

Effects of Enhanced Resistance and Transcriptome Analysis of Twig Blight Disease by Exogenous Brassinolide in Myrica rubra
(This article belongs to the Section Health Outcomes of Antioxidants and Oxidative Stress)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
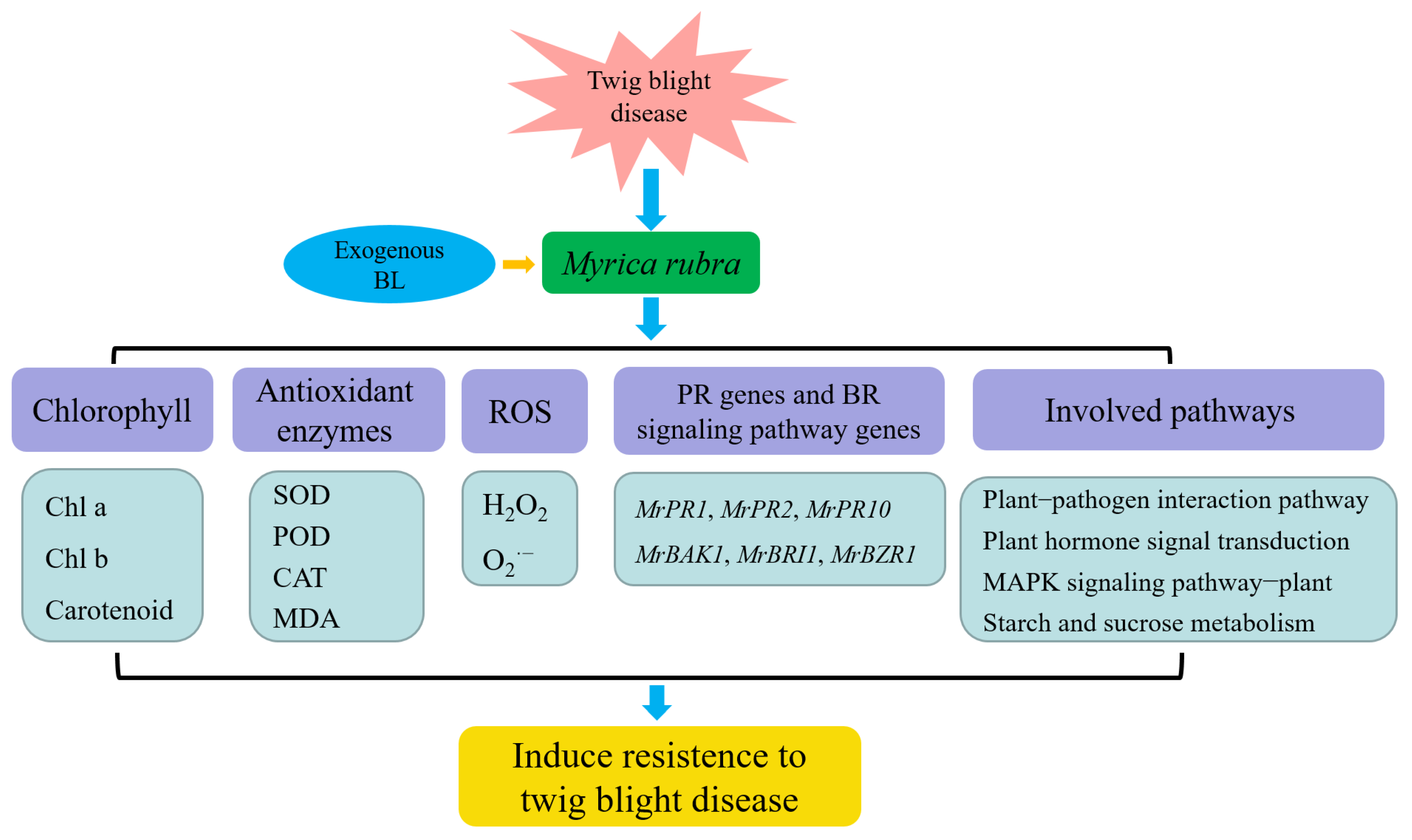
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Materials
2.2. Inoculation
2.3. Determination of Chlorophyll Content
2.4. Determination of Antioxidant Enzyme Activity and ROS Accumulation
2.5. qRT-PCR Analysis
2.6. RNA-seq Analysis
2.7. Data Analysis
3. Results
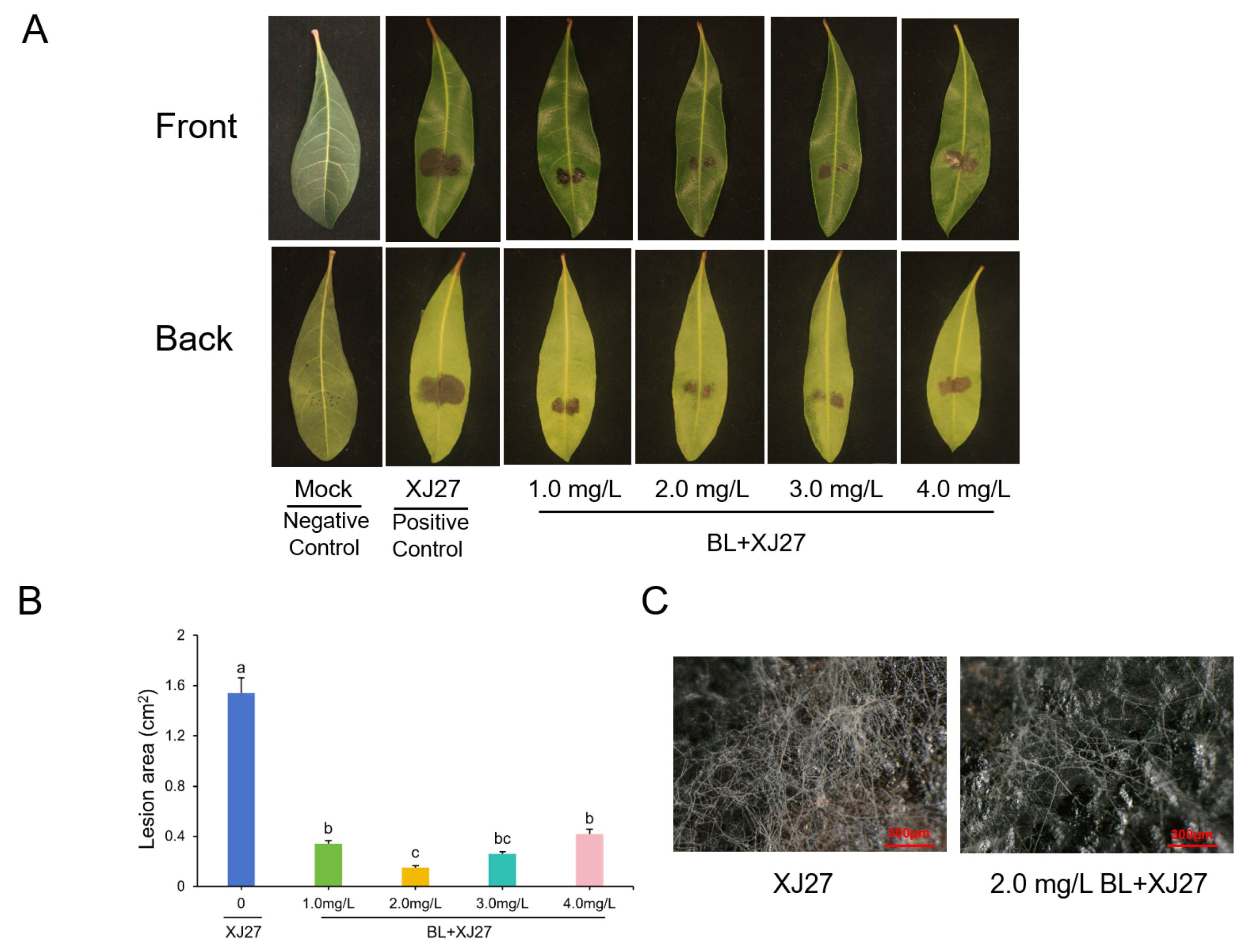
3.1. Exogenous BL Significantly Reduces the Level of Twig Blight Disease in M. rubra Leaves
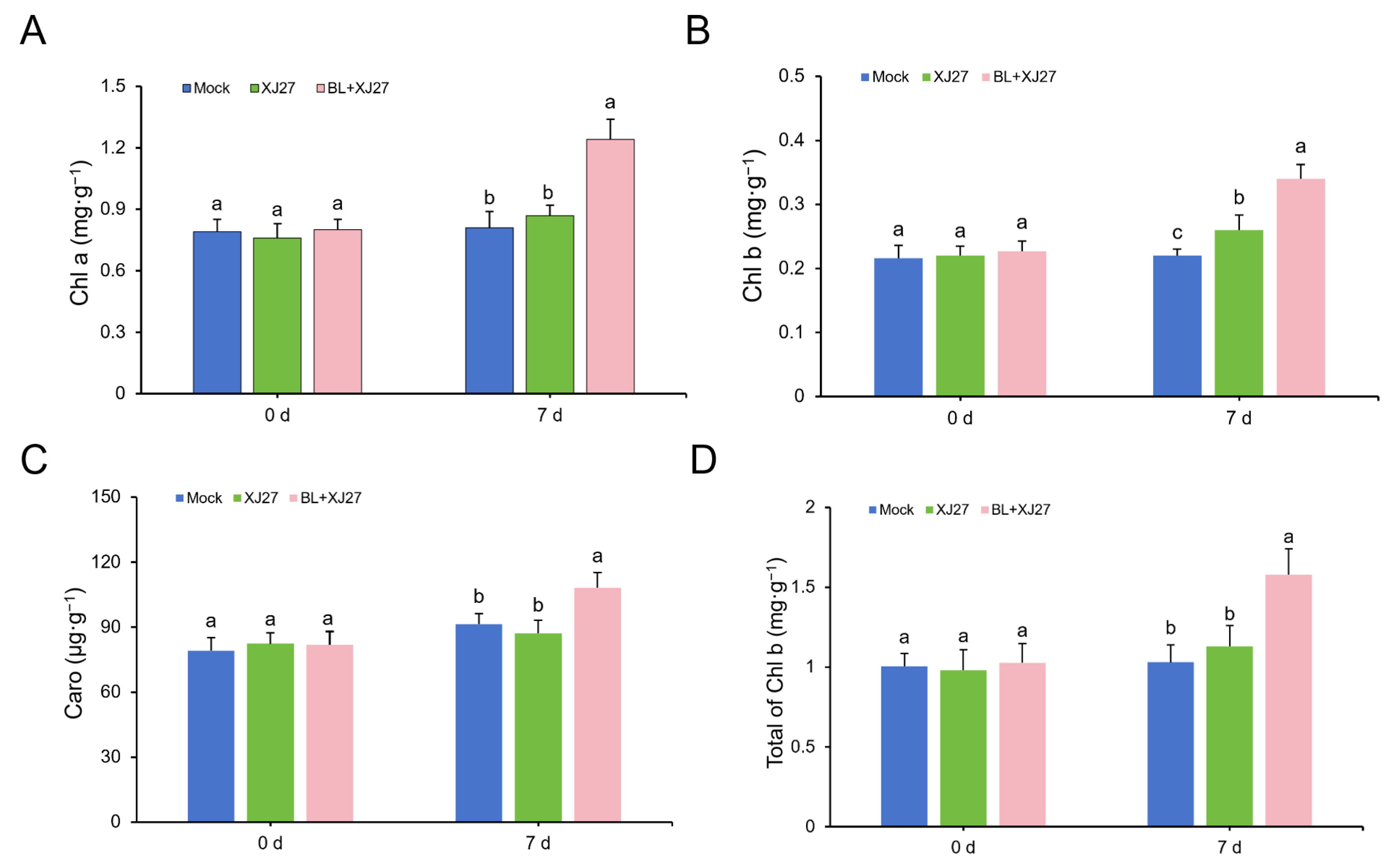
3.2. Exogenous BL Treatment Increases Chlorophyll Content in M. rubra Leaves
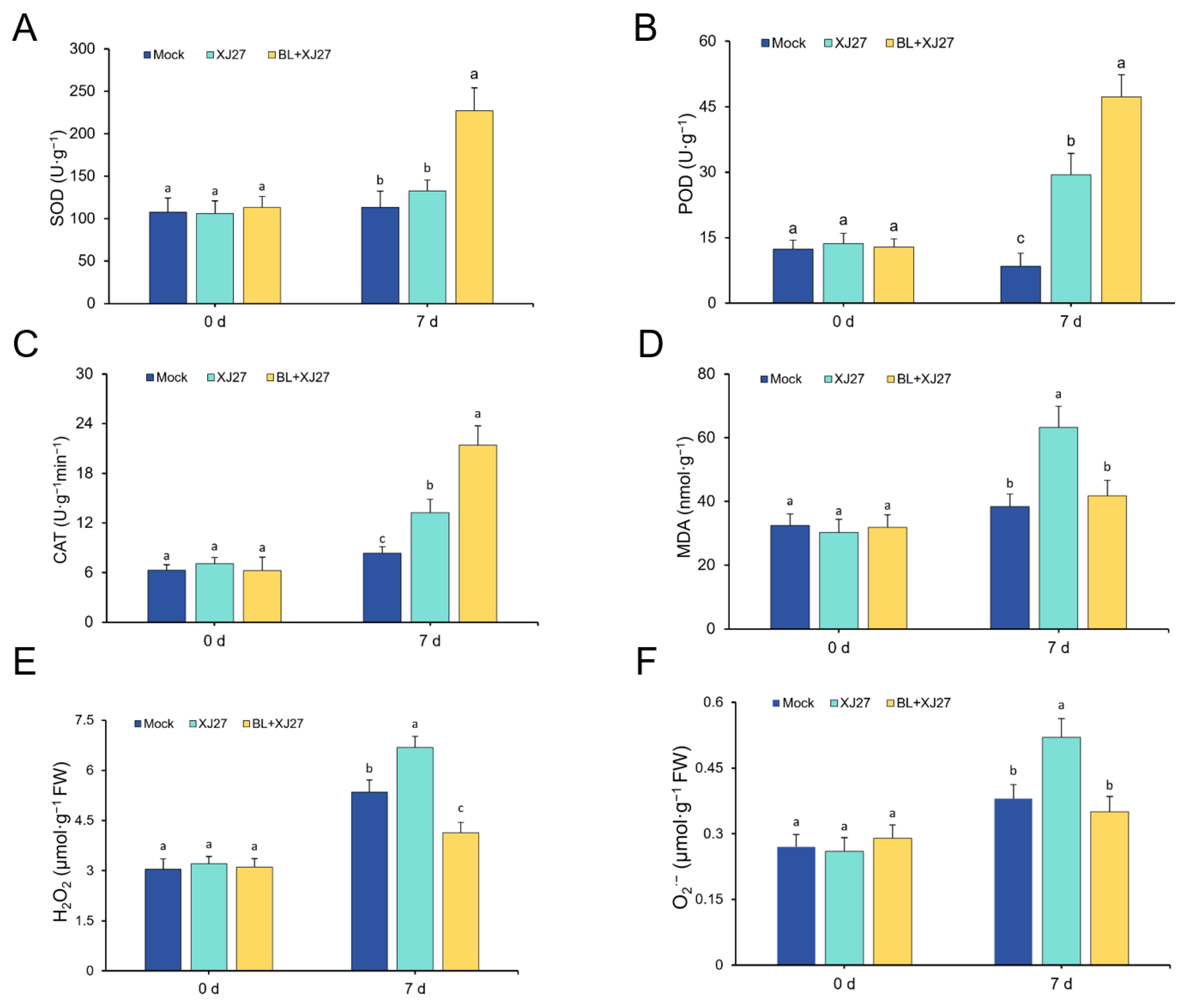
3.3. Exogenous BL Increases Antioxidant Enzyme Activity and ROS Accumulation in M. rubra
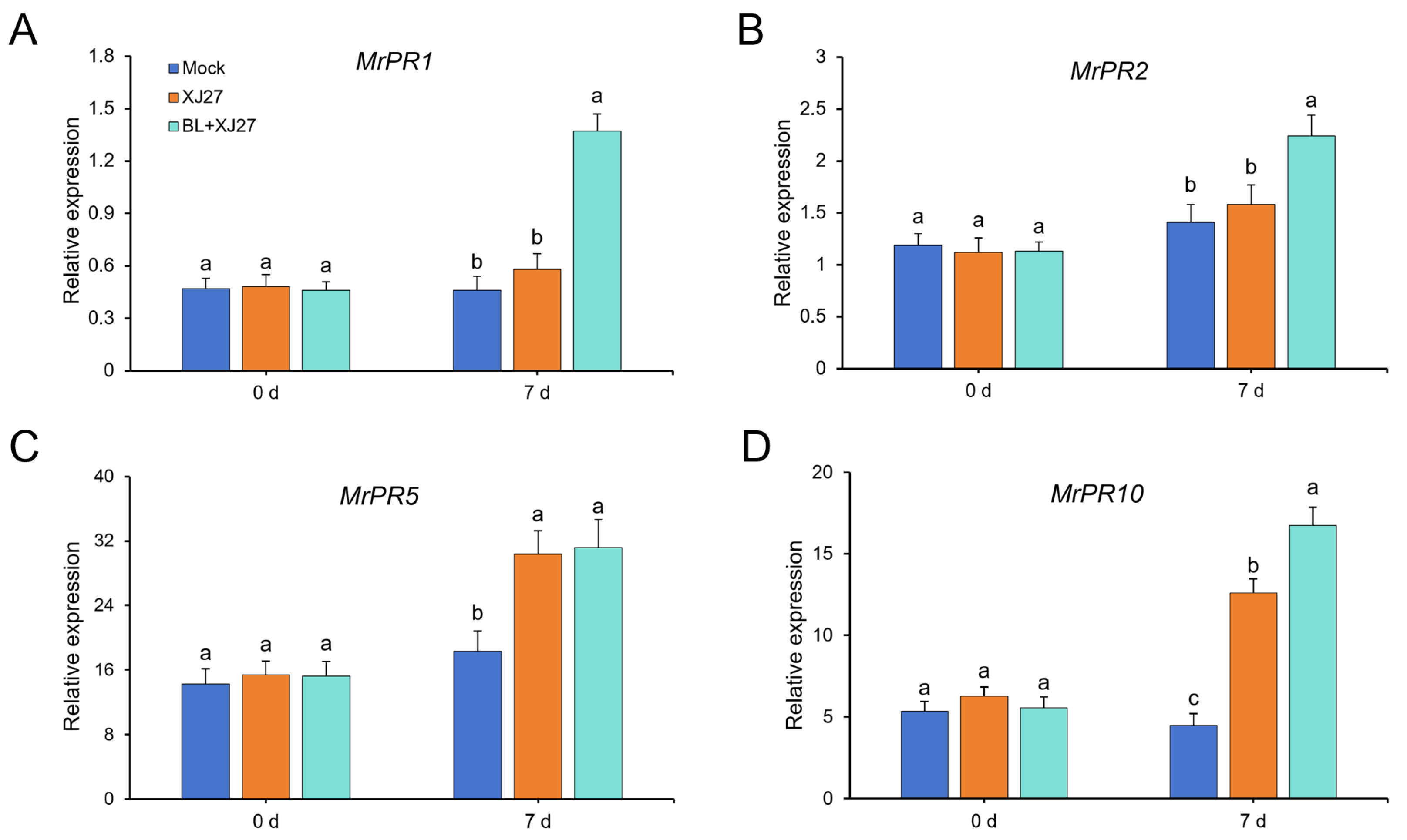
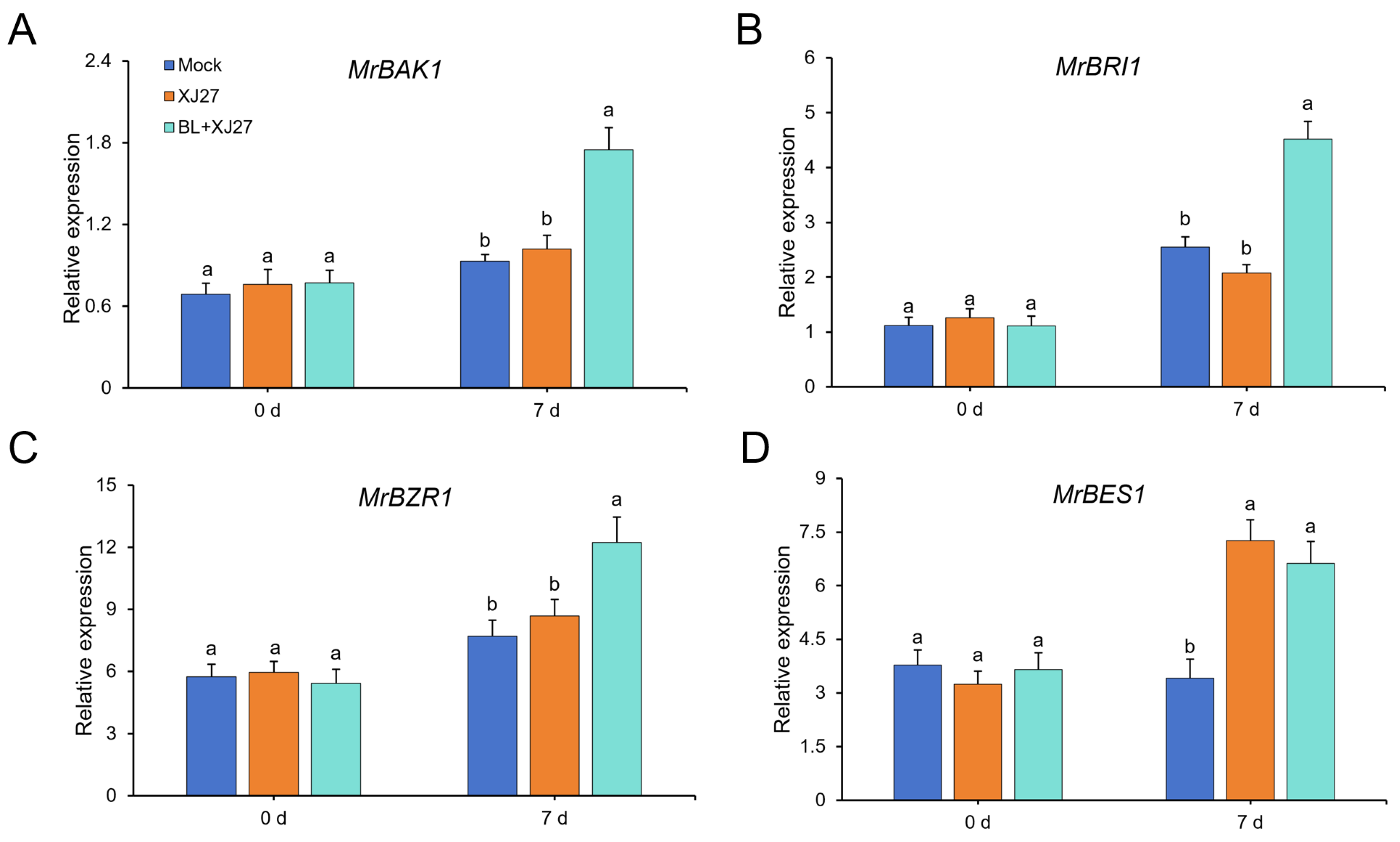
3.4. Effects of Exogenous BR on Expression of PR Genes and BR Signaling-Related Genes
3.5. RNA-seq Analysis of M. rubra Leaves under Exogenous BR Treatment
3.5.1. Transcriptome Data Quality Analysis
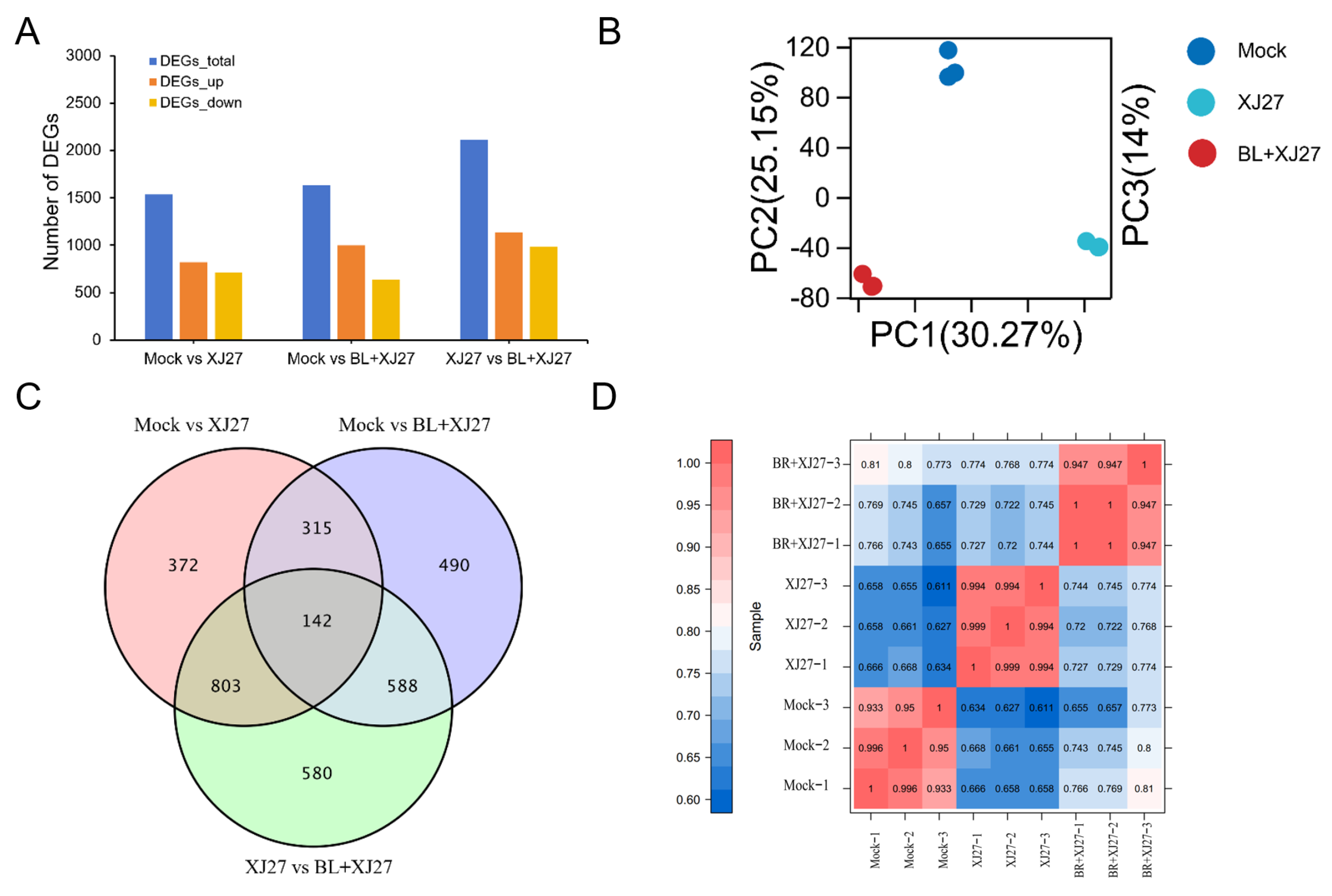
3.5.2. Analysis of DEGs
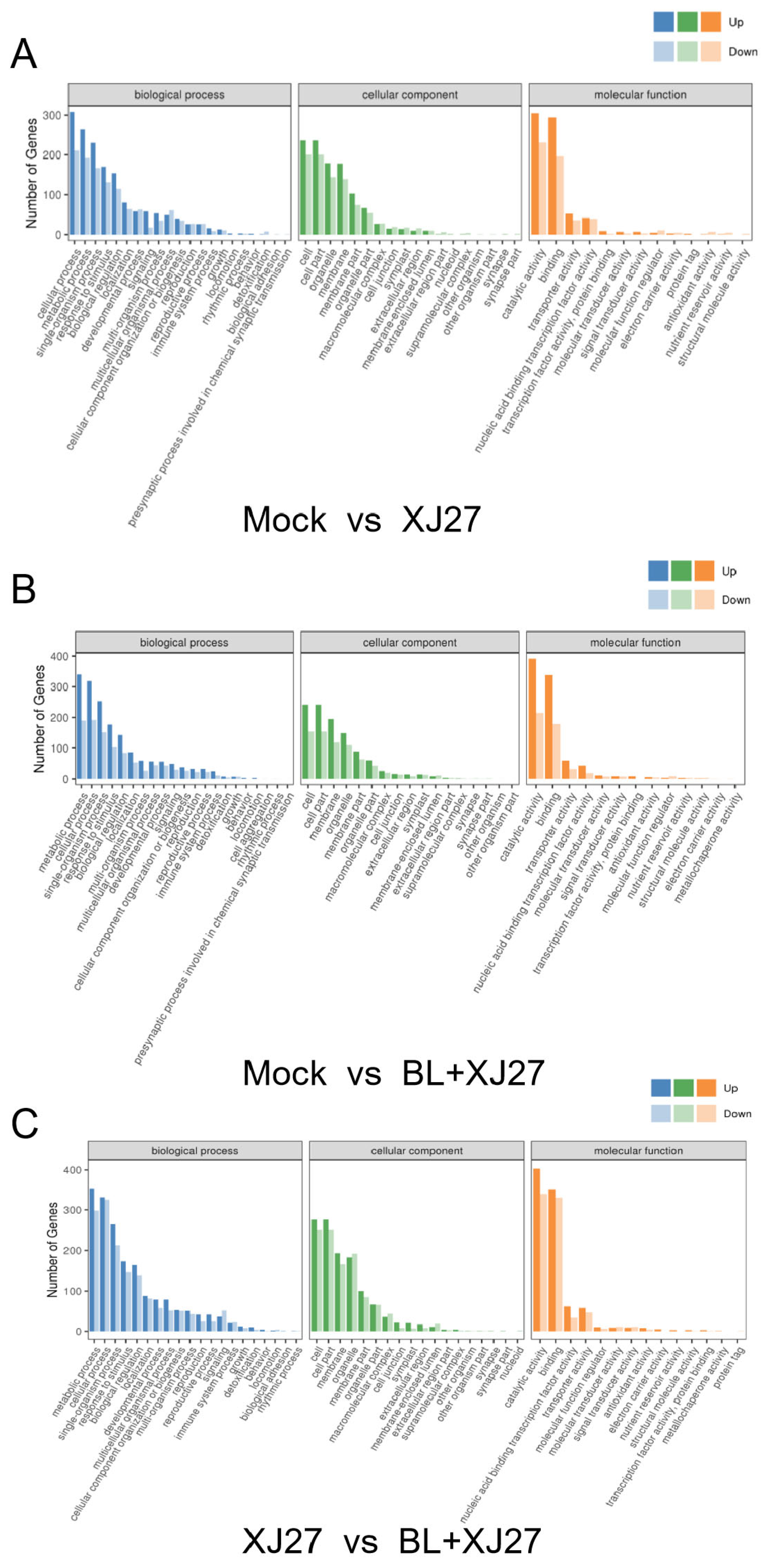
3.5.3. GO Enrichment Analysis
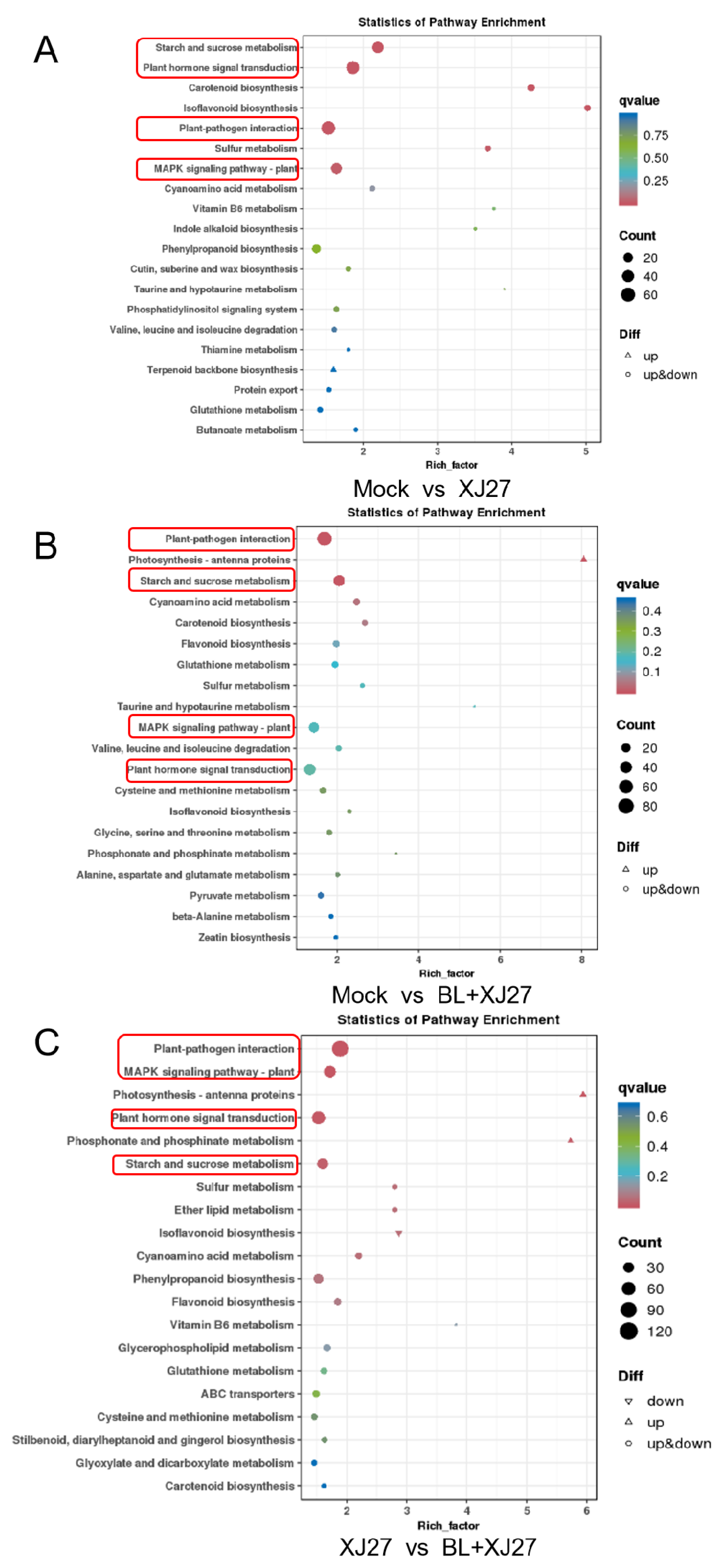
3.5.4. KEGG Enrichment Analysis
3.5.5. Identification of Candidate Genes Responding to BR in M. rubra
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhang, S.W.; Yu, Z.P.; Qi, X.J.; Wang, Z.; Zheng, Y.Y.; Ren, H.Y.; Liang, S.M.; Zheng, X.L. Construction of a high-density genetic map and identification of leaf trait-related QTLs in Chinese Bayberry (Myrica rubra). Front. Plant Sci. 2021, 12, 675855. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.W.; Yu, Z.P.; Sun, L.; Ren, H.Y.; Zheng, X.L.; Liang, S.M.; Qi, X.J. An overview of the nutritional value, health properties, and future challenges of Chinese bayberry. PeerJ 2022, 10, e13070. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.Y.; Yu, Z.P.; Li, Y.G.; Zhong, S.; Sun, Y.Q.; Sun, L.; Zheng, X.L.; Qi, X.J.; Zhang, S.W. Alcohol extracts of Chinese bayberry branch induce S-phase arrest and apoptosis in HepG2 cells. Food Sci. Nutr. 2022, 11, 493–503. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.D.; Huang, H.Z.; Xu, C.J.; Li, X.; Chen, K.S. Biological activities of extracts from Chinese Bayberry (Myrica rubra Sieb. et Zucc.): A review. Plant Foods Hum. Nutr. 2013, 68, 97–106. [Google Scholar] [CrossRef] [PubMed]
- Ren, H.Y.; Li, G.; Qi, X.J.; Li, F.; Wang, H.R.; Wei, J.G.; Zhong, S.B. Identification and characterization of Pestalotiopsis spp. causing twig blight disease of bayberry (Myrica rubra Sieb. & Zucc) in China. Eur. J. Plant Pathol. 2013, 137, 451–461. [Google Scholar]
- Fang, L.; Wang, H.R.; Feng, J.J. Branch blight of loquat caused by Pestalotiopsis sydowiana in China. Plant Dis. 2013, 97, 990. [Google Scholar] [CrossRef] [PubMed]
- Espinoza, J.G.; Briceño, E.X.; Keith, L.M.; Latorre, B.A. Canker and twig dieback of blueberry caused by Pestalotiopsis spp. and a Truncatella sp. in Chile. Plant Dis. 2008, 92, 1407–1414. [Google Scholar] [CrossRef] [PubMed]
- Chamorro, M.; Aguado, A.; Santos, B.D.L. First report of root and crown rot caused by Pestalotiopsis clavispora (Neopestalotiopsis clavispora) on strawberry in Spain. Plant Dis. 2015, 100, 3. [Google Scholar] [CrossRef]
- Wang, Y.; Xiong, F.; Lu, Q. Diversity of Pestalotiopsis-like species causing gray blight disease of tea plants (Camellia sinensis) in China, including two novel Pestalotiopsis Species, and analysis of their pathogenicity. Plant Dis. 2019, 103, 2548–2558. [Google Scholar] [CrossRef]
- Goode, K.; Mitchum, M.G. Pattern-triggered immunity against root-knot nematode infection: A minireview. Physiol. Plant. 2022, 174, e13680. [Google Scholar] [CrossRef]
- Yu, J.J.; Gonzalez, J.M.; Dong, Z.P.; Shan, Q.R.; Tan, B.W.; Koh, J.; Zhang, T.; Zhu, N.; Dufresne, C.P.; Martin, G.B.; et al. Integrative proteomic and phosphoproteomic analyses of pattern- and effector-triggered immunity in tomato. Front. Plant Sci. 2021, 12, 768693. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.C.; Dolatabadian, A.; Fernando, W.G.D. The wonderful world of intrinsic and intricate immunity responses in plants against pathogens. Can. J. Plant Pathol. 2021, 44, 1–20. [Google Scholar] [CrossRef]
