Regulation of Hydrogen Sulfide Metabolism by Nitric Oxide Inhibitors and the Quality of Peaches during Cold Storage


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Treatments
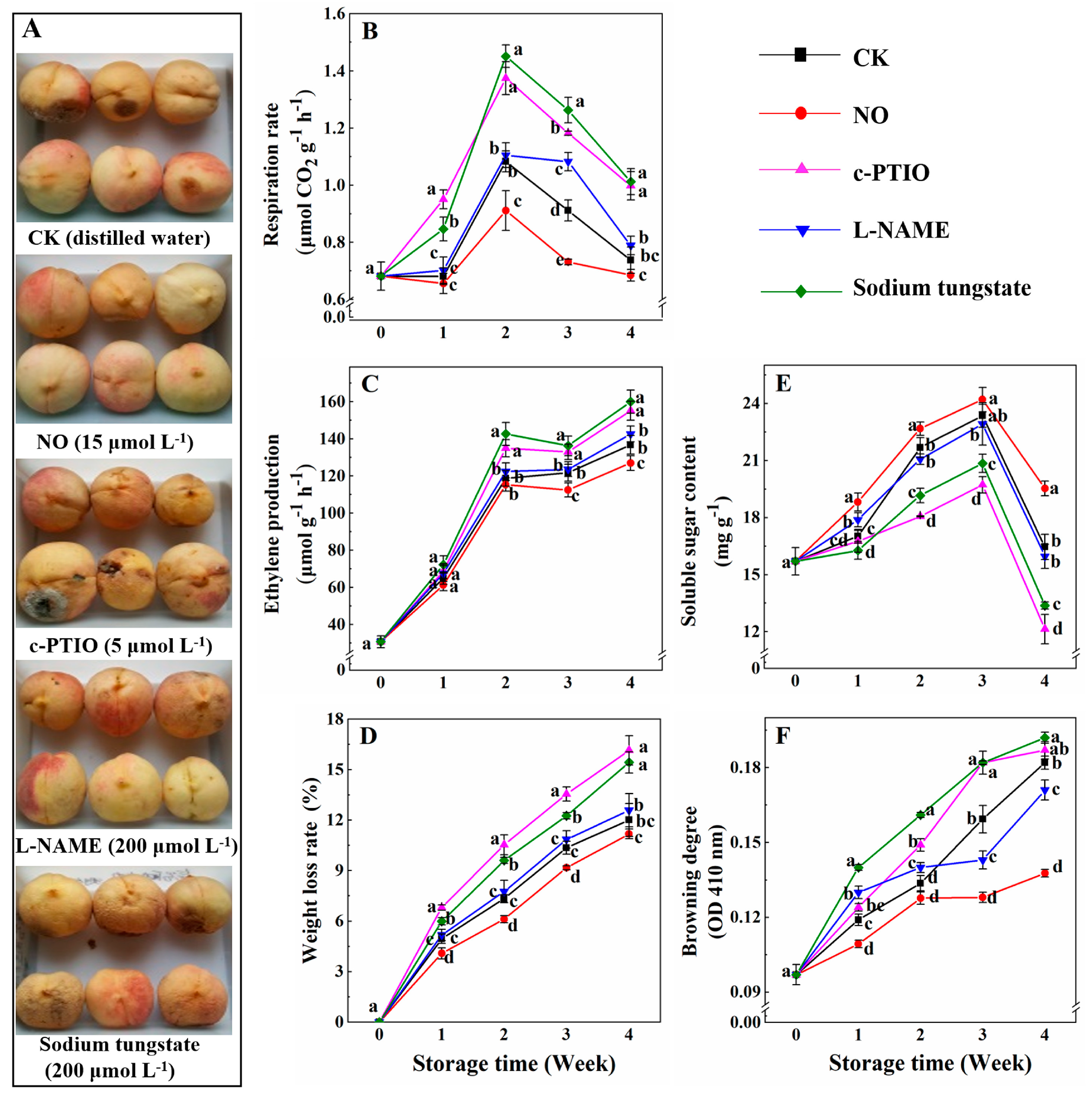
2.2. Determination of the Rates of Respiration, Ethylene Production, Water Loss, the Content of Soluble Sugar, and Browning Degree
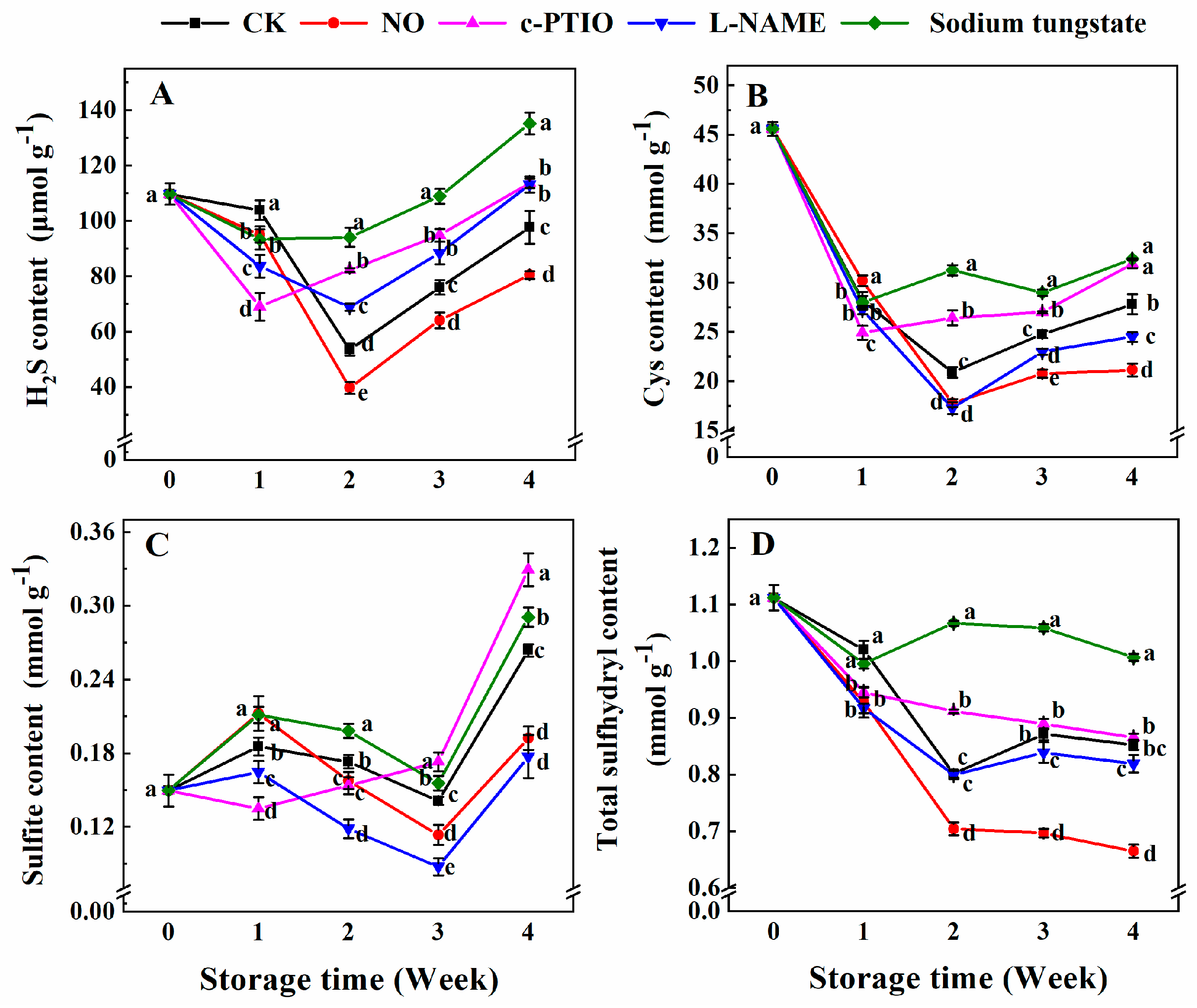
2.3. Determination of the Contents of Endogenous H2S, Total Sulfhydryl, Cys, and Sulfite
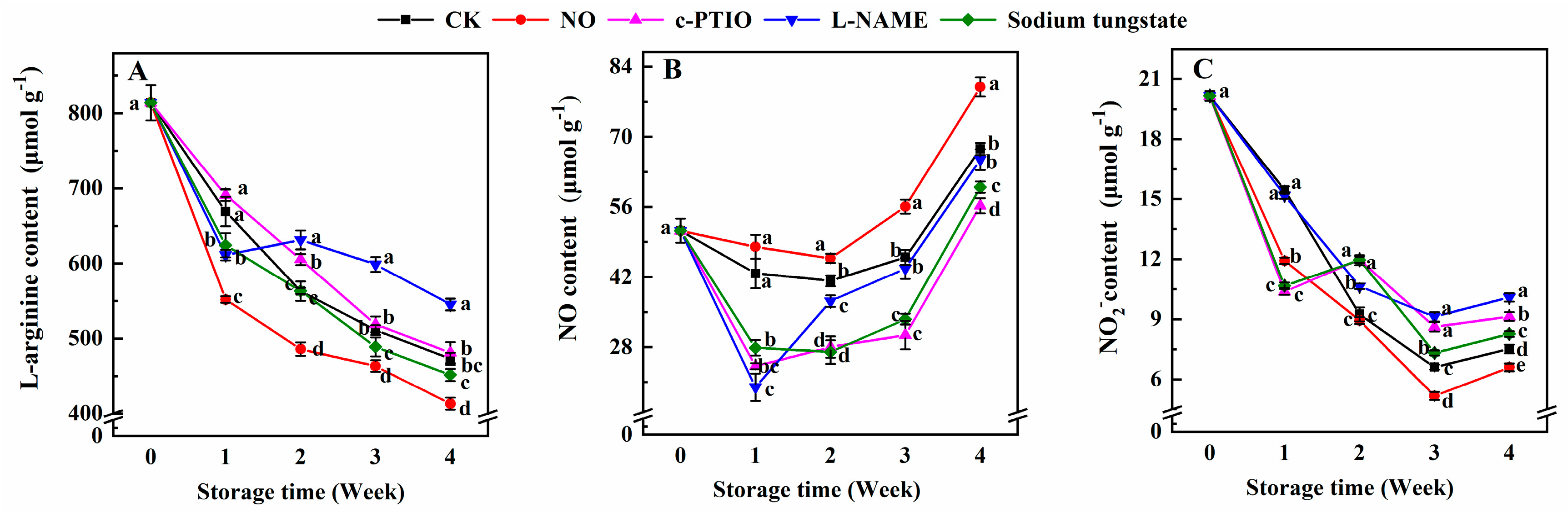
2.4. Determination of the Contents of Endogenous l-Arginine, NO, and NO2−
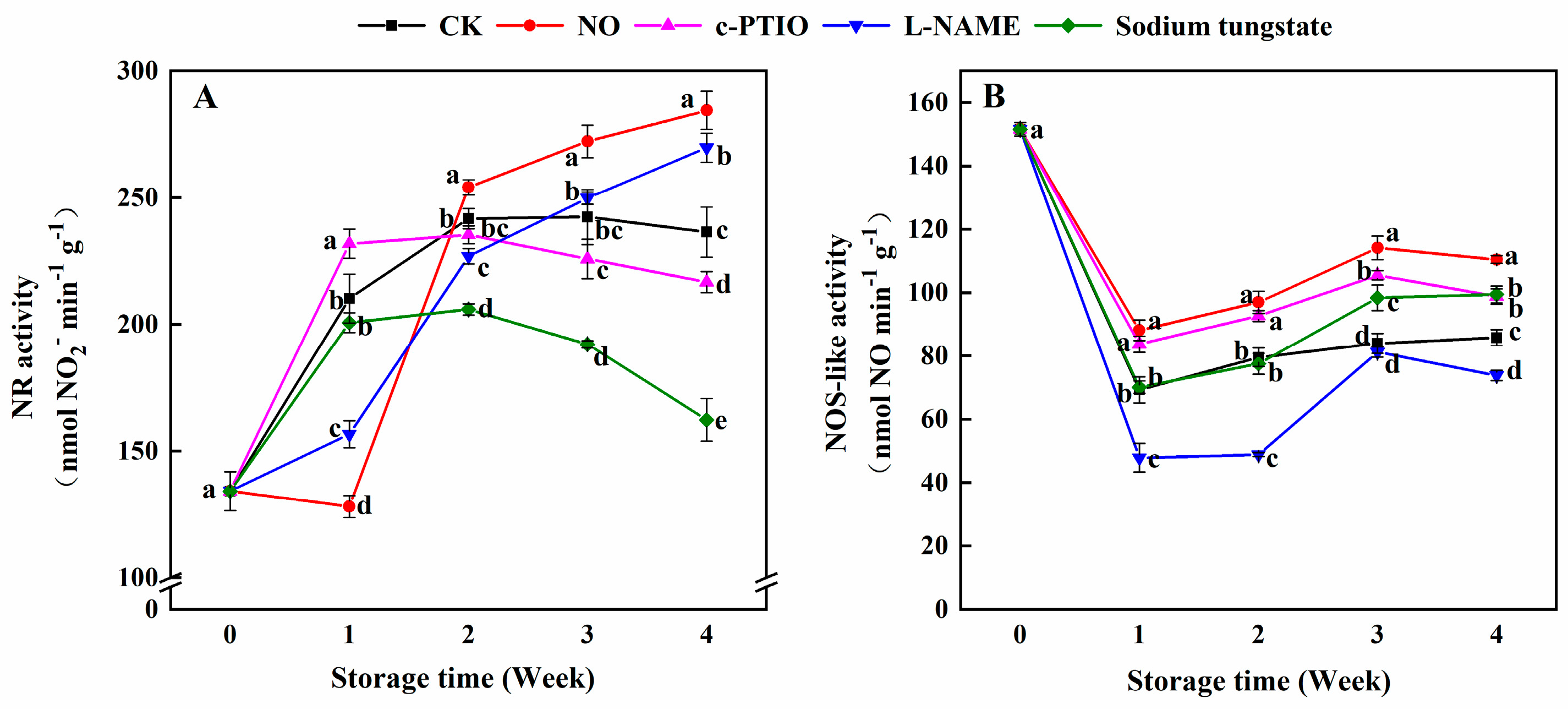
2.5. Determination of Nitrate Reductase (NR) and NOS-Like Activity
2.6. Determination of the Activities of β-Cyanoalanine Synthase (β-CAS), Sulfite Reductase (SiR), l/d-Cysteine Desulfhydrase (L/D -CDes), and O-Acetylserine(thiol)lyase (OAS-TL)
2.7. Statistical Analysis
3. Results
3.1. Changes in the Rates of Respiration, ethylene Production, Weight Loss, Soluble Sugar Content, and Browning Degree
3.2. Changes in the Contents of Endogenous H2S, Cys, Sulfite, and Total Sulfhydryl
3.3. Changes in the Contents of Endogenous l-Arginine, NO, and NO2−
3.4. Changes in the NR and NOS-Like Activities
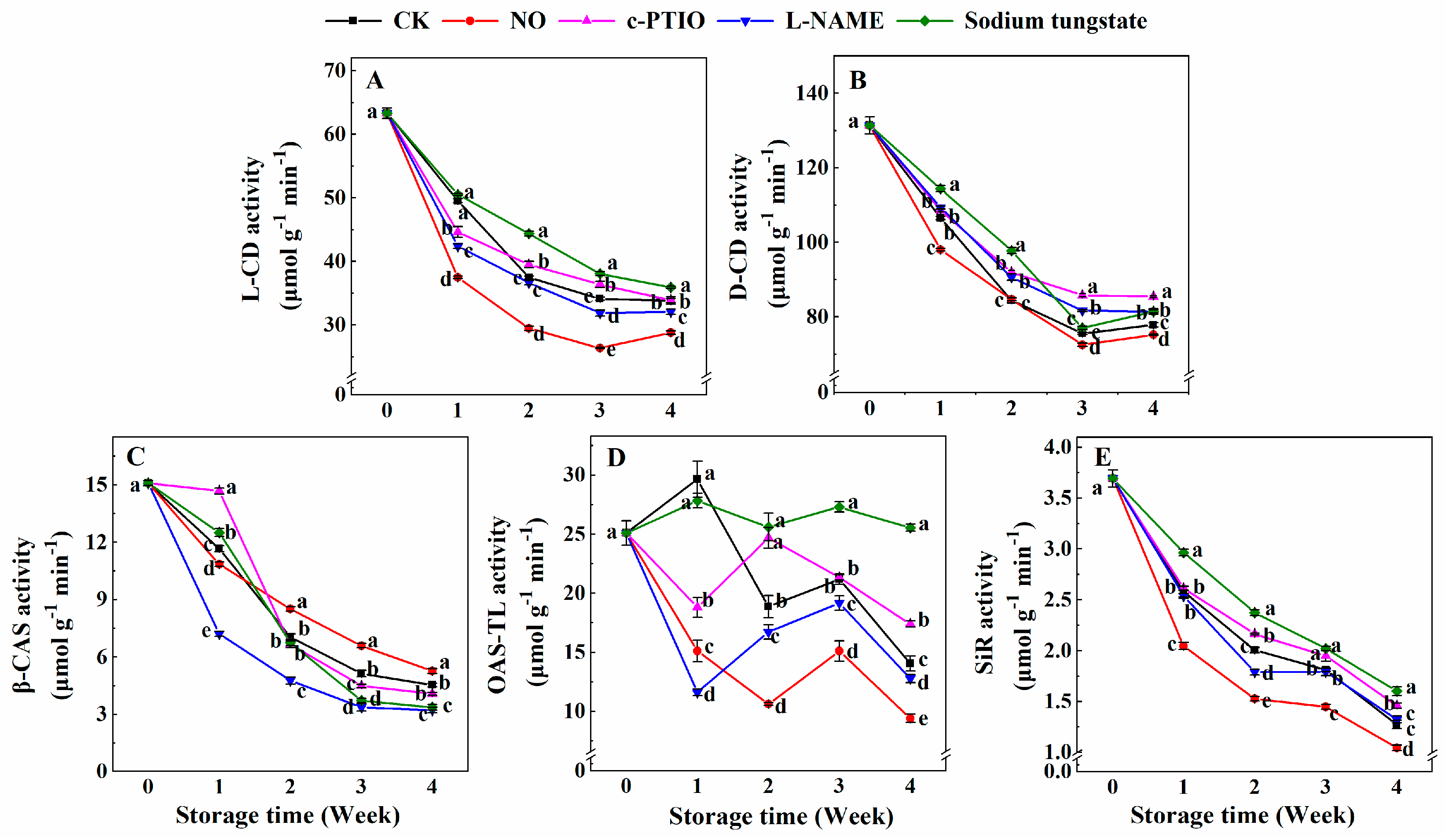
3.5. Changes in the Activities of l-CD, d-CD, β-CAS, OAS-TL, and SiR
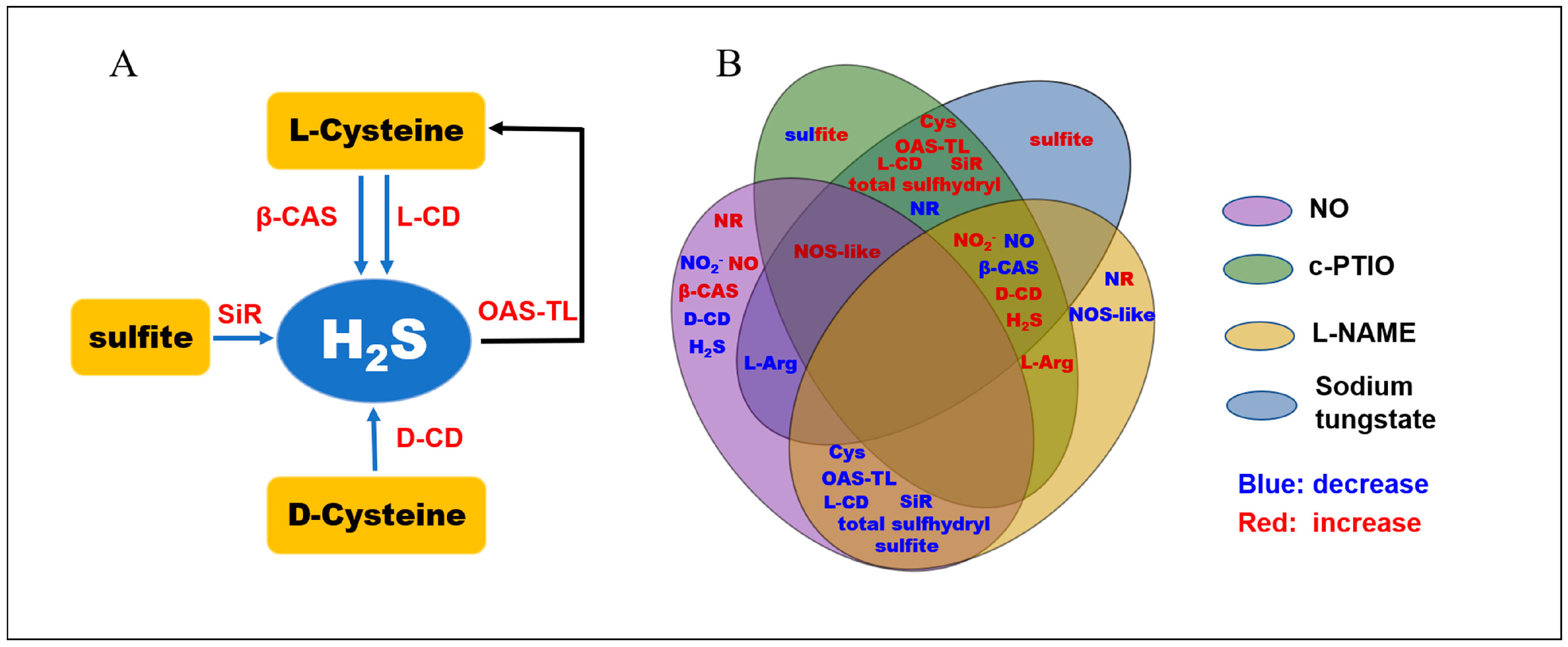
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Hancock, J.T. Hydrogen sulfide and environmental stresses. Environ. Exp. Bot. 2019, 161, 50–56. [Google Scholar] [CrossRef]
- Li, Z.-G. Analysis of some enzymes activities of hydrogen sulfide metabolism in plants. Methods Enzymol. 2015, 555, 253–269. [Google Scholar] [CrossRef] [PubMed]
