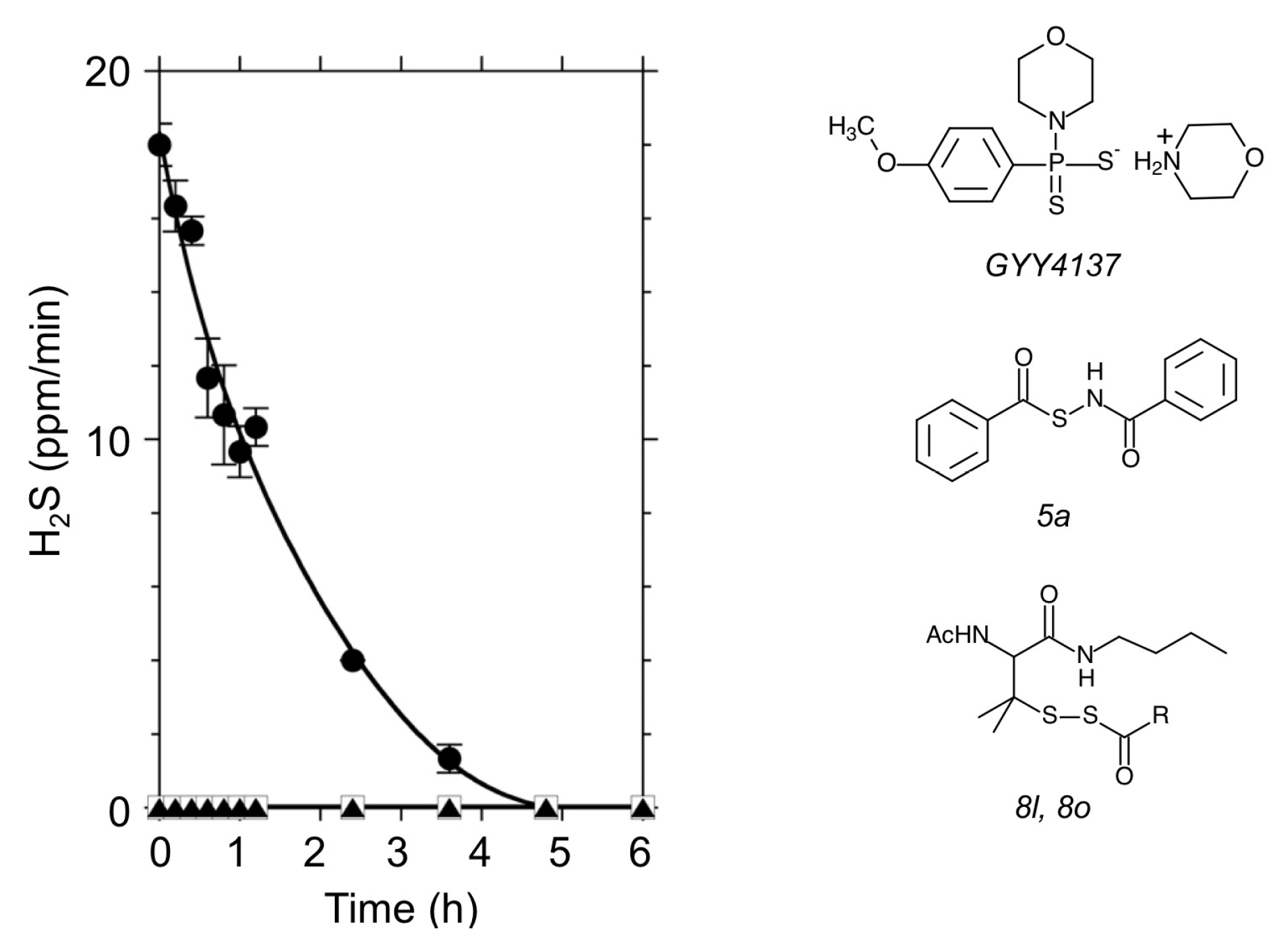
3.1. Effects of H2S Donors on the Root Abscission in Azolla
3.2. Effects of Polysulfides on the Root Abscission


3.3. d-Cysteine-Induced Root Abscission
4. Discussion
4.1. Application of Chemical H2S Donors for Inducing Root Abscission
4.2. Effects of Polysulfides (Na2Sn) on the Root Abscission
4.3. d-Cysteine is a Novel Inducer of Rapid Root Abscsision in Azolla
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Yamasaki, H. The NO world for plants: Achieving balance in an open system. Plant Cell Environ. 2005, 28, 78–84. [Google Scholar] [CrossRef]
- Sakihama, Y.; Murakami, S.; Yamasaki, H. Involvement of nitric oxide in the mechanism for stomatal opening in Vicia faba leaves. Biol. Plant. 2003, 46, 117–119. [Google Scholar] [CrossRef]
- Antoniou, C.; Savvides, A.; Christou, A.; Fotopoulos, V. Unravelling chemical priming machinery in plants: The role of reactive oxygen–nitrogen–sulfur species in abiotic stress tolerance enhancement. Curr. Opin. Plant Biol. 2016, 33, 101–107. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhang, L.; Dong, F.; Gao, J.; Galbraith, D.W.; Song, C.-P. Hydrogen peroxide is involved in abscisic acid-induced stomatal closure in Vicia faba. Plant Physiol. 2001, 126, 1438–1448. [Google Scholar] [CrossRef] [PubMed]
- Quan, L.J.; Zhang, B.; Shi, W.W.; Li, H.Y. Hydrogen peroxide in plants: A versatile molecule of the reactive oxygen species network. J. Integr. Plant Biol. 2008, 50, 2–18. [Google Scholar] [CrossRef] [PubMed]
- Neill, S.J.; Desikan, R.; Clarke, A.; Hurst, R.D.; Hancock, J.T. Hydrogen peroxide and nitric oxide as signalling molecules in plants. J. Exp. Bot. 2002, 53, 1237–1247. [Google Scholar] [CrossRef] [PubMed]
- Desikan, R.; Griffiths, R.; Hancock, J.; Neill, S. A new role for an old enzyme: Nitrate reductase-mediated nitric oxide generation is required for abscisic acid-induced stomatal closure in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2002, 99, 16314–16318. [Google Scholar] [CrossRef] [PubMed]
- Desikan, R.; Cheung, M.K.; Bright, J.; Henson, D.; Hancock, J.T.; Neill, S.J. ABA, hydrogen peroxide and nitric oxide signalling in stomatal guard cells. J. Exp. Bot. 2004, 55, 205–212. [Google Scholar] [CrossRef]
- Ozgur, R.; Uzilday, B.; Iwata, Y.; Koizumi, N.; Turkan, I. Interplay between the unfolded protein response and reactive oxygen species: A dynamic duo. J. Exp. Bot. 2018, 69, 3333–3345. [Google Scholar] [CrossRef]
- Giles, G.I.; Tasker, K.M.; Jacob, C. Hypothesis: The role of reactive sulfur species in oxidative stress. Free Radic. Biol. Med. 2001, 31, 1279–1283. [Google Scholar] [CrossRef]
