

Identification of Oxidative-Stress-Reducing Plant Extracts from a Novel Extract Library—Comparative Analysis of Cell-Free and Cell-Based In Vitro Assays to Quantitate Antioxidant Activity
 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Sample Collection, Categorization, and Pretreatment
2.2. Plant Extract Preparation
2.3. Determination of Total Phenolic Content
2.4. Determination of Ferric Reducing Antioxidant Power
2.5. Cell Culture
2.6. Detection of ROS Production in Caco-2 Cells
2.7. Detection of NO Production in RAW264.7 Cells
2.8. Determination of Cell Viability
2.9. Statistical Analysis
3. Results
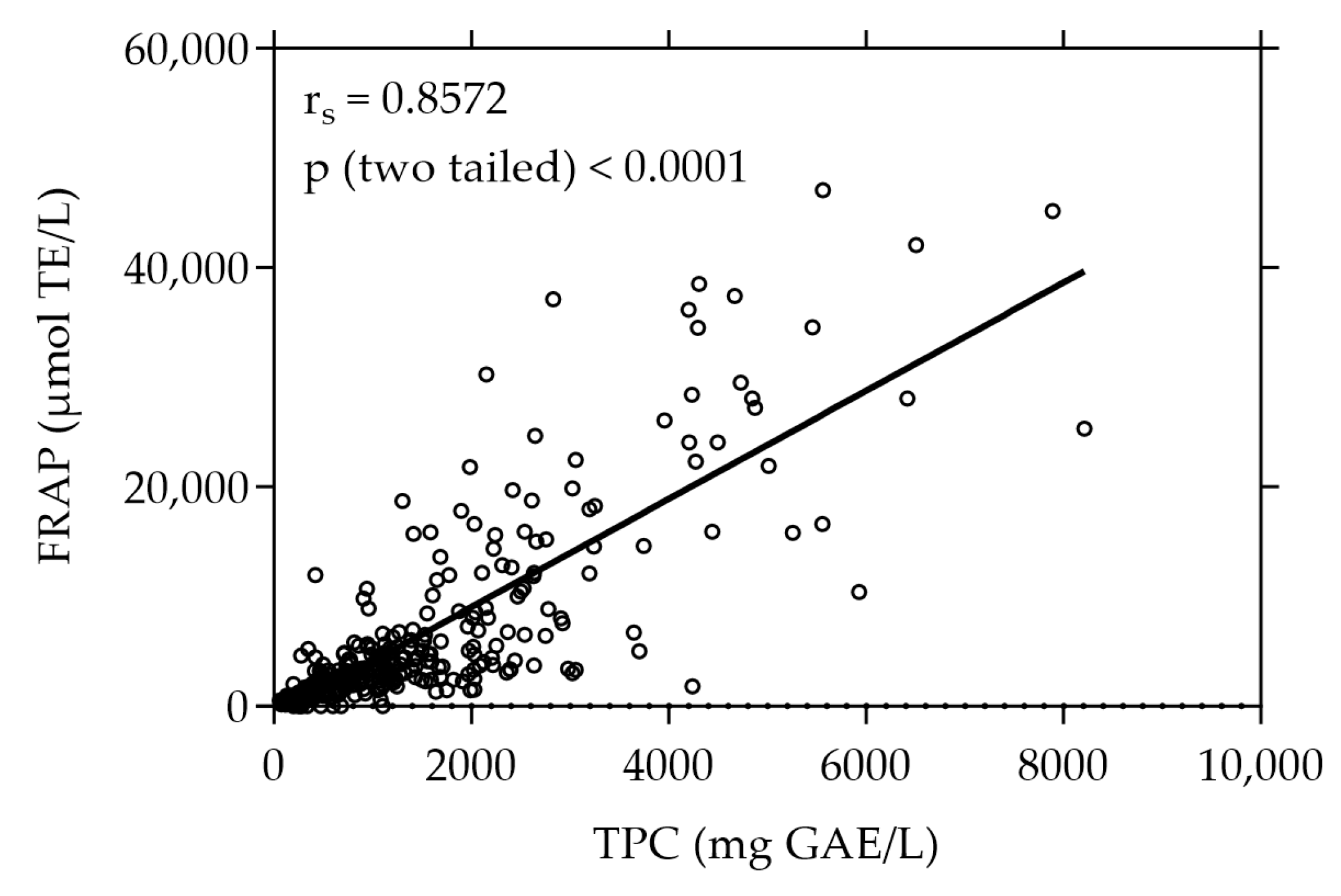
3.1. TPC and FRAP Values of Plant Extracts under Study
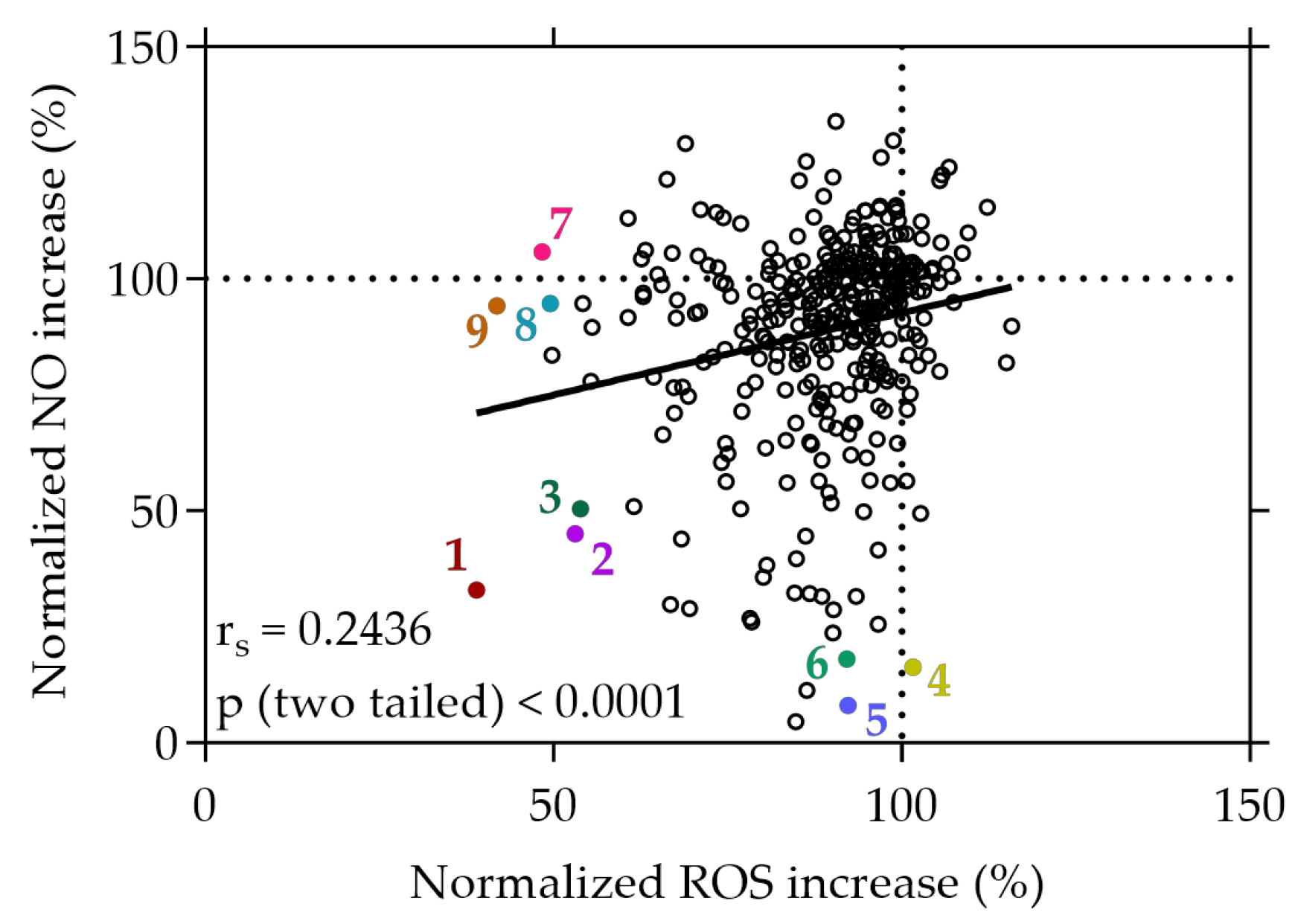
3.2. Inhibition of Cellular ROS and NO by the Plant Extracts
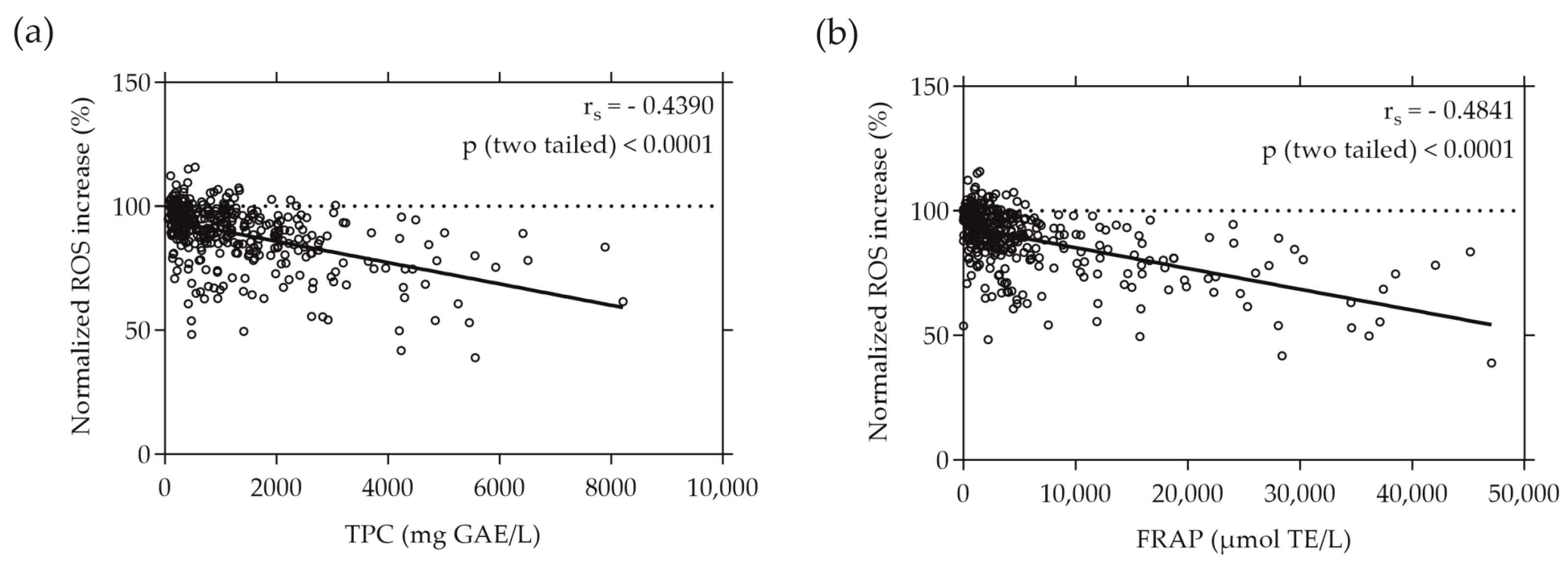
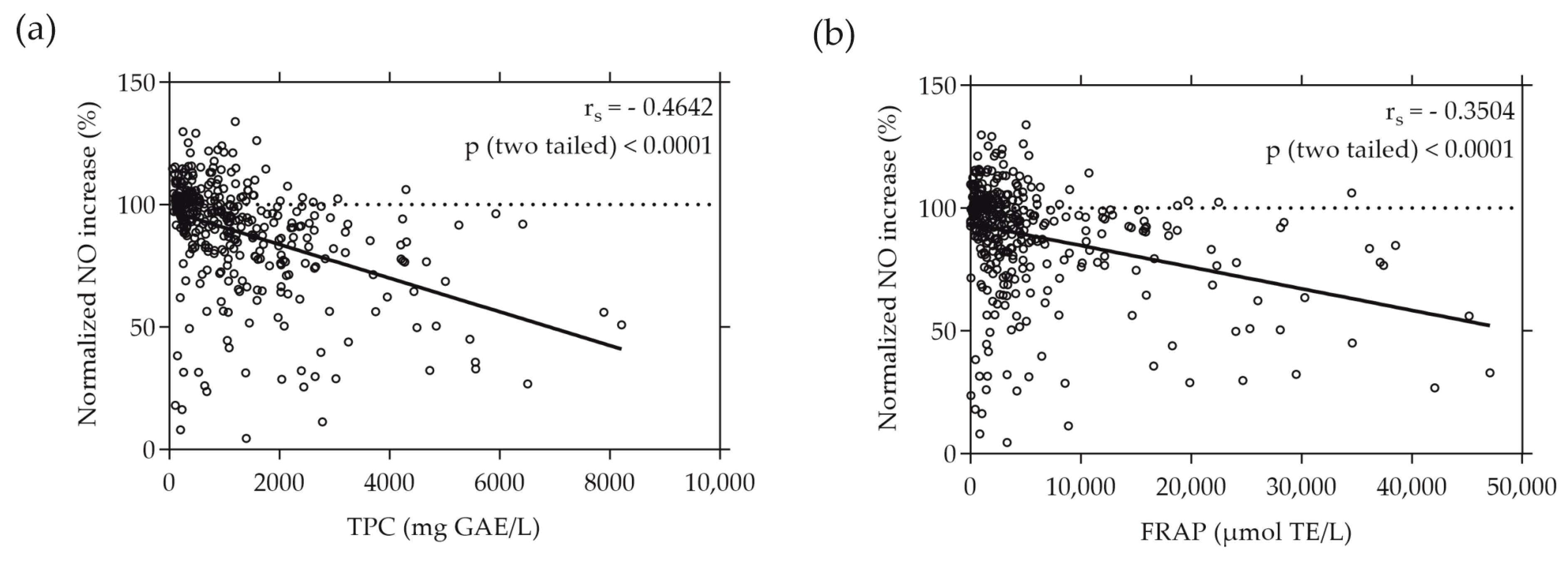
3.3. Moderate Correlations Found between Cell-Free and Cell-Based Antioxidant Assays
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Atanasov, A.G.; Waltenberger, B.; Pferschy-Wenzig, E.-M.; Linder, T.; Wawrosch, C.; Uhrin, P.; Temml, V.; Wang, L.; Schwaiger, S.; Heiss, E.H.; et al. Discovery and Resupply of Pharmacologically Active Plant-Derived Natural Products: A Review. Biotechnol. Adv. 2015, 33, 1582–1614. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Torres, I.; Castrejón-Téllez, V.; Soto, M.E.; Rubio-Ruiz, M.E.; Manzano-Pech, L.; Guarner-Lans, V. Oxidative Stress, Plant Natural Antioxidants, and Obesity. Int. J. Mol. Sci. 2021, 22, 1786. [Google Scholar] [CrossRef] [PubMed]
