
High Heme and Low Heme Oxygenase-1 Are Associated with Mast Cell Activation/Degranulation in HIV-Induced Chronic Widespread Pain
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
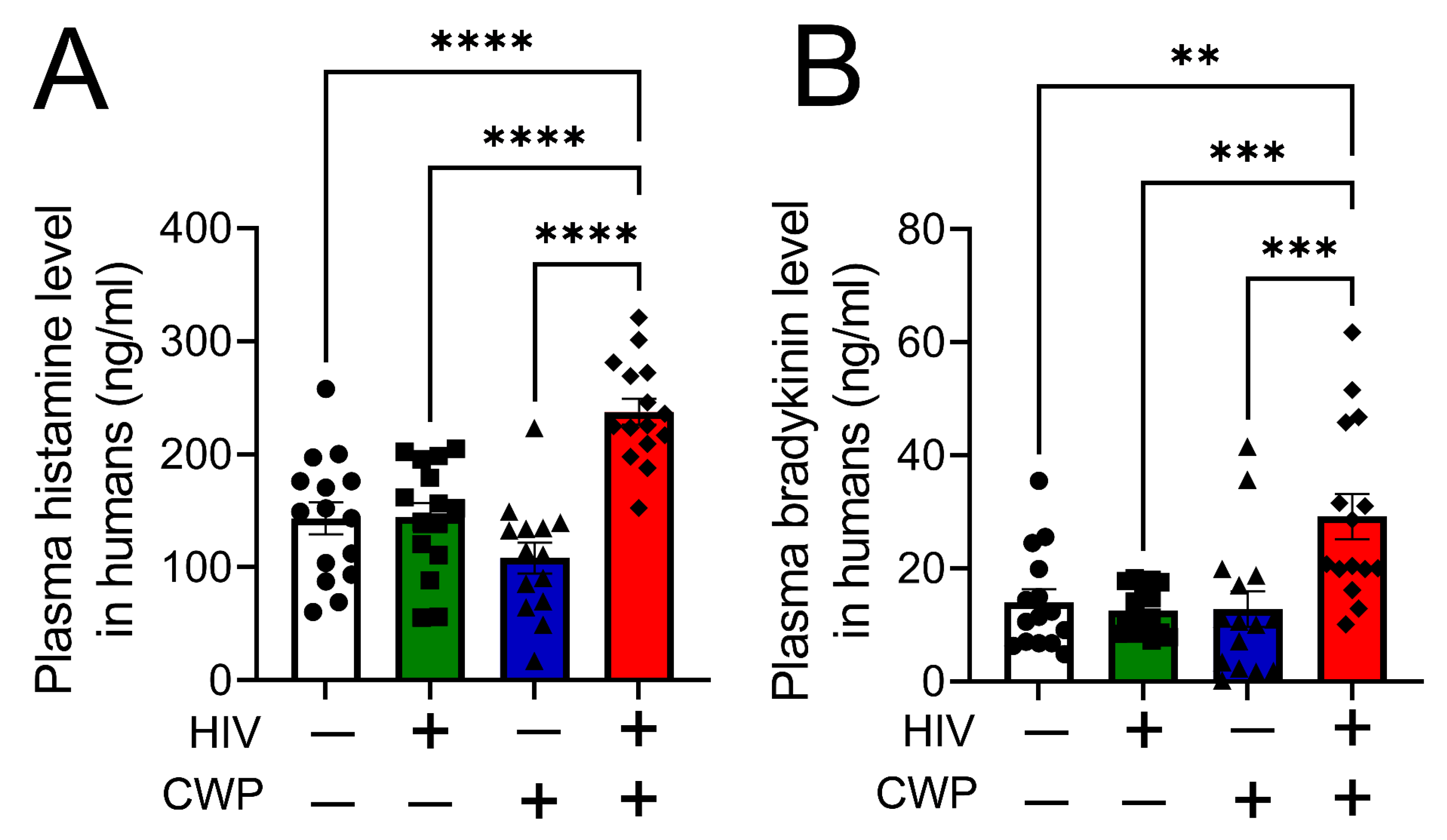
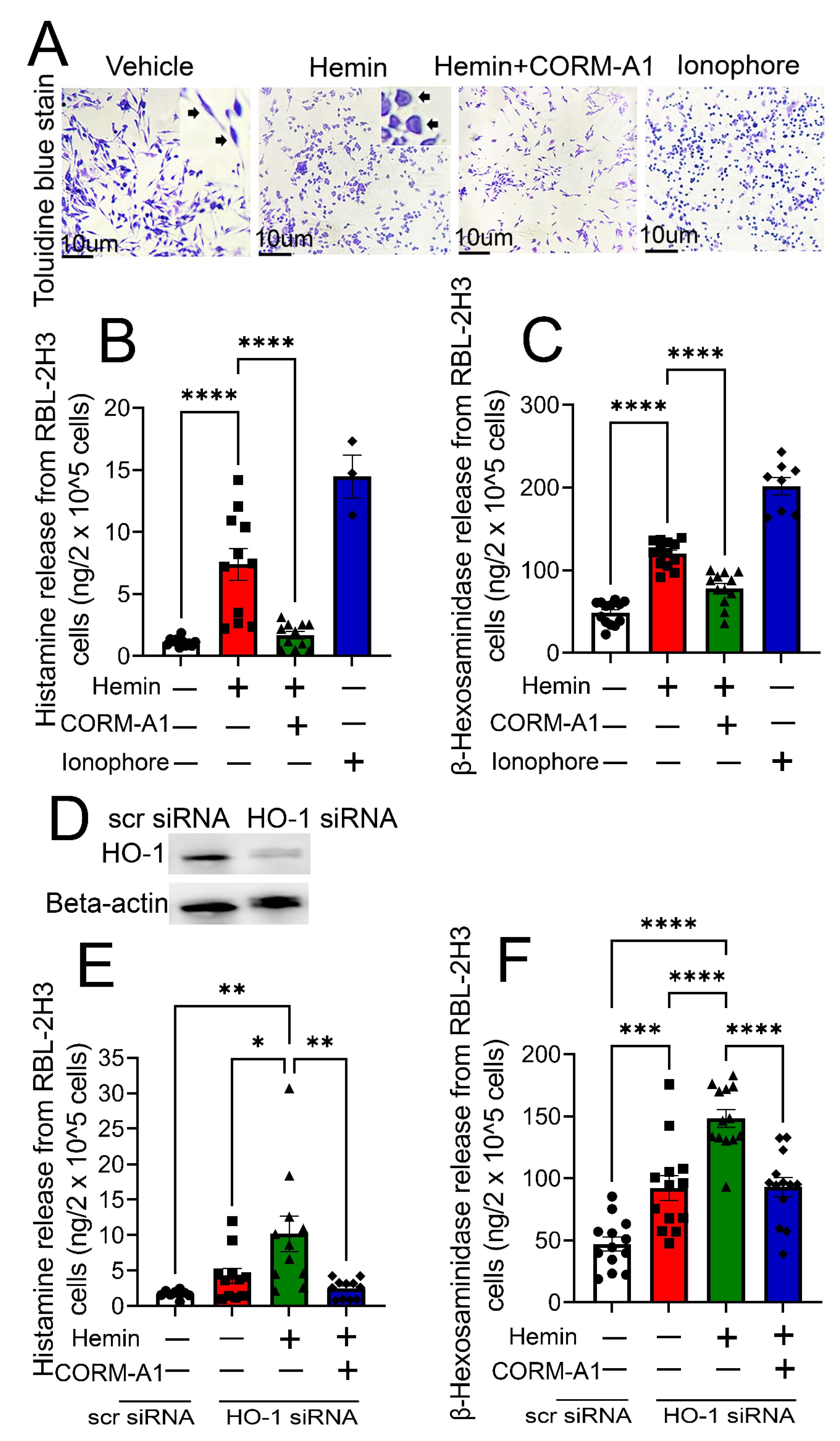
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Aggarwal, S.; DeBerry, J.J.; Ahmad, I.; Lynn, P.; Dewitte, C.; Malik, S.; Merlin, J.S.; Goodin, B.R.; Heath, S.L.; Matalon, S. Heme attenuates beta-endorphin levels in leukocytes of HIV positive individuals with chronic widespread pain. Redox Biol. 2020, 36, 101684. [Google Scholar] [CrossRef]
- Addis, D.R.; DeBerry, J.J.; Aggarwal, S. Chronic Pain in HIV. Mol. Pain 2020, 16, 1744806920927276. [Google Scholar] [CrossRef] [PubMed]
- Oguntibeju, O.O. Quality of life of people living with HIV and AIDS and antiretroviral therapy. HIV AIDS 2012, 4, 117–124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miaskowski, C.; Penko, J.M.; Guzman, D.; Mattson, J.E.; Bangsberg, D.R.; Kushel, M.B. Occurrence and characteristics of chronic pain in a community-based cohort of indigent adults living with HIV infection. J. Pain. 2011, 12, 1004–1016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Merlin, J.S.; Westfall, A.; Raper, J.L.; Zinski, A.; Norton, W.E.; Willig, J.H.; Gross, R.; Ritchie, C.S.; Saag, M.S.; Mugavero, M.J. Pain, mood, and substance abuse in HIV: Implications for clinic visit utilization, antiretroviral therapy adherence, and virologic failure. J. Acquir. Immune Defic. Syndr. 2012, 61, 164–170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Merlin, J.S.; Cen, L.; Praestgaard, A.; Turner, M.; Obando, A.; Alpert, C.; Woolston, S.; Casarett, D.; Kostman, J.; Gross, R.; et al. Pain and physical and psychological symptoms in ambulatory HIV patients in the current treatment era. J. Pain Symptom Manag. 2012, 43, 638–645. [Google Scholar] [CrossRef] [PubMed]
- Cunningham, C.O. Opioids and HIV Infection: From Pain Management to Addiction Treatment. Top. Antivir. Med. 2018, 25, 143–146. [Google Scholar]
- Kempuraj, D.; Mentor, S.; Thangavel, R.; Ahmed, M.E.; Selvakumar, G.P.; Raikwar, S.P.; Dubova, I.; Zaheer, S.; Iyer, S.S.; Zaheer, A. Mast Cells in Stress, Pain, Blood-Brain Barrier, Neuroinflammation and Alzheimer’s Disease. Front. Cell. Neurosci. 2019, 13, 54. [Google Scholar] [CrossRef] [Green Version]