- Gruhlke, M.C.H.; Slusarenko, A.J. The biology of reactive sulfur species (RSS). Plant Physiol. Biochem. 2012, 59, 98–107. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Rose, P.; Moore, P.K. Hydrogen sulfide and cell signaling. Annu. Rev. Pharmacol. Toxicol. 2011, 51, 169–187. [Google Scholar] [CrossRef] [PubMed]
- Lisjak, M.; Srivastava, N.; Teklic, T.; Civale, L.; Lewandowski, K.; Wilson, I.; Wood, M.E.; Whiteman, M.; Hancock, J.T. A novel hydrogen sulfide donor causes stomatal opening and reduces nitric oxide accumulation. Plant Physiol. Biochem. 2010, 48, 931–935. [Google Scholar] [CrossRef] [PubMed]
- Lisjak, M.; Teklic, T.; Wilson, I.D.; Whiteman, M.; Hancock, J.T. Hydrogen sulfide: Environmental factor or signalling molecule? Plant Cell Environ. 2013, 36, 1607–1616. [Google Scholar] [CrossRef] [PubMed]
- De Kok, L.; Bosma, W.; Maas, F.; Kuiper, P. The effect of short-term H2S fumigation on water-soluble sulphydryl and glutathione levels in spinach. Plant Cell Environ. 1985, 8, 189–194. [Google Scholar] [CrossRef]
- Zhang, H.; Ye, Y.-K.; Wang, S.-H.; Luo, J.-P.; Tang, J.; Ma, D.-F. Hydrogen sulfide counteracts chlorophyll loss in sweetpotato seedling leaves and alleviates oxidative damage against osmotic stress. Plant Growth Regul. 2009, 58, 243–250. [Google Scholar] [CrossRef]
- Min, Y.; Qin, B.-P.; Ping, W.; Li, M.-L.; Chen, L.-L.; Chen, L.-T.; Sun, A.-Q.; Wang, Z.-L.; Yin, Y.-P. Foliar application of sodium hydrosulfide (NaHS), a hydrogen sulfide (H2S) donor, can protect seedlings against heat stress in wheat (Triticum aestivum L.). J. Integr. Agric. 2016, 15, 2745–2758. [Google Scholar]
- Kaya, C.; Ashraf, M.; Akram, N.A. Hydrogen sulfide regulates the levels of key metabolites and antioxidant defense system to counteract oxidative stress in pepper (Capsicum annuum L.) plants exposed to high zinc regime. Environ. Sci. Pollut. Res. 2018, 25, 1–7. [Google Scholar] [CrossRef]
- Cohen, M.F.; Sakihama, Y.; Takagi, Y.C.; Ichiba, T.; Yamasaki, H. Synergistic effect of deoxyanthocyanins from symbiotic fern Azolla spp. on hrmA gene induction in the cyanobacterium Nostoc punctiforme. Mol. Plant Microbe Interact. 2002, 15, 875–882. [Google Scholar] [CrossRef]
- Gurung, S.; Cohen, M.F.; Fukuto, J.; Yamasaki, H. Polyamine-induced rapid root abscission in Azolla pinnata. J. Amino Acids 2012, 2012, 493209. [Google Scholar] [CrossRef]
- Gurung, S.; Cohen, M.F.; Yamasaki, H. Azide-dependent nitric oxide emission from the water fern Azolla pinnata. Russ. J. Plant Physiol. 2014, 61, 543–547. [Google Scholar] [CrossRef]
- Lumpkin, T.A.; Plucknett, D.L. Azolla: Botany, physiology, and use as a green manure. Econ. Bot. 1980, 34, 111–153. [Google Scholar] [CrossRef]
- Leavitt, R.G. The root-hairs, cap, and sheath of Azolla. Bot. Gaz. 1902, 34, 414–419. [Google Scholar] [CrossRef]