- Benchoam, D.; Cuevasanta, E.; Möller, M.N.; Alvarez, B. Hydrogen sulfide and persulfides oxidation by biologically relevant oxidizing species. Antioxidants 2019, 8, 48. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.-G.; Min, X.; Zhou, Z.-H. Hydrogen sulfide: A signal molecule in plant cross-adaptation. Front. Plant Sci. 2016, 7, 1621. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.-Q.; Bao, J.; Yuan, F.; Liang, X.; Feng, Z.-T.; Wang, B.-S. Exogenous hydrogen sulfide alleviates salt stress in wheat seedlings by decreasing Na+ content. Plant Growth Regul. 2016, 79, 391–399. [Google Scholar] [CrossRef]
- Zhou, Z.-H.; Wang, Y.; Ye, X.-Y.; Li, Z.-G. Signaling molecule hydrogen sulfide improves seed germination and seedling growth of maize (Zea mays L.) under high temperature by inducing antioxidant system and osmolyte biosynthesis. Front. Plant Sci. 2018, 9, 1288. [Google Scholar] [CrossRef]
- Corpas, F.J.; Barroso, J.B.; González-Gordo, S.; Muñoz-Vargas, M.A.; Palma, J.M. Hydrogen sulfide: A novel component in Arabidopsis peroxisomes which triggers catalase inhibition. J. Integr. Plant Biol. 2019, 61, 871–883. [Google Scholar] [CrossRef]
- Jin, Z.; Wang, Z.; Yang, G.; Pei, Y. Diversity of hydrogen sulfide concentration in plant: A little spark to start a prairie fire. Sci. Bull. 2018, 63, 1314–1316. [Google Scholar] [CrossRef]
- Ziogas, V.; Molassiotis, A.; Fotopoulos, V.; Tanou, G. Hydrogen sulfide: A potent tool in postharvest fruit biology and possible mechanism of action. Front. Plant Sci. 2018, 9, 1375. [Google Scholar] [CrossRef]
- Corpas, F.J.; González-Gordo, S.; Cañas, A.; Palma, J.M. Nitric oxide and hydrogen sulfide in plants: Which comes first? J. Exp. Bot. 2019. [Google Scholar] [CrossRef]
- Hancock, J.T.; Whiteman, M. Reactive oxygen species, nitric oxide and hydrogen sulfide in plant cell regulation. Free Radic. Biol. Med. 2018, 120, S9. [Google Scholar] [CrossRef]