- Mittal, M.; Siddiqui, M.R.; Tran, K.; Reddy, S.P.; Malik, A.B. Reactive Oxygen Species in Inflammation and Tissue Injury. Antioxid. Redox Signal 2014, 20, 1126–1167. [Google Scholar] [CrossRef] [PubMed]
- Modun, D.; Giustarini, D.; Tsikas, D. Nitric Oxide-Related Oxidative Stress and Redox Status in Health and Disease. Oxid. Med. Cell Longev. 2014, 2014, 129651. [Google Scholar] [CrossRef] [PubMed]
- Förstermann, U.; Xia, N.; Li, H. Roles of Vascular Oxidative Stress and Nitric Oxide in the Pathogenesis of Atherosclerosis. Circ. Res. 2017, 120, 713–735. [Google Scholar] [CrossRef] [PubMed]
- Hedayati, N.; Naeini, M.B.; Nezami, A.; Hosseinzadeh, H.; Wallace Hayes, A.; Hosseini, S.; Imenshahidi, M.; Karimi, G. Protective Effect of Lycopene against Chemical and Natural Toxins: A Review. BioFactors 2019, 45, 5–23. [Google Scholar] [CrossRef] [PubMed]
- Nishino, A.; Yasui, H.; Maoka, T. Reaction and Scavenging Mechanism of β-Carotene and Zeaxanthin with Reactive Oxygen Species. J. Oleo Sci. 2017, 66, 77–84. [Google Scholar] [CrossRef]
- Musial, C.; Kuban-Jankowska, A.; Gorska-Ponikowska, M. Beneficial Properties of Green Tea Catechins. Int. J. Mol. Sci. 2020, 21, 1744. [Google Scholar] [CrossRef]
- Ma, Z.; Zhang, H. Phytochemical Constituents, Health Benefits, and Industrial Applications of Grape Seeds: A Mini-Review. Antioxidants 2017, 6, 71. [Google Scholar] [CrossRef]
- Goswami, M.J.; Dutta, U.; Seema, T.; Bharali, S.J.; Yanka, H.; Tag, H.; Bharali, P.; Kakati, D. Antioxidant and Antidiabetic Properties of Extracts from Three Underutilized Food Plants of North East India. Chem. Biodivers. 2023, 20, e2022007. [Google Scholar] [CrossRef]
