
Impaired NRF2 Inhibits Recovery from Ischemic Reperfusion Injury in the Aging Kidney
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Model
2.2. Measurement of Urinary Albumin Excretion
2.3. Determining Oxidative Stress
2.4. Histological Examination and Immunohistochemical Analysis
2.5. Isolation of Mitochondria
2.6. Measurement of ATP Content
2.7. Generation of Senescent Cells
2.8. Senescence-Associated β-Galactosidase (SA-β-gal) Staining
2.9. Cell Culture
2.10. Hypoxia/Reoxygenation Cell Model
2.11. Western Blot Analysis
2.12. Cell Viability Assay
2.13. Measurement of Reactive Oxygen Species (ROS)
2.14. Assessment of the Mitochondrial Membrane Potential (MMP)
2.15. Statistics
3. Results
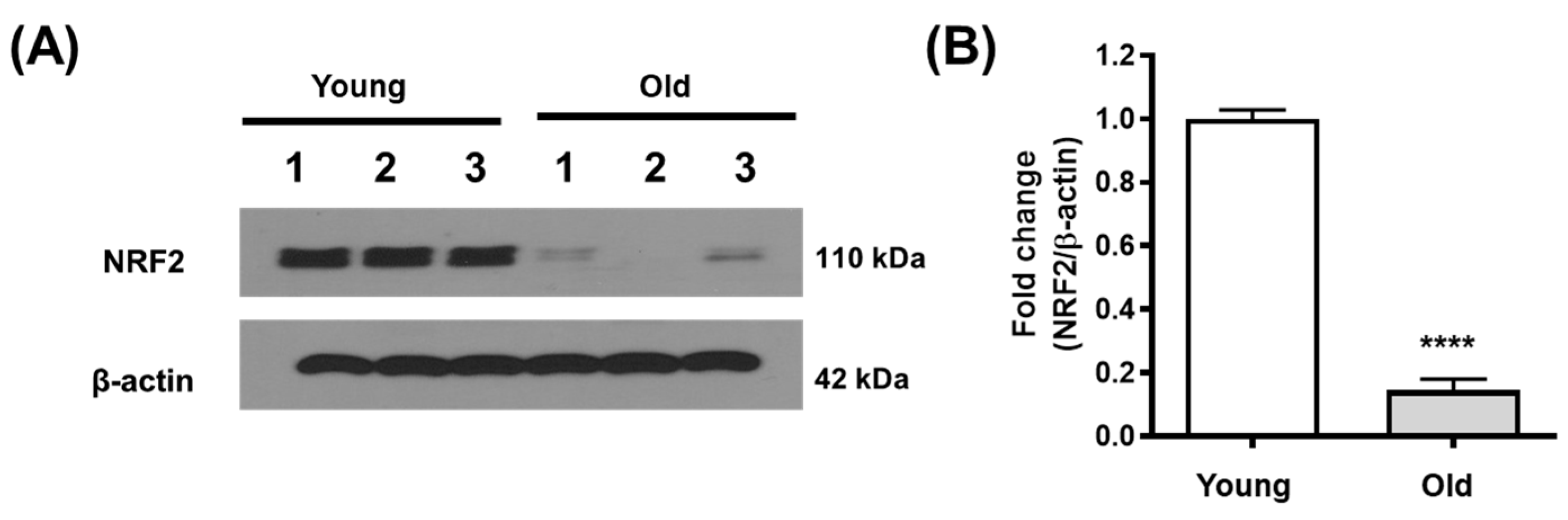
3.1. NRF2 Expression Decreases in the Aging Kidney
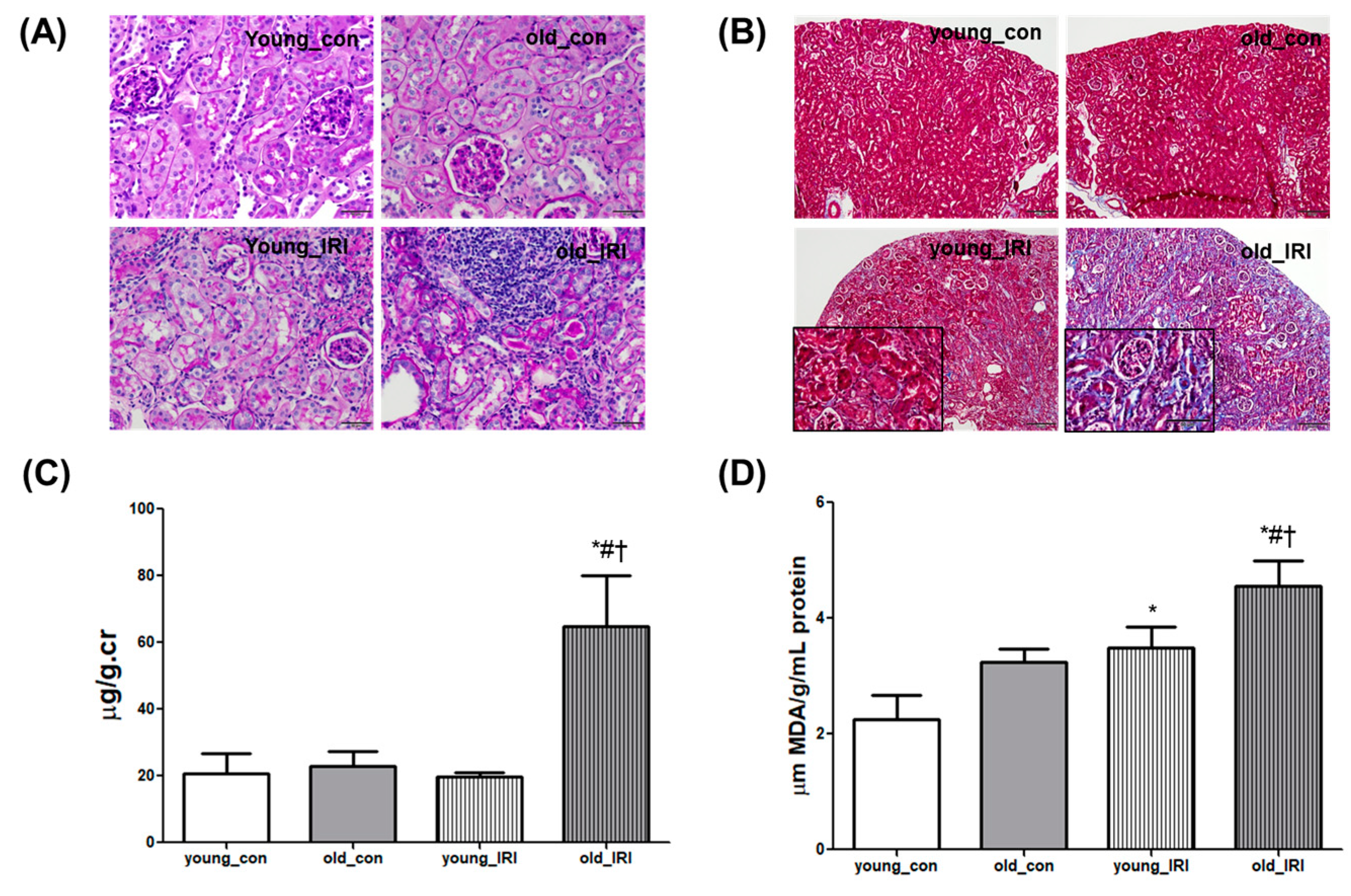
3.2. Delayed Renal Recovery after IRI in Old Mice
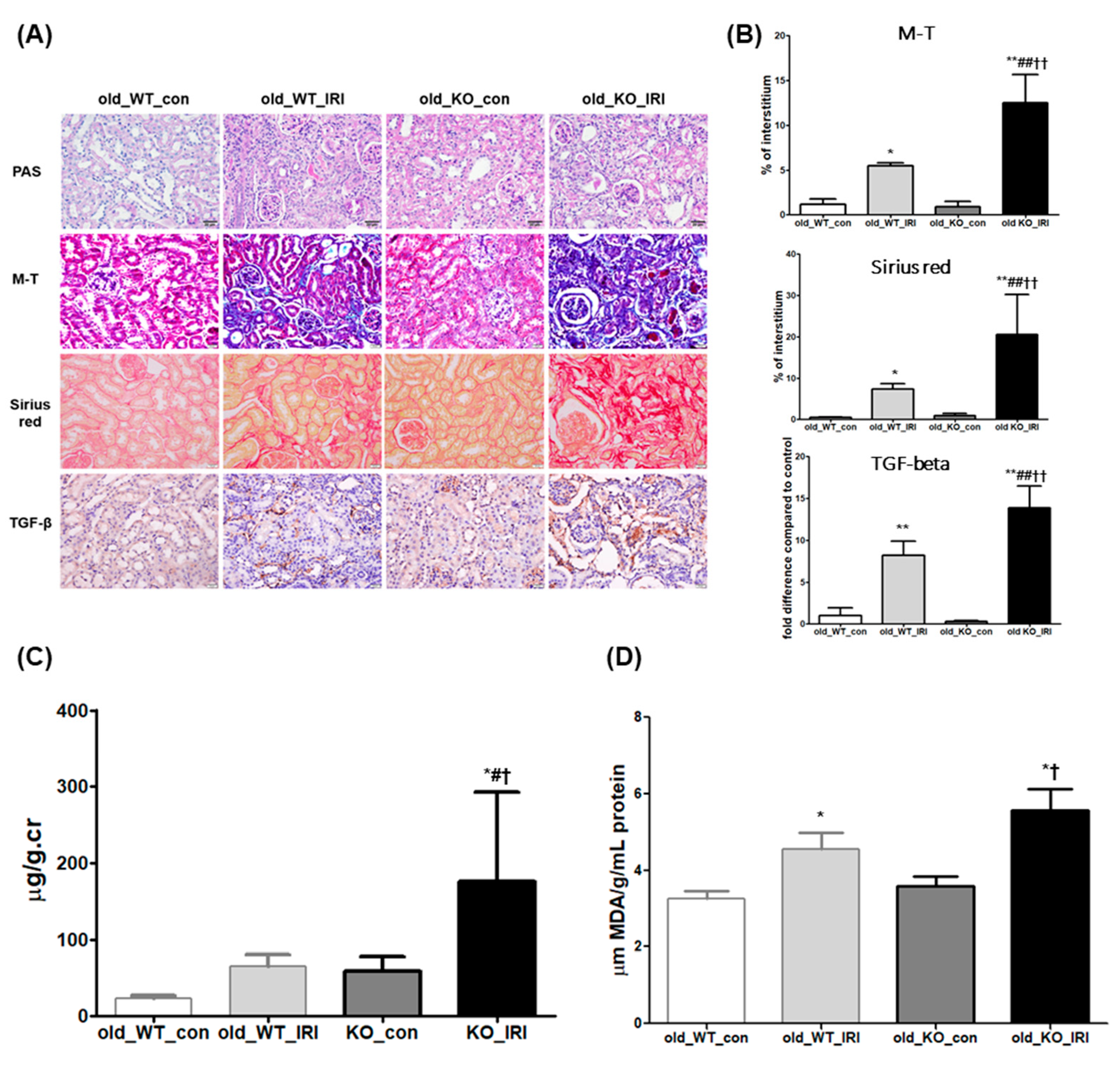
3.3. Exacerbation of Renal Dysfunction after IRI in NRF2 Knockout Mice
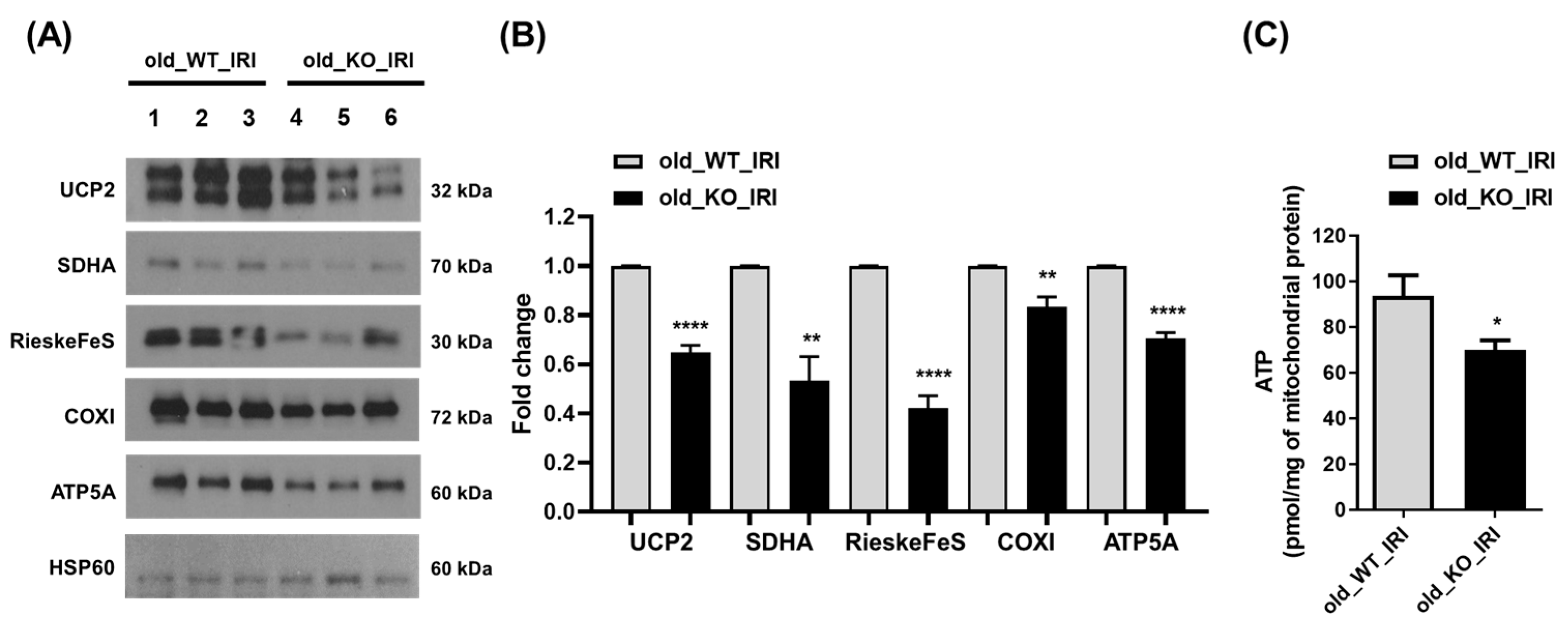
3.4. Reduced Expression of Mitochondrial Protein in NRF2 Knockout Mice
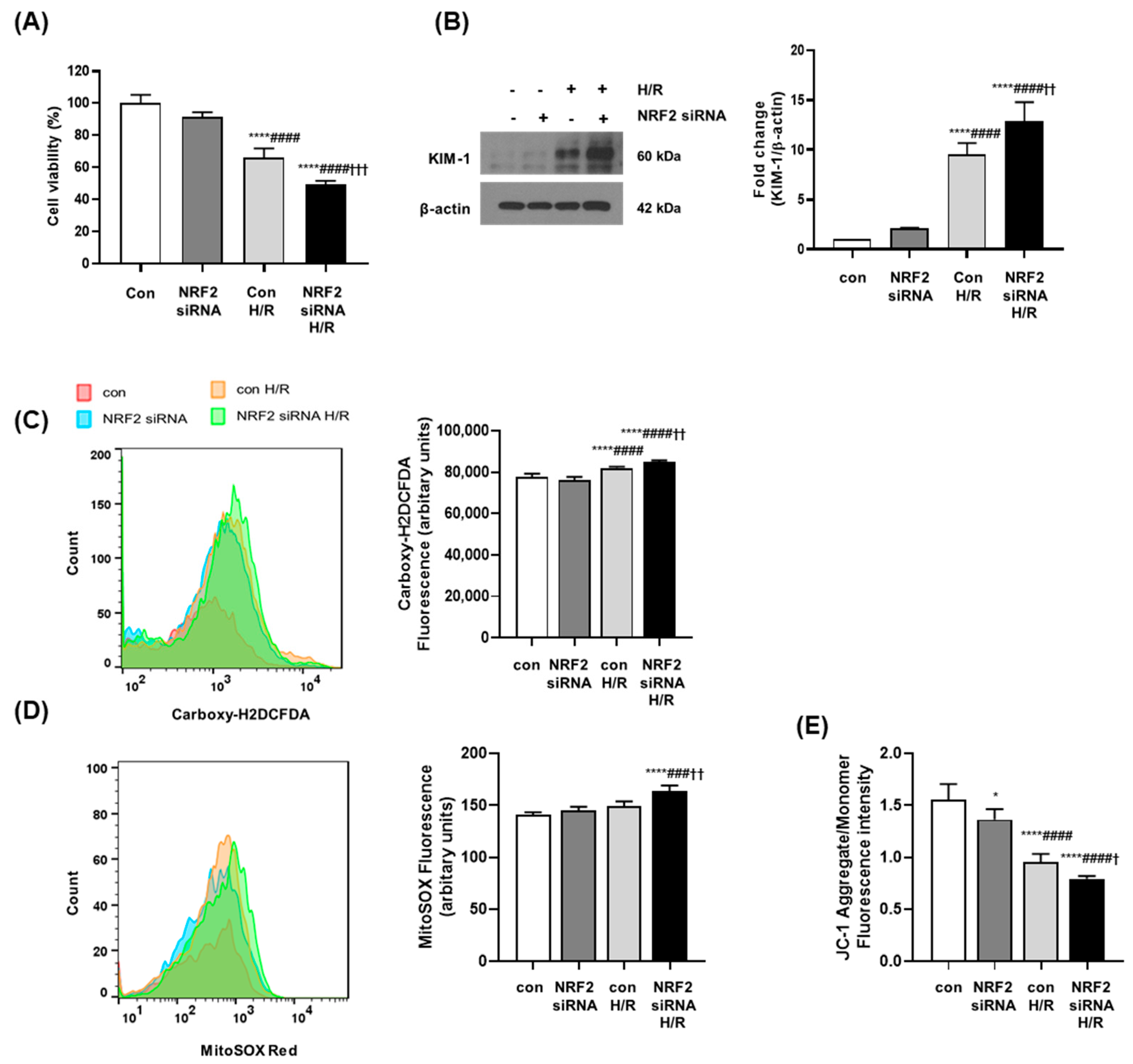
3.5. NRF2 Deficiency Increases Oxidative Stress and Causes Mitochondrial Dysfunction in Senescent Cells
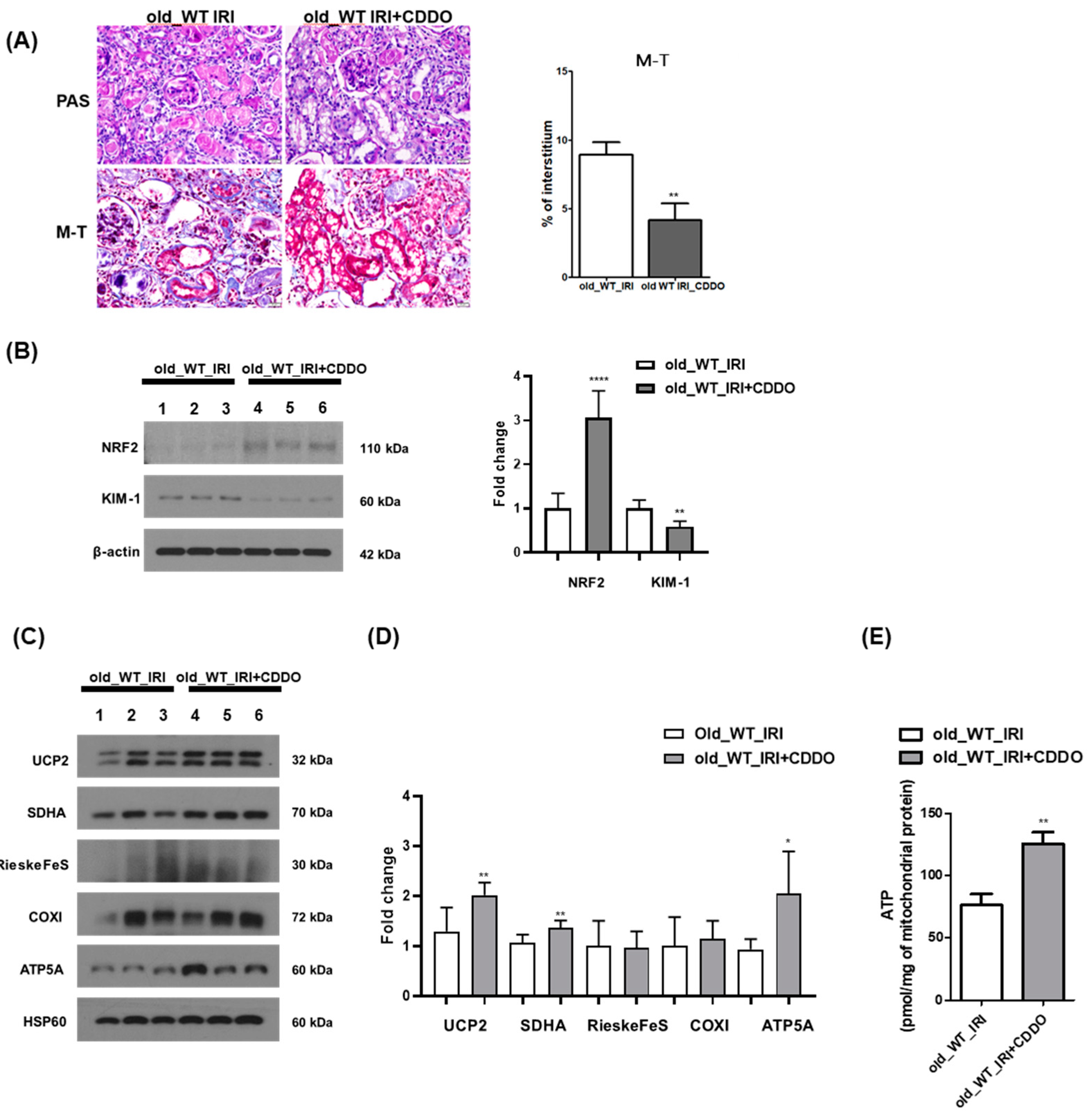
3.6. CDDO-Me Treatment Contributes to Renal Recovery
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bolignano, D.; Mattace-Raso, F.; Sijbrands, E.J.; Zoccali, C. The aging kidney revisited: A systematic review. Ageing Res. Rev. 2014, 14, 65–80. [Google Scholar] [CrossRef] [PubMed]
- Levin, A.; Stevens, P.E. Summary of Recommendation Statements. Kidney Int. Suppl. 2013, 3, 5–14. [Google Scholar]
- Bellomo, R.; Kellum, J.A.; Ronco, C. Acute Kidney Injury. Lancet 2012, 380, 756–766. [Google Scholar] [CrossRef] [PubMed]