- Conti, P.; D’ovidio, C.; Conti, C.; Gallenga, C.E.; Lauritano, D.; Caraffa, A.; Kritas, S.K.; Ronconi, G. Progression in migraine: Role of mast cells and pro-inflammatory and anti-inflammatory cytokines. Eur. J. Pharmacol. 2019, 844, 87–94. [Google Scholar] [CrossRef]
- Gordon, J.R.; Galli, S.J. Mast cells as a source of both preformed and immunologically inducible TNF-alpha/cachectin. Nature 1990, 346, 274–276. [Google Scholar] [CrossRef]
- Gupta, K.; Harvima, I.T. Mast cell-neural interactions contribute to pain and itch. Immunol. Rev. 2018, 282, 168–187. [Google Scholar] [CrossRef] [PubMed]
- Lei, J.; Paul, J.; Wang, Y.; Gupta, M.; Vang, D.; Thompson, S.; Jha, R.; Nguyen, J.; Valverde, Y.; Lamarre, Y.; et al. Heme Causes Pain in Sickle Mice via Toll-Like Receptor 4-Mediated Reactive Oxygen Species- and Endoplasmic Reticulum Stress-Induced Glial Activation. Antioxid. Redox Signal. 2021, 34, 279–293. [Google Scholar] [CrossRef] [PubMed]
- Hofman, Z.; de Maat, S.; Hack, C.E.; Maas, C. Bradykinin: Inflammatory Product of the Coagulation System. Clin. Rev. Allergy Immunol. 2016, 51, 152–161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tran, H.; Gupta, M.; Gupta, K. Targeting novel mechanisms of pain in sickle cell disease. Blood 2017, 130, 2377–2385. [Google Scholar] [CrossRef]
- Ren, K.; Torres, R. Role of interleukin-1beta during pain and inflammation. Brain Res. Rev. 2009, 60, 57–64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conti, P.; Pregliasco, F.E.; Bellomo, R.G.; Gallenga, C.E.; Caraffa, A.; Kritas, S.K.; Lauritano, D.; Ronconi, G. Mast Cell Cytokines IL-1, IL-33, and IL-36 Mediate Skin Inflammation in Psoriasis: A Novel Therapeutic Approach with the Anti-Inflammatory Cytokines IL-37, IL-38, and IL-1Ra. Int. J. Mol. Sci. 2021, 22, 8076. [Google Scholar] [CrossRef]
- Castany, S.; Carcolé, M.; Leánez, S.; Pol, O. The Induction of Heme Oxygenase 1 Decreases Painful Diabetic Neuropathy and Enhances the Antinociceptive Effects of Morphine in Diabetic Mice. PLoS ONE 2016, 11, e0146427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Merlin, J.S.; Westfall, A.O.; Heath, S.L.; Goodin, B.R.; Stewart, J.C.; Sorge, R.E.; Younger, J. Brief Report: IL-1β Levels Are Associated With Chronic Multisite Pain in People Living With HIV. J. Acquir. Immune Defic. Syndr. 2017, 75, e99–e103. [Google Scholar] [CrossRef]
- Takamiya, R.; Murakami, M.; Kajimura, M.; Goda, N.; Makino, N.; Takamiya, Y.; Yamaguchi, T.; Ishimura, Y.; Hozumi, N.; Suematsu, M. Stabilization of mast cells by heme oxygenase-1: An anti-inflammatory role. Am. J. Physiol. Heart Circ. Physiol. 2002, 283, H861–H870. [Google Scholar] [CrossRef] [Green Version]