- Junsathian, P.; Nakamura, S.; Katayama, S.; Rawdkuen, S. Antioxidant and Antimicrobial Activities of Thai Edible Plant Extracts Prepared Using Different Extraction Techniques. Molecules 2022, 27, 6489. [Google Scholar] [CrossRef] [PubMed]
- Sandner, G.; Stadlbauer, V.; Sadova, N.; Neuhauser, C.; Schwarzinger, B.; Karlsberger, L.; Hangweirer, K.; Antensteiner, K.; Stallinger, A.; Aumiller, T.; et al. Grape Seed Extract Improves Intestinal Barrier Integrity and Performance: Evidence from in Vitro, Caenorhabditis Elegans and Drosophila Melanogaster Experiments and a Study with Growing Broilers. Food Biosci. 2023, 52, 102483. [Google Scholar] [CrossRef]
- Aqil, F.; Ahmad, I.; Mehmood, Z. Antioxidant and Free Radical Scavenging Properties of Twelve Traditionally Used Indian Medicinal Plants. Turk. J. Biol. 2006, 30, 177–183. [Google Scholar]
- Liu, X.; Zhao, M.; Wang, J.; Yang, B.; Jiang, Y. Antioxidant Activity of Methanolic Extract of Emblica Fruit (Phyllanthus Emblica L.) from Six Regions in China. J. Food Compos. Anal. 2008, 21, 219–228. [Google Scholar] [CrossRef]
- Muflihah, Y.M.; Gollavelli, G.; Ling, Y.-C. Correlation Study of Antioxidant Activity with Phenolic and Flavonoid Compounds in 12 Indonesian Indigenous Herbs. Antioxidants 2021, 10, 1530. [Google Scholar] [CrossRef] [PubMed]
- Dobrinas, S.; Soceanu, A.; Popescu, V.; Carazeanu Popovici, I.; Jitariu, D. Relationship between Total Phenolic Content, Antioxidant Capacity, Fe and Cu Content from Tea Plant Samples at Different Brewing Times. Processes 2021, 9, 1311. [Google Scholar] [CrossRef]
- Dasgupta, N.; De, B. Antioxidant Activity of Some Leafy Vegetables of India: A Comparative Study. Food Chem. 2007, 101, 471–474. [Google Scholar] [CrossRef]