- Lafrance, J.-P.; Miller, D.R. Acute Kidney Injury Associates with Increased Long-Term Mortality. J. Am. Soc. Nephrol. 2010, 21, 345–352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chawla, L.S.; Kimmel, P.L. Acute Kidney Injury and Chronic Kidney Disease: An Integrated Clinical Syndrome. Kidney Int. 2012, 82, 516–524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murtagh, F.E.; Burns, A.; Moranne, O.; Morton, R.L.; Naicker, S. Supportive Care: Comprehensive Conservative Care in End-Stage Kidney Disease. Clin. J. Am. Soc. Nephrol. 2016, 11, 1909–1914. [Google Scholar] [CrossRef] [Green Version]
- Chertow, G.M.; Burdick, E.; Honour, M.; Bonventre, J.V.; Bates, D.W. Acute Kidney Injury, Mortality, Length of Stay, and Costs in Hospitalized Patients. J. Am. Soc. Nephrol. 2005, 16, 3365–3370. [Google Scholar] [CrossRef] [Green Version]
- Chao, C.-T.; Tsai, H.-B.; Lin, Y.-F.; Ko, W.-J. Acute kidney injury in the elderly: Only the tip of the iceberg. J. Clin. Gerontol. Geriatr. 2014, 5, 7–12. [Google Scholar] [CrossRef] [Green Version]
- Yokota, L.G.; Sampaio, B.M.; Rocha, E.P.; Balbi, A.L.; Prado, I.R.S.; Ponce, D. Acute kidney injury in elderly patients: Narrative review on incidence, risk factors, and mortality. Int. J. Nephrol. Renov. Dis. 2018, 11, 217–224. [Google Scholar] [CrossRef] [Green Version]
- Coca, S.G. Acute Kidney Injury in Elderly Persons. Am. J. Kidney Dis. 2010, 56, 122–131. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Bonventre, J.V.; Parrish, A.R. The Aging Kidney: Increased Susceptibility to Nephrotoxicity. Int. J. Mol. Sci. 2014, 15, 15358–15376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmitt, R.; Coca, S.; Kanbay, M.; Tinetti, M.E.; Cantley, L.G.; Parikh, C.R. Recovery of Kidney Function After Acute Kidney Injury in the Elderly: A Systematic Review and Meta-analysis. Am. J. Kidney Dis. 2008, 52, 262–271. [Google Scholar] [CrossRef]
- He, F.; Ru, X.; Wen, T. NRF2, a Transcription Factor for Stress Response and Beyond. Int. J. Mol. Sci. 2020, 21, 4777. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Jia, Z.; Zhu, H. Regulation of Nrf2 Signaling. React. Oxyg. Species 2019, 8, 312–322. [Google Scholar] [CrossRef]