- Hosokawa, J.; Suzuki, K.; Nakagomi, D.; Tamachi, T.; Takatori, H.; Suto, A.; Nakajima, H. Role of calcium ionophore A23187-induced activation of IkappaB kinase 2 in mast cells. Int. Arch. Allergy Immunol. 2013, 161 (Suppl. S2), 37–43. [Google Scholar] [CrossRef]
- Kim, J.; Zarjou, A.; Traylor, A.M.; Bolisetty, S.; Jaimes, E.A.; Hull, T.D.; George, J.F.; Mikhail, F.M.; Agarwal, A. In vivo regulation of the heme oxygenase-1 gene in humanized transgenic mice. Kidney Int. 2012, 82, 278–291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonin, R.P.; Bories, C.; De Koninck, Y. A simplified up-down method (SUDO) for measuring mechanical nociception in rodents using von Frey filaments. Mol. Pain 2014, 10, 26. [Google Scholar] [CrossRef] [PubMed]
- Puebla-Osorio, N.; Sarchio, S.N.E.; Ullrich, S.E.; Byrne, S.N. Detection of Infiltrating Mast Cells Using a Modified Toluidine Blue Staining. Methods Mol. Biol. 2017, 1627, 213–222. [Google Scholar] [CrossRef] [PubMed]
- Aich, A.; Afrin, L.B.; Gupta, K. Mast Cell-Mediated Mechanisms of Nociception. Int. J. Mol. Sci. 2015, 16, 29069–29092. [Google Scholar] [CrossRef] [Green Version]
- Varvara, G.; Tettamanti, L.; E Gallenga, C.; Caraffa, A.; D’Ovidio, C.; Mastrangelo, F.; Ronconi, G.; Kritas, S.K.; Conti, P. Stimulated mast cells release inflammatory cytokines: Potential suppression and therapeutical aspects. J. Biol. Regul. Homeost. Agents 2018, 32, 1355–1360. [Google Scholar]
- Marshall, J.S. Mast-cell responses to pathogens. Nat. Rev. Immunol. 2004, 4, 787–799. [Google Scholar] [CrossRef]
- E Gallenga, C.; Pandolfi, F.; Caraffa, A.; Kritas, S.K.; Ronconi, G.; Toniato, E.; Martinotti, S.; Conti, P. Interleukin-1 family cytokines and mast cells: Activation and inhibition. J. Biol. Regul. Homeost. Agents 2019, 33, 1–6. [Google Scholar]
- Nigrovic, P.A.; Binstadt, B.A.; Monach, P.A.; Johnsen, A.; Gurish, M.; Iwakura, Y.; Benoist, C.; Mathis, D.; Lee, D.M. Mast cells contribute to initiation of autoantibody-mediated arthritis via IL-1. Proc. Natl. Acad. Sci. USA 2007, 104, 2325–2330. [Google Scholar] [CrossRef] [Green Version]
- King, C.A.; Anderson, R.; Marshall, J.S. Dengue virus selectively induces human mast cell chemokine production. J. Virol. 2002, 76, 8408–8419. [Google Scholar] [CrossRef] [Green Version]