- Katagiri, F.; Tsuda, K. Understanding the plant immune system. Mol. Plant Microbe Interact. 2010, 23, 1531–1536. [Google Scholar] [CrossRef] [PubMed]
- Kamiya, Y. Plant hormones: Versatile regulators of plant growth and development. Annu. Rev. Plant Biol. 2010, 61. [Google Scholar] [CrossRef]
- Pieterse, C.M.J.; Does, D.V.D.; Zamioudis, C.; Reyes, A.L.; Wees, S.C.M.V. Hormonal modulation of plant immunity. Annu. Rev. Cell Dev. Biol. 2012, 28, 489–521. [Google Scholar] [CrossRef] [PubMed]
- Hussain, M.A.; Fahad, S.; Sharif, R.; Jan, M.F.; Mujtaba, M.; Ali, Q.; Ahmad, A.; Ahmad, H.; Amin, N.; Ajayo, B.S.; et al. Multifunctional role of brassinosteroid and its analogues in plants. Plant Growth Regul. 2020, 92, 141–156. [Google Scholar] [CrossRef]
- Mao, J.P.; Zhang, D.; Li, K.; Liu, Z.; Liu, X.J.; Song, C.H.; Li, G.F.; Zhao, C.P.; Ma, J.J.; Han, M.Y. Effect of exogenous Brassinolide (BR) application on the morphology, hormone status, and gene expression of developing lateral roots in Malus hupehensis. Plant Growth Regul. 2017, 82, 391–401. [Google Scholar] [CrossRef]
- Ainoa, P.R.; Aditi, G.; Isabel, B.P.; Nadja, B.; Marta, I.; Ana, I.C.D. Brassinosteroid signaling in plant development and adaptation to stress. Development 2019, 146, dev151894. [Google Scholar]
- Nolan, T.M.; Vukašinović, N.; Liu, D.R.; Russinova, E.; Yin, Y.H. Brassinosteroids: Multidimensional regulators of plant growth, development, and stress responses. Plant Cell 2019, 32, 295–318. [Google Scholar] [CrossRef]
- Fariduddin, Q.; Khanam, S.; Hasan, S.A.; Ali, B.; Hayat, S.; Ahmad, A. Effect of 28-homobrassinolide on the drought stress-induced changes in photosynthesis and antioxidant system of Brassica juncea L. Acta Physiol. Plant 2009, 31, 889–897. [Google Scholar] [CrossRef]
- Mahesh, K.; Balaraju, P.; Ramakrishna, B.; Rao, S.S.R. Effect of brassinosteroids on germination and seedling growth of radish (Raphanus sativus L.) under PEG-6000 induced water stress. Am. J. Plant Sci. 2013, 4, 2305–2313. [Google Scholar] [CrossRef]
