
Repurposing Glutathione Transferases: Directed Evolution Combined with Chemical Modification for the Creation of a Semisynthetic Enzyme with High Hydroperoxidase Activity

Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Methods
2.2.1. DNA Shuffling and Construction of GmGSTUs Library
2.2.2. Expression and Purification of Recombinant Enzymes
2.2.3. Assay of Enzyme Activity and Kinetic Analysis
2.2.4. Site-Directed Mutagenesis
2.2.5. Chemical Modification for the Creation of Selenium-Containing Enzymes
3. Results and Discussion
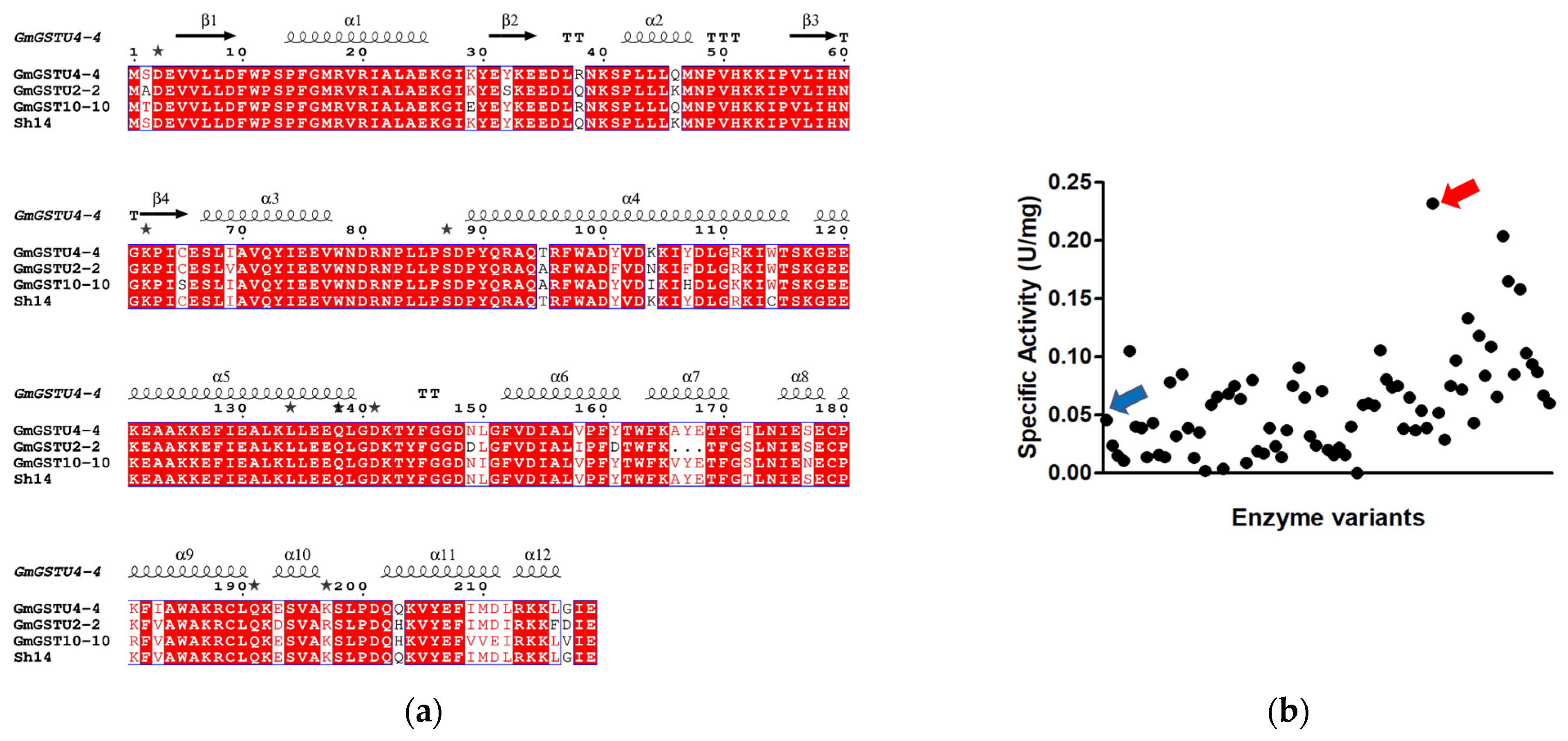
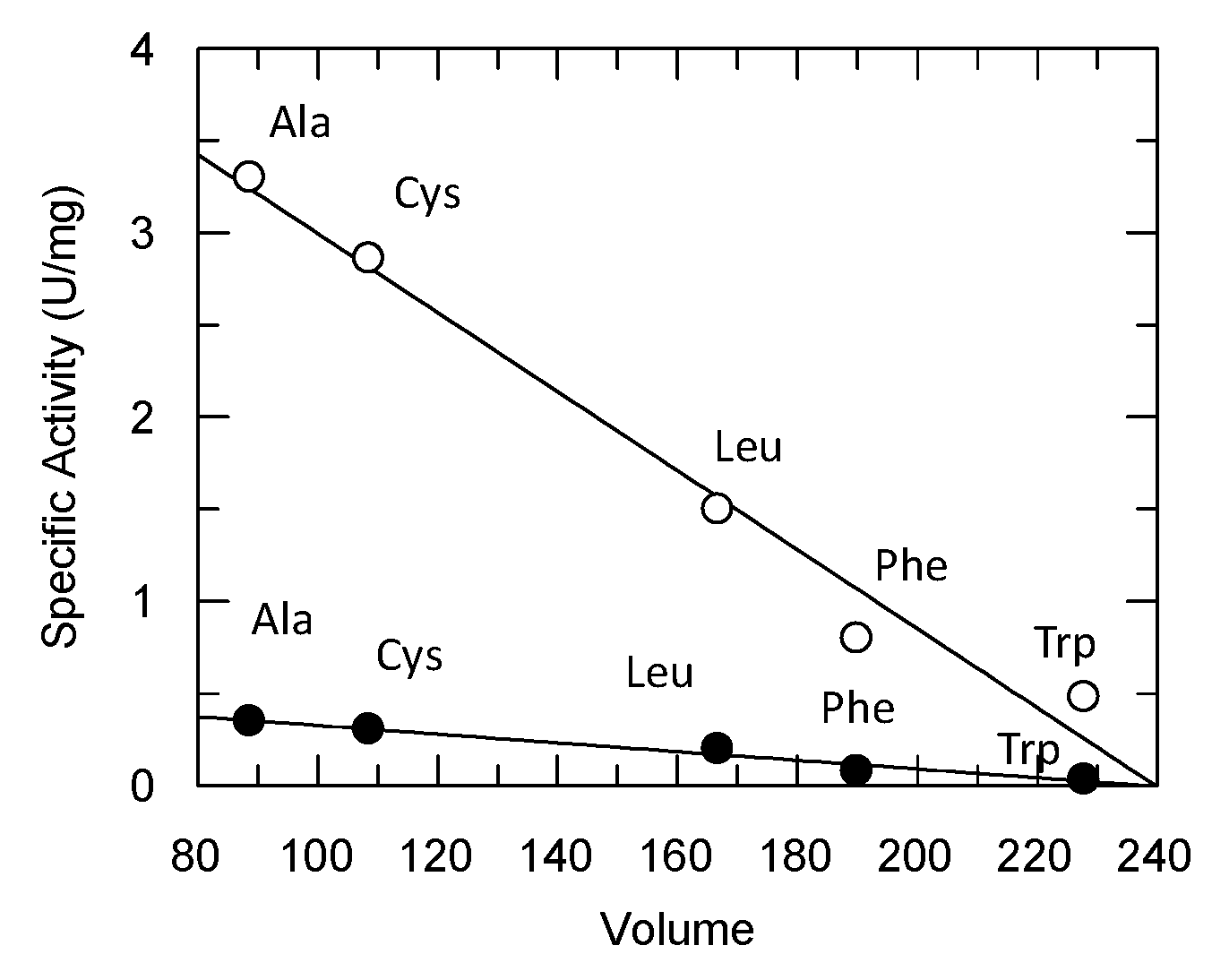
3.1. Directed Evolution of Three Homologous GmGSTUs for the Isolation of an Enzyme Variant with High Hydroperoxidase Activity
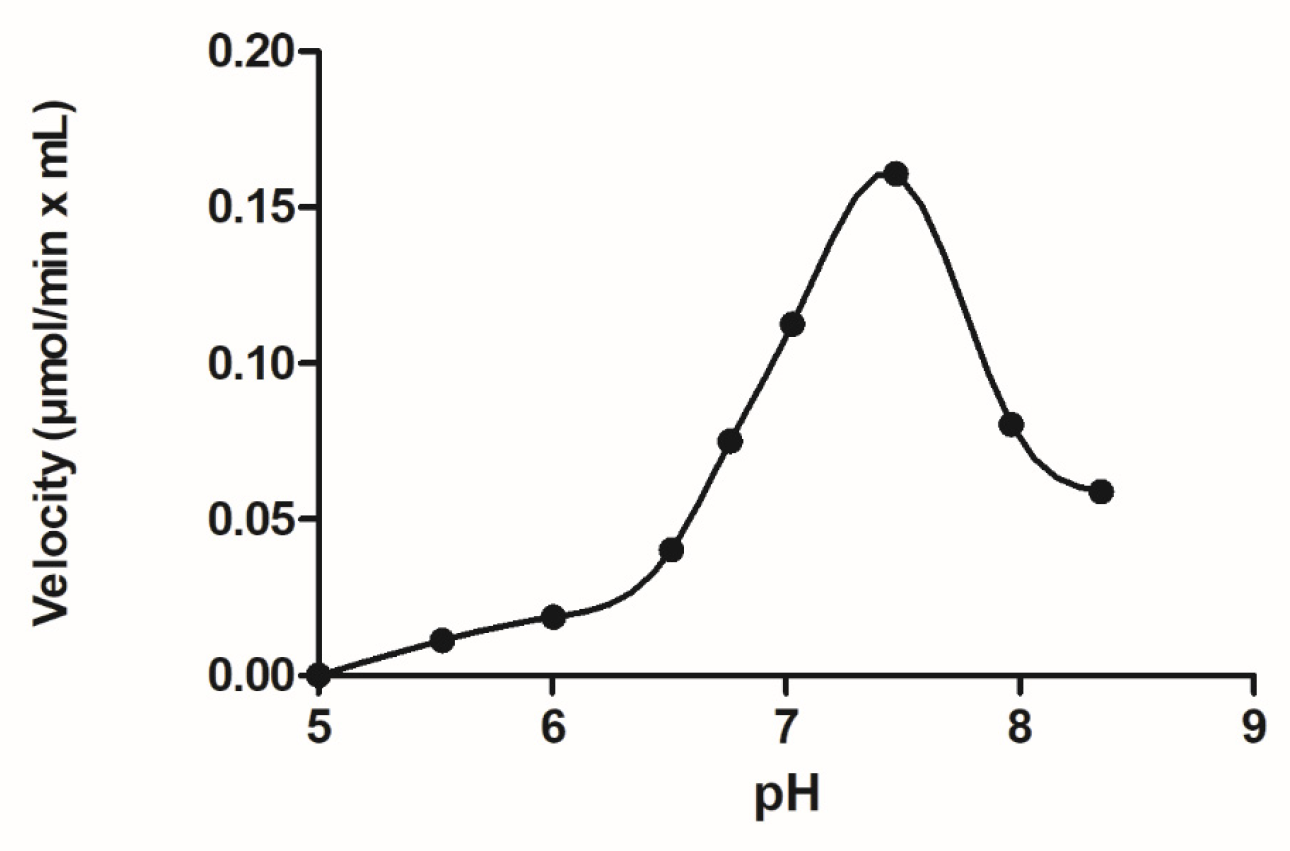
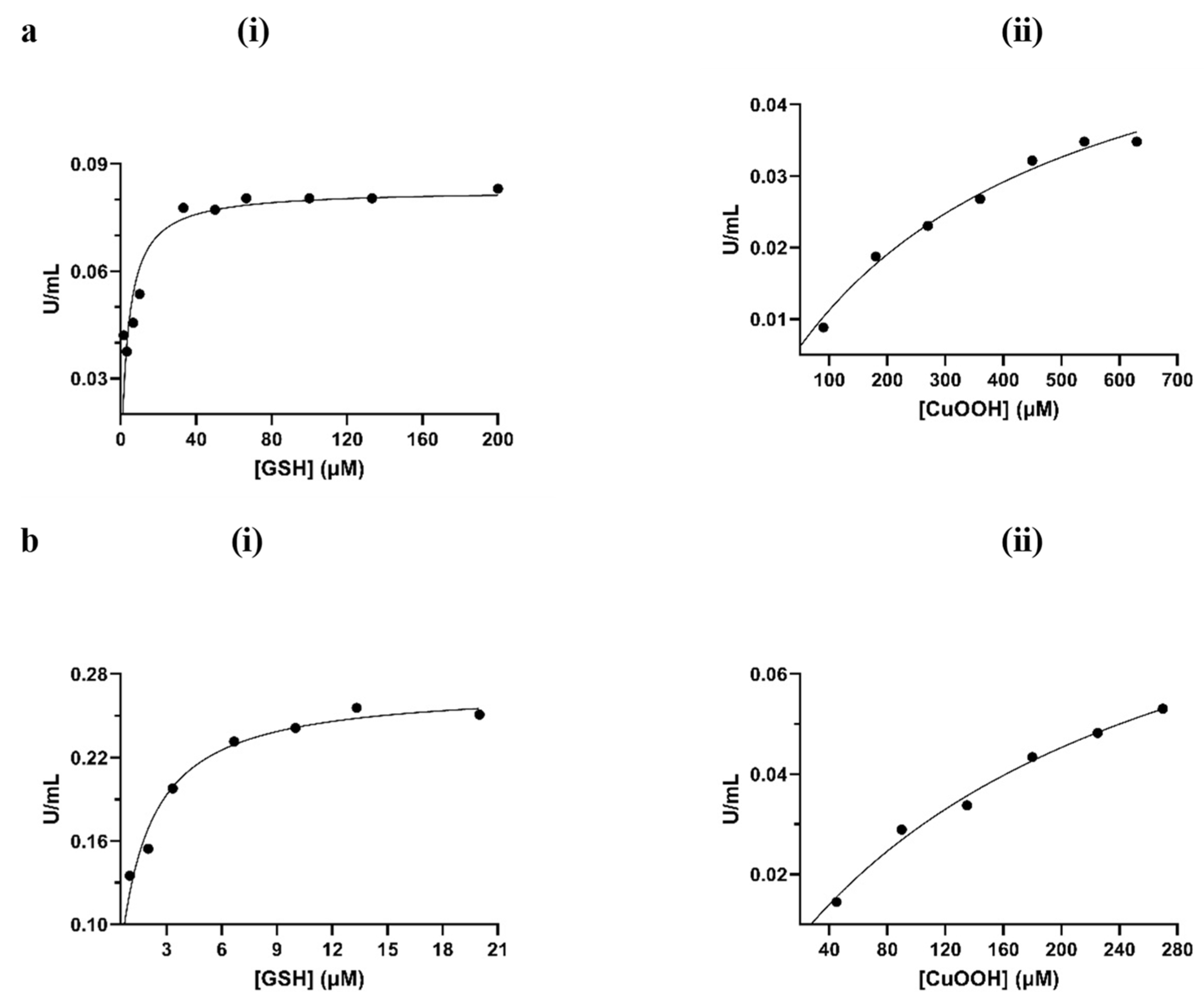
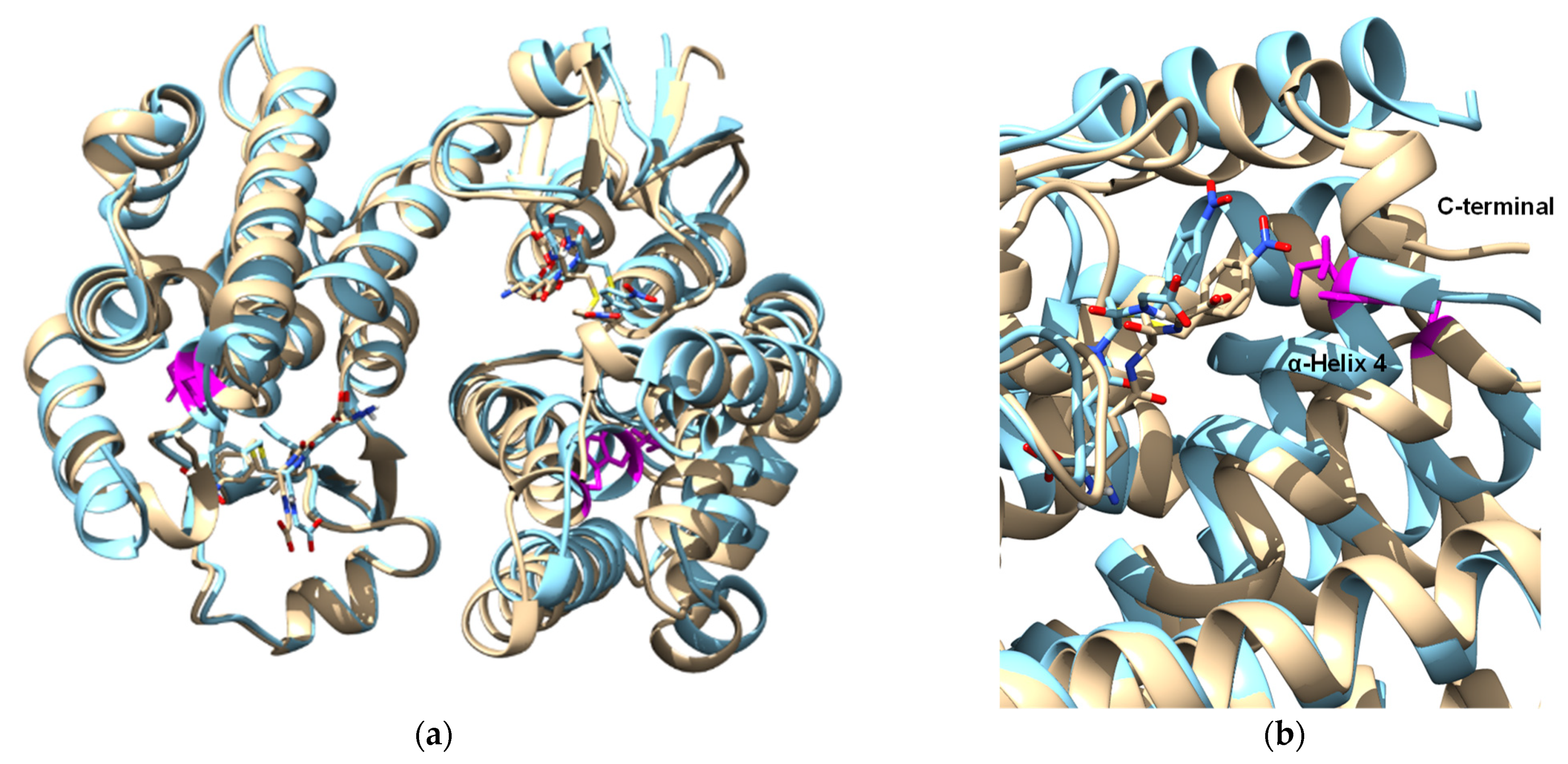
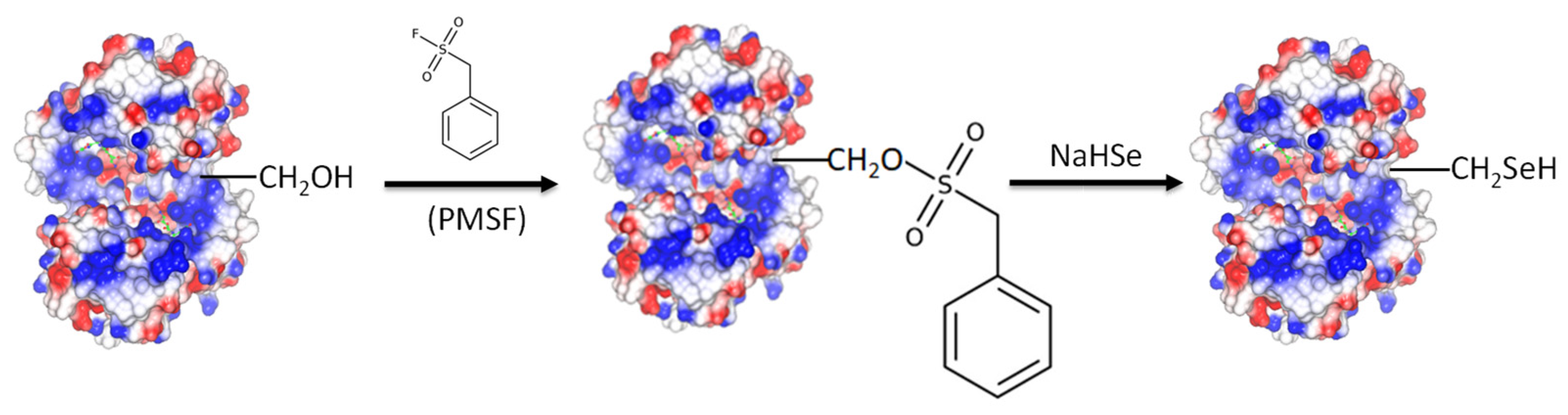
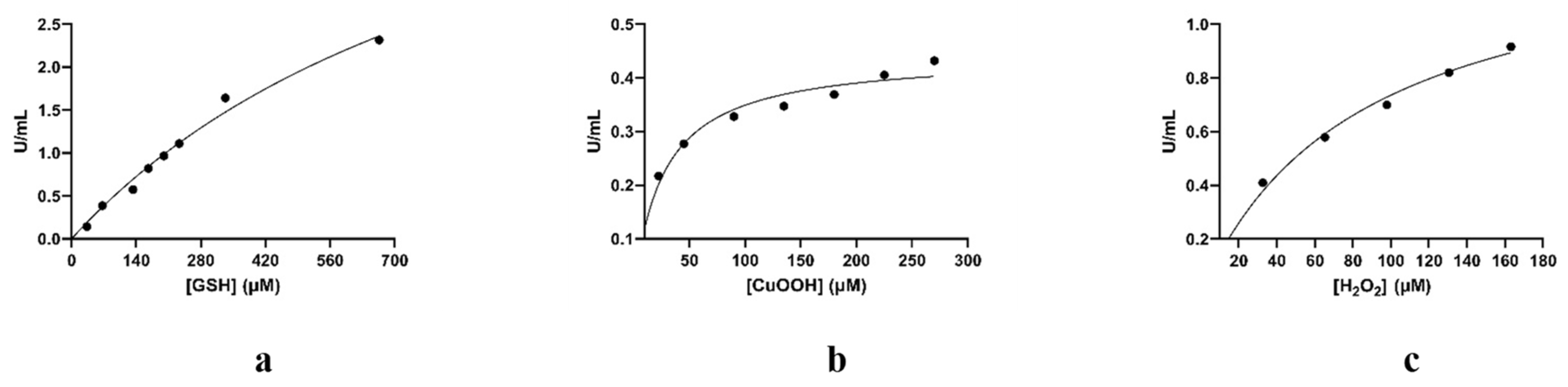
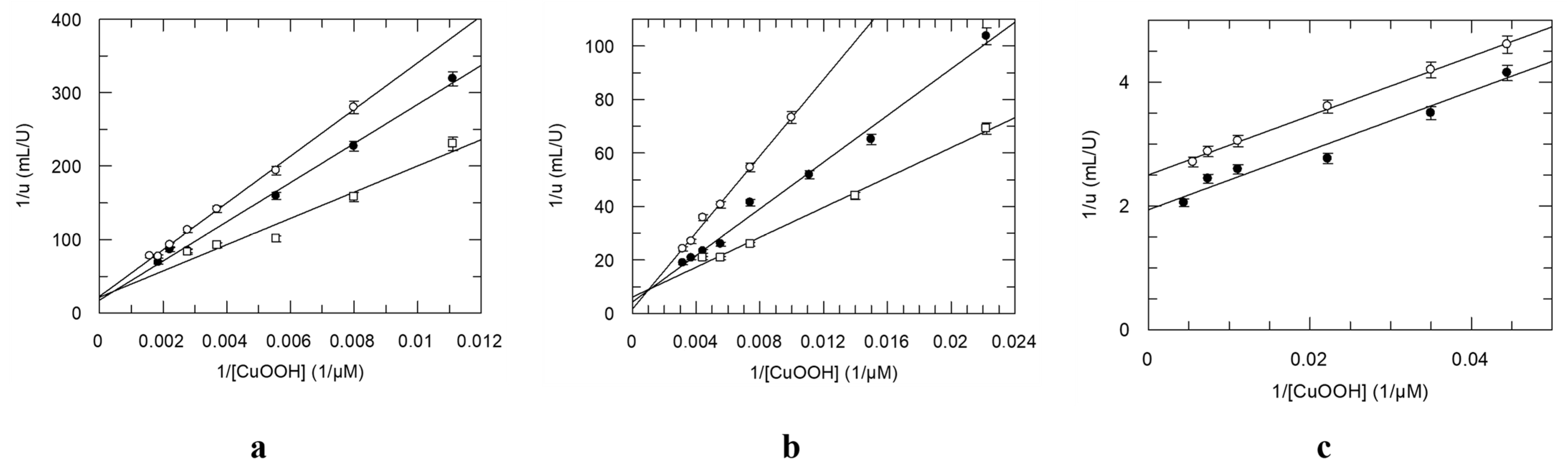
3.2. Chemical Modification for the Creation of Selenium-Containing Enzymes
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dogaru, C.B.; Muscurel, C.; Duță, C.; Stoian, I. “Alphabet” Selenoproteins: Their Characteristics and Physiological Roles. Int. J. Mol. Sci. 2023, 24, 15992. [Google Scholar] [CrossRef] [PubMed]
- Averill-Bates, D. Reactive oxygen species and cell signaling. Review. Biochim. Biophys. Acta Mol. Cell Res. 2023, 2023, 119573. [Google Scholar] [CrossRef] [PubMed]
- Lapenna, D. Glutathione and Glutathione-dependent Enzymes: From Biochemistry to Gerontology and Successful Aging. Ageing Res. Rev. 2023, 92, 102066. [Google Scholar] [CrossRef] [PubMed]
- Herbette, S.; Roeckel-Drevet, P.; Drevet, J.R. Seleno-independent glutathione peroxidases: More than simple antioxidant scavengers. FEBS J. 2007, 274, 2163–2180. [Google Scholar] [CrossRef] [PubMed]