- Gunning, B.; Hardham, A.; Hughes, J. Evidence for initiation of microtubules in discrete regions of the cell cortex in Azolla root-tip cells, and an hypothesis on the development of cortical arrays of microtubules. Planta 1978, 143, 161–179. [Google Scholar] [PubMed]
- Gunning, B.; Hughes, J.; Hardham, A. Formative and proliferative cell divisions, cell differentiation, and developmental changes in the meristem of Azolla roots. Planta 1978, 143, 121–144. [Google Scholar] [CrossRef]
- Sela, M.; Tel-Or, E.; Fritz, E.; Huttermann, A. Localization and toxic effects of cadmium, copper, and uranium in Azolla. Plant Physiol. 1988, 88, 30–36. [Google Scholar] [CrossRef]
- Cohen, M.F.; Yamasaki, H.; Mazzola, M. Bioremediation of soils by plant-microbe systems. Int. J. Green Energy 2004, 1, 301–312. [Google Scholar] [CrossRef]
- Rao, H. The structure and life-history of Azolla pinnata R. Brown with remarks on the fossil history of the hydropterideæ. Proc. Plant Sci. 1935, 2, 175–201. [Google Scholar]
- Kitoh, S.; Shiomi, N.; Uheda, E. The growth and nitrogen fixation of Azolla filiculoides Lam. in polluted water. Aquat Bot. 1993, 46, 129–139. [Google Scholar] [CrossRef]
- Uheda, E.; Kitoh, S. Rapid shedding of roots from Azolla filiculoides plants in response to inhibitors of respiration. Plant Cell Physiol. 1994, 35, 37–43. [Google Scholar]
- Yamasaki, H.; Sakihama, Y.; Takahashi, S. An alternative pathway for nitric oxide production in plants: New features of an old enzyme. Trends Plant Sci. 1999, 4, 128–129. [Google Scholar] [CrossRef]
- Lundberg, J.O.; Weitzberg, E.; Gladwin, M.T. The nitrate–nitrite–nitric oxide pathway in physiology and therapeutics. Nat. Rev. Drug Discov. 2008, 7, 156. [Google Scholar] [CrossRef] [PubMed]
- Yamasaki, H.; Watanabe, N.S.; Fukuto, J.; Cohen, M.F. Nitrite-dependent nitric oxide production pathway: Diversity of NO production systems. In Studies on Pediatric Disorders, Oxidative Stress in Applied Basic Research and Clinical Practice; Tsukahara, H., Kaneko, K., Eds.; Springer: New York, NY, USA, 2014; pp. 35–54. [Google Scholar]
- Cohen, M.F.; Gurung, S.; Birarda, G.; Holman, H.Y.N.; Yamasaki, H. Bimodal effect of hydrogen peroxide and oxidative events in nitrite-induced rapid root abscission by the water fern Azolla pinnata. Front. Plant Sci. 2015, 6, 518. [Google Scholar] [CrossRef]
- Cohen, M.F.; Gurung, S.; Fukuto, J.M.; Yamasaki, H. Controlled free radical attack in the apoplast: A hypothesis for roles of O, N and S species in regulatory and polysaccharide cleavage events during rapid abscission by Azolla. Plant Sci. 2014, 217, 120–126. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Whiteman, M.; Guan, Y.Y.; Neo, K.L.; Cheng, Y.; Lee, S.W.; Zhao, Y.; Baskar, R.; Tan, C.