- Fariduddin, Q.; Khalil, R.R.A.E.; Mir, B.A.; Yusuf, M.; Ahmad, A. 24-Epibrassinolide regulates photosynthesis, antioxidant enzyme activities and proline content of Cucumis sativus under salt and/or copper stress. Environ. Monit. Assess. 2013, 185, 7845–7856. [Google Scholar] [CrossRef] [PubMed]
- Ogweno, J.O.; Song, X.S.; Shi, K.; Hu, W.H.; Mao, W.H.; Zhou, Y.H.; Yu, J.Q.; Nogués, S. Brassinosteroids alleviate heat-induced inhibition of photosynthesis by increasing carboxylation efficiency and enhancing antioxidant systems in Lycopersicon esculentum. J. Plant Growth Regul. 2008, 27, 49–57. [Google Scholar] [CrossRef]
- Wang, Q.; Ding, T.; Gao, L.; Pang, J.; Yang, N. Effect of brassinolide on chilling injury of green bell pepper in storage. Sci. Hortic. 2012, 144, 195–200. [Google Scholar] [CrossRef]
- Nakashita, H.; Yasuda, M.; Nitta, T.; Asami, T.; Fujioka, S.; Arai, Y.; Sekimata, K.; Takatsuto, S.; Yamaguchi, I.; Yoshida, S. Brassinosteroid functions in a broad range of disease resistance in tobacco and rice. Plant J. 2003, 33, 887–898. [Google Scholar] [CrossRef] [PubMed]
- Thirunahari, U.; Bala, R.P.; Ayodhya, R.; Mandaloju, V. Influence of peg imposed water stress and exogenous application of brassinosteroids on metabolites in radish. Asian J. Sci. Technol. 2015, 6, 951–955. [Google Scholar]
- Sun, Y.; Asghari, M.; Zahedipour-Sheshgelani, P. Foliar spray with 24-epibrassinolide enhanced strawberry fruit quality, phytochemical content, and postharvest life. J. Plant Growth Regul. 2019, 39, 920–929. [Google Scholar] [CrossRef]
- Wang, X.F.; Kota, U.; He, K.; Blackburn, K.; Li, J.; Goshe, M.B.; Huber, S.C.; Clouse, S.D. Sequential transphosphorylation of the BRI1/BAK1 receptor kinase complex impacts early events in brassinosteroid signaling. Dev. Cell 2008, 15, 220–235. [Google Scholar] [CrossRef]
- Ryu, H.; Kim, K.; Hwang, I. Spatial redistribution of key transcriptional regulators in brassinosteroid signaling. Plant Signal. Behav. 2008, 3, 278–280. [Google Scholar] [CrossRef]