- Chaudière, J. Biological and Catalytic Properties of Selenoproteins. Int. J. Mol. Sci. 2023, 24, 10109. [Google Scholar] [CrossRef] [PubMed]
- Genchi, G.; Lauria, G.; Catalano, A.; Sinicropi, M.S.; Carocci, A. Biological Activity of Selenium and Its Impact on Human Health. Int. J. Mol. Sci. 2023, 24, 2633. [Google Scholar] [CrossRef] [PubMed]
- Ren, X.; Jemth, P.; Board, P.G.; Luo, G.; Mannervik, B.; Liu, J.; Zhang, K.; Shen, J. A semisynthetic glutathione peroxidase with high catalytic efficiency: Selenoglutathione transferase. Chem. Biol. 2002, 9, 789–794. [Google Scholar] [CrossRef] [PubMed]
- Zheng, K.; Board, P.G.; Fei, X.; Sun, Y.; Lv, S.; Yan, G.; Liu, J.; Shen, J.; Luo, G. A novel selenium-containing glutathione transferase zeta1-1, the activity of which surpasses the level of some native glutathione peroxidases. Int. J. Biochem. Cell Biol. 2008, 40, 2090–2097. [Google Scholar] [CrossRef]
- Yu, H.-J.; Liu, J.-Q.; Böck, A.; Li, J.; Luo, G.-M.; Shen, J.-C. Engineering glutathione transferase to a novel glutathione peroxidase mimic with high catalytic efficiency: Incorporation of selenocysteine into a glutathione-binding scaffold using an auxotrophic expression system. J. Biol. Chem. 2005, 280, 11930–11935. [Google Scholar] [CrossRef]
- Yin, L.; Song, J.; Board, P.G.; Yu, Y.; Han, X.; Wei, J. Characterization of selenium-containing glutathione transferase zeta1-1 with high GPX activity prepared in eukaryotic cells. J. Mol. Recognit. 2013, 26, 38–45. [Google Scholar] [CrossRef]
- Mazari, A.M.A.; Zhang, L.; Ye, Z.-W.; Zhang, J.; Tew, K.D.; Townsend, D.M. The Multifaceted Role of Glutathione S-Transferases in Health and Disease. Biomolecules 2023, 13, 688. [Google Scholar] [CrossRef] [PubMed]
