
A Metabolomic Signature of Ischemic Stroke Showing Acute Oxidative and Energetic Stress
 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethic Statements
2.2. Study Participants
2.3. Studied Parameters
2.4. Blood Sample Collection
2.5. Metabolomic Analysis
2.6. Data Analyses
3. Results
3.1. Clinical Features
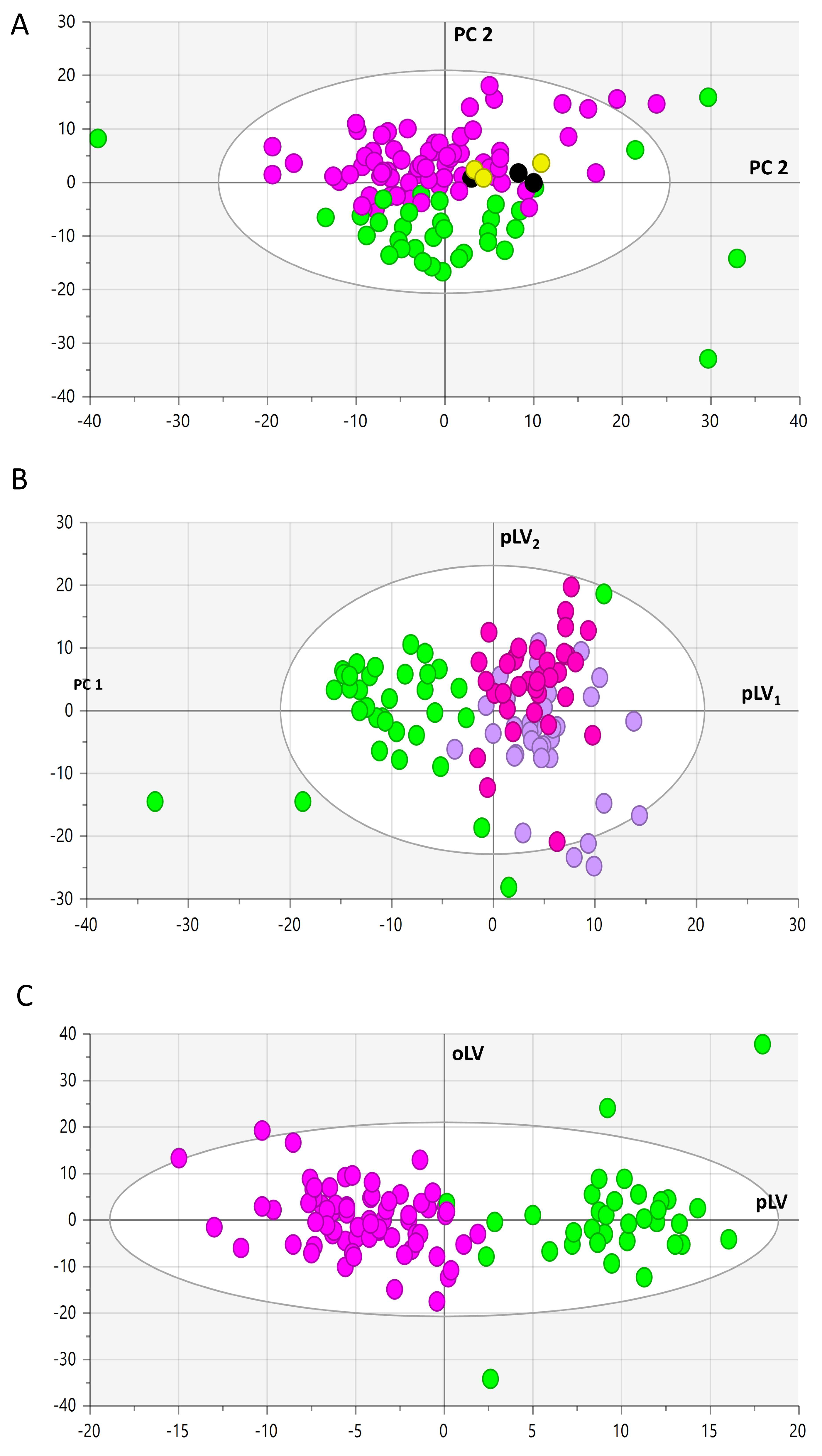
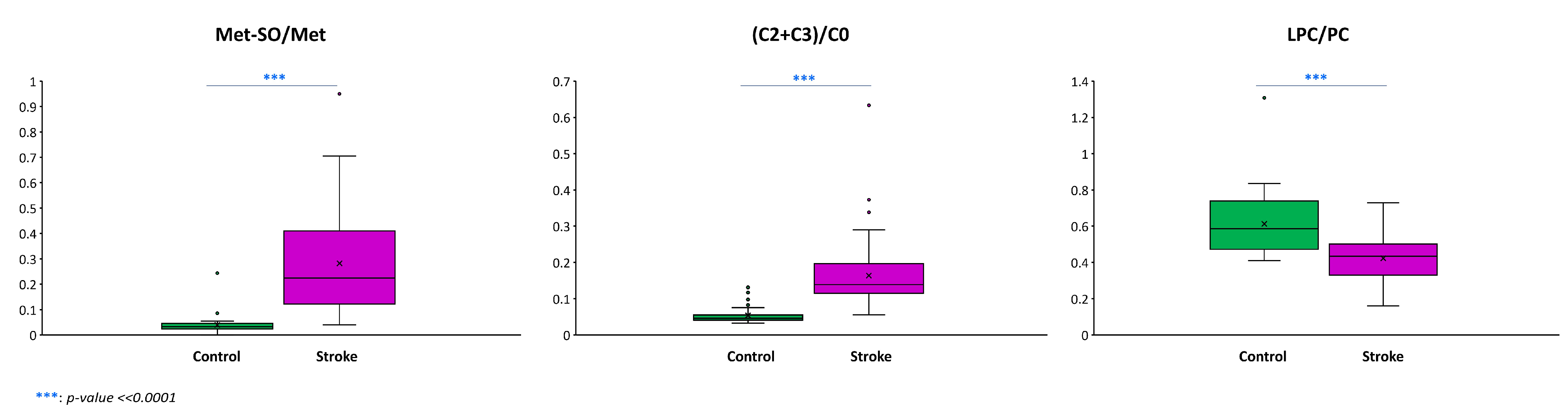
3.2. Metabolomic Analysis
- (1)
- Diminished alkaloid trigonelline.
- (2)
- Diminished non-proteinogenic (ornithine and citrulline) and proteinogenic amino acids (save cysteine with irrelevant variation).
- (3)
- Amino-acid-related molecules: increased levels of methionine sulfoxide, homocysteine, dopamine, serotonin, GABA, α-aminobutyrate, α-aminoadipate, 1-methylhistidine, and polyamines (putrescine, spermidine, and spermine) and decreased levels of betaine, pro-betaine, tryptophane-betaine, sarcosine, and β-alanine.
- (4)
- Other polar metabolites: decreased levels of choline, hippurate, 3-indolepropionate, and xanthine.
- (5)
- Acylcarnitines: increased levels of short-chain acetyl (C2), propionyl (C3), and butyryl (C4) carnitines and long-chain oleoyl (C18:1) and linoleyl (C18:2) carnitines.
- (6)
- Free or non-esterified fatty acids: diminished concentration of arachidonic (AA or FA 20:4 ω6) and docosahexaenoic (DHA or FA 22:6 ω3) acids.
- (7)
- Acylglycerols: diminished concentration of 3 diacylglycerols and increased concentration of 1 diacylglycerol molecule and decreased concentration of 28 triacylglycerol (TAG) species and increased concentration of 57 TAG molecules. Diminished TAGs were mainly polyunsaturated whilst the opposite happened for the increased TAG with a median sum of double bonds for the 3 acyl moieties of 7 and 2, respectively (non-parametric Wilcoxon test’s p-value = 1.062 × 10−13).
