
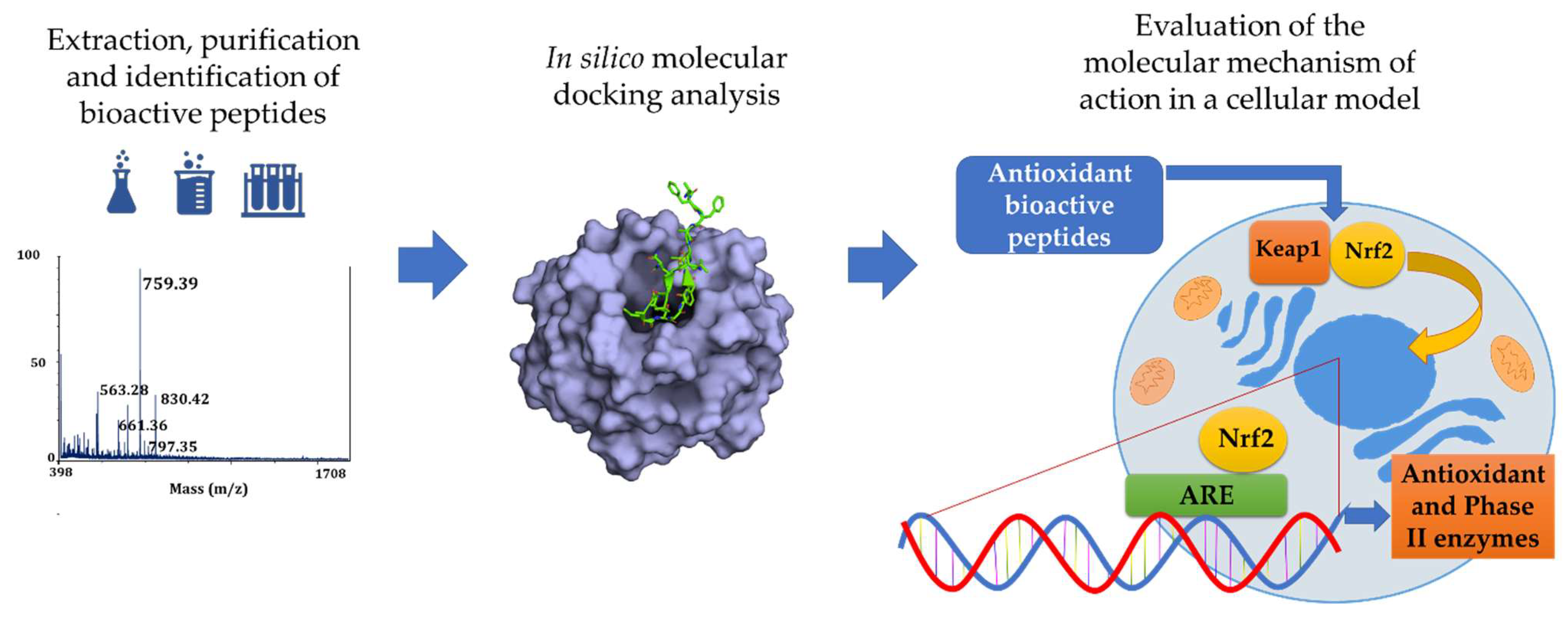
From In Silico to a Cellular Model: Molecular Docking Approach to Evaluate Antioxidant Bioactive Peptides

Abstract
:1. Introduction
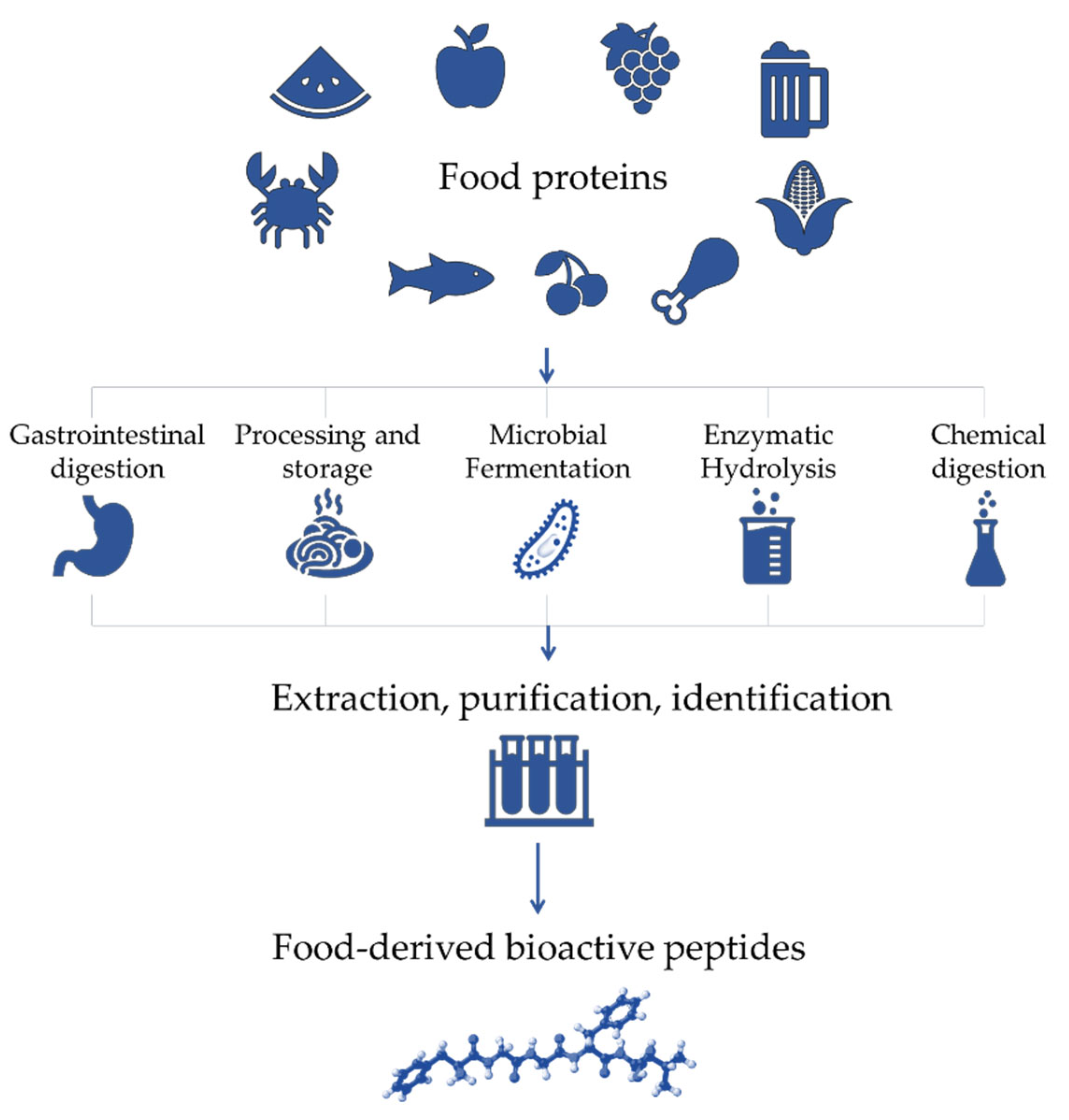
2. Antioxidant Bioactive Peptides
2.1. Applications of Antioxidant Bioactive Peptides
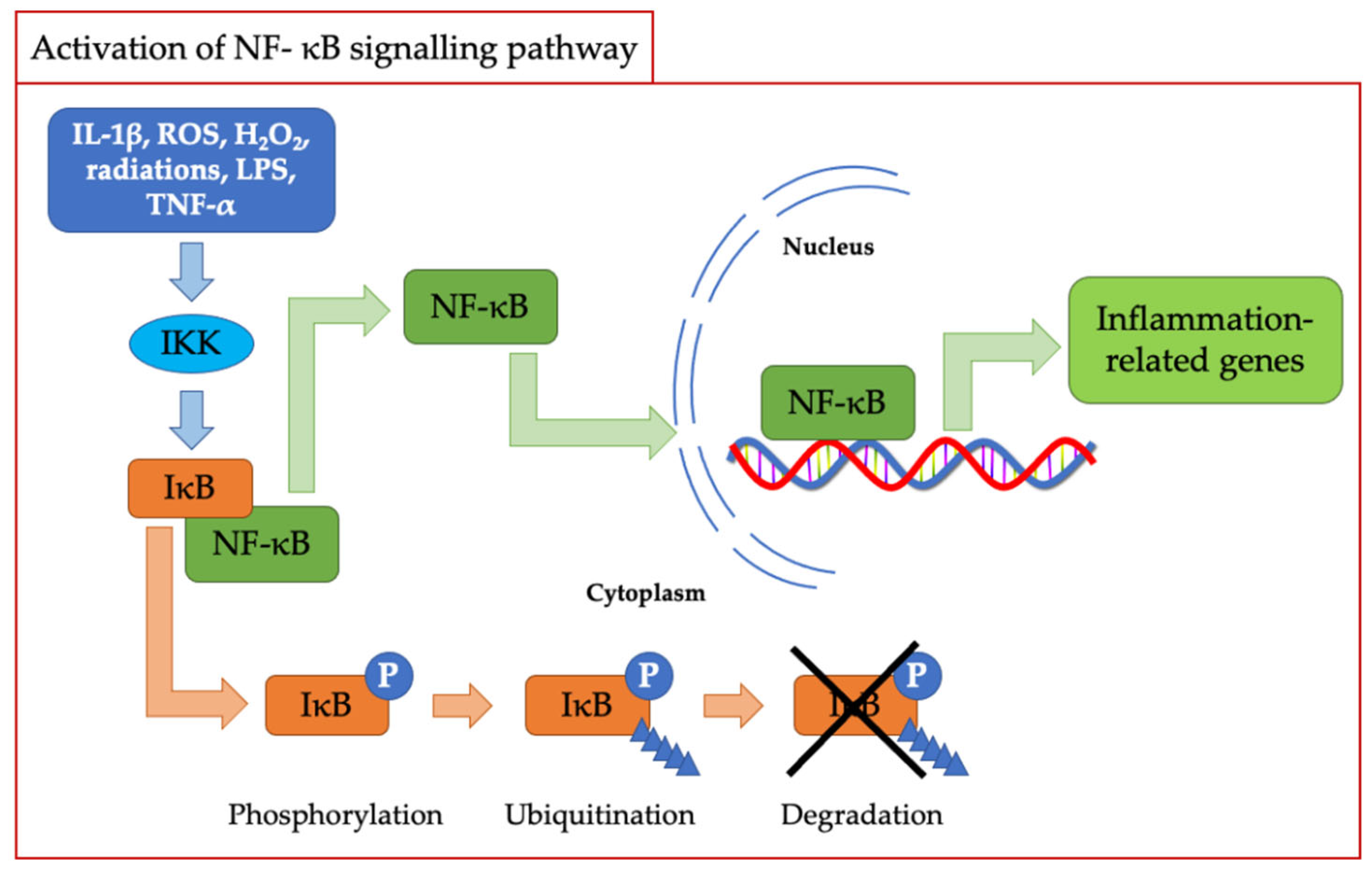
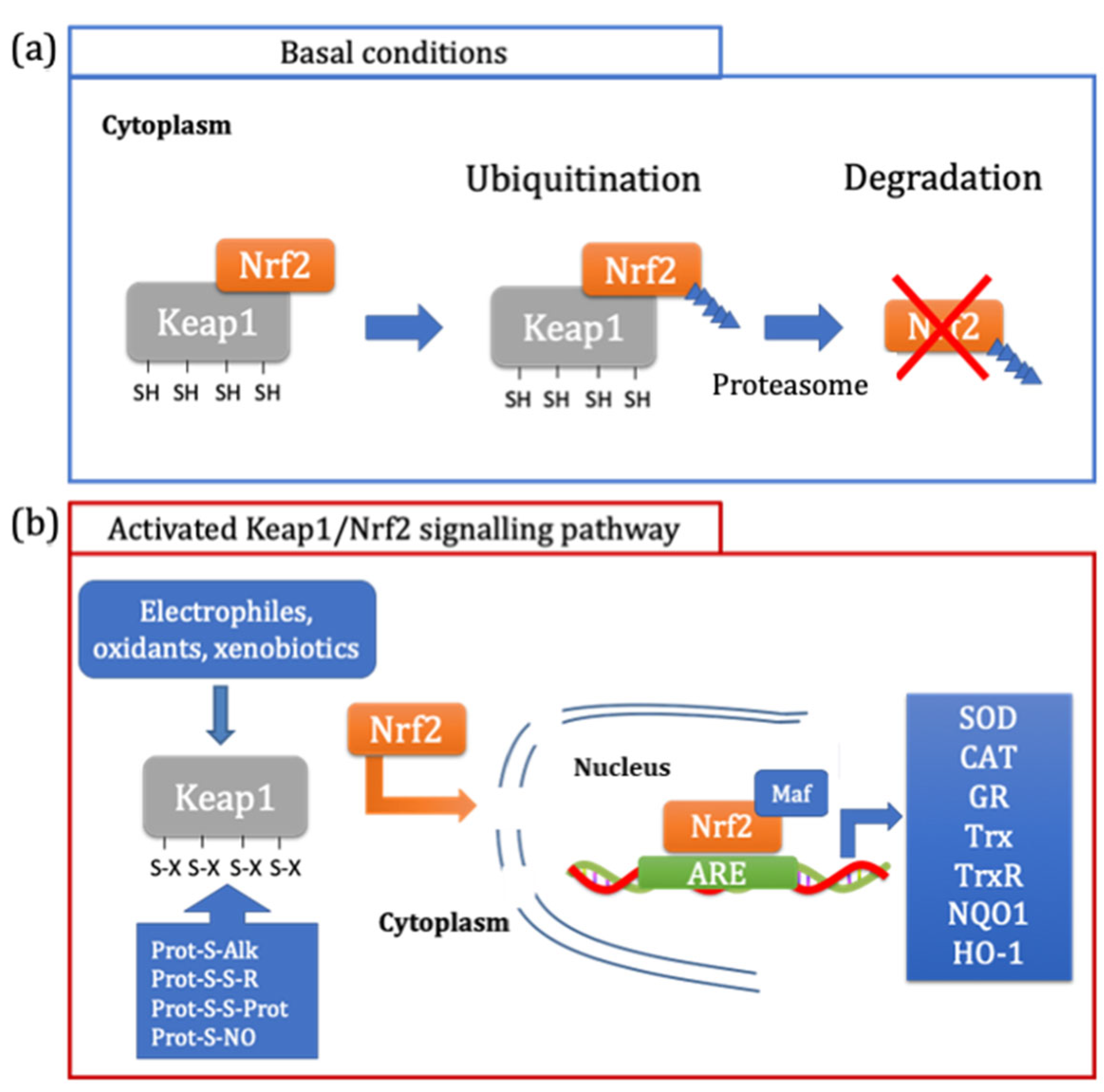
2.2. Redox Signalling
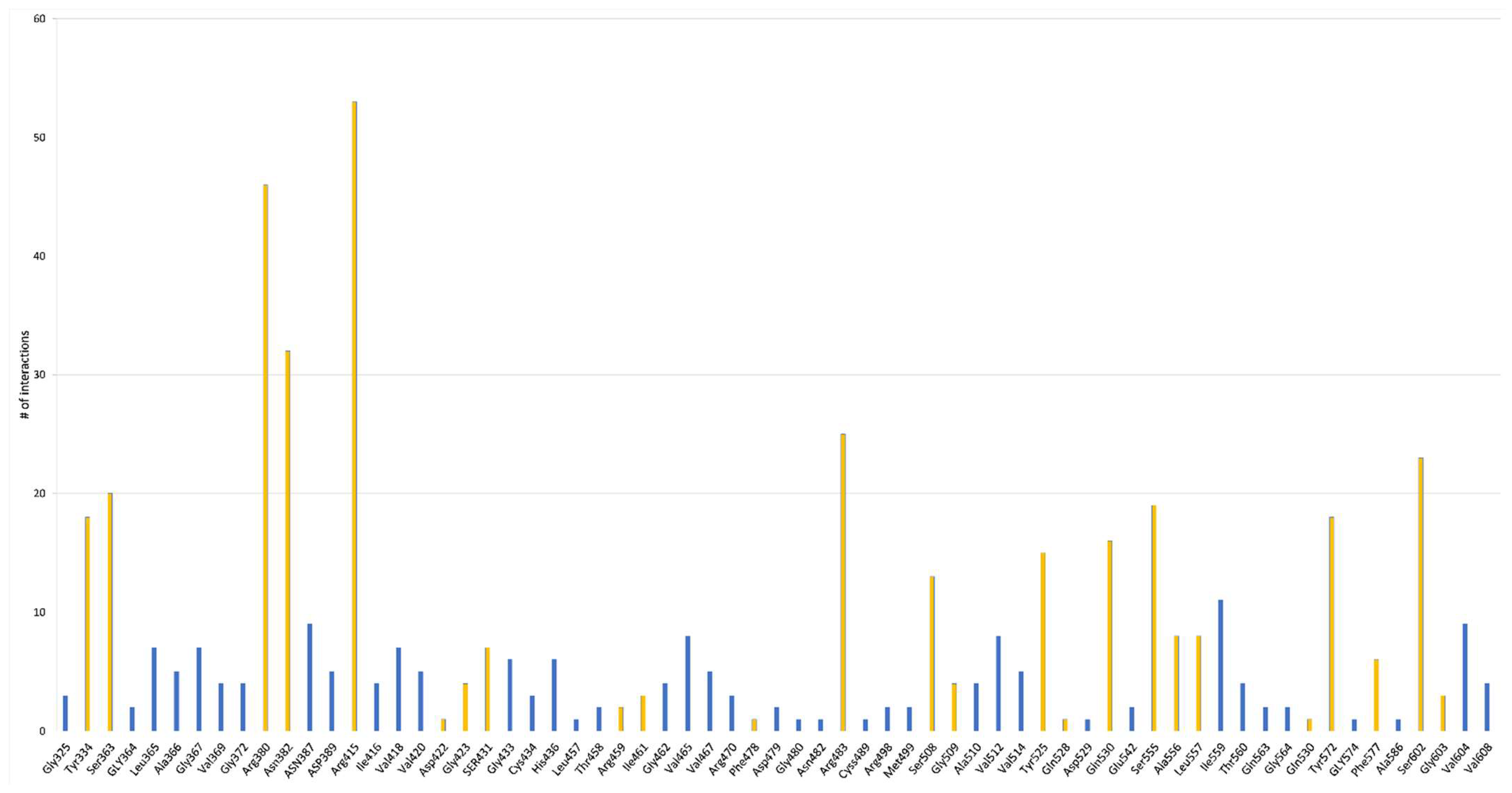
3. In Silico Prediction of Bioactive Peptides Docking
In Silico Prediction of Antioxidant Peptides Activity

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| aa | Sequence | pI | Net Charge | Kelch Domain Interaction | Validation in Cellular Studies | Ref. | Ref. |
|---|---|---|---|---|---|---|---|
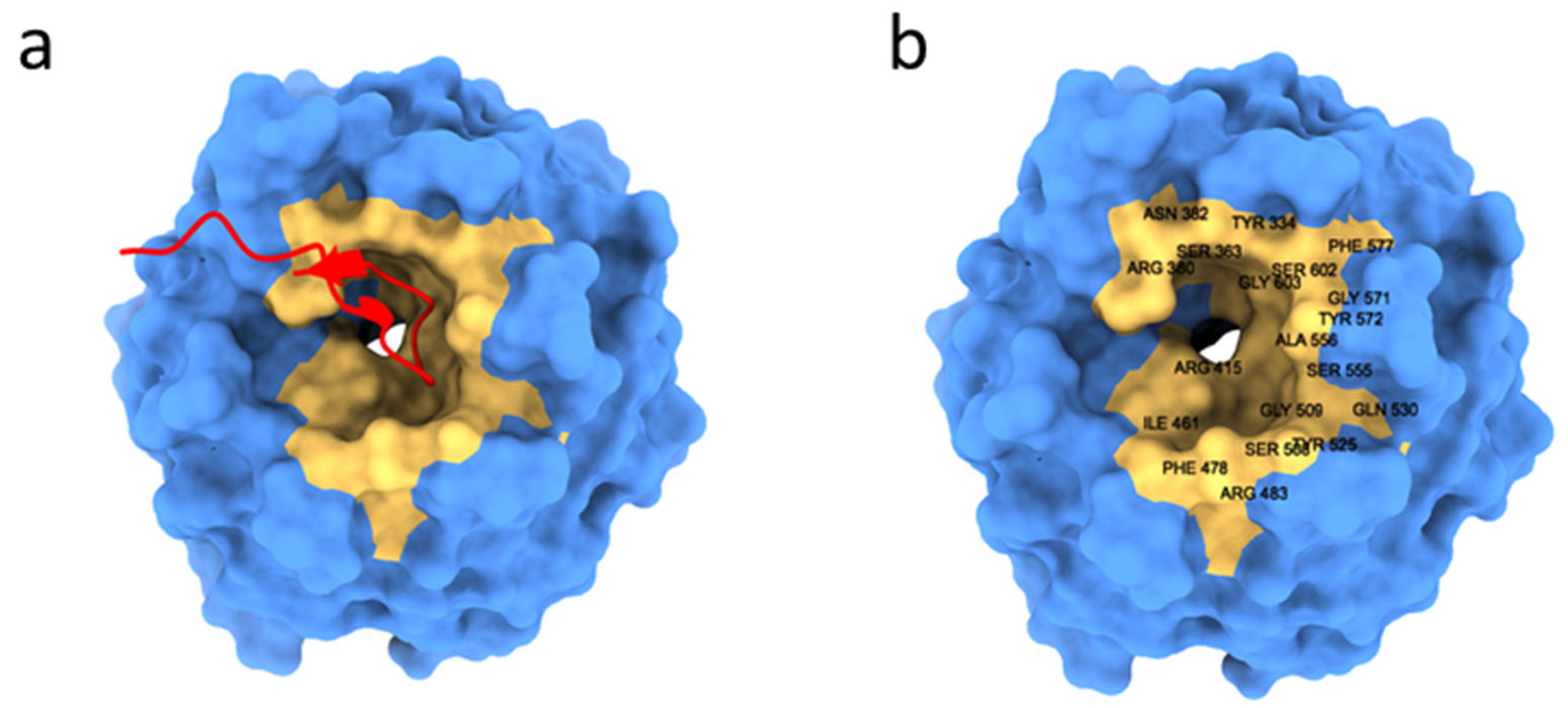
| 8 | KVLPVPEK | 9.63 | +1 | Gln337, Ser383, Asn382, Asn387, Tyr334, Arg380 and Ser363 | yes | [60] | |
| 5 | EDYGA | 2.87 | −3 | Arg415 | no | [61] | |
| 10 | DEQIPSHPPR | 5.21 | −1 | Arg380, Asn382 and Arg415 | yes | [62] | |
| 10 | SLVNNDDRDS | 3.53 | −2 | Tyr334, Arg380, Asn414, Arg415 and Tyr525 | yes | [62] | |
| 11 | VNPESQQGSPR | 6.99 | 0 | Tyr334, Arg415, Arg483 and Tyr572 | yes | [62] | |
| 11 | IGINAENNQRN | 6.99 | 0 | Ser363, Asn382, Asp385, Arg415, Arg483, Ser508, Gln530 and Ser602 | yes | [62] | |
| 12 | FVDAQPQQKEEG | 3.77 | −2 | Tyr334, Arg380, Asn382, Asn387, Arg415, Arg483 and Gln530 | yes | [62] | |
| 12 | FGREEGQQQGEE | 3.7 | −3 | Arg336, Ser363, Arg380, Asn382, Arg415, Arg483, Tyr525, Tyr572 and Ser602 | yes | [62] | |
| 13 | MRKPQQEEDDDDE | 3.53 | −5 | Arg380, Asp389, Arg415, Ser431, His436, Arg483 and Ser602 | yes | [62] | |
| 9 | YLAGNQEQE | 3.09 | −2 | Arg380 and Arg415 | yes | [62] | |
| 14 | NALEPDHRVESEGG | 4.18 | −3 | Tyr334, Arg380, Asn414, Arg415, His432 and Ser602 | yes | [62] | |
| 14 | KEQQQEQQQEEQPL | 3.70 | -3 | Tyr334, Arg380, Asn414, Arg415, Ser431, Arg483 and Ser602 | yes | [62] | |
| 14 | HEQKEEHEWHRKEE | 5.31 | −3 | Arg336, Ser363, Arg380, Asn382, Asp385, Asn387, Asp389, Asn414, Arg415, Arg483 and Ser602 | yes | [62] | |
| 14 | GKHQQEEENEGGSI | 4.28 | −3 | Ser363, Arg380, Asp389, Asn414, Tyr572 and Ser602 | yes | [62] | |
| 14 | QGPIVLNPWDQVKR | 10.12 | 1 | Arg380, Asn382, Asn387, Arg415, His432, Ser508, Tyr525, Gln530, His575 and Thr576 | yes | [63] | |
| 15 | NTVPAKSCQAQPTTM | 8.97 | 1 | Arg336, Arg380, Asn382, Arg415, Gly433, Ile435, Gly509, Tyr572, Thr576 and Ser602 | yes | [63] | |
| 17 | APSFSDIPNPIGSENSE | 2.93 | −3 | Arg336, Ser363, Arg415 and Tyr572 | yes | [63] | |
| 9 | VLSTSFCPK | 8.67 | +1 | Cys434, Asp479, Thr458, Leu457, Met499, Cys489, Glu542, Arg459, Met499, Arg498 and Glu542 | yes | [67] | |
| 9 | VLSTSFYPK | 9.48 | +1 | Cys434, Asp459, Met499, Cys489, Glu542, His436, Gly480, Arg459 and Thr458 | yes | [67] | |
| 8 | IVLPDEGK | 1.01 | −1 | Arg380 and Arg415, His436, Ile461, Arg483, Ser508, Ser555 and Tyr572 | yes | [68] | |
| 10 | SDGSNIHFPN | 4.98 | −1 | Leu365, Arg380 and Arg415. Additionally, Gly462, Arg483, Ala510, Tyr525, Ala556, Leu557, Tyr572 and Gly603 | yes | [68] | |