- Perperopoulou, F.; Pouliou, F.; Labrou, N.E. Recent advances in protein engineering and biotechnological applications of glutathione transferases. Crit. Rev. Biotechnol. 2018, 38, 511–528. [Google Scholar] [CrossRef] [PubMed]
- Atkinson, H.J.; Babbitt, P.C. Glutathione transferases are structural and functional outliers in the thioredoxin fold. Biochemistry 2009, 48, 11108–11116. [Google Scholar] [CrossRef] [PubMed]
- Axarli, I.; Muleta, A.W.; Vlachakis, D.; Kossida, S.; Kotzia, G.; Maltezos, A.; Dhavala, P.; Papageorgiou, A.C.; Labrou, N.E. Directed evolution of Tau class glutathione transferases reveals a site that regulates catalytic efficiency and masks co-operativity. Biochem. J. 2016, 473, 559–570. [Google Scholar] [CrossRef] [PubMed]
- Axarli, I.; Dhavala, P.; Papageorgiou, A.C.; Labrou, N.E. Crystallographic and functional characterization of the fluorodifen-inducible glutathione transferase from glycine max reveals an active site topography suited for diphenylether herbicides and a novel L-site. J. Mol. Biol. 2009, 385, 984–1002. [Google Scholar] [CrossRef] [PubMed]
- Cummins, I.; Dixon, D.P.; Freitag-Pohl, S.; Skipsey, M.; Edwards, R. Multiple roles for plant glutathione transferases in xenobiotic detoxification. Drug Metab. Rev. 2011, 43, 266–280. [Google Scholar] [CrossRef]
- Mannervik, B. Five Decades with Glutathione and the GSTome. J. Biol. Chem. 2012, 287, 6072–6083. [Google Scholar] [CrossRef]
- Deponte, M. Glutathione catalysis and the reaction mechanisms of glutathione-dependent enzymes. Biochim. Biophys. Acta 2013, 1830, 3217–3266. [Google Scholar] [CrossRef]
- Mannervik, B. Glutathione peroxidase. Methods Enzymol. 1985, 113, 490–495. [Google Scholar]