- Joshi, V.; Joshi, N.; Vyas, A.; Jadhav, S.K. 25-Pathogenesis-related proteins: Role in plant defense. In Biocontrol Agents and Secondary Metabolites; Woodhead Publishing: Cambridge, UK, 2021; pp. 573–590. [Google Scholar]
- Santos, C.D.; Franco, O.L. Pathogenesis-related proteins (PRs) with enzyme activity activating plant defense responses. Plants 2023, 12, 2226. [Google Scholar] [CrossRef]
- Raji, M.R.; Lotfi, M.; Tohidfar, M.; Ramshini, H.; Sahebani, N.; Alifar, M.; Baratian, M.; Mercati, F.; Michele, R.D.; Carimi, F. Multiple fungal diseases resistance induction in Cucumis melo through co-transformation of different pathogenesis related (PR) protein genes. Sci. Hortic. 2022, 297, 110924. [Google Scholar] [CrossRef]
- Goto, K.; Yabuta, S.; Ssenyonga, P.; Tamaru, S.; Sakagami, J.I. Response of leaf water potential, stomatal conductance and chlorophyll content under different levels of soil water, air vapor pressure deficit and solar radiation in chili pepper (Capsicum chinense). Sci. Hortic. 2021, 281, 109943. [Google Scholar] [CrossRef]
- Dai, X.; Wang, Y.; Chen, Y.; Li, H.; Xu, S.; Yang, T.; Zhang, X.; Su, X.; Xia, Z. Overexpression of NtDOG1L-T improves heat stress tolerance by modulation of antioxidant capability and defense-, heat-, and ABA-related gene expression in tobacco. Front. Plant Sci. 2020, 11, 568489. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.Y.; Li, F.F.; Xu, T.; Cai, S.J.; Chu, W.Y.; Qiu, H.; Sha, S.; Cheng, G.Y.; Xu, Q.S. Bioaccumulation, subcellular, and molecular localization and damage to physiology and ultrastructure in Nymphoides peltata (Gmel.) O. Kuntze exposed to yttrium. Environ Sci. Pollut. Res. Int. 2013, 21, 2935–2942. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.F.; Miao, J.J.; Pan, L.Q.; Li, Y.; Lin, Y.F.; Wu, J.Y. Toxicity effects of p-choroaniline on the growth, photosynthesis, respiration capacity and antioxidant enzyme activities of a diatom, Phaeodactylum tricornutu. Ecotox. Environ. Safe. 2019, 169, 654–661. [Google Scholar] [CrossRef] [PubMed]
- Gautam, V.; Kohli, S.; Kapoor, D.; Bakshi, P.; Sharma, P.; Arora, S.; Bhardwaj, R.; Ahmad, P. Stress protective effect of Rhododendron arboretum leaves (MEL) on Chromium-Treated Vigna radiate plants. J. Plant Growth Regul. 2021, 40, 423–435. [Google Scholar] [CrossRef]