- Gulcin, İ. Antioxidants and Antioxidant Methods: An Updated Overview. Arch. Toxicol. 2020, 94, 651–715. [Google Scholar] [CrossRef]
- Munteanu, I.G.; Apetrei, C. Analytical Methods Used in Determining Antioxidant Activity: A Review. Int. J. Mol. Sci. 2021, 22, 3380. [Google Scholar] [CrossRef]
- Wolfe, K.L.; Liu, R.H. Cellular Antioxidant Activity (CAA) Assay for Assessing Antioxidants, Foods, and Dietary Supplements. J. Agric. Food Chem. 2007, 55, 8896–8907. [Google Scholar] [CrossRef]
- López-Alarcón, C.; Denicola, A. Evaluating the Antioxidant Capacity of Natural Products: A Review on Chemical and Cellular-Based Assays. Anal. Chim. Acta 2013, 763, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Heckmann, M.; Sadova, N.; Drotarova, I.; Atzmüller, S.; Schwarzinger, B.; Guedes, R.M.C.; Correia, P.A.; Hirtenlehner, S.; Potthast, C.; Klanert, G.; et al. Extracts Prepared from Feed Supplements Containing Wood Lignans Improve Intestinal Health by Strengthening Barrier Integrity and Reducing Inflammation. Molecules 2022, 27, 6327. [Google Scholar] [CrossRef] [PubMed]
- König, A.; Sadova, N.; Dornmayr, M.; Schwarzinger, B.; Neuhauser, C.; Stadlbauer, V.; Wallner, M.; Woischitzschläger, J.; Müller, A.; Tona, R.; et al. Combined Acid Hydrolysis and Fermentation Improves Bioactivity of Citrus Flavonoids in Vitro and in Vivo. Commun. Biol. 2023, 6, 1083. [Google Scholar] [CrossRef]
- Mahdi-Pour, B.; Jothy, S.L.; Latha, L.Y.; Chen, Y.; Sasidharan, S. Antioxidant Activity of Methanol Extracts of Different Parts of Lantana Camara. Asian Pac. J. Trop. Biomed. 2012, 2, 960–965. [Google Scholar] [CrossRef]