| 17 | PGMLGGSPPGLLGGSPP | 5.25 | 0 | Gly364, Leu365, Ala366 and Arg380, Asn382, Arg415, Ile416, Gly433, Arg483, Cys434, Ala510, Tyr525, Leu557, Tyr572, Gly603 and Val604 | yes | [68] | |
| 6 | VLFSNY | 5.53 | 0 | Arg380, Asn382, Arg415, Arg483, Ser508, Ser555 and Ser602 | yes | [69] | |
| 7 | FYSLHTF | 7.64 | 0 | Arg380, Asn414, Arg415, Ser431, Gln530 and Ser602 | yes | [69] | |
| 7 | VYGYADK | 6.41 | 0 | Arg336, Arg380, Asn414, Arg415, Gln530 and Ser602 | yes | [69] | |
| 8 | TFQGPPHG | 7.91 | 0 | Arg380, Asn382, Asn414, Arg415, Ser431, Gly433, Ser555 and Ser602 | yes | [69] | |
| 8 | YTPEYQTK | 6.5 | 0 | Tyr334, Ser363, Arg380, Asn382, Arg415, Ser431, His436, Arg483, Tyr525, Gln530, Ser555 and Ser602 | yes | [69] | |
| 10 | SSGHTLPAGV | 7.89 | 0 | Arg380, Arg415, Arg483, Tyr525, Gln530 and Ser555 | yes | [69] | |
| 10 | SGDWSDIGGR | 3.92 | −1 | Tyr334, Gly364, Arg380, Arg483, Tyr525, Gln530, gly574 and Ser602 | yes | [70] | |
| 6 | RDPEER | 4.32 | −1 | Asn382, Arg380 and Tyr334 | no | [71] | |
| 5 | SPSSS | 5.38 | 0 | Ser363, Asn382, Asn387 and Ser555 | yes | [72] | |
| 5 | SGTAV | 5.54 | 0 | Tyr334, Asn382, Ser383, Asn414, Arg415, Ser555 and Tyr572 | yes | [72] | |
| 5 | NSVAA | 5.38 | 0 | Ser363, Asn387, Asn414, Arg415, Ser508, Ser555 and Gly603 | yes | [72] | |
| 4 | DLEE | 2.74 | −3 | Val418, Val465, Ile416, Arg415 and Val420 | yes | [73] | |
| 5 | LWNPR | 10.73 | +1 | Ser363, Arg380, Asn382, Arg415, His436, Tyr572 and Phe577 | yes | [74] | |
| 6 | KPLCPP | 9.29 | +1 | Arg380, Arg415, Gln530, Tyr525, Ala556 and Ser602 | yes | [74] | |
| 8 | YSNQNGRF | 9.69 | +1 | Tyr334, Arg380, Asn382, Arg415, Ser508, Tyr525, Gln530, Ser555 and Ser602 | no | [75] | |
| 3 | SPW | 5.42 | 0 | Arg380, Asn382 and Ser 602 | no | [76] | |
| 3 | STW | 5.42 | 0 | Arg380 and Asn382 | no | [76] | |
| 3 | QKW | 9.98 | +1 | Arg380, Asn387, Asp389, Arg415, Ser431 and Gly433 | no | [76] | |
| 3 | MKW | 9.98 | +1 | Tyr525, Gln530 and Ser555 | no | [76] | |
| 3 | ETW | 3.09 | −1 | Tyr334, Arg380 and Asn382 | no | [76] | |
| 3 | SVW | 5.42 | 0 | Tyr334, Arg336 and asn382 | no | [76] | |
| 3 | CNW | 4.94 | 0 | Gln528, Gln530 and Ser555 | no | [76] | |
| 3 | DHW | 4.98 | −1 | Ser363, Arg380, Asn382 and Arg415 | no | [76] | |
| 3 | GQW | 5.55 | 0 | Gly480, Arg483, Arg415 and Ser508 | no | [76] | |
| 3 | SQW | 5.42 | 0 | Arg380, Asn382 and Tyr572 | no | [76] | |
| 4 | EGCG | 3.09 | −1 | Asn414, Arg389, Ser 555 and Ser602 | yes | [76] | |
| 3 | VPN | 5.4 | 0 | Tyr334, Ser363, Asn382 and Gln530 | no | [77] | |
| 4 | DREL | 4.0 | −1 | Arg135 and Gly148 | no | [77] | |
| 3 | DKK | 9.63 | +1 | Arg380 and Asn382 | no | [78] | |
| 3 | DDW | 2.78 | −2 | Arg415, Arg380, Asn382, Ser508 and Arg483 | no | [78] | |
| 5 | LYSPH | 7.65 | 0 | Tyr334, Ser363, Arg380, Asn382, Arg415, Arg483, Ser508, Tyr525, Gln530, Ala556, Tyr572, Phe577 and Ser602 | no | [78] | |
| 6 | LPHFNS | 7.63 | 0 | Tyr334, Ser363, Arg380, Asn382, Asn414, Arg415, Arg483, Tyr525, Gln530, Ser555, Ala556, Tyr572, Phe577 and Ser602 | no | [78] | |
| 7 | AEHGSLH | 6.05 | −1 | Tyr334, Arg336, Ser363, Arg380, Asn382, Ser383, Pro384, Arg415, Ile461, Arg483, Ser508, Tyr525, Gln530, Ala556, Tyr572 and Ser602 | no | [78] | |