- Wang, F.B.; Liu, J.C.; Zhou, L.J.; Pan, G.; Li, Z.W.; Zaidi, S.H.R.; Cheng, F.M. Senescence-specific change in ROS scavenging enzyme activities and regulation of various SOD isozymes to ROS levels in psf mutant rice leaves. Plant Physiol. Biochem. 2016, 109, 248–261. [Google Scholar] [CrossRef]
- Martins, L.; Fernandes, C.B.; Albuquerque, P.Z.; Tavares, F. Assessment of Xanthomonas arboricola pv. juglandis Bacterial load in infected walnut fruits by quantitative PCR. Plant Dis. 2019, 103, 2577–2586. [Google Scholar] [CrossRef]
- Young, M.D.; Wakefield, M.J.; Smyth, G.K.; Oshlack, A. Gene ontology analysis for RNAseq: Accounting for selection bias. Genome Biol. 2010, 11, R14. [Google Scholar] [CrossRef]
- Mao, X.Z.; Cai, T.; Olyarchuk, J.G.; Wei, L.P. Automated genome annotation and pathway identification using the KEGG Orthology (KO) as a controlled vocabulary. Bioinformatics 2005, 21, 3787–3793. [Google Scholar] [CrossRef]
- Liu, Q.; Xi, Z.M.; Gao, J.M.; Meng, Y.; Lin, S.; Zhang, Z.W. Effects of exogenous 24-epibrassinolide to control grey mould and maintain postharvest quality of table grapes. Int. J. Food Sci. Technol. 2016, 51, 1236–1243. [Google Scholar] [CrossRef]
- Song, L.X.; Xu, X.C.; Wang, F.N.; Wang, Y.; Xia, X.J.; Shi, K.; Zhou, Y.H.; Zhou, J.; Yu, J.Q. Brassinosteroids act as a positive regulator for resistance against root-knot nematode involving RESPIRATORY BURST OXIDASE HOMOLOG-dependent activation of MAPKs in tomato. Plant Cell Environ. 2018, 41, 1113–1125. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.F.; Jiang, T.L.; Xiang, Y.; He, X.W.; Zhang, Z.; Wen, S.J.; Zhang, J.J. Epi-brassinolide positively affects chlorophyll content and dark-reaction enzymes of maize seedlings. Phyton 2021, 90, 1465–1476. [Google Scholar] [CrossRef]
- Guo, J.K.; Zhou, R.; Ren, X.H.; Jia, H.L.; Hua, L.; Xu, H.H.; Lv, X.; Zhao, J.; Wei, T. Effects of salicylic acid, Epi-brassinolide and calcium on stress alleviation and Cd accumulation in tomato plants. Ecotox. Environ. Saf. 2018, 15, 491–496. [Google Scholar] [CrossRef] [PubMed]