- Laxa, M.; Liebthal, M.; Telman, W.; Chibani, K.; Dietz, K.-J. The role of the plant antioxidant system in drought tolerance. Antioxidants 2019, 8, 94. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.N.; Mobin, M.; Abbas, Z.K.; Siddiqui, M.H. Nitric oxide-induced synthesis of hydrogen sulfide alleviates osmotic stress in wheat seedlings through sustaining antioxidant enzymes, osmolyte accumulation and cysteine homeostasis. Nitric Oxide 2017, 68, 91–102. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Wang, W.-H.; Wu, F.-H.; He, E.-M.; Liu, X.; Shangguan, Z.-P.; Zheng, H.-L. Hydrogen sulfide enhances salt tolerance through nitric oxide-mediated maintenance of ion homeostasis in barley seedling roots. Sci. Rep. 2015, 5, 12516. [Google Scholar] [CrossRef] [PubMed]
- Peng, R.; Bian, Z.; Zhou, L.; Cheng, W.; Hai, N.; Yang, C.; Yang, T.; Wang, X.; Wang, C. Hydrogen sulfide enhances nitric oxide-induced tolerance of hypoxia in maize (Zea mays L.). Plant Cell Rep. 2016, 35, 2325–2340. [Google Scholar] [CrossRef] [PubMed]
- Kaya, C.; Higgs, D.; Ashraf, M.; Alyemeni, M.N.; Ahmad, P. Integrative roles of nitric oxide and hydrogen sulfide in melatonin-induced tolerance of pepper (Capsicum annuum L.) plants to iron deficiency and salt stress alone or in combination. Physiol. Plant. 2019. [Google Scholar] [CrossRef]
- Fotopoulos, V.; Christou, A.; Antoniou, C.; Manganaris, G.A. Hydrogen sulphide: A versatile tool for the regulation of growth and defence responses in horticultural crops. J. Hortic. Sci. Biotechnol. 2015, 90, 227–234. [Google Scholar] [CrossRef]
- Huo, J.; Huang, D.; Zhang, J.; Fang, H.; Wang, B.; Wang, C.; Liao, W. Hydrogen sulfide: A gaseous molecule in postharvest freshness. Front. Plant Sci. 2018, 9, 1172. [Google Scholar] [CrossRef]
- Li, D.; Limwachiranon, J.; Li, L.; Du, R.; Luo, Z. Involvement of energy metabolism to chilling tolerance induced by hydrogen sulfide in cold-stored banana fruit. Food Chem. 2016, 208, 272–278. [Google Scholar] [CrossRef]
- Zhu, L.; Wang, W.; Shi, J.; Zhang, W.; Shen, Y.; Du, H.; Wu, S. Hydrogen sulfide extends the postharvest life and enhances antioxidant activity of kiwifruit during storage. J. Sci. Food Agric. 2014, 94, 2699–2704. [Google Scholar] [CrossRef]