- Furger, C. Live Cell Assays for the Assessment of Antioxidant Activities of Plant Extracts. Antioxidants 2021, 10, 944. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Zhang, X.; Broderick, M.; Fein, H. Measurement of Nitric Oxide Production in Biological Systems by Using Griess Reaction Assay. Sensors 2003, 3, 276–284. [Google Scholar] [CrossRef]
- Nakagawa, T.; Yokozawa, T. Direct Scavenging of Nitric Oxide and Superoxide by Green Tea. Food Chem. Toxicol. 2002, 40, 1745–1750. [Google Scholar] [CrossRef]
- Sreejayan, X.X.; Rao, M.N.A. Nitric Oxide Scavenging by Curcuminoids. J. Pharm. Pharmacol. 2011, 49, 105–107. [Google Scholar] [CrossRef]
- Hussain, T.; Tan, B.; Yin, Y.; Blachier, F.; Tossou, M.C.B.; Rahu, N. Oxidative Stress and Inflammation: What Polyphenols Can Do for Us? Oxid. Med. Cell Longev. 2016, 2016, 7432797. [Google Scholar] [CrossRef]
- Sultanbawa, Y.; Sivakumar, D. Enhanced Nutritional and Phytochemical Profiles of Selected Underutilized Fruits, Vegetables, and Legumes. Curr. Opin. Food Sci. 2022, 46, 100853. [Google Scholar] [CrossRef]
- Cragg, G.M.; Newman, D.J. Biodiversity: A Continuing Source of Novel Drug Leads. Pure Appl. Chem. 2005, 77, 7–24. [Google Scholar] [CrossRef]
- Arenas-Jal, M.; Suñé-Negre, J.M.; Pérez-Lozano, P.; García-Montoya, E. Trends in the Food and Sports Nutrition Industry: A Review. Crit. Rev. Food Sci. Nutr. 2020, 60, 2405–2421. [Google Scholar] [CrossRef]
- Secretariat of the Convention on Biological Diversity. Nagoya Protocol on Access to Genetic Resources and the Fair and Equitable Sharing of Benefits Arising from Their Utilization to the Convention on Biological Diversity. Available online: https://www.cbd.int/abs/doc/protocol/nagoya-protocol-en.pdf (accessed on 28 November 2023).