- Ebert, S.; Becker, M.; Lemmermann, N.; Büttner, J.K.; Michel, A.; Taube, C.; Podlech, J.; Böhm, V.; Freitag, K.; Thomas, D.; et al. Mast cells expedite control of pulmonary murine cytomegalovirus infection by enhancing the recruitment of protective CD8 T cells to the lungs. PLoS Pathog. 2014, 10, e1004100. [Google Scholar] [CrossRef] [Green Version]
- Gasser, B.; Russenberger, M.E.; Berchtold, R.; Wyder, D.; Perren, S.M. Value of coining on the improvement of endurance in slotted intramedullary nails. J. Biomed. Eng. 1991, 13, 287–292. [Google Scholar] [CrossRef]
- Jiang, A.-P.; Jiang, J.-F.; Wei, J.-F.; Guo, M.-G.; Qin, Y.; Guo, Q.-Q.; Ma, L.; Liu, B.-C.; Wang, X.; Veazey, R.S.; et al. Human Mucosal Mast Cells Capture HIV-1 and Mediate Viral trans-Infection of CD4+ T Cells. J. Virol. 2015, 90, 2928–2937. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guma, M.; Kashiwakura, J.-I.; Crain, B.; Kawakami, Y.; Beutler, B.; Firestein, G.S.; Kawakami, T.; Karin, M.; Corr, M. JNK1 controls mast cell degranulation and IL-1beta production in inflammatory arthritis. Proc. Natl. Acad. Sci. USA 2010, 107, 22122–22127. [Google Scholar] [CrossRef] [Green Version]
- Kandere-Grzybowska, K.; Letourneau, R.; Kempuraj, D.; Donelan, J.; Poplawski, S.; Boucher, W.; Athanassiou, A.; Theoharides, T.C. IL-1 induces vesicular secretion of IL-6 without degranulation from human mast cells. J. Immunol. 2003, 171, 4830–4836. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burd, P.R.; Rogers, H.W.; Gordon, J.; A Martin, C.; Jayaraman, S.; Wilson, S.D.; Dvorak, A.M.; Galli, S.J.; E Dorf, M. Interleukin 3-dependent and -independent mast cells stimulated with IgE and antigen express multiple cytokines. J. Exp. Med. 1989, 170, 245–257. [Google Scholar] [CrossRef]
- Okayama, Y.; Ono, Y.; Nakazawa, T.; Church, M.K.; Mori, M. Human skin mast cells produce TNF-alpha by substance P. Int. Arch. Allergy Immunol. 1998, 117 (Suppl. S1), 48–51. [Google Scholar] [CrossRef] [PubMed]
- Tangudu, N.K.; Alan, B.; Vinchi, F.; Wörle, K.; Lai, D.; Vettorazzi, S.; Leopold, K.; Spasić, M.V. Scavenging Reactive Oxygen Species Production Normalizes Ferroportin Expression and Ameliorates Cellular and Systemic Iron Disbalances in Hemolytic Mouse Model. Antioxid. Redox Signal. 2018, 29, 484–499. [Google Scholar] [CrossRef] [Green Version]
- Couret, J.; Chang, T.L. Reactive Oxygen Species in HIV Infection. EC Microbiol. 2016, 3, 597–604. [Google Scholar]
- Elbim, C.; Pillet, S.; Prevost, M.H.; Preira, A.; Girard, P.M.; Rogine, N.; Matusani, H.; Hakim, J.; Israel, N.; Gougerot-Pocidalo, M.A. Redox and activation status of monocytes from human immunodeficiency virus-infected patients: Relationship with viral load. J. Virol. 1999, 73, 4561–4566. [Google Scholar] [CrossRef] [Green Version]