- Ge, Y.; Hu, K.-D.; Wang, S.-S.; Hu, L.-Y.; Chen, X.-Y.; Li, Y.-H.; Yang, Y.; Yang, F.; Zhang, H. Hydrogen sulfide alleviates postharvest ripening and senescence of banana by antagonizing the effect of ethylene. PLoS ONE 2017, 12, e0180113. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Xu, S.; Hu, H.; Pan, J.; Li, P.; Shen, W. Endogenous hydrogen sulfide homeostasis is responsible for the alleviation of senescence of postharvest daylily flower via increasing antioxidant capacity and maintained energy status. J. Agric. Food Chem. 2017, 65, 718–726. [Google Scholar] [CrossRef] [PubMed]
- González-Gordo, S.; Bautista, R.; Claros, M.G.; Cañas, A.; Palma, J.M.; Corpas, F.J. Nitric oxide-dependent regulation of sweet pepper fruit ripening. J. Exp. Bot. 2019. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Xu, J.; Chen, Y.; Wei, J.; Wu, B. Nitric oxide treatment maintains postharvest quality of table grapes by mitigation of oxidative damage. Postharvest Biol. Technol. 2019, 152, 9–18. [Google Scholar] [CrossRef]
- Ma, Y.; Fu, L.; Hussain, Z.; Huang, D.; Zhu, S. Enhancement of storability and antioxidant systems of sweet cherry fruit by nitric oxide-releasing chitosan nanoparticles (GSNO-CS NPs). Food Chem. 2019, 285, 10–21. [Google Scholar] [CrossRef] [PubMed]
- Mukherjee, S. Recent advancements in the mechanism of nitric oxide signaling associated with hydrogen sulfide and melatonin crosstalk during ethylene-induced fruit ripening in plants. Nitric Oxide 2019, 82, 25–34. [Google Scholar] [CrossRef]
- Liu, H.; Jiang, W.; Cao, J.; Li, Y. Effect of chilling temperatures on physiological properties, phenolic metabolism and antioxidant level accompanying pulp browning of peach during cold storage. Sci. Hortic. 2019, 255, 175–182. [Google Scholar] [CrossRef]
- Koushesh Saba, M.; Moradi, S. Sodium nitroprusside (SNP) spray to maintain fruit quality and alleviate postharvest chilling injury of peach fruit. Sci. Hortic. 2017, 216, 193–199. [Google Scholar] [CrossRef]