- Onur, S.O.; Stöckmann, H.; Zenthoefer, M.; Piker, L.; Döring, F. The Plant Extract Collection Kiel in Schleswig-Holstein (PECKISH) Is an Open Access Screening Library. J. Food Res. 2013, 2, 101. [Google Scholar] [CrossRef]
- Chhabra, S.C.; Uiso, F.C.; Mshiu, E.N. Phytochemical Screening of Tanzanian Medicinal Plants. I. J. Ethnopharmacol. 1984, 11, 157–179. [Google Scholar] [CrossRef] [PubMed]
- He, M.; Grkovic, T.; Evans, J.R.; Thornburg, C.C.; Akee, R.K.; Thompson, J.R.; Whitt, J.A.; Harris, M.J.; Loyal, J.A.; Britt, J.R.; et al. The NCI Library of Traditional Chinese Medicinal Plant Extracts—Preliminary Assessment of the NCI-60 Activity and Chemical Profiling of Selected Species. Fitoterapia 2019, 137, 104285. [Google Scholar] [CrossRef] [PubMed]
- Yen, C.-H.; Chang, H.-S.; Yang, T.-H.; Wang, S.-F.; Wu, H.-C.; Chen, Y.-C.; Lin, K.-J.; Wang, S. High-Content Screening of a Taiwanese Indigenous Plant Extract Library Identifies Syzygium Simile Leaf Extract as an Inhibitor of Fatty Acid Uptake. Int. J. Mol. Sci. 2018, 19, 2130. [Google Scholar] [CrossRef] [PubMed]
- Nesterov, A.; Hong, M.; Hertel, C.; Jiao, P.; Brownell, L.; Cannon, E. Screening a Plant Extract Library for Inhibitors of Cholecystokinin Receptor CCK1 Pathways. SLAS Discov. 2010, 15, 518–527. [Google Scholar] [CrossRef]
- McCloud, T.G. High Throughput Extraction of Plant, Marine and Fungal Specimens for Preservation of Biologically Active Molecules. Molecules 2010, 15, 4526–4563. [Google Scholar] [CrossRef]
- Mäder, P.; Boho, D.; Rzanny, M.; Seeland, M.; Wittich, H.C.; Deggelmann, A.; Wäldchen, J. The Flora Incognita App—Interactive Plant Species Identification. Methods Ecol. Evol. 2021, 12, 1335–1342. [Google Scholar] [CrossRef]
- Hohla, M.; Stöhr, O.; Brandstätter, G.; Danner, J.; Diewald, W.; Essl, F.; Fiereder, H.; Grims, F.; Höglinger, F.; Kleesadl, G.; et al. Katalog und Rote Liste der Gefäßpflanzen Oberösterreichs. Stapfia Land. Oberösterreich Linz. 2009, 91, 1–324. [Google Scholar]
- Singleton, V.L.; Rossi, J.A. Colorimetry of Total Phenolics with Phosphomolybdic-Phosphotungstic Acid Reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar] [CrossRef]
- Benzie, I.F.F.; Strain, J.J. Ferric Reducing/Antioxidant Power Assay: Direct Measure of Total Antioxidant Activity of Biological Fluids and Modified Version for Simultaneous Measurement of Total Antioxidant Power and Ascorbic Acid Concentration. Methods Enzym. Enzymol. 1999, 299, 15–27. [Google Scholar] [CrossRef]
- Wan, H.; Liu, D.; Yu, X.; Sun, H.; Li, Y. A Caco-2 Cell-Based Quantitative Antioxidant Activity Assay for Antioxidants. Food Chem. 2015, 175, 601–608. [Google Scholar] [CrossRef] [PubMed]
- Zeb, A. Concept, Mechanism, and Applications of Phenolic Antioxidants in Foods. J. Food Biochem. 2020, 44, e13394. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, B.; Marques, A.; Ramos, C.; Serrano, C.; Matos, O.; Neng, N.R.; Nogueira, J.M.F.; Saraiva, J.A.; Nunes, M.L. Chemical Composition and Bioactivity of Different Oregano (Origanum vulgare) Extracts and Essential Oil. J. Sci. Food Agric. 2013, 93, 2707–2714. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, M.R.A.; Krause, L.C.; Caramão, E.B.; dos Santos, J.G.; Dariva, C.; Vladimir de Oliveira, J. Chemical Composition and Extraction Yield of the Extract of Origanum vulgare Obtained from Sub- and Supercritical CO 2. J. Agric. Food Chem. 2004, 52, 3042–3047. [Google Scholar] [CrossRef]