- Nagy, E.; Eaton, J.W.; Jeney, V.; Soares, M.; Varga, Z.; Galajda, Z.; Szentmiklósi, J.A.; Méhes, G.; Csonka, T.; Smith, A.; et al. Red cells, hemoglobin, heme, iron, and atherogenesis. Arter. Thromb. Vasc. Biol. 2010, 30, 1347–1353. [Google Scholar] [CrossRef]
- Smith, A.; McCulloh, R.J. Hemopexin and haptoglobin: Allies against heme toxicity from hemoglobin not contenders. Front. Physiol. 2015, 6, 187. [Google Scholar] [CrossRef]
- Hvidberg, V.; Maniecki, M.B.; Jacobsen, C.; Højrup, P.; Møller, H.J.; Moestrup, S.K. Identification of the receptor scavenging hemopexin-heme complexes. Blood 2005, 106, 2572–2579. [Google Scholar] [CrossRef]
- Maines, M.D. Heme oxygenase: Function, multiplicity, regulatory mechanisms, and clinical applications. FASEB J. 1988, 2, 2557–2568. [Google Scholar] [CrossRef] [Green Version]
- Nath, K.A. Heme oxygenase-1: A provenance for cytoprotective pathways in the kidney and other tissues. Kidney Int. 2006, 70, 432–443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gozzelino, R.; Jeney, V.; Soares, M.P. Mechanisms of cell protection by heme oxygenase-1. Annu. Rev. Pharmacol. Toxicol. 2010, 50, 323–354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dunn, L.L.; Midwinter, R.G.; Ni, J.; Hamid, H.A.; Parish, C.R.; Stocker, R. New insights into intracellular locations and functions of heme oxygenase-1. Antioxid. Redox Signal. 2014, 20, 1723–1742. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yachie, A. Heme Oxygenase-1 Deficiency and Oxidative Stress: A Review of 9 Independent Human Cases and Animal Models. Int. J. Mol. Sci. 2021, 22, 1514. [Google Scholar] [CrossRef]
- Yasui, Y.; Nakamura, M.; Onda, T.; Uehara, T.; Murata, S.; Matsui, N.; Fukuishi, N.; Akagi, R.; Suematsu, M.; Akagi, M. Heme oxygenase-1 inhibits cytokine production by activated mast cells. Biochem. Biophys. Res. Commun. 2007, 354, 485–490. [Google Scholar] [CrossRef]
- Chelombitko, M.A.; Fedorov, A.V.; Ilyinskaya, O.P.; Zinovkin, R.A.; Chernyak, B.V. Role of Reactive Oxygen Species in Mast Cell Degranulation. Biochemistry 2016, 81, 1564–1577. [Google Scholar] [CrossRef]
- Swindle, E.J.; Coleman, J.W.; DeLeo, F.R.; Metcalfe, D.D. FcepsilonRI- and Fcgamma receptor-mediated production of reactive oxygen species by mast cells is lipoxygenase- and cyclooxygenase-dependent and NADPH oxidase-independent. J. Immunol. 2007, 179, 7059–7071. [Google Scholar] [CrossRef] [Green Version]