- Robertson, G.J.; Stoychev, S.H.; Sayed, Y.; Achilonu, I.; Dirr, H.W. The effects of mutating Tyr9 and Arg15 on the structure, stability, conformational dynamics and mechanism of GSTA3-3. Biophys. Chem. 2017, 224, 40–48. [Google Scholar] [CrossRef]
- Pégeot, H.; Mathiot, S.; Perrot, T.; Gense, F.; Hecker, A.; Didierjean, C.; Rouhier, N. Structural plasticity among glutathione transferase Phi members: Natural combination of catalytic residues confers dual biochemical activities. FEBS J. 2017, 284, 2442–2463. [Google Scholar] [CrossRef] [PubMed]
- Osman, W.H.W.; Mikami, B.; Saka, N.; Kondo, K.; Nagata, T.; Katahira, M. Structure of a serine-type glutathione S-transferase of Ceriporiopsis subvermispora and identification of the enzymatically important non-canonical residues by functional mutagenesis. Biochem. Biophys. Res. Commun. 2019, 510, 177–183. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Wu, Z.; Gao, J. Effects of conserved Arg20, Glu74 and Asp77 on the structure and function of a tau class glutathione S-transferase in rice. Plant Mol. Biol. 2021, 105, 451–462. [Google Scholar] [CrossRef] [PubMed]
- Blikstad, C.; Shokeer, A.; Kurtovic, S.; Mannervik, B. Emergence of a novel highly specific and catalytically efficient enzyme from a naturally promiscuous glutathione transferase. Biochim. Biophys. Acta 2008, 1780, 1458–1463. [Google Scholar] [CrossRef]
- Runarsdottir, A.; Mannervik, B. A novel quasi-species of glutathione transferase with high activity towards naturally occurring isothiocyanates evolves from promiscuous low-activity variants. J. Mol. Biol. 2010, 401, 451–464. [Google Scholar] [CrossRef] [PubMed]
- Aravilli, R.K.; Kohila, V.; Vikram, S.L. Heuristics in Role of Human Glutathione S-transferase Mu 1 as Nitric Oxide Carrier and its Engineered Variants for Enhanced Activity. Curr. Pharm. Biotechnol. 2021, 22, 2071–2084. [Google Scholar] [CrossRef] [PubMed]