- Zhu, L.Q.; Zhou, J.; Zhu, S.H. Effect of a combination of nitric oxide treatment and intermittent warming on prevention of chilling injury of ‘Feicheng’ peach fruit during storage. Food Chem. 2010, 121, 165–170. [Google Scholar] [CrossRef]
- Huang, D.; Hu, S.; Zhu, S.; Feng, J. Regulation by nitric oxide on mitochondrial permeability transition of peaches during storage. Plant Physiol. Biochem. 2019, 138, 17–25. [Google Scholar] [CrossRef]
- Ma, Y.; Huang, D.; Chen, C.; Zhu, S.; Gao, J. Regulation of ascorbate-glutathione cycle in peaches via nitric oxide treatment during cold storage. Sci. Hortic. 2019, 247, 400–406. [Google Scholar] [CrossRef]
- Lim, M.D.; Lorković, I.M.; Ford, P.C. The preparation of anaerobic nitric oxide solutions for the study of heme model systems in aqueous and nonaqueous media: Some consequences of NOx impurities. Methods Enzymol. 2005, 396, 3–17. [Google Scholar] [CrossRef]
- Zhang, Q.; Zhang, L.; Geng, B.; Feng, J.; Zhu, S. Interactive effects of abscisic acid and nitric oxide on chilling resistance and active oxygen metabolism in peach fruit during cold storage. J. Sci. Food Agric. 2019, 99, 3367–3380. [Google Scholar] [CrossRef]
- Jing, G.; Zhou, J.; Zhu, S. Effects of nitric oxide on mitochondrial oxidative defence in postharvest peach fruits. J. Sci. Food Agric. 2016, 96, 1997–2003. [Google Scholar] [CrossRef] [PubMed]
- Zhu, S.H.; Zhou, J. Effect of nitric oxide on ethylene production in strawberry fruit during storage. Food Chem. 2007, 100, 1517–1522. [Google Scholar] [CrossRef]
- Jermyn, M.A. Increasing the sensitivity of the anthrone method for carbohydrate. Anal. Biochem. 1975, 68, 332–335. [Google Scholar] [CrossRef]
- Lei, J.; Li, B.; Zhang, N.; Yan, R.; Guan, W.; Brennan, C.S.; Gao, H.; Peng, B. Effects of UV-C treatment on browning and the expression of polyphenol oxidase (PPO) genes in different tissues of Agaricus bisporus during cold storage. Postharvest Biol. Technol. 2018, 139, 99–105. [Google Scholar] [CrossRef]