- Kaspar, J.W.; Niture, S.K.; Jaiswal, A.K. Nrf2:INrf2 (Keap1) signaling in oxidative stress. Free. Radic. Biol. Med. 2009, 47, 1304–1309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keum, Y.-S.; Choi, B.Y. Molecular and Chemical Regulation of the Keap1-Nrf2 Signaling Pathway. Molecules 2014, 19, 10074–10089. [Google Scholar] [CrossRef] [Green Version]
- Osburn, W.O.; Kensler, T.W. Nrf2 signaling: An adaptive response pathway for protection against environmental toxic insults. Mutat. Res. Mol. Mech. Mutagen. 2008, 659, 31–39. [Google Scholar] [CrossRef] [Green Version]
- Nezu, M.; Suzuki, N.; Yamamoto, M. Targeting the KEAP1-NRF2 System to Prevent Kidney Disease Progression. Am. J. Nephrol. 2017, 45, 473–483. [Google Scholar] [CrossRef]
- Yamamoto, M.; Kensler, T.W.; Motohashi, H. The KEAP1-NRF2 System: A Thiol-Based Sensor-Effector Apparatus for Maintaining Redox Homeostasis. Physiol. Rev. 2018, 98, 1169–1203. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.C.; Fogo, A.B. Fibrosis and Renal Aging. Kidney Int. Suppl. 2014, 4, 75–78. [Google Scholar] [CrossRef] [Green Version]
- Muroya, Y.; He, X.; Fan, L.; Wang, S.; Xu, R.; Fan, F.; Roman, R.J. Enhanced renal ischemia-reperfusion injury in aging and diabetes. Am. J. Physiol.-Ren. Physiol. 2018, 315, F1843–F1854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.; Ko, Y.S.; Lee, H.Y.; Yang, J.; Oh, S.; Jo, S.-K.; Cho, W.; Kim, M.-G. The role of senescence of bone marrow cells in acute kidney injury. Kidney Res. Clin. Pr. 2019, 38, 25–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amdur, R.L.; Chawla, L.S.; Amodeo, S.; Kimmel, P.L.; Palant, C.E. Outcomes following diagnosis of acute renal failure in U.S. veterans: Focus on acute tubular necrosis. Kidney Int. 2009, 76, 1089–1097. [Google Scholar] [CrossRef]