- Musdal, Y.; Govindarajan, S.; Mannervik, B. Exploring sequence-function space of a poplar glutathione transferase using designed information-rich gene variants. Protein Eng. Des. Sel. 2017, 30, 543–549. [Google Scholar] [CrossRef]
- Zhang, W.; Dourado, D.F.; Mannervik, B. Evolution of the active site of human glutathione transferase A2-2 for enhanced activity with dietary isothiocyanates. Biochim. Biophys. Acta 2015, 1850, 742–749. [Google Scholar] [CrossRef]
- Kurtovic, S.; Mannervik, B. Identification of emerging quasi-species in directed enzyme evolution. Biochemistry 2009, 48, 9330–9339. [Google Scholar] [CrossRef]
- Mannervik, B.; Runarsdottir, A.; Kurtovic, S. Multi-substrate–activity space and quasi-species in enzyme evolution: Ohno’s dilemma, promiscuity and functional orthogonality. Biochem. Soc. Trans. 2009, 37 Pt 4, 740–744. [Google Scholar] [CrossRef]
- Govindarajan, S.; Mannervik, B.; Silverman, J.A.; Wright, K.; Regitsky, D.; Hegazy, U.; Purcell, T.J.; Welch, M.; Minshull, J.; Gustafsson, C. Mapping of amino acid substitutions conferring herbicide resistance in wheat glutathione transferase. ACS Synth. Biol. 2015, 4, 221–227. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Dourado, D.F.A.R.; Fernandes, P.A.; Ramos, M.J.; Mannervik, B. Multidimensional epistasis and fitness landscapes in enzyme evolution. Biochem. J. 2012, 445, 39–46. [Google Scholar] [CrossRef]
- Sylvestre-Gonon, E.; Law, S.R.; Schwartz, M.; Robe, K.; Keech, O.; Didierjean, C.; Dubos, C.; Rouhier, N.; Hecker, A. Functional, Structural and Biochemical Features of Plant Serinyl-Glutathione Transferases. Front. Plant Sci. 2019, 10, 608. [Google Scholar] [CrossRef] [PubMed]