- Sekiya, J.; Schmidt, A.; Wilson, L.G.; Filner, P. Emission of hydrogen sulfide by leaf tissue in response to L-cysteine. Plant Physiol. 1982, 70, 430–436. [Google Scholar] [CrossRef]
- Zhang, H.; Ye, Y.-K.; Wang, S.-H.; Luo, J.-P.; Tang, J.; Ma, D.-F. Hydrogen sulfide counteracts chlorophyll loss in sweetpotato seedling leaves and alleviates oxidative damage against osmotic stress. Plant Growth Regul. 2009, 58, 243–250. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Li, J.; Jia, H.; Wang, J.; Cao, Q.; Wen, Z. Hydrogen sulfide is involved in maintaining ion homeostasis via regulating plasma membrane Na+/H+ antiporter system in the hydrogen peroxide-dependent manner in salt-stress Arabidopsis thaliana root. Protoplasma 2014, 251, 899–912. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.L.; Zhu, S.H.; Chen, C.B.; Zhou, J. Metabolism of endogenous nitric oxide during growth and development of apple fruit. Sci. Hortic. 2011, 127, 500–506. [Google Scholar] [CrossRef]
- Beda, N.; Nedospasov, A. A spectrophotometric assay for nitrate in an excess of nitrite. Nitric Oxide 2005, 13, 93–97. [Google Scholar] [CrossRef] [PubMed]
- Warrilow, A.G.S.; Hawkesford, M.J. Separation, subcellular location and influence of sulphur nutrition on isoforms of cysteine synthase in spinach. J. Exp. Bot. 1998, 49, 1625–1636. [Google Scholar] [CrossRef]
- Riemenschneider, A.; Nikiforova, V.; Hoefgen, R.; De Kok, L.J.; Papenbrock, J. Impact of elevated H2S on metabolite levels, activity of enzymes and expression of genes involved in cysteine metabolism. Plant Physiol. Biochem. 2005, 43, 473–483. [Google Scholar] [CrossRef]
- Salbitani, G.; Carfagna, S. Extraction and activity of O-acetylserine(thiol)lyase (OASTL) from microalga Chlorella sorokiniana. Bio-Protocol 2017, 7, e2342. [Google Scholar] [CrossRef]