-H.; Moore, P.K. Characterization of a novel, water-soluble hydrogen sulfide-releasing molecule (GYY4137). Circulation 2008, 117, 2351–2360. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Bhushan, S.; Yang, C.; Otsuka, H.; Stein, J.D.; Pacheco, A.; Peng, B.; Devarie-Baez, N.O.; Aguilar, H.C.; Lefer, D.J. Controllable hydrogen sulfide donors and their activity against myocardial ischemia-reperfusion injury. ACS Chem. Biol. 2013, 8, 1283–1290. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Wang, H.; Xian, M. Cysteine-activated hydrogen sulfide (H2S) donors. J. Am. Chem. Soc. 2010, 133, 15–17. [Google Scholar] [CrossRef] [PubMed]
- Yamasaki, H.; Cohen, M.F. Biological consilience of hydrogen sulfide and nitric oxide in plants: Gases of primordial earth linking plant, microbial and animal physiologies. Nitric Oxide 2016, 55, 91–100. [Google Scholar] [CrossRef]
- Kimura, H. Signaling by hydrogen sulfide (H2S) and polysulfides (H2Sn) in the central nervous system. Neurochem. Int. 2019, 126, 118–125. [Google Scholar] [CrossRef]
- Takata, T.; Tsukuda, A.; Tsuchiya, Y.; Akaike, T.; Watanabe, Y. The active-site cysteine residue of Ca2+/calmodulin-dependent protein kinase I is protected from irreversible modification via generation of polysulfidation. Nitric Oxide 2019, 86, 68–75. [Google Scholar] [CrossRef]
- Ihara, H.; Kasamatsu, S.; Kitamura, A.; Nishimura, A.; Tsutsuki, H.; Ida, T.; Ishizaki, K.; Toyama, T.; Yoshida, E.; Hamid, H.A. Exposure to electrophiles impairs reactive persulfide-dependent redox signaling in neuronal cells. Chem. Res. Toxicol. 2017, 30, 1673–1684. [Google Scholar] [CrossRef] [PubMed]
- Akaike, T.; Ida, T.; Wei, F.-Y.; Nishida, M.; Kumagai, Y.; Alam, M.M.; Ihara, H.; Sawa, T.; Matsunaga, T.; Kasamatsu, S. Cysteinyl-tRNA synthetase governs cysteine polysulfidation and mitochondrial bioenergetics. Nat. Commun. 2017, 8, 1177. [Google Scholar] [CrossRef] [PubMed]
- Shibuya, N.; Koike, S.; Tanaka, M.; Ishigami-Yuasa, M.; Kimura, Y.; Ogasawara, Y.; Fukui, K.; Nagahara, N.; Kimura, H. A novel pathway for the production of hydrogen sulfide from D-cysteine in mammalian cells. Nat. Commun. 2013, 4, 1366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papenbrock, J.; Riemenschneider, A.; Kamp, A.; Schulz-Vogt, H.N.; Schmidt, A. Characterization of cysteine-degrading and H2S-releasing enzymes of higher plants-from the field to the test tube and back. Plant Biol. 2007, 9, 582–588. [Google Scholar] [CrossRef] [PubMed]
- Azolla filiculoides Genome Encoding Azfi_s0002.g001593. Available online: https://www.fernbase.org/jbrowse_fernbase/?data=data%2Fjson%2FAzolla_asm_v1.1&loc=Azfi_s0002%3A9528035..9533485&tracks=DNA%2CGene%20models-high%20confidence&highlight= (accessed on 17 September 2019).