- Skopelitou, K.; Muleta, A.W.; Papageorgiou, A.C.; Chronopoulou, E.G.; Pavli, O.; Flemetakis, E.; Skaracis, G.N.; Labrou, N.E. Characterization and functional analysis of a recombinant tau class glutathione transferase GmGSTU2-2 from Glycine max. Int. J. Biol. Macromol. 2017, 94 Pt B, 802–812. [Google Scholar] [CrossRef] [PubMed]
- Skopelitou, K.; Muleta, A.W.; Papageorgiou, A.C.; Chronopoulou, E.; Labrou, N.E. Catalytic features and crystal structure of a tau class glutathione transferase from Glycine max specifically upregulated in response to soybean mosaic virus infections. Biochim. Biophys. Acta 2015, 1854, 166–177. [Google Scholar] [CrossRef] [PubMed]
- Axarli, I.; Georgiadou, C.; Dhavala, P.; Papageorgiou, A.C.; Labrou, N.E. Investigation of the role of conserved residues Ser13, Asn48 and Pro49 in the catalytic mechanism of the tau class glutathione transferase from Glycine max. Biochim. Biophys. Acta 2010, 1804, 662–667. [Google Scholar] [CrossRef] [PubMed]
- Klayman, D.L.; Griffin, T.S. Reaction of selenium with sodium borohydride in protic solvents: A facile method for the introduction of selenium into organic molecules. J. Am. Chem. Soc. 1973, 95, 197–199. [Google Scholar] [CrossRef]
- E Labrou, N.; Rigden, D.J.; Clonis, Y.D. Engineering the pH-dependence of kinetic parameters of maize glutathione S-transferase I by site-directed mutagenesis. Biomol. Eng. 2004, 21, 61–66. [Google Scholar] [CrossRef]
- Yamamoto, Y.; Takahashi, K. Glutathione peroxidase isolated from plasma reduces phospholipid hydroperoxides. Arch. Biochem. Biophys. 1993, 305, 541–545. [Google Scholar] [CrossRef]
- Robert, X.; Gouet, P. Deciphering key features in protein structures with the new ENDscript server. Nucleic Acids Res. 2014, 42, W320–W324. [Google Scholar] [CrossRef]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera?A visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef] [PubMed]