| 7 | FGPEMEQ | 2.97 | −2 | Tyr334, Ser363, Arg380, Asn382, Asn387, Asp389, Asn414, Arg415, Gly433, Ile461, Ser555, Ala556, Tyr572 and Phe577 | no | [78] | |
| 9 | PSYLNTPLL | 5.22 | 0 | Tyr334, Ser363, Gly364, Arg380, Asn382, Arg415, Arg483, Tyr525, Gln530, Ser555, Ala556, Tyr572 and Phe577 | no | [78] | |
| 3 | DDL | 2.91 | −2 | Ala366, Gly367, Arg415, Val465, Val512, Ile559 and Val604 | yes | [79] | |
| 4 | LSEE | 3.09 | −2 | Ala366, Gly367, Arg415, Ile416, Gly462, Arg483, Gly509, Val512 and Val604 | yes | [79] | |
| 4 | TGEV | 3.27 | −1 | Gly367, Arg415, Val418, Gly462, Leu557 and Val604 | yes | [79] | |
| 4 | TVEE | 3.09 | −2 | Leu365, Val420, val514, Leu557, Ile559 and Val604 | yes | [79] | |
| 4 | TVET | 3.27 | −1 | Leu365, Ala366, Arg415, Val418, Val465, Ile 559 and Val604 | yes | [79] | |
| 4 | TFEE | 3.09 | −2 | Ala366, Arg380, Arg415, Val418, Ala510, Val512, Leu557 and Ile559 | yes | [79] | |
| 4 | LEHL | 5.11 | −1 | Arg415, Val418, Val465, Val512, Leu557 and Ile559 | yes | [79] | |
| 4 | HELE | 4.27 | −2 | Gly367, Arg380, Leu557, Leu559 and Val604 | yes | [79] | |
| 5 | NEGPQ | 3.27 | −1 | Leu365, Arg380, Arg415, Val418, Val465, Val512 and Ile559 | yes | [79] | |
| 7 | WGDAGAE | 3.01 | −2 | Gly367, Arg415, Ile416, Arg483, Val512, Ile559 and Val604 | yes | [79] | |
| 4 | ICRD | 6.09 | 0 | Tyr85, Ala 88, His129, Lys131, Val132, Arg135, Cys151, His154 and Val 155 | yes | [80] | |
| 5 | LCGEC | 3.20 | −1 | His129, Lys131, Val132, Arg135, Met147, Gly148, Lys150, Cys151, His154 and Val 155 | yes | [80] | |
| 6 | RVIEPR | 10.58 | +1 | Val369, Val467 and Val561 | yes | [81] | |
| 7 | SGFSTEL | 3.13 | −1 | Val465, Ile559 and Val608 | yes | [81] | |
| 7 | ISREEAQ | 4.09 | −1 | Gly367, Val418, Val465, Val467, Val512, Thr560, Val561, Val606 and Val608 | yes | [81] | |
| 9 | ERYQEQGYQ | 4.08 | −1 | Gly372, Arg470, Val514, Ile559, Thr560 and Val608 | yes | [81] | |
| 9 | ERYQEQGYQ | 4.08 | −1 | Gly325, Val369, Gly371, Gly372, Gly423 and Gln563 | yes | [81] | |
| 11 | LQEQEQGQVQS | 3.03 | −2 | Gly325, Val369, Gly371, Val420, Val467, Val514, Thr560 and Met610 | yes | [81] | |
| 11 | KEEQTQAYLPT | 4.08 | −1 | Gly325, Arg470, Ile559, Val561 and Val606 | yes | [81] | |
| 13 | IDNPNRADTYNPR | 6.56 | 0 | Gly371, Gly372, Gly423, Val514 and Gly564 | yes | [81] | |
| 13 | IDNPQSSDIFNPH | 3.92 | −2 | Ser363, Asn382, Ser508 and Gly509 | yes | [81] | |
| 14 | NIDNPQSSDIFNPH | 3.91 | −2 | Val369, Val420, Asp422, Gly423, Val467, Arg470, Val514, Thr560, Gln563 and Gly564 | yes | [81] | |
| 6 | SGFDAE | 3.01 | −2 | Ser363, Leu365, Asn414, Ala510, Ser555, Ala556, Tyr572, Phe577 and Ser602 | yes | [82] | |
| 8 | YPFPGPIH | 7.83 | 0 | Arg 415 and Gly 367 | yes | [83] | |
| 9 | VTSALVGPR | 11.6 | 1 | Gly423, Val420 and Asn469 | yes | [84] | |
| 10 | DEQIPSHPPR | 5.1 | −1 | Tyr334, Ser363, Arg380, Arg382, Arg415, Ser431, Gly433, His436, Gly462, Phe478, Arg483, Ser508, Gly509, Tyr525, Leu557 and Ser602 | yes | [85] |
4. From In Silico Analysis to Cellular Studies