- Shelton, L.M.; Park, B.K.; Copple, I.M. Role of Nrf2 in protection against acute kidney injury. Kidney Int. 2013, 84, 1090–1095. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Davies, K.J.; Forman, H.J. Oxidative stress response and Nrf2 signaling in aging. Free. Radic. Biol. Med. 2015, 88, 314–336. [Google Scholar] [CrossRef] [Green Version]
- Schmidlin, C.J.; Dodson, M.B.; Madhavan, L.; Zhang, D.D. Redox Regulation by Nrf2 in Aging and Disease. Free Radic. Biol. Med. 2019, 134, 702–707. [Google Scholar] [CrossRef]
- Junqueira, V.B.; Barros, S.B.; Chan, S.S.; Rodrigues, L.; Giavarotti, L.; Abud, R.L.; Deucher, G.P. Aging and Oxidative Stress. Mol. Asp. Med. 2004, 25, 5–16. [Google Scholar] [CrossRef]
- Shih, P.-H.; Yen, G.-C. Differential expressions of antioxidant status in aging rats: The role of transcriptional factor Nrf2 and MAPK signaling pathway. Biogerontology 2006, 8, 71–80. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.-J.; Nel, A.E. The Role of Phase II Antioxidant Enzymes in Protecting Memory T Cells from Spontaneous Apoptosis in Young and Old Mice. J. Immunol. 2005, 175, 2948–2959. [Google Scholar] [CrossRef] [Green Version]
- George, M.; Tharakan, M.; Culberson, J.; Reddy, A.P.; Reddy, P.H. Role of Nrf2 in Aging, Alzheimer’s and Other Neurodegenerative Diseases. Ageing Res. Rev. 2022, 82, 101756. [Google Scholar] [CrossRef]
- Liu, Y.; Uruno, A.; Saito, R.; Matsukawa, N.; Hishinuma, E.; Saigusa, D.; Liu, H.; Yamamoto, M. Nrf2 deficiency deteriorates diabetic kidney disease in Akita model mice. Redox Biol. 2022, 58, 102525. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.; Murakami, S.; Matsumaru, D.; Kawauchi, T.; Nabeshima, Y.I.; Motohashi, H. Nrf2 Pathway Activation Attenuates Ageing-Related Renal Phenotypes due to Alpha-Klotho Deficiency. J. Biochem. 2022, 171, 579–589. [Google Scholar] [CrossRef] [PubMed]