- Swindle, E.J.; Metcalfe, D.D. The role of reactive oxygen species and nitric oxide in mast cell-dependent inflammatory processes. Immunol. Rev. 2007, 217, 186–205. [Google Scholar] [CrossRef] [PubMed]
- Kramkowski, K.; Leszczynska, A.; Mogielnicki, A.; Chlopicki, S.; Fedorowicz, A.; Grochal, E.; Mann, B.; Brzoska, T.; Urano, T.; Motterlini, R.; et al. Antithrombotic properties of water-soluble carbon monoxide-releasing molecules. Arter. Thromb. Vasc. Biol. 2012, 32, 2149–2157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adach, W.; Olas, B. Carbon monoxide and its donors—Their implications for medicine. Future Med. Chem. 2019, 11, 61–73. [Google Scholar] [CrossRef] [PubMed]
- Srisook, K.; Han, S.-S.; Choi, H.-S.; Li, M.-H.; Ueda, H.; Kim, C.; Cha, Y.-N. CO from enhanced HO activity or from CORM-2 inhibits both O2− and NO production and downregulates HO-1 expression in LPS-stimulated macrophages. Biochem. Pharmacol. 2006, 71, 307–318. [Google Scholar] [CrossRef] [PubMed]
- Adach, W.; Olas, B. The role of CORM-2 as a modulator of oxidative stress and hemostatic parameters of human plasma in vitro. PLoS ONE 2017, 12, e0184787. [Google Scholar] [CrossRef] [Green Version]
- Upadhyay, K.K.; Jadeja, R.N.; Vyas, H.S.; Pandya, B.; Joshi, A.; Vohra, A.; Thounaojam, M.C.; Martin, P.M.; Bartoli, M.; Devkar, R.V. Carbon monoxide releasing molecule-A1 improves nonalcoholic steatohepatitis via Nrf2 activation mediated improvement in oxidative stress and mitochondrial function. Redox Biol. 2020, 28, 101314. [Google Scholar] [CrossRef] [PubMed]
- Motterlini, R.; Sawle, P.; Bains, S.; Hammad, J.; Alberto, R.; Foresti, R.; Green, C.J. CORM-A1: A new pharmacologically active carbon monoxide-releasing molecule. FASEB J. 2005, 19, 284–286. [Google Scholar] [CrossRef] [Green Version]
- Green, D.P.; Limjunyawong, N.; Gour, N.; Pundir, P.; Dong, X. A Mast-Cell-Specific Receptor Mediates Neurogenic Inflammation and Pain. Neuron 2019, 101, 412–420.e3. [Google Scholar] [CrossRef] [Green Version]
- Aich, A.; Jones, M.K.; Gupta, K. Pain and sickle cell disease. Curr. Opin. Hematol. 2019, 26, 131–138. [Google Scholar] [CrossRef]
- Khalilzadeh, E.; Azarpey, F.; Hazrati, R.; Vafaei Saiah, G. Evaluation of different classes of histamine H(1) and H(2) receptor antagonist effects on neuropathic nociceptive behavior following tibial nerve transection in rats. Eur. J. Pharmacol. 2018, 834, 221–229. [Google Scholar] [CrossRef]