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- United Nations, Department of Economic and Social Affairs, Populate Division. World Population Prospects 2022: Summary of the Results; United Nations: New York, NY, USA, 2022. [Google Scholar]
- Mada, S.B.; Ugwu, C.P.; Abarshi, M.M. Health Promoting Effects of Food-Derived Bioactive Peptides: A Review. Int. J. Pept. Res. Ther. 2020, 26, 831–848. [Google Scholar] [CrossRef]
- Lv, R.; Dong, Y.; Bao, Z.; Zhang, S.; Lin, S.; Sun, N. Advances in the Activity Evaluation and Cellular Regulation Pathways of Food-Derived Antioxidant Peptides. Trends Food Sci. Technol. 2022, 122, 171–186. [Google Scholar] [CrossRef]
- Manzoor, M.; Singh, J.; Gani, A. Exploration of Bioactive Peptides from Various Origin as Promising Nutraceutical Treasures: In Vitro, in Silico and in Vivo Studies. Food Chem. 2022, 373, 131395. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Lao, F.; Pan, X.; Wu, J. Food Protein-Derived Antioxidant Peptides: Molecular Mechanism, Stability and Bioavaila-bility. Biomolecules 2022, 12, 1622. [Google Scholar] [CrossRef] [PubMed]
- Naeem, M.; Malik, M.I.; Umar, T.; Ashraf, S.; Ahmad, A. A Comprehensive Review About Bioactive Peptides: Sources to Future Perspective. Int. J. Pept. Res. Ther. 2022, 28, 155. [Google Scholar] [CrossRef]
- Du, Z.; Li, Y. Review and Perspective on Bioactive Peptides: A Roadmap for Research, Development, and Future Opportunities. J. Agric. Food Res. 2022, 9, 100353. [Google Scholar] [CrossRef]
- Chakrabarti, S.; Guha, S.; Majumder, K. Food-Derived Bioactive Peptides in Human Health: Challenges and Opportunities. Nutrients 2018, 10, 1738. [Google Scholar] [CrossRef] [Green Version]
- Korhonen, H.; Pihlanto, A. Food-Derived Bioactive Peptides--Opportunities for Designing Future Foods. Curr. Pharm. Des. 2003, 9, 1297–1308. [Google Scholar] [CrossRef] [Green Version]
- Daliri, E.B.-M.; Lee, B.H.; Oh, D.H. Current Trends and Perspectives of Bioactive Peptides. Crit. Rev. Food Sci. Nutr. 2018, 58, 2273–2284. [Google Scholar] [CrossRef]
- Gallego, M.; Mora, L.; Toldrá, F. Health Relevance of Antihypertensive Peptides in Foods. Curr. Opin. Food Sci. 2018, 19, 8–14. [Google Scholar] [CrossRef]
- Jahandideh, F.; Wu, J. Perspectives on the Potential Benefits of Antihypertensive Peptides towards Metabolic Syndrome. Int. J. Mol. Sci. 2020, 21, 2192. [Google Scholar] [CrossRef] [Green Version]
- Yan, J.; Zhao, J.; Yang, R.; Zhao, W. Bioactive Peptides with Antidiabetic Properties: A Review. Int. J. Food Sci. Technol. 2019, 54, 1909–1919. [Google Scholar] [CrossRef] [Green Version]
- Mora, L.; González-Rogel, D.; Heres, A.; Toldrá, F. Iberian Dry-Cured Ham as a Potential Source of α-Glucosidase-Inhibitory Peptides. J. Funct. Foods 2020, 67, 103840. [Google Scholar] [CrossRef]
- Nongonierma, A.B.; Mooney, C.; Shields, D.C.; FitzGerald, R.J. In Silico Approaches to Predict the Potential of Milk Pro-tein-Derived Peptides as Dipeptidyl Peptidase IV (DPP-IV) Inhibitors. Peptides 2014, 57, 43–51. [Google Scholar] [CrossRef]
- Zaky, A.A.; Simal-Gandara, J.; Eun, J.-B.; Shim, J.-H.; Abd El-Aty, A.M. Bioactivities, Applications, Safety, and Health Benefits of Bioactive Peptides From Food and By-Products: A Review. Front. Nutr. 2021, 8, 815640. [Google Scholar] [CrossRef]