- (8)
- Cholesteryl esters (CE): decreased concentration of 7 CE molecules with only one having a saturated acyl moiety and the remaining 6 having acyl moieties with at least 2 double bonds, of which one is an acyl moiety of 20 carbons in length with 4 double bonds (i.e., FA 20:4) and another 22 carbons in length with 6 double bonds (i.e., FA 22:6) (median double bond in acyl moieties of decreased CE of 3) and increased levels of 5 CE species, two of which had a saturated acyl chain, while another two had only one double bond and the other had two double bonds (median double bond in acyl moieties in increased CE of 1). The difference between the median double bonds in acyl moieties of decreased versus increased CE was also statistically significant (non-parametric Wilcoxon test’s p-value = 0.03985).
- (9)
- Decreased levels of dehydroepiandrosterone sulfate (DHEAS) and the bile acid glycolithocholate (GLCA).
- (10)
- Ceramide and hexosylceramides: 7 ceramides and 17 hexosylceramides, of which 10 monohexosylceramides, 6 dihexosylceramides, and 1 trihexosylceramide were found to be all relatively diminished in the stroke group.
- (11)
- Sphingomyelins (SM): diminished concentration of 6 sphingomyelin species and 4 hydroxysphingomyelin molecules.
- (12)
- Lysophosphatidylcholines (LPC): reduced levels of 7 LPCs and increased levels of the less abundant LPC 28:1.
- (13)
- Phosphatidylcholines (PC): diminished concentration of 20 PC species with a median number for the sum of the double bonds of the two acyl moieties of 4 and increased concentration of 20 PC species with a median number for the sum of the double bonds of the two acyl moieties of 2 (non-parametric Wilcoxon test’s p-value = 0.01945).
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Akinyemi, R.O.; Ovbiagele, B.; Adeniji, O.A.; Sarfo, F.S.; Abd-Allah, F.; Adoukonou, T.; Ogah, O.S.; Naidoo, P.; Damasceno, A.; Walker, R.W.; et al. Stroke in Africa: Profile, progress, prospects and priorities. Nat. Rev. Neurol. 2021, 17, 634–656. [Google Scholar] [CrossRef]
- Walker, R.W.; McLarty, D.G.; Kitange, H.M.; Whiting, D.; Masuki, G.; Mtasiwa, D.M.; Machibya, H.; Unwin, N.; Alberti, K.G. Stroke mortality in urban and rural Tanzania. Adult Morbidity and Mortality Project. Lancet 2000, 355, 1684–1687. [Google Scholar] [CrossRef]
- Owolabi, M.O.; Akarolo-Anthony, S.; Akinyemi, R.; Arnett, D.; Gebregziabher, M.; Jenkins, C.; Tiwari, H.; Arulogun, O.; Akpalu, A.; Sarfo, F.S.; et al. The burden of stroke in Africa: A glance at the present and a glimpse into the future. Cardiovasc. J. Afr. 2015, 26, S27–S38. [Google Scholar] [CrossRef]
- Owolabi, M.O.; Sarfo, F.; Akinyemi, R.; Gebregziabher, M.; Akpa, O.; Akpalu, A.; Wahab, K.; Obiako, R.; Owolabi, L.; Ovbiagele, B.; et al. Dominant modifiable risk factors for stroke in Ghana and Nigeria (SIREN): A case-control study. Lancet Glob. Health. 2018, 6, e436–e446. [Google Scholar] [CrossRef]