- Pacheco, J.H.L.; Gonsebatt, M.E. The role of antioxidants and antioxidant-related enzymes in protective responses to environmentally induced oxidative stress. Mutat. Res. 2009, 674, 137–147. [Google Scholar] [CrossRef] [PubMed]
- Moghaddam, M.G.F. Seed germination, antioxidant enzymes activity and proline content in medicinal plant tagetes minuta under salinity stress. Plant Biosyst. 2020, 154, 835–842. [Google Scholar] [CrossRef]
- Kumutha, D.; Ezhilmathi, K.; Sairam, R.K.; Srivastava, G.C.; Deshmukh, P.S.; Meena, R. Waterlogging induced oxidative stress and antioxidant activity in pigeonpea genotypes. Biol. Plantarum. 2009, 53, 75–84. [Google Scholar] [CrossRef]
- Sairam, R.K.; Deshmukh, P.S.; Shukla, D.S. Tolerance of drought and temperature stress in relation to increased antioxidant enzyme activity in wheat. J. Agron. Crop Sci. 2008, 178, 171–178. [Google Scholar] [CrossRef]
- Zhang, D.W.; Deng, X.G.; Fu, F.Q.; Lin, H.H. Induction of plant virus defense response by brassinosteroids and brassinosteroid signaling in Arabidopsis thaliana. Planta 2015, 241, 875–885. [Google Scholar] [CrossRef]
- You, J.; Chan, Z.L. ROS regulation during abiotic stress responses in crop plants. Front. Plant Sci. 2015, 6, 690–695. [Google Scholar] [CrossRef]
- Dwivedi, S.K.; Basu, S.; Kumar, S.; Kumari, S.; Kumar, A.; Jha, S.; Mishra, J.S.; Bhatt, B.P.; Kumar, G. Enhanced antioxidant enzyme activities in developing anther contributes to heat stress alleviation and sustains grain yield in wheat. Funct. Plant Biol. 2019, 46, 1090–1102. [Google Scholar] [CrossRef] [PubMed]
- Dinakar, C.; Abhaypratap, V.; Yearla, S.R.; Raghavendra, A.S.; Padmasree, K. Importance of ROS and antioxidant system during the beneficial interactions of mitochondrial metabolism with photosynthetic carbon assimilation. Planta 2010, 231, 461–474. [Google Scholar] [CrossRef] [PubMed]
- Wei, H.Y.; Yang, M.; Ke, Y.G.; Liu, J.N.; Chen, Z.B.; Zhao, J.R.; Zhao, Y.T.; Huang, F.Y.; Yu, L. Comparative physiological and transcriptomic profiles reveal regulatory mechanisms of soft rot disease resistance in Amorphophallus spp. Physiol. Mol. Plant Pathol. 2022, 118, 101807. [Google Scholar] [CrossRef]