- Choi, S.I.; Hwang, S.W. Depolarizing Effectors of Bradykinin Signaling in Nociceptor Excitation in Pain Perception. Biomol. Ther. 2018, 26, 255–267. [Google Scholar] [CrossRef] [PubMed]
- Allali, S.; Lionnet, F.; Mattioni, S.; Callebert, J.; Stojanovic, K.S.; Bachmeyer, C.; Arlet, J.; Brousse, V.; de Montalembert, M.; Chalumeau, M.; et al. Plasma histamine elevation in a large cohort of sickle cell disease patients. Br. J. Haematol. 2019, 186, 125–129. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.J.; Wilkie, D.J.; Molokie, R. Neurobiological mechanisms of pain in sickle cell disease. Hematol. Am. Soc. Hematol. Educ. Program 2010, 2010, 403–408. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group (Number of Participants) | HIVneg, Painneg (14) | HIVneg, Painpos (14) | HIVpos, Painneg (14) | HIVpos, Painpos (13) |
|---|---|---|---|---|
| Avg. age (SD) | 50.1 (12.6) | 45.3 (15.0) | 48.6 (8.4) | 54.1 (5.8) |
| Percent Females | 64.3 | 64.3 | 50 | 38.5 |
| Percent Afr. Amer. | 64.3 | 64.3 | 71.4 | 76.9 |
| Avg. CD4+ (SD) | 715.4 (421.2) | 637.5 (385.0) | ||
| Avg. Nadir CD4+ (SD) | 165.0 (231.5) | 204.5 (202.4) | ||
| Avg. VL (SD) | 19.5 (1.9) | 59.2 (130.7) | ||
| Avg. Highest VL (SD) | 199,115 (249,762) | 428,995 (1,220,712) | ||
| Plasma heme (µM) (SD) | 14.4 (4.0) | 26.3 (13.0) | 27.4 (12.4) | 80.9 (62.4) a,b,c |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chatterjee, T.; Arora, I.; Underwood, L.; Gryshyna, A.; Lewis, T.L.; Masjoan Juncos, J.X.; Goodin, B.R.; Heath, S.; Aggarwal, S. High Heme and Low Heme Oxygenase-1 Are Associated with Mast Cell Activation/Degranulation in HIV-Induced Chronic Widespread Pain. Antioxidants 2023, 12, 1213. https://doi.org/10.3390/antiox12061213
Chatterjee T, Arora I, Underwood L, Gryshyna A, Lewis TL, Masjoan Juncos JX, Goodin BR, Heath S, Aggarwal S. High Heme and Low Heme Oxygenase-1 Are Associated with Mast Cell Activation/Degranulation in HIV-Induced Chronic Widespread Pain. Antioxidants. 2023; 12(6):1213. https://doi.org/10.3390/antiox12061213
Chicago/Turabian StyleChatterjee, Tanima, Itika Arora, Lilly Underwood, Anastasiia Gryshyna, Terry L. Lewis, Juan Xavier Masjoan Juncos, Burel R. Goodin, Sonya Heath, and Saurabh Aggarwal. 2023. "High Heme and Low Heme Oxygenase-1 Are Associated with Mast Cell Activation/Degranulation in HIV-Induced Chronic Widespread Pain" Antioxidants 12, no. 6: 1213. https://doi.org/10.3390/antiox12061213