- Sene-Diouf, F.; Basse, A.M.; Ndiaye, M.; Toure, K.; Diop, M.S.; Thiam, A.; Diop, A.G.; Ndiaye, M.M.; Ndiaye, I.P. The management of cerebrovascular events in Senegal. Rev. Neurol. 2007, 163, 823–827. [Google Scholar] [CrossRef]
- Sacco, R.L.; Kasner, S.E.; Broderick, J.P.; Caplan, L.R.; Connors, J.J.; Culebras, A.; Elkind, M.S.; George, M.G.; Hamdan, A.D.; Higashida, R.T.; et al. An updated definition of stroke for the 21st century: A statement for healthcare professionals from the American Heart Association/AmericanStroke Association. Stroke 2013, 44, 2064–2089. [Google Scholar] [CrossRef]
- Yaghi, S.; Elkind, M.S. Cryptogenic stroke: A diagnostic challenge. Neurol. Clin. Pract. 2014, 4, 386–393. [Google Scholar] [CrossRef]
- Au, A. Metabolomics and Lipidomics of Ischemic Stroke. Adv. Clin. Chem. 2018, 85, 31–69. [Google Scholar]
- Sidorov, E.; Sanghera, D.K.; Vanamala, J.K.P. Biomarker for Ischemic Stroke Using Metabolome: A Clinician Perspective. J. Stroke 2019, 21, 31–41. [Google Scholar] [CrossRef] [PubMed]
- Shin, T.H.; Lee, D.Y.; Basith, S.; Manavalan, B.; Paik, M.J.; Rybinnik, I.; Mouradian, M.M.; Ahn, J.H.; Lee, G. Metabolome Changes in Cerebral Ischemia. Cells 2020, 9, 1630. [Google Scholar] [CrossRef] [PubMed]
- Adams, H.P., Jr.; Bendixen, B.H.; Kappelle, L.J.; Biller, J.; Love, B.B.; Gordon, D.L.; Marsh, E.E., 3rd. Classification of subtype of acute ischemic stroke. Definitions for use in a multicenter clinical trial. TOAST. Trial of Org 10172 in Acute Stroke Treatment. Stroke 1993, 24, 35–41. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: https://www.R-project.org/ (accessed on 7 February 2023).
- Sun, R.; Li, Y.; Cai, M.; Cao, Y.; Piao, X. Discovery of a New Biomarker Pattern for Differential Diagnosis of Acute Ischemic Stroke Using Targeted Metabolomics. Front. Neurol. 2019, 10, 1011. [Google Scholar] [CrossRef] [PubMed]
- Balasubramanian, R.; Hu, J.; Guasch-Ferre, M.; Li, J.; Sorond, F.; Zhao, Y.; Shutta, K.H.; Salas-Salvado, J.; Hu, F.; Clish, C.B.; et al. Metabolomic Profiles Associated With Incident Ischemic Stroke. Neurology 2022, 98, e483–e492. [Google Scholar] [CrossRef] [PubMed]
- Gu, S.X.; Blokhin, I.O.; Wilson, K.M.; Dhanesha, N.; Doddapattar, P.; Grumbach, I.M.; Chauhan, A.K.; Lentz, S.R. Protein methionine oxidation augments reperfusion injury in acute ischemic stroke. JCI Insight 2016, 1, e86460. [Google Scholar] [CrossRef]
- Pinzon, R.T.; Wijaya, V.O.; Veronica, V. The role of homocysteine levels as a risk factor of ischemic stroke events: A systematic review and meta-analysis. Front. Neurol. 2023, 14, 1144584. [Google Scholar] [CrossRef] [PubMed]
- Pravalika, K.; Sarmah, D.; Kaur, H.; Vats, K.; Saraf, J.; Wanve, M.; Kalia, K.; Borah, A.; Yavagal, D.R.; Dave, K.R.; et al. Trigonelline therapy confers neuroprotection by reduced glutathione mediated myeloperoxidase expression in animal model of ischemic stroke. Life Sci. 2019, 216, 49–58. [Google Scholar] [CrossRef] [PubMed]