- Guha, S.; Majumder, K. Structural-FeatuRes. of Food-Derived Bioactive Peptides with Anti-Inflammatory Activity: A Brief Review. J. Food Chem. Biochem. 2019, 43, e12531. [Google Scholar] [CrossRef] [Green Version]
- Rivera-Jiménez, J.; Berraquero-García, C.; Pérez-Gálvez, R.; García-Moreno, P.J.; Javier Espejo-Carpio, F.; Guadix, A.; Guadix, E.M. Peptides and Protein Hydrolysates Exhibiting Anti-Inflammatory Activity: Sources, Structural FeatuRes. and Modulation Mechanisms. Food Funct. 2022, 13, 12510–12540. [Google Scholar] [CrossRef]
- Chakrabarti, S.; Jahandideh, F.; Wu, J. Food-Derived Bioactive Peptides on Inflammation and Oxidative Stress. BioMed. Res. Int. 2014, 2014, 608979. [Google Scholar] [CrossRef] [Green Version]
- Karami, Z.; Akbari-adergani, B. Bioactive Food Derived Peptides: A Review on Correlation between Structure of Bioactive Peptides and Their Functional Properties. J. Food Sci. Technol. 2019, 56, 535–547. [Google Scholar] [CrossRef]
- Manzanares, P.; Gandía, M.; Garrigues, S.; Marcos, J.F. Improving Health-Promoting Effects of Food-Derived Bioactive Peptides through Rational Design and Oral Delivery Strategies. Nutrients 2019, 11, 2545. [Google Scholar] [CrossRef] [Green Version]
- Akbarian, M.; Khani, A.; Eghbalpour, S.; Uversky, V.N. Bioactive Peptides: Synthesis, Sources, Applications, and Proposed Mechanisms of Action. Int. J. Mol. Sci. 2022, 23, 1445. [Google Scholar] [CrossRef] [PubMed]
- Pan, M.; Liu, K.; Yang, J.; Liu, S.; Wang, S.; Wang, S. Advances on Food-Derived Peptidic Antioxidants—A Review. Antioxidants 2020, 9, 799. [Google Scholar] [CrossRef] [PubMed]
- Toldrá, F.; Reig, M.; Aristoy, M.-C.; Mora, L. Generation of Bioactive Peptides during Food Processing. Food Chem. 2018, 267, 395–404. [Google Scholar] [CrossRef] [PubMed]
- Freitas, A.C.; Andrade, J.C.; Silva, F.M.; Rocha-Santos, T.A.P.; Duarte, A.C.; Gomes, A.M. Antioxidative Peptides: Trends and Perspectives for Future Research. Curr. Med. Chem. 2013, 20, 4575–4594. [Google Scholar] [CrossRef]
- Park, Y.W.; Nam, M.S. Bioactive Peptides in Milk and Dairy Products: A Review. Korean J. Food Sci. Anim. Resour. 2015, 35, 831–840. [Google Scholar] [CrossRef] [Green Version]
- Mardani, M.; Badakné, K.; Farmani, J.; Aluko, R.E. Antioxidant Peptides: Overview of Production, Properties, and Applications in Food Systems. Compr. Rev. Food Sci. Food Saf. 2023, 22, 46–106. [Google Scholar] [CrossRef]
- Liao, W.; Chakrabarti, S.; Davidge, S.T.; Wu, J. Modulatory Effects of Egg White Ovotransferrin-Derived Tripeptide IRW (Ile-Arg-Trp) on Vascular Smooth Muscle Cells against Angiotensin II Stimulation. J. Agric. Food Chem. 2016, 64, 7342–7347. [Google Scholar] [CrossRef]
- Chicken Muscle Protein-Derived Peptide VVHPKESF Reduces TNFα-Induced Inflammation and Oxidative Stress by Sup-pressing TNFR1 Signaling in Human Vascular Endothelial Cells—Fan—2022—Molecular Nutrition & Food Research—Wiley Online Library. Available online: https://onlinelibrary.wiley.com/doi/full/10.1002/mnfr.202200184 (accessed on 27 February 2023).