- Du, H.S.; Wang, Y.Q.; Yang, J.J.; Yang, W.C. Comparative transcriptome analysis of resistant and susceptible tomato lines in response to infection by xanthomonas perforans race T3. Front. Plant Sci. 2015, 6, 161–171. [Google Scholar] [CrossRef] [PubMed]
- Lantican, D.V.; Nocum, J.D.L.; Manohar, A.N.C.; Mendoza, J.V.S.; Gardoce, R.R.; Lachica, G.C.; Gueco, L.S.; Cueva, F.M.D. Comparative RNA-seq analysis of resistant and susceptible banana genotypes reveals molecular mechanisms in response to banana bunchy top virus(BBTV) infection. Sci. Rep. 2023, 13, 18719. [Google Scholar] [CrossRef]
- Ali, M.A.; Ren, H.Y.; Ahmed, T.; Luo, J.Y.; An, Q.L.; Qi, X.J.; Li, B. Antifungal effects of rhizospheric Bacillus species against bayberry twig blight pathogen pestalotiopsis versicolor. Agronomy 2022, 10, 1811. [Google Scholar] [CrossRef]
- Ahmed, T.; Ren, H.R.; Noman, M.; Shahid, M.; Liu, M.J.; Ali, M.A.; Zhang, J.N.; Tian, Y.; Qi, X.J.; Li, B. Green synthesis and characterization of zirconium oxide nanoparticles by using a native Enterobacter sp and its antifungal activity against bayberry twig blight disease pathogen Pestalotiopsis versicolor. NanoImpact 2021, 21, 100281. [Google Scholar] [CrossRef]










Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, Z.; Zhang, S.; Sun, L.; Liang, S.; Zheng, X.; Ren, H.; Qi, X. Effects of Enhanced Resistance and Transcriptome Analysis of Twig Blight Disease by Exogenous Brassinolide in Myrica rubra. Antioxidants 2024, 13, 61. https://doi.org/10.3390/antiox13010061
Yu Z, Zhang S, Sun L, Liang S, Zheng X, Ren H, Qi X. Effects of Enhanced Resistance and Transcriptome Analysis of Twig Blight Disease by Exogenous Brassinolide in Myrica rubra. Antioxidants. 2024; 13(1):61. https://doi.org/10.3390/antiox13010061
Chicago/Turabian StyleYu, Zheping, Shuwen Zhang, Li Sun, Senmiao Liang, Xiliang Zheng, Haiying Ren, and Xingjiang Qi. 2024. "Effects of Enhanced Resistance and Transcriptome Analysis of Twig Blight Disease by Exogenous Brassinolide in Myrica rubra" Antioxidants 13, no. 1: 61. https://doi.org/10.3390/antiox13010061