- Flohe, L. The selenoprotein glutathione peroxidase. In Glutathione: Chemical, Biochemical and Medical Aspects: Part A; Dolphin, D., Poulson, R., Avramovic, O., Eds.; Wiley Interscience: New York, NY, USA, 1989; pp. 643–731. [Google Scholar]
- Axarli, I.; Dhavala, P.; Papageorgiou, A.C.; Labrou, N.E. Crystal structure of Glycine max glutathione transferase in complex with glutathione: Investigation of the mechanism operating by the Tau class glutathione transferases. Biochem. J. 2009, 422, 247–256. [Google Scholar] [CrossRef] [PubMed]
- Jakobson, I.; Warholm, M.; Mannervik, B. The binding of substrates and a product of the enzymatic reaction to glutathione S-transferase A. J. Biol. Chem. 1979, 254, 7085–7089. [Google Scholar] [CrossRef] [PubMed]
- Jakobson, I.; Warholm, M.; Askelof, P.; Mannervik, B. A steady-state-kinetic random mechanism for glutathione S-transferase a from rat liver: A model involving kinetically significant enzyme-product complexes in the forward reaction. JBIC J. Biol. Inorg. Chem. 1977, 77, 253–262. [Google Scholar] [CrossRef]
- Yu, H.; Liu, J.; Liu, X.; Zang, T.; Luo, G.; Shen, J. Kinetic studies on the glutathione peroxidase activity of selenium-containing glutathione transferase. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2005, 141, 382–389. [Google Scholar] [CrossRef]
- Pannala, V.R.; Bazil, J.N.; Camara, A.K.S.; Dash, R.K. A mechanistic mathematical model for the catalytic action ofglutathione peroxidase. Free Radic. Res. 2014, 48, 487–502. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Enzyme | kcat (min−1) (CuOOH) | kcat (min−1) (H2O2) | Κm (μΜ) GSH | Κm (μΜ) CuOOH | Κm (μΜ) H2O2 | kcat/Κm (×10−3) (μΜ−1·min−1) (CuOOH) | kcat/Κm (×10−3) (μΜ−1·min−1) (H2O2) |
|---|---|---|---|---|---|---|---|