- López-García, G.; Dublan-García, O.; Arizmendi-Cotero, D.; Gómez Oliván, L.M. Antioxidant and Antimicrobial Peptides Derived from Food Proteins. Molecules 2022, 27, 1343. [Google Scholar] [CrossRef]
- Abeyrathne, E.D.N.S.; Nam, K.; Huang, X.; Ahn, D.U. Plant- and Animal-Based Antioxidants’ Structure, Efficacy, Mechanisms, and Applications: A Review. Antioxidants 2022, 11, 1025. [Google Scholar] [CrossRef]
- Lorenzo, J.M.; Munekata, P.E.S.; Gómez, B.; Barba, F.J.; Mora, L.; Pérez-Santaescolástica, C.; Toldrá, F. Bioactive Peptides as Natural Antioxidants in Food Products—A Review. Trends Food Sci. Technol. 2018, 79, 136–147. [Google Scholar] [CrossRef]
- Bhandari, D.; Rafiq, S.; Gat, Y.; Gat, P.; Waghmare, R.; Kumar, V. A Review on Bioactive Peptides: Physiological Functions, Bioavailability and Safety. Int. J. Pept. Res. Ther. 2020, 26, 139–150. [Google Scholar] [CrossRef]
- Punia, H.; Tokas, J.; Malik, A.; Sangwan, S.; Baloda, S.; Singh, N.; Singh, S.; Bhuker, A.; Singh, P.; Yashveer, S.; et al. Identi-fication and Detection of Bioactive Peptides in Milk and Dairy Products: Remarks about Agro-Foods. Molecules 2020, 25, 3328. [Google Scholar] [CrossRef]
- Bindoli, A.; Rigobello, M.P. Principles in Redox Signaling: From Chemistry to Functional Significance. Antioxid. Redox Signal. 2012, 18, 1557–1593. [Google Scholar] [CrossRef]
- Schieber, M.; Chandel, N.S. ROS Function in Redox Signaling and Oxidative Stress. Curr. Biol. 2014, 24, R453–R462. [Google Scholar] [CrossRef] [Green Version]
- Schreck, R.; Rieber, P.; Baeuerle, P.A. Reactive Oxygen Intermediates as Apparently Widely Used Messengers in the Activation of the NF-Kappa B Transcription Factor and HIV-1. EMBO J. 1991, 10, 2247–2258. [Google Scholar] [CrossRef]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.-C. NF-ΚB Signaling in Inflammation. Sig. Transduct. Target. Ther. 2017, 2, 17023. [Google Scholar] [CrossRef] [Green Version]
- Baird, L.; Dinkova-Kostova, A.T. The Cytoprotective Role of the Keap1–Nrf2 Pathway. Arch. Toxicol. 2011, 85, 241–272. [Google Scholar] [CrossRef]
- Dinkova-Kostova, A.T.; Kostov, R.V.; Canning, P. Keap1, the Cysteine-Based Mammalian Intracellular Sensor for Electrophiles and Oxidants. Arch. Biochem. Biophys. 2017, 617, 84–93. [Google Scholar] [CrossRef] [Green Version]
- Baird, L.; Yamamoto, M. The Molecular Mechanisms Regulating the KEAP1-NRF2 Pathway. Mol. Cell. Biol. 2020, 40, e00099-20. [Google Scholar] [CrossRef]
- Tonelli, C.; Chio, I.I.C.; Tuveson, D.A. Transcriptional Regulation by Nrf2. Antioxid. Redox Signal. 2017, 29, 1727–1745. [Google Scholar] [CrossRef] [Green Version]
- Aguilera-Mendoza, L.; Marrero-Ponce, Y.; Beltran, J.A.; Tellez Ibarra, R.; Guillen-Ramirez, H.A.; Brizuela, C.A. Graph-Based Data Integration from Bioactive Peptide Databases of Pharmaceutical Interest: Toward an Organized Collection Enabling Visual Network Analysis. Bioinformatics 2019, 35, 4739–4747. [Google Scholar] [CrossRef] [PubMed]
- Minkiewicz, P.; Iwaniak, A.; Darewicz, M. BIOPEP-UWM Database of Bioactive Peptides: Current Opportunities. Int. J. Mol. Sci. 2019, 20, 5978. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gentile, F.; Agrawal, V.; Hsing, M.; Ton, A.-T.; Ban, F.; Norinder, U.; Gleave, M.E.; Cherkasov, A. Deep Docking: A Deep Learning Platform for Augmentation of Structure Based Drug Discovery. ACS Cent. Sci. 2020, 6, 939–949. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Yao, K.; Repasky, M.P.; Leswing, K.; Abel, R.; Shoichet, B.K.; Jerome, S.V. Efficient Exploration of Chemical Space with Docking and Deep Learning. J. Chem. Theory Comput. 2021, 17, 7106–7119. [Google Scholar] [CrossRef]
- Torres, P.H.M.; Sodero, A.C.R.; Jofily, P.; Silva-Jr, F.P. Key Topics in Molecular Docking for Drug Design. Int. J. Mol. Sci. 2019, 20, 4574. [Google Scholar] [CrossRef] [Green Version]
- Proctor, E.A.; Yin, S.; Tropsha, A.; Dokholyan, N.V. Discrete Molecular Dynamics Distinguishes Nativelike Binding Poses from Decoys in Difficult Targets. Biophys. J. 2012, 102, 144–151. [Google Scholar] [CrossRef] [Green Version]
- Ciemny, M.; Kurcinski, M.; Kamel, K.; Kolinski, A.; Alam, N.; Schueler-Furman, O.; Kmiecik, S. Protein–Peptide Docking: Opportunities and Challenges. Drug Discov. Today 2018, 23, 1530–1537. [Google Scholar] [CrossRef]
- Kundrotas, P.J.; Zhu, Z.; Janin, J.; Vakser, I.A. Templates Are Available to Model Nearly All Complexes of Structurally Characterized Proteins. Proc. Natl. Acad. Sci. USA 2012, 109, 9438–9441. [Google Scholar] [CrossRef] [Green Version]
- Lensink, M.F.; Velankar, S.; Wodak, S.J. Modeling Protein–Protein and Protein–Peptide Complexes: CAPRI 6th Edition. Proteins Struct. Funct. Bioinform. 2017, 85, 359–377. [Google Scholar] [CrossRef]
- Marcu, O.; Dodson, E.-J.; Alam, N.; Sperber, M.; Kozakov, D.; Lensink, M.F.; Schueler-Furman, O. FlexPepDock Lessons from CAPRI Peptide–Protein Rounds and Suggested New Criteria for Assessment of Model Quality and Utility. Proteins Struct. Funct. Bioinform. 2017, 85, 445–462. [Google Scholar] [CrossRef] [Green Version]
- Pallara, C.; Jiménez-García, B.; Romero, M.; Moal, I.H.; Fernández-Recio, J. PyDock Scoring for the New Modeling Challenges in Docking: Protein–Peptide, Homo-Multimers, and Domain–Domain Interactions. Proteins Struct. Funct. Bioinform. 2017, 85, 487–496. [Google Scholar] [CrossRef] [Green Version]
- de Vries, S.J.; Rey, J.; Schindler, C.E.M.; Zacharias, M.; Tuffery, P. The PepATTRACT Web Server for Blind, Large-Scale Peptide–Protein Docking. Nucleic Acids Res. 2017, 45, W361–W364. [Google Scholar] [CrossRef]
- Ben-Shimon, A.; Niv, M.Y. AnchorDock: Blind and Flexible Anchor-Driven Peptide Docking. Structure 2015, 23, 929–940. [Google Scholar] [CrossRef] [Green Version]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the Speed and Accuracy of Docking with a New Scoring Function, Efficient Optimization, and Multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef] [Green Version]