- Kim, G.H.; Komotar, R.J.; McCullough-Hicks, M.E.; Otten, M.L.; Starke, R.M.; Kellner, C.P.; Garrett, M.C.; Merkow, M.B.; Rynkowski, M.; Dash, K.A.; et al. The role of polyamine metabolism in neuronal injury following cerebral ischemia. Can. J. Neurol. Sci. 2009, 36, 14–19. [Google Scholar] [CrossRef] [PubMed]
- Takano, K.; Ogura, M.; Nakamura, Y.; Yoneda, Y. Neuronal and glial responses to polyamines in the ischemic brain. Curr. Neurovasc. Res. 2005, 2, 213–223. [Google Scholar] [CrossRef]
- Hu, Z.; Fan, S.; Liu, M.; Zhong, J.; Cao, D.; Zheng, P.; Wang, Y.; Wei, Y.; Fang, L.; Xie, P. Objective diagnosis of post-stroke depression using NMR-based plasma metabonomics. Neuropsychiatr. Dis. Treat. 2019, 15, 867–881. [Google Scholar] [CrossRef]
- Wang, X.; Zhang, L.; Sun, W.; Pei, L.L.; Tian, M.; Liang, J.; Liu, X.; Zhang, R.; Fang, H.; Wu, J.; et al. Changes of Metabolites in Acute Ischemic Stroke and Its Subtypes. Front. Neurosci. 2021, 14, 580929. [Google Scholar] [CrossRef]
- Jiménez, M.C.; Sun, Q.; Schürks, M.; Chiuve, S.; Hu, F.B.; Manson, J.E.; Rexrode, K.M. Low dehydroepiandrosterone sulfate is associated with increased risk of ischemic stroke among women. Stroke 2013, 44, 1784–1789. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.J.; Ngo, D.; Psychogios, N.; Dejam, A.; Larson, M.G.; Vasan, R.S.; Ghorbani, A.; O’Sullivan, J.; Cheng, S.; Rhee, E.P.; et al. 2-Aminoadipic acid is a biomarker for diabetes risk. J. Clin. Investig. 2013, 123, 4309–4317. [Google Scholar] [CrossRef] [PubMed]
- Lee, T.H.; Cheng, M.L.; Shiao, M.S.; Lin, C.N. Metabolomics study in severe extracranial carotid artery stenosis. BMC Neurol. 2019, 19, 138. [Google Scholar] [CrossRef] [PubMed]
- Zhong, C.; Miao, M.; Che, B.; Du, J.; Wang, A.; Peng, H.; Bu, X.; Zhang, J.; Ju, Z.; Xu, T.; et al. Plasma choline and betaine and risks of cardiovascular events and recurrent stroke after ischemic stroke. Am. J. Clin. Nutr. 2021, 114, 1351–1359. [Google Scholar] [CrossRef] [PubMed]
- Tiedje, K.E.; Stevens, K.; Barnes, S.; Weaver, D.F. Beta-alanine as a small molecule neurotransmitter. Neurochem. Int. 2010, 57, 177–188. [Google Scholar] [CrossRef]
- Kopach, O.; Rusakov, D.A.; Sylantyev, S. Multi-target action of β-alanine protects cerebellar tissue from ischemic damage. Cell Death Dis. 2022, 13, 747. [Google Scholar] [CrossRef]
- Yu, H.; Chen, X.; Guo, X.; Chen, D.; Jiang, L.; Qi, Y.; Shao, J.; Tao, L.; Hang, J.; Lu, G.; et al. The clinical value of serum xanthine oxidase levels in patients with acute ischemic stroke. Redox Biol. 2023, 60, 102623. [Google Scholar] [CrossRef]
- Maciejczyk, M.; Nesterowicz, M.; Zalewska, A.; Biedrzycki, G.; Gerreth, P.; Hojan, K.; Gerreth, K. Salivary Xanthine Oxidase as a Potential Biomarker in Stroke Diagnostics. Front. Immunol. 2022, 13, 897413. [Google Scholar] [CrossRef]