- Aminzadeh, M.A.; Nicholas, S.B.; Norris, K.C.; Vaziri, N.D. Role of impaired Nrf2 activation in the pathogenesis of oxidative stress and inflammation in chronic tubulo-interstitial nephropathy. Nephrol. Dial. Transplant. 2013, 28, 2038–2045. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Wang, S.; Sun, Q.W.; Zhang, B.; Ullah, M.F.; Sun, Z. Klotho Deficiency Causes Heart Aging via Impairing the Nrf2-GR Pathway. Circ. Res. 2021, 128, 492–507. [Google Scholar] [CrossRef]
- Lee, S.M.; Koh, D.H.; Jun, D.W.; Roh, Y.J.; Kang, H.T.; Oh, J.H.; Kim, H.S. Auranofin Attenuates Hepatic Steatosis and Fibrosis in Nonalcoholic Fatty Liver Disease Via Nrf2 and Nf- Kappab Signaling Pathways. Clin. Mol. Hepatol. 2022, 28, 827–840. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.N.; Lim, J.H.; Kim, M.Y.; Ban, T.H.; Jang, I.A.; Yoon, H.E.; Park, C.W.; Chang, Y.S.; Choi, B.S. Resveratrol, an Nrf2 activator, ameliorates aging-related progressive renal injury. Aging 2018, 10, 83–99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nie, H.; Xue, X.; Liu, G.; Guan, G.; Liu, H.; Sun, L.; Zhao, L.; Wang, X.; Chen, Z. Nitro-oleic acid ameliorates oxygen and glucose deprivation/re-oxygenation triggered oxidative stress in renal tubular cells via activation of Nrf2 and suppression of NADPH oxidase. Free. Radic. Res. 2016, 50, 1200–1213. [Google Scholar] [CrossRef]
- Liu, M.; Reddy, N.M.; Higbee, E.M.; Potteti, H.R.; Noel, S.; Racusen, L.; Kensler, T.W.; Sporn, M.B.; Reddy, S.P.; Rabb, H. The Nrf2 Triterpenoid Activator, Cddo-Imidazolide, Protects Kidneys from Ischemia–Reperfusion Injury in Mice. Kidney Int. 2014, 85, 134–141. [Google Scholar] [CrossRef] [Green Version]
- Kocak, C.; Kocak, F.E.; Akcilar, R.; Bayat, Z.; Aras, B.; Metineren, M.H.; Yucel, M.; Simsek, H. Effects of Captopril, Telmisartan and Bardoxolone Methyl (Cddo-Me) in Ischemia-Reperfusion-Induced Acute Kidney Injury in Rats: An Experimental Comparative Study. Clin. Exp. Pharmacol. Physiol. 2016, 43, 230–241. [Google Scholar] [CrossRef] [PubMed]