- Ivetac, A.; McCammon, J.A. Molecular Recognition in the Case of Flexible Targets. Curr. Pharm. Des. 2011, 17, 1663–1671. [Google Scholar] [CrossRef] [Green Version]
- Huang, S.-Y.; Zou, X. Advances and Challenges in Protein-Ligand Docking. Int. J. Mol. Sci. 2010, 11, 3016–3034. [Google Scholar] [CrossRef] [Green Version]
- Pinzi, L.; Rastelli, G. Molecular Docking: Shifting Paradigms in Drug Discovery. Int. J. Mol. Sci. 2019, 20, 4331. [Google Scholar] [CrossRef] [Green Version]
- Tonolo, F.; Folda, A.; Cesaro, L.; Scalcon, V.; Marin, O.; Ferro, S.; Bindoli, A.; Rigobello, M.P. Milk-Derived Bioactive Peptides Exhibit Antioxidant Activity through the Keap1-Nrf2 Signaling Pathway. J. Funct. Food 2019, 64, 103696. [Google Scholar] [CrossRef]
- Wang, N.; Wang, W.; Sadiq, F.A.; Wang, S.; Caiqin, L.; Jianchang, J. Involvement of Nrf2 and Keap1 in the Activation of Antioxidant Responsive Element (ARE) by Chemopreventive Agent Peptides from Soft-Shelled Turtle. Process Biochem. 2020, 92, 174–181. [Google Scholar] [CrossRef]
- Tonolo, F.; Moretto, L.; Grinzato, A.; Fiorese, F.; Folda, A.; Scalcon, V.; Ferro, S.; Arrigoni, G.; Bellamio, M.; Feller, E.; et al. Fermented Soy-Derived Bioactive Peptides Selected by a Molecular Docking Approach Show Antioxidant Properties Involving the Keap1/Nrf2 Pathway. Antioxidants 2020, 9, 1306. [Google Scholar] [CrossRef]
- Tonolo, F.; Fiorese, F.; Moretto, L.; Folda, A.; Scalcon, V.; Grinzato, A.; Ferro, S.; Arrigoni, G.; Bindoli, A.; Feller, E.; et al. Identification of New Peptides from Fermented Milk Showing Antioxidant Properties: Mechanism of Action. Antioxidants 2020, 9, 117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Zhang, D.; Hannink, M.; Beamer, L.J. Crystal Structure of the Kelch Domain of Human Keap1. J. Biol. Chem. 2004, 279, 54750–54758. [Google Scholar] [CrossRef] [Green Version]
- Tong, K.I.; Katoh, Y.; Kusunoki, H.; Itoh, K.; Tanaka, T.; Yamamoto, M. Keap1 Recruits Neh2 through Binding to ETGE and DLG Motifs: Characterization of the Two-Site Molecular Recognition Model. Mol. Cell Biol. 2006, 26, 2887–2900. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crisman, E.; Duarte, P.; Dauden, E.; Cuadrado, A.; Rodríguez-Franco, M.I.; López, M.G.; León, R. KEAP1-NRF2 Protein–Protein Interaction Inhibitors: Design, Pharmacological Properties and Therapeutic Potential. Med. Res. Rev. 2023, 43, 237–287. [Google Scholar] [CrossRef] [PubMed]
- Mirdamadi, S.; Mirzaei, M.; Soleymanzadeh, N.; Safavi, M.; Bakhtiari, N.; Zandi, M. Antioxidant and Cytoprotective Effects of Synthetic Peptides Identified from Kluyveromyces Marxianus Protein Hydrolysate: Insight into the Molecular Mechanism. LWT 2021, 148, 111792. [Google Scholar] [CrossRef]
- Zhang, J.; Li, M.; Zhang, G.; Tian, Y.; Kong, F.; Xiong, S.; Zhao, S.; Jia, D.; Manyande, A.; Du, H. Identification of Novel Antioxidant Peptides from Snakehead (Channa argus) Soup Generated during Gastrointestinal Digestion and Insights into the Anti-Oxidation Mechanisms. Food Chem. 2021, 337, 127921. [Google Scholar] [CrossRef]
- Zhang, J.; Wu, S.; Wang, Q.; Yuan, Q.; Li, Y.; Reboredo-Rodríguez, P.; Varela-López, A.; He, Z.; Wu, F.; Hu, H.; et al. Oxidative Stress Amelioration of Novel Peptides Extracted from Enzymatic Hydrolysates of Chinese Pecan Cake. Int. J. Mol. Sci. 2022, 23, 12086. [Google Scholar] [CrossRef]
- Ren, L.-K.; Yang, Y.; Ma, C.-M.; Fan, J.; Bian, X.; Liu, B.-X.; Wang, D.-F.; Zhu, P.-Y.; Fu, Y.; Zhang, N. Identification and in Silico Analysis of Novel Antioxidant Peptides in Broken Rice Protein Hydrolysate and Its Cytoprotective Effect against H2O2-Induced 2BS Cell Model. Food Res. Int. 2022, 162, 112108. [Google Scholar] [CrossRef]
- Wen, C.; Zhang, J.; Zhang, H.; Duan, Y.; Ma, H. Study on the Structure–Activity Relationship of Watermelon Seed Antioxidant Peptides by Using Molecular Simulations. Food Chem. 2021, 364, 130432. [Google Scholar] [CrossRef]
- Huang, P.; Miao, J.; Li, J.; Li, Y.; Wang, X.; Yu, Y.; Cao, Y. Novel Antioxidant Peptides from Pearl Shell Meat Hydrolysate and Their Antioxidant Activity Mechanism. Molecules 2023, 28, 864. [Google Scholar] [CrossRef]
- Xing, L.; Fu, L.; Hao, Y.; Zhang, W. Dry-Cured Ham-Derived Peptide (Asp–Leu–Glu–Glu) Exerts Cytoprotective Capacity in Human Intestinal Epithelial Caco-2 Cells. Antioxidants 2021, 10, 1354. [Google Scholar] [CrossRef]
- Zhu, L.; Xiong, H.; Huang, X.; Guyonnet, V.; Ma, M.; Chen, X.; Zheng, Y.; Wang, L.; Hu, G. Identification and Molecular Mechanisms of Novel Antioxidant Peptides from Two Sources of Eggshell Membrane Hydrolysates Showing Cytoprotection against Oxidative Stress: A Combined in Silico and in Vitro Study. Food Res. Int. 2022, 157, 111266. [Google Scholar] [CrossRef]
- Wang, L.; Ma, M.; Yu, Z.; Du, S. Preparation and Identification of Antioxidant Peptides from Cottonseed Proteins. Food Chem. 2021, 352, 129399. [Google Scholar] [CrossRef] [PubMed]
- Yin, J.-Y.; Han, Y.-N.; Liu, M.-Q.; Piao, Z.-H.; Zhang, X.; Xue, Y.-T.; Zhang, Y.-H. Structure-Guided Discovery of Antioxidant Peptides Bounded to the Keap1 Receptor as Hunter for Potential Dietary Antioxidants. Food Chem. 2022, 373, 130999. [Google Scholar] [CrossRef]
- Jiang, Y.; Wang, R.; Yin, Z.; Sun, J.; Wang, B.; Zhao, D.; Zeng, X.; Li, H.; Huang, M.; Sun, B. Optimization of Jiuzao Protein Hydrolysis Conditions and Antioxidant Activity in Vivo of Jiuzao Tetrapeptide Asp-Arg-Glu-Leu by Elevating the Nrf2/Keap1-P38/PI3K-MafK Signaling Pathway. Food Funct. 2021, 12, 4808–4824. [Google Scholar] [CrossRef]