- Hartmann, T.; Mult, S.; Suter, M.; Rennenberg, H.; Herschbach, C. Leaf age-dependent differences in sulphur assimilation and allocation in poplar (Populus tremula × P. alba) leaves. J. Exp. Bot. 2000, 51, 1077–1088. [Google Scholar] [CrossRef]
- Corpas, F.J.; Palma, J.M. Nitric oxide on/off in fruit ripening. Plant Biol. 2018, 20, 805–807. [Google Scholar] [CrossRef]
- Li, Z.-G.; Yang, S.-Z.; Long, W.-B.; Yang, G.-X.; Shen, Z.-Z. Hydrogen sulphide may be a novel downstream signal molecule in nitric oxide-induced heat tolerance of maize (Zea mays L.) seedlings. Plant Cell Environ. 2013, 36, 1564–1572. [Google Scholar] [CrossRef]
- Wang, H.; Ji, F.; Zhang, Y.; Hou, J.; Liu, W.; Huang, J.; Liang, W. Interactions between hydrogen sulphide and nitric oxide regulate two soybean citrate transporters during the alleviation of aluminium toxicity. Plant Cell Environ. 2019, 42, 2340–2356. [Google Scholar] [CrossRef]
- Liang, Y.; Zheng, P.; Li, S.; Li, K.; Xu, H. Nitrate reductase-dependent NO production is involved in H2S-induced nitrate stress tolerance in tomato via activation of antioxidant enzymes. Sci. Hortic. 2018, 229, 207–214. [Google Scholar] [CrossRef]
- Munoz-Vargas, M.A.; Gonzalez-Gordo, S.; Canas, A.; Lopez-Jaramillo, J.; Palma, J.M.; Corpas, F.J. Endogenous hydrogen sulfide (H2S) is up-regulated during sweet pepper (Capsicum annuum L.) fruit ripening. In vitro analysis shows that NADP-dependent isocitrate dehydrogenase (ICDH) activity is inhibited by H2S and NO. Nitric Oxide 2018, 81, 36–45. [Google Scholar] [CrossRef] [PubMed]
- Chang, Z.; Jingying, S.; Liqin, Z.; Changle, L.; Qingguo, W. Cooperative effects of hydrogen sulfide and nitric oxide on delaying softening and decay of strawberry. Int. J. Agric. Biol. Eng. 2014, 7, 114–122. [Google Scholar]