- Reisman, S.A.; Goldsberry, A.R.; Lee, C.-Y.I.; O’grady, M.L.; Proksch, J.W.; Ward, K.W.; Meyer, C.J. Topical application of RTA 408 lotion activates Nrf2 in human skin and is well-tolerated by healthy human volunteers. BMC Dermatol. 2015, 15, 10. [Google Scholar] [CrossRef] [Green Version]
- Zahran, M.H.; Hussein, A.M.; Barakat, N.; Awadalla, A.; Khater, S.; Harraz, A.; Shokeir, A.A. Sildenafil activates antioxidant and antiapoptotic genes and inhibits proinflammatory cytokine genes in a rat model of renal ischemia/reperfusion injury. Int. Urol. Nephrol. 2015, 47, 1907–1915. [Google Scholar] [CrossRef] [PubMed]
- Wei, W.; Ma, N.; Fan, X.; Yu, Q.; Ci, X. The role of Nrf2 in acute kidney injury: Novel molecular mechanisms and therapeutic approaches. Free Radic. Biol. Med. 2020, 158, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Nezu, M.; Suzuki, N. Nrf2 Activation for Kidney Disease Treatment—A Mixed Blessing? Kidney Int. 2021, 99, 20–22. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.-Y.; Yang, Y.-X.; Zhe, H.; He, Z.-X.; Zhou, S.-F. Bardoxolone methyl (CDDO-Me) as a therapeutic agent: An update on its pharmacokinetic and pharmacodynamic properties. Drug Des. Dev. Ther. 2014, 8, 2075–2088. [Google Scholar] [CrossRef] [Green Version]
- De Zeeuw, D.; Akizawa, T.; Audhya, P.; Bakris, G.L.; Chin, M.; Christ-Schmidt, H.; Goldsberry, A.; Houser, M.; Krauth, M.; Lambers Heerspink, H.J. Bardoxolone Methyl in Type 2 Diabetes and Stage 4 Chronic Kidney Disease. N. Engl. J. Med. 2013, 369, 2492–2503. [Google Scholar] [CrossRef] [Green Version]
- Trifunovic, A.; Larsson, N.-G. Mitochondrial dysfunction as a cause of ageing. J. Intern. Med. 2008, 263, 167–178. [Google Scholar] [CrossRef]
- Jankauskas, S.S.; Silachev, D.N.; Andrianova, N.V.; Pevzner, I.B.; Zorova, L.D.; Popkov, V.A.; Plotnikov, E.Y.; Zorov, D.B. Aged kidney: Can we protect it? Autophagy, mitochondria and mechanisms of ischemic preconditioning. Cell Cycle 2018, 17, 1291–1309. [Google Scholar] [CrossRef] [Green Version]