- Chyan, Y.J.; Poeggeler, B.; Omar, R.A.; Chain, D.G.; Frangione, B.; Ghiso, J.; Pappolla, M.A. Potent neuroprotective properties against the Alzheimer beta-amyloid by an endogenous melatonin-related indole structure, indole-3-propionic acid. J. Biol. Chem. 1999, 274, 21937–21942. [Google Scholar] [CrossRef]
- Xie, Y.; Zou, X.; Han, J.; Zhang, Z.; Feng, Z.; Ouyang, Q.; Hua, S.; Liu, Z.; Li, C.; Cai, Y.; et al. Indole-3-propionic acid alleviates ischemic brain injury in a mouse middle cerebral artery occlusion model. Exp. Neurol. 2022, 353, 114081. [Google Scholar] [CrossRef]
- Rodrigues, C.M.; Spellman, S.R.; Solá, S.; Grande, A.W.; Linehan-Stieers, C.; Low, W.C.; Steer, C.J. Neuroprotection by a bile acid in an acute stroke model in the rat. J. Cereb. Blood Flow Metab. 2002, 22, 463–471. [Google Scholar] [CrossRef]
- Yamagata, K. Docosahexaenoic acid inhibits ischemic stroke to reduce vascular dementia and Alzheimer’s disease. Prostaglandins Other Lipid Mediat. 2023, 167, 106733. [Google Scholar] [CrossRef]
- Rink, C.; Khanna, S. Significance of brain tissue oxygenation and the arachidonic acid cascade in stroke. Antioxid. Redox Signal. 2011, 14, 1889–1903. [Google Scholar] [CrossRef]
- Jové, M.; Mota-Martorell, N.; Pradas, I.; Galo-Licona, J.D.; Martín-Gari, M.; Obis, È.; Sol, J.; Pamplona, R. The Lipidome Fingerprint of Longevity. Molecules 2020, 25, 4343. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cryptogenic Stroke (n = 40) | Determined Stroke (n = 39) | Controls (n = 40) | p-Value | |
|---|---|---|---|---|
| Epidemiological parameters | ||||
| Mean age (years) | 47 ± 11 | 57.02 ± 14.91 | 47 ± 12 | <0.001 |
| Sex ratio | 1.35 | 1.35 | 1.35 | - |
| Biological parameters | ||||
| Total cholesterol (mmol/L) | 5.08 ± 1.78 | 4.98 ± 1.78 | 5.06 ± 0.80 | 0.904 |
| HDL-c (mmol/L) | 1.19 ± 0.33 | 1.14 ± 0.33 | 1.52 ± 0.25 | 0.353 |
| LDL-c (mmol/L) | 2.71 ± 1.26 | 2.68 ± 1.21 | 3.33 ± 0.62 | 0.762 |
| Triglycerides (mmol/L) | 0.98 ± 0.44 | 1.04 ± 0.54 | 0.89 ± 0.41 | 0.725 |
| Blood glucose (mmol/L) | 5.3 ± 0.99 | 6.6 ± 2.03 | - | <0.001 |
| Urea (mmol/L) | 3.7 ± 2.66 | 4.80 ± 2.81 | 2.7 ± 0.63 | 0.007 |
| Creatinine (µmol/L) | 65.68 ± 13.61 | 66.12 ± 25.72 | 70.54 ± 10.96 | 0.254 |
| Uric acid (µmol/L) | 332.9 ± 73.75 | 337.6 ± 128.47 | 286 ± 53.53 | 0.912 |
| CRP (nmol/L) | 32.47 ± 24.76 | 409.52 ± 620.96 | 18.38 ± 18.09 | <0.001 |
| AST (µkat/L) | 0.57 ± 0.26 | 0.97 ± 0.99 | 0.39 ± 0.09 | 0.012 |
| ALT (µkat/L) | 0.42 ± 0.39 | 0.50 ± 0.65 | 0.24 ± 0.15 | 0.123 |
| GGT (µkat/L) | 1.01 ± 0.97 | 1.70 ± 3.13 | 0.54 ± 0.24 | 0.080 |
| ALP (µkat/L) | 1 ± 0.35 | 1.50 ± 1.50 | - | 0.026 |
| Clinical and lifestyle parameters | ||||
| HTA (%) | 45 | 84.61 | - | 0.002 |
| Diabetes (%) | 15 | 61.53 | - | 0.040 |
| Dyslipidemia (%) | 22.50 | 64.10 | - | 0.012 |
| Sedentary lifestyle (%) | 25 | 51.28 | - | <0.001 |
| Smoking (%) | 25 | 17.94 | - | <0.001 |
| Alcoholism (%) | 15 | 2.56 | - | <0.001 |