- Xia, Z.; Wang, M.; Xu, Z. The maize sulfite reductase is involved in cold and oxidative stress responses. Front. Plant Sci. 2018, 9, 1680. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Tang, J.; Yao, G.-F.; Huang, Z.-Q.; Li, Y.-H.; Han, Z.; Chen, X.-Y.; Hu, L.-Y.; Hu, K.-D.; Zhang, H. Central role of adenosine 5′-phosphosulfate reductase in the control of plant hydrogen sulfide metabolism. Front. Plant Sci. 2018, 9, 1404. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Hou, L.; Liu, G.; Liu, X.; Wang, X. Hydrogen sulfide induced by nitric oxide mediates ethylene-induced stomatal closure of Arabidopsis thaliana. Chin. Sci. Bull. 2011, 56, 3547–3553. [Google Scholar] [CrossRef]
- Zhang, P.; Luo, Q.; Wang, R.; Xu, J. Hydrogen sulfide toxicity inhibits primary root growth through the ROS-NO pathway. Sci. Rep. 2017, 7, 868. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Hou, Z.-H.; Liu, G.-H.; Hou, L.-X.; Liu, X. Hydrogen sulfide may function downstream of nitric oxide in ethylene-induced stomatal closure in Vicia faba L. J. Integr. Agric. 2012, 11, 1644–1653. [Google Scholar] [CrossRef]
- Scuffi, D.; Álvarez, C.; Laspina, N.; Gotor, C.; Lamattina, L.; García-Mata, C. Hydrogen sulfide generated by L-cysteine desulfhydrase acts upstream of nitric oxide to modulate abscisic acid-dependent stomatal closure. Plant Physiol. 2014, 166, 2065–2076. [Google Scholar] [CrossRef] [PubMed]






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Geng, B.; Huang, D.; Zhu, S. Regulation of Hydrogen Sulfide Metabolism by Nitric Oxide Inhibitors and the Quality of Peaches during Cold Storage. Antioxidants 2019, 8, 401. https://doi.org/10.3390/antiox8090401
Geng B, Huang D, Zhu S. Regulation of Hydrogen Sulfide Metabolism by Nitric Oxide Inhibitors and the Quality of Peaches during Cold Storage. Antioxidants. 2019; 8(9):401. https://doi.org/10.3390/antiox8090401
Chicago/Turabian StyleGeng, Biao, Dandan Huang, and Shuhua Zhu. 2019. "Regulation of Hydrogen Sulfide Metabolism by Nitric Oxide Inhibitors and the Quality of Peaches during Cold Storage" Antioxidants 8, no. 9: 401. https://doi.org/10.3390/antiox8090401