| GmGSTU4-4 | 10.7 ± 1.0 | - | 3.6 ± 0.8 | 454.3 ± 83.6 | - | 23.6 | - |
| Sh14 | 35.6 ± 3.7 | - | 1.2 ± 0.1 | 255.8 ± 46.4 | - | 139.2 | - |
| SeSh14 | 1898.7 ± 79.9 | 5659.4 ± 442.3 | 999.3 ± 232.1 | 29.3 ± 5.3 | 83.8 ± 14.7 | 64,802 | 67,510 |
| Enzyme | Substrate System | Specific Activity (μmol·min−1·mg−1) |
|---|---|---|
| GmGSTU4-4 | CDNB/GSH | 11.2 |
| CuOOH/GSH | 0.5 | |
| H2O2/GSH | ND 1 | |
| SeGmGSTU4-4 | CDNB/GSH | ND |
| CuOOH/GSH | 90.0 | |
| H2O2/GSH | 67.5 | |
| Sh14 | CDNB/GSH | 48.4 |
| CuOOH/GSH | 2.9 | |
| H2O2/GSH | ND | |
| SeSh14 | CDNB/GSH | ND |
| CuOOH/GSH | 349.2 | |
| H2O2/GSH | 244.5 | |
| Ser13Ala | CDNB/GSH | 0.04 |
| CuOOH/GSH | ND | |
| H2O2/GSH | ND | |
| SeSer13Ala | CDNB/GSH | ND |
| CuOOH/GSH | 0.3 | |
| H2O2/GSH | 0.7 | |
| SerGST T2-2 (seleno rat GSTT2-2) | CuOOH | 23.00 2 |
| H2O2 | 102.00 2 | |
| GPx (human erythrocytes) | H2O2 | 100.00 3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Axarli, I.; Ataya, F.; Labrou, N.E. Repurposing Glutathione Transferases: Directed Evolution Combined with Chemical Modification for the Creation of a Semisynthetic Enzyme with High Hydroperoxidase Activity. Antioxidants 2024, 13, 41. https://doi.org/10.3390/antiox13010041
Axarli I, Ataya F, Labrou NE. Repurposing Glutathione Transferases: Directed Evolution Combined with Chemical Modification for the Creation of a Semisynthetic Enzyme with High Hydroperoxidase Activity. Antioxidants. 2024; 13(1):41. https://doi.org/10.3390/antiox13010041
Chicago/Turabian StyleAxarli, Irene, Farid Ataya, and Nikolaos E. Labrou. 2024. "Repurposing Glutathione Transferases: Directed Evolution Combined with Chemical Modification for the Creation of a Semisynthetic Enzyme with High Hydroperoxidase Activity" Antioxidants 13, no. 1: 41. https://doi.org/10.3390/antiox13010041