| Drugs (%) | 5 | - | - | - |
| Related to oxidative stress | Methionine sulfoxide, Met-SO/Met ratio, homocysteine, trigonelline, spermine, spermidine, putrescine, amino acids, α-aminoadipate, xanthine, 3-Indolepropionic, overall increase in the saturation of fatty acids present in complex lipids |
| Related to energetic crisis | Acylcarnitines C2, C3, C4, C18:1, and C18:2 (C2 + C3)/C0 ratio, trigonelline, amino acids |
| Related to cell membrane damage | Choline, betaine, Pro-betaine, and Trp-betaine, docosahexaenoic acid, phosphatidylcholines, lysophosphatidylcholines, sphingomyelins, hydroxysphingomyelins, ceramides, hexosylceramides, cholesterol esters, diacylglycerols, diacylglycerols |
| Related to one-carbon metabolism | Methionine, methionine sulfoxide, homocysteine, choline, betaine, Pro-betaine, and Trp-betaine, sarcosine |
| Related to neurotransmission and cell signaling | Spermine, spermidine, putrescine, amino acids, dopamine, serotonin, GABA, α-aminobutyric acid, choline, beta-alanine, dehydroepiandrosterone sulfate, α-aminobutyric acid, arachidonic acid |
| Gut microbiota and or diet-related metabolites | Trigonelline, hippuric acid, 3-indolepropionic, glycolithocholic acid sulfate (GLCA), docosahexaenoic acid (DHA) |
| Related to inflammation | Methionine sulfoxide, homocysteine, docosahexaenoic acid (DHA), arachidonic acid |
| Decreased concentration of neuroprotective metabolites | Trigonelline, beta-alanine, 3-indolepropionic, glycolithocholic acid sulfate (GLCA), dehydroepiandrosterone sulfate (DHEAS), docosahexaenoic acid (DHA) |
| New metabolites related to acute ischemic stroke | Methionine sulfoxide, Met-SO/Met ratio, trigonelline, 1-methyl-histidine, dopamine, serotonin, GABA, α-aminobutyric acid, α-aminoadipate, sarcosine, hippuric acid, xanthine, glycolithocholic acid sulfate (GLCA), dehydroepiandrosterone sulfate (DHEAS) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Djite, M.; Chao de la Barca, J.M.; Bocca, C.; Gaye, N.M.; Barry, N.O.K.; Mbacke, M.N.; Cissé, O.; Kandji, P.M.; Thioune, N.M.; Coly-Gueye, N.F.; et al. A Metabolomic Signature of Ischemic Stroke Showing Acute Oxidative and Energetic Stress. Antioxidants 2024, 13, 60. https://doi.org/10.3390/antiox13010060
Djite M, Chao de la Barca JM, Bocca C, Gaye NM, Barry NOK, Mbacke MN, Cissé O, Kandji PM, Thioune NM, Coly-Gueye NF, et al. A Metabolomic Signature of Ischemic Stroke Showing Acute Oxidative and Energetic Stress. Antioxidants. 2024; 13(1):60. https://doi.org/10.3390/antiox13010060
Chicago/Turabian StyleDjite, Moustapha, Juan Manuel Chao de la Barca, Cinzia Bocca, Ndiaga Matar Gaye, Néné Oumou Kesso Barry, Mame Ndoumbé Mbacke, Ousmane Cissé, Pape Matar Kandji, Ndèye Marème Thioune, Najah Fatou Coly-Gueye, and et al. 2024. "A Metabolomic Signature of Ischemic Stroke Showing Acute Oxidative and Energetic Stress" Antioxidants 13, no. 1: 60. https://doi.org/10.3390/antiox13010060