- Chai, T.-T.; Koh, J.-A.; Wong, C.C.-C.; Sabri, M.Z.; Wong, F.-C. Computational Screening for the Anticancer Potential of Seed-Derived Antioxidant Peptides: A Cheminformatic Approach. Molecules 2021, 26, 7396. [Google Scholar] [CrossRef]
- Wang, Q.; Yang, Z.; Zhuang, J.; Zhang, J.; Shen, F.; Yu, P.; Zhong, H.; Feng, F. Antiaging Function of Chinese Pond Turtle (Chinemys reevesii) Peptide through Activation of the Nrf2/Keap1 Signaling Pathway and Its Structure-Activity Relationship. Front. Nutr. 2022, 9, 961922. [Google Scholar] [CrossRef]
- Han, J.; Huang, Z.; Tang, S.; Lu, C.; Wan, H.; Zhou, J.; Li, Y.; Ming, T.; Jim Wang, Z.; Su, X. The Novel Peptides ICRD and LCGEC Screened from Tuna Roe Show Antioxidative Activity via Keap1/Nrf2-ARE Pathway Regulation and Gut Microbiota Modulation. Food Chem. 2020, 327, 127094. [Google Scholar] [CrossRef]
- Yang, W.; Huang, Z.; Xiong, H.; Wang, J.; Zhang, H.; Guo, F.; Wang, C.; Sun, Y. Rice Protein Peptides Alleviate Dextran Sulfate Sodium-Induced Colitis via the Keap1–Nrf2 Signaling Pathway and Regulating Gut Microbiota. J. Agric. Food Chem. 2022, 70, 12469–12483. [Google Scholar] [CrossRef]
- Wang, S.; Su, G.; Zhang, X.; Song, G.; Zhang, L.; Zheng, L.; Zhao, M. Characterization and Exploration of Potential Neuro-protective Peptides in Walnut (Juglans regia) Protein Hydrolysate against Cholinergic System Damage and Oxidative Stress in Scopolamine-Induced Cognitive and Memory Impairment Mice and Zebrafish. J. Agric. Food Chem. 2021, 69, 2773–2783. [Google Scholar] [CrossRef]
- Wei, G.; Zhao, Q.; Wang, D.; Fan, Y.; Shi, Y.; Huang, A. Novel ACE Inhibitory, Antioxidant and α-Glucosidase Inhibitory Peptides Identified from Fermented Rubing Cheese through Peptidomic and Molecular Docking. LWT 2022, 159, 113196. [Google Scholar] [CrossRef]
- Li, J.; Lu, J.; Asakiya, C.; Huang, K.; Zhou, X.; Liu, Q.; He, X. Extraction and Identification of Three New Urechis Unicinctus Visceral Peptides and Their Antioxidant Activity. Mar. Drugs 2022, 20, 293. [Google Scholar] [CrossRef] [PubMed]
- Ong, J.-H.; Koh, J.-A.; Cao, H.; Tan, S.-A.; Abd Manan, F.; Wong, F.-C.; Chai, T.-T. Purification, Identification and Charac-terization of Antioxidant Peptides from Corn Silk Tryptic Hydrolysate: An Integrated In Vitro-In Silico Approach. Antioxidants 2021, 10, 1822. [Google Scholar] [CrossRef] [PubMed]
- Tong, K.I.; Padmanabhan, B.; Kobayashi, A.; Shang, C.; Hirotsu, Y.; Yokoyama, S.; Yamamoto, M. Different Electrostatic Potentials Define ETGE and DLG Motifs as Hinge and Latch in Oxidative Stress Response. Mol. Cell. Biol. 2007, 27, 7511–7521. [Google Scholar] [CrossRef] [Green Version]
- Beamer, L.J.; Li, X.; Bottoms, C.A.; Hannink, M. Conserved Solvent and Side-Chain Interactions in the 1.35 Å Structure of the Kelch Domain of Keap1. Acta Crystallogr. Sect. D Biol. Crystallogr. 2005, 61, 1335–1342. [Google Scholar] [CrossRef] [Green Version]
- Padmanabhan, B.; Tong, K.I.; Ohta, T.; Nakamura, Y.; Scharlock, M.; Ohtsuji, M.; Kang, M.-I.; Kobayashi, A.; Yokoyama, S.; Yamamoto, M. Structural Basis for Defects of Keap1 Activity Provoked by Its Point Mutations in Lung Cancer. Mol. Cell 2006, 21, 689–700. [Google Scholar] [CrossRef]
- Lo, S.-C.; Li, X.; Henzl, M.T.; Beamer, L.J.; Hannink, M. Structure of the Keap1:Nrf2 Interface Provides Mechanistic Insight into Nrf2 Signaling. EMBO J. 2006, 25, 3605–3617. [Google Scholar] [CrossRef] [Green Version]
- Fukutomi, T.; Takagi, K.; Mizushima, T.; Ohuchi, N.; Yamamoto, M. Kinetic, Thermodynamic, and Structural Characterizations of the Association between Nrf2-DLGex Degron and Keap1. Mol. Cell. Biol. 2014, 34, 832–846. [Google Scholar] [CrossRef] [Green Version]
- Wen, C.; Zhang, J.; Zhang, H.; Duan, Y.; Ma, H. Plant Protein-Derived Antioxidant Peptides: Isolation, Identification, Mechanism of Action and Application in Food Systems: A Review. Trends Food Sci. Technol. 2020, 105, 308–322. [Google Scholar] [CrossRef]
- Tonolo, F.; Sandre, M.; Ferro, S.; Folda, A.; Scalcon, V.; Scutari, G.; Feller, E.; Marin, O.; Bindoli, A.; Rigobello, M.P. Milk-Derived Bioactive Peptides Protect against Oxidative Stress in a Caco-2 Cell Model. Food Funct. 2018, 9, 1245–1253. [Google Scholar] [CrossRef]
- Tonolo, F.; Folda, A.; Scalcon, V.; Marin, O.; Bindoli, A.; Rigobello, M.P. Nrf2-Activating Bioactive Peptides Exert An-ti-Inflammatory Activity through Inhibition of the NF-ΚB Pathway. Int. J. Mol. Sci. 2022, 23, 4382. [Google Scholar] [CrossRef]
- Suo, S.-K.; Zheng, S.-L.; Chi, C.-F.; Luo, H.-Y.; Wang, B. Novel Angiotensin-Converting Enzyme Inhibitory Peptides from Tuna Byproducts-Milts: Preparation, Characterization, Molecular Docking Study, and Antioxidant Function on H2O2-Damaged Human Umbilical Vein Endothelial Cells. Front. Nutr. 2022, 9, 957778. [Google Scholar] [CrossRef]
- Suo, S.-K.; Zhao, Y.-Q.; Wang, Y.-M.; Pan, X.-Y.; Chi, C.-F.; Wang, B. Seventeen Novel Angiotensin Converting Enzyme (ACE) Inhibitory Peptides from the Protein Hydrolysate of Mytilus Edulis: Isolation, Identification, Molecular Docking Study, and Protective Function on HUVECs. Food Funct. 2022, 13, 7831–7846. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tonolo, F.; Grinzato, A.; Bindoli, A.; Rigobello, M.P. From In Silico to a Cellular Model: Molecular Docking Approach to Evaluate Antioxidant Bioactive Peptides. Antioxidants 2023, 12, 665. https://doi.org/10.3390/antiox12030665
Tonolo F, Grinzato A, Bindoli A, Rigobello MP. From In Silico to a Cellular Model: Molecular Docking Approach to Evaluate Antioxidant Bioactive Peptides. Antioxidants. 2023; 12(3):665. https://doi.org/10.3390/antiox12030665
Chicago/Turabian StyleTonolo, Federica, Alessandro Grinzato, Alberto Bindoli, and Maria Pia Rigobello. 2023. "From In Silico to a Cellular Model: Molecular Docking Approach to Evaluate Antioxidant Bioactive Peptides" Antioxidants 12, no. 3: 665. https://doi.org/10.3390/antiox12030665