- Jankauskas, S.S.; Pevzner, I.B.; Andrianova, N.V.; Zorova, L.D.; Popkov, V.A.; Silachev, D.N.; Kolosova, N.G.; Plotnikov, E.Y.; Zorov, D.B. The age-associated loss of ischemic preconditioning in the kidney is accompanied by mitochondrial dysfunction, increased protein acetylation and decreased autophagy. Sci. Rep. 2017, 7, srep44430. [Google Scholar] [CrossRef] [Green Version]
- Mohammad, R.S.; Lokhandwala, M.F.; Banday, A.A. Age-Related Mitochondrial Impairment and Renal Injury Is Ameliorated by Sulforaphane via Activation of Transcription Factor NRF2. Antioxidants 2022, 11, 156. [Google Scholar] [CrossRef]
- Csiszar, A.; Toth, J.; Peti-Peterdi, J.; Ungvari, Z. The aging kidney: Role of endothelial oxidative stress and inflammation. Acta Physiol. Hung. 2007, 94, 107–115. [Google Scholar] [CrossRef]
- Romano, A.D.; Serviddio, G.; de Matthaeis, A.; Bellanti, F.; Vendemiale, G. Oxidative Stress and Aging. J. Nephrol. 2010, 23, S29–S36. [Google Scholar]
- Bian, A.; Neyra, J.A.; Zhan, M.; Hu, M.C. Klotho, Stem Cells, and Aging. Clin. Interv. Aging 2015, 10, 1233. [Google Scholar] [PubMed] [Green Version]
- Kuro-o, M. Klotho and the Aging Process. Korean J. Intern. Med. 2011, 26, 113. [Google Scholar] [CrossRef] [PubMed]
- Nabeshima, Y.-I. Klotho: A fundamental regulator of aging. Ageing Res. Rev. 2002, 1, 627–638. [Google Scholar] [CrossRef] [PubMed]
- Briganti, S.; Flori, E.; Bellei, B.; Picardo, M. Modulation of PPARγ Provides New Insights in a Stress Induced Premature Senescence Model. PLoS ONE 2014, 9, e104045. [Google Scholar] [CrossRef] [Green Version]
- Sung, B.; Park, S.; Yu, B.P.; Chung, H.Y. Modulation of Ppar in Aging, Inflammation, and Calorie Restriction. J. Gerontol. Ser. A Biol. Sci. Med. Sci. 2004, 59, B997–B1006. [Google Scholar] [CrossRef] [Green Version]
- Chen, D.; Xie, R.; Shu, B.; Landay, A.L.; Wei, C.; Reiser, J.; Spagnoli, A.; Torquati, A.; Forsyth, C.B.; Keshavarzian, A.; et al. Wnt signaling in bone, kidney, intestine, and adipose tissue and interorgan interaction in aging. Ann. N. Y. Acad. Sci. 2019, 1442, 48–60. [Google Scholar] [CrossRef]