- Álvarez, C.; García, I.; Moreno, I.; Pérez-Pérez, M.E.; Crespo, J.L.; Romero, L.C.; Gotor, C. Cysteine-generated sulfide in the cytosol negatively regulates autophagy and modulates the transcriptional profile in Arabidopsis. Plant Cell 2012, 24, 4621–4634. [Google Scholar] [CrossRef] [PubMed]
- Li, F.-W.; Brouwer, P.; Carretero-Paulet, L.; Cheng, S.; De Vries, J.; Delaux, P.-M.; Eily, A.; Koppers, N.; Kuo, L.-Y.; Li, Z. Fern genomes elucidate land plant evolution and cyanobacterial symbioses. Nat. Plants 2018, 4, 460. [Google Scholar] [CrossRef] [PubMed]
- Eily, A.N.; Pryer, K.M.; Li, F.W. A first glimpse at genes important to the Azolla-Nostoc symbiosis. Symbiosis 2019, 78, 149–162. [Google Scholar] [CrossRef]
- Corpas, F.J.; Chaki, M.; Fernandez-Ocana, A.; Valderrama, R.; Palma, J.M.; Carreras, A.; Begara-Morales, J.C.; Airaki, M.; del Río, L.A.; Barroso, J.B. Metabolism of reactive nitrogen species in pea plants under abiotic stress conditions. Plant Cell Physiol. 2008, 49, 1711–1722. [Google Scholar] [CrossRef] [PubMed]
- Yamasaki, H.; Watanabe, N.S.; Sakihama, Y.; Cohen, M.F. An overview of methods in plant nitric oxide (NO) research: Why do we always need to use multiple methods? In Plant Nitric Oxide: Methods and Protocols; Gupta, K.J., Ed.; Springer: New York, NY, USA, 2016; pp. 1–14. [Google Scholar]







© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yamasaki, H.; Ogura, M.P.; Kingjoe, K.A.; Cohen, M.F. d-Cysteine-Induced Rapid Root Abscission in the Water Fern Azolla Pinnata: Implications for the Linkage between d-Amino Acid and Reactive Sulfur Species (RSS) in Plant Environmental Responses. Antioxidants 2019, 8, 411. https://doi.org/10.3390/antiox8090411
Yamasaki H, Ogura MP, Kingjoe KA, Cohen MF. d-Cysteine-Induced Rapid Root Abscission in the Water Fern Azolla Pinnata: Implications for the Linkage between d-Amino Acid and Reactive Sulfur Species (RSS) in Plant Environmental Responses. Antioxidants. 2019; 8(9):411. https://doi.org/10.3390/antiox8090411
Chicago/Turabian StyleYamasaki, Hideo, Masahiro P. Ogura, Katsumi A. Kingjoe, and Michael F. Cohen. 2019. "d-Cysteine-Induced Rapid Root Abscission in the Water Fern Azolla Pinnata: Implications for the Linkage between d-Amino Acid and Reactive Sulfur Species (RSS) in Plant Environmental Responses" Antioxidants 8, no. 9: 411. https://doi.org/10.3390/antiox8090411