- Wang, S.Y.; Lin, H.-S. Antioxidant Activity in Fruits and Leaves of Blackberry, Raspberry, and Strawberry Varies with Cultivar and Developmental Stage. J. Agric. Food Chem. 2000, 48, 140–146. [Google Scholar] [CrossRef]
- Li, H.; Tsao, R.; Deng, Z. Factors Affecting the Antioxidant Potential and Health Benefits of Plant Foods. Can. J. Plant Sci. 2012, 92, 1101–1111. [Google Scholar] [CrossRef]
- Huang, D.; Ou, B.; Prior, R.L. The Chemistry behind Antioxidant Capacity Assays. J. Agric. Food Chem. 2005, 53, 1841–1856. [Google Scholar] [CrossRef]
- Priyanthi, C.; Sivakanesan, R. The Total Antioxidant Capacity and the Total Phenolic Content of Rice Using Water as a Solvent. Int. J. Food Sci. 2021, 2021, 5268584. [Google Scholar] [CrossRef]
- Diep, T.; Pook, C.; Yoo, M. Phenolic and Anthocyanin Compounds and Antioxidant Activity of Tamarillo (Solanum betaceum Cav.). Antioxidants 2020, 9, 169. [Google Scholar] [CrossRef] [PubMed]
- Wong, S.; Leong, L.; William Koh, J. Antioxidant Activities of Aqueous Extracts of Selected Plants. Food Chem. 2006, 99, 775–783. [Google Scholar] [CrossRef]
- Tymczewska, A.; Klebba, J.; Szydłowska-Czerniak, A. Antioxidant Capacity and Total Phenolic Content of Spice Extracts Obtained by Ultrasound-Assisted Extraction Using Deep Eutectic and Conventional Solvents. Appl. Sci. 2023, 13, 6987. [Google Scholar] [CrossRef]
- Staszowska-Karkut, M.; Materska, M. Phenolic Composition, Mineral Content, and Beneficial Bioactivities of Leaf Extracts from Black Currant (Ribes nigrum L.), Raspberry (Rubus idaeus), and Aronia (Aronia melanocarpa). Nutrients 2020, 12, 463. [Google Scholar] [CrossRef] [PubMed]
- Liberal, J.; Francisco, V.; Costa, G.; Figueirinha, A.; Amaral, M.T.; Marques, C.; Girão, H.; Lopes, M.C.; Cruz, M.T.; Batista, M.T. Bioactivity of Fragaria Vesca Leaves through Inflammation, Proteasome and Autophagy Modulation. J. Ethnopharmacol. 2014, 158, 113–122. [Google Scholar] [CrossRef]
- Şöhretoğlu, D.; Genç, Y.; Harput, Ş. Comparative Evaluation of Phenolic Profile, Antioxidative and Cytotoxic Activities of Different Geranium Species. Iran. J. Pharm. Res. 2017, 16, 178–187. [Google Scholar] [PubMed]
- Koss-Mikołajczyk, I.; Kusznierewicz, B.; Wiczkowski, W.; Płatosz, N.; Bartoszek, A. Phytochemical Composition and Biological Activities of Differently Pigmented Cabbage (Brassica oleracea Var. Capitata) and Cauliflower (Brassica oleracea Var. Botrytis) Varieties. J. Sci. Food Agric. 2019, 99, 5499–5507. [Google Scholar] [CrossRef] [PubMed]
- Ray, L.R.; Alam, M.S.; Junaid, M.; Ferdousy, S.; Akter, R.; Hosen, S.M.Z.; Mouri, N.J. Brassica oleracea Var. Capitata f. Alba: A Review on Its Botany, Traditional Uses, Phytochemistry and Pharmacological Activities. Mini-Rev. Med. Chem. 2021, 21, 2399–2417. [Google Scholar] [CrossRef]
- Szewczyk, A.; Zagaja, M.; Bryda, J.; Kosikowska, U.; Stępień-Pyśniak, D.; Winiarczyk, S.; Andres-Mach, M. Topinambur—New Possibilities for Use in a Supplementation Diet. Ann. Agric. Environ. Med. 2019, 26, 24–28. [Google Scholar] [CrossRef]
- Sittisart, P.; Chitsomboon, B. Intracellular ROS Scavenging Activity and Downregulation of Inflammatory Mediators in RAW264.7 Macrophage by Fresh Leaf Extracts of Pseuderanthemum palatiferum. Evid. Based Complement. Altern. Med. 2014, 2014, 309095. [Google Scholar] [CrossRef]