- Miao, J.; Liu, J.; Niu, J.; Zhang, Y.; Shen, W.; Luo, C.; Liu, Y.; Li, C.; Li, H.; Yang, P. Wnt/Β-Catenin/Ras Signaling Mediates Age-Related Renal Fibrosis and Is Associated with Mitochondrial Dysfunction. Aging Cell 2019, 18, e13004. [Google Scholar] [CrossRef] [Green Version]
- O’Sullivan, E.D.; Hughes, J.; Ferenbach, D.A. Renal Aging: Causes and Consequences. J. Am. Soc. Nephrol. 2017, 28, 407–420. [Google Scholar] [CrossRef] [Green Version]
- Gomes, P.; Simão, S.; Silva, E.; Pinto, V.; Amaral, J.S.; Afonso, J.; Serrão, M.P.; Pinho, M.J.; Soares-Da-Silva, P. Aging increases oxidative stress and renal expression of oxidant and antioxidant enzymes that are associated with an increased trend in systolic blood pressure. Oxid. Med. Cell. Longev. 2009, 2, 138–145. [Google Scholar] [CrossRef] [Green Version]
- Zuo, Z.; Lei, H.; Wang, X.; Wang, Y.; Sonntag, W.; Sun, Z. Aging-related kidney damage is associated with a decrease in klotho expression and an increase in superoxide production. Age 2011, 33, 261–274. [Google Scholar] [CrossRef] [PubMed] [Green Version]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jo, M.J.; Kim, J.E.; Bae, S.Y.; Cho, E.; Ahn, S.Y.; Kwon, Y.J.; Ko, G.-J. Impaired NRF2 Inhibits Recovery from Ischemic Reperfusion Injury in the Aging Kidney. Antioxidants 2023, 12, 1440. https://doi.org/10.3390/antiox12071440
Jo MJ, Kim JE, Bae SY, Cho E, Ahn SY, Kwon YJ, Ko G-J. Impaired NRF2 Inhibits Recovery from Ischemic Reperfusion Injury in the Aging Kidney. Antioxidants. 2023; 12(7):1440. https://doi.org/10.3390/antiox12071440
Chicago/Turabian StyleJo, Min Jee, Ji Eun Kim, So Yon Bae, Eunjung Cho, Shin Young Ahn, Young Joo Kwon, and Gang-Jee Ko. 2023. "Impaired NRF2 Inhibits Recovery from Ischemic Reperfusion Injury in the Aging Kidney" Antioxidants 12, no. 7: 1440. https://doi.org/10.3390/antiox12071440