- Guo, G.-X.; Wu, K.-Y.; Zhang, X.-Y.; Lai, F.-X.; Tsim, K.W.-K.; Qin, Q.-W.; Hu, W.-H. The Extract of Curcumae Longae Rhizoma Suppresses Angiogenesis via VEGF-Induced PI3K/Akt-ENOS-NO Pathway. J. Ethnopharmacol. 2023, 308, 116299. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Mu, B.; Song, Z.; Ma, Z.; Mu, T. The In Vitro Antioxidant Activity and Inhibition of Intracellular Reactive Oxygen Species of Sweet Potato Leaf Polyphenols. Oxid. Med. Cell Longev. 2018, 2018, 9017828. [Google Scholar] [CrossRef] [PubMed]
- Xiong, L.; Yang, J.; Jiang, Y.; Lu, B.; Hu, Y.; Zhou, F.; Mao, S.; Shen, C. Phenolic Compounds and Antioxidant Capacities of 10 Common Edible Flowers from China. J. Food Sci. 2014, 79, C517–C525. [Google Scholar] [CrossRef] [PubMed]
- Chung, Y.; Park, J.-Y.; Lee, J.-E.; Kim, K.-T.; Paik, H.-D. Antioxidant Activity and Inhibitory Effect on Nitric Oxide Production of Hydroponic Ginseng Fermented with Lactococcus Lactis KC24. Antioxidants 2021, 10, 1614. [Google Scholar] [CrossRef] [PubMed]
- Xue, T.-T.; Yang, Y.-G.; Tang, Z.-S.; Duan, J.-A.; Song, Z.-X.; Hu, X.-H.; Yang, H.-D.; Xu, H.-B. Evaluation of Antioxidant, Enzyme Inhibition, Nitric Oxide Production Inhibitory Activities and Chemical Profiles of the Active Extracts from the Medicinal and Edible Plant: Althaea Officinalis. Food Res. Int. 2022, 156, 111166. [Google Scholar] [CrossRef] [PubMed]
- Leong, S.W.; Mohd Faudzi, S.M.; Abas, F.; Mohd Aluwi, M.F.F.; Rullah, K.; Lam, K.W.; Abdul Bahari, M.N.; Ahmad, S.; Tham, C.L.; Shaari, K.; et al. Nitric Oxide Inhibitory Activity and Antioxidant Evaluations of 2-Benzoyl-6-Benzylidenecyclohexanone Analogs, a Novel Series of Curcuminoid and Diarylpentanoid Derivatives. Bioorg Med. Chem. Lett. 2015, 25, 3330–3337. [Google Scholar] [CrossRef] [PubMed]
- Habashy, N.H.; Abu Serie, M.M.; Attia, W.E.; Abdelgaleil, S.A.M. Chemical Characterization, Antioxidant and Anti-Inflammatory Properties of Greek Thymus Vulgaris Extracts and Their Possible Synergism with Egyptian Chlorella Vulgaris. J. Funct. Foods 2018, 40, 317–328. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Heckmann, M.; Stadlbauer, V.; Drotarova, I.; Gramatte, T.; Feichtinger, M.; Arnaut, V.; Atzmüller, S.; Schwarzinger, B.; Röhrl, C.; Blank-Landeshammer, B.; et al. Identification of Oxidative-Stress-Reducing Plant Extracts from a Novel Extract Library—Comparative Analysis of Cell-Free and Cell-Based In Vitro Assays to Quantitate Antioxidant Activity. Antioxidants 2024, 13, 297. https://doi.org/10.3390/antiox13030297
Heckmann M, Stadlbauer V, Drotarova I, Gramatte T, Feichtinger M, Arnaut V, Atzmüller S, Schwarzinger B, Röhrl C, Blank-Landeshammer B, et al. Identification of Oxidative-Stress-Reducing Plant Extracts from a Novel Extract Library—Comparative Analysis of Cell-Free and Cell-Based In Vitro Assays to Quantitate Antioxidant Activity. Antioxidants. 2024; 13(3):297. https://doi.org/10.3390/antiox13030297
Chicago/Turabian StyleHeckmann, Mara, Verena Stadlbauer, Ivana Drotarova, Theresa Gramatte, Michaela Feichtinger, Verena Arnaut, Stefanie Atzmüller, Bettina Schwarzinger, Clemens Röhrl, Bernhard Blank-Landeshammer, and et al. 2024. "Identification of Oxidative-Stress-Reducing Plant Extracts from a Novel Extract Library—Comparative Analysis of Cell-Free and Cell-Based In Vitro Assays to Quantitate Antioxidant Activity" Antioxidants 13, no. 3: 297. https://doi.org/10.3390/antiox13030297