
Accumulation of Proline in Plants under Contaminated Soils—Are We on the Same Page?
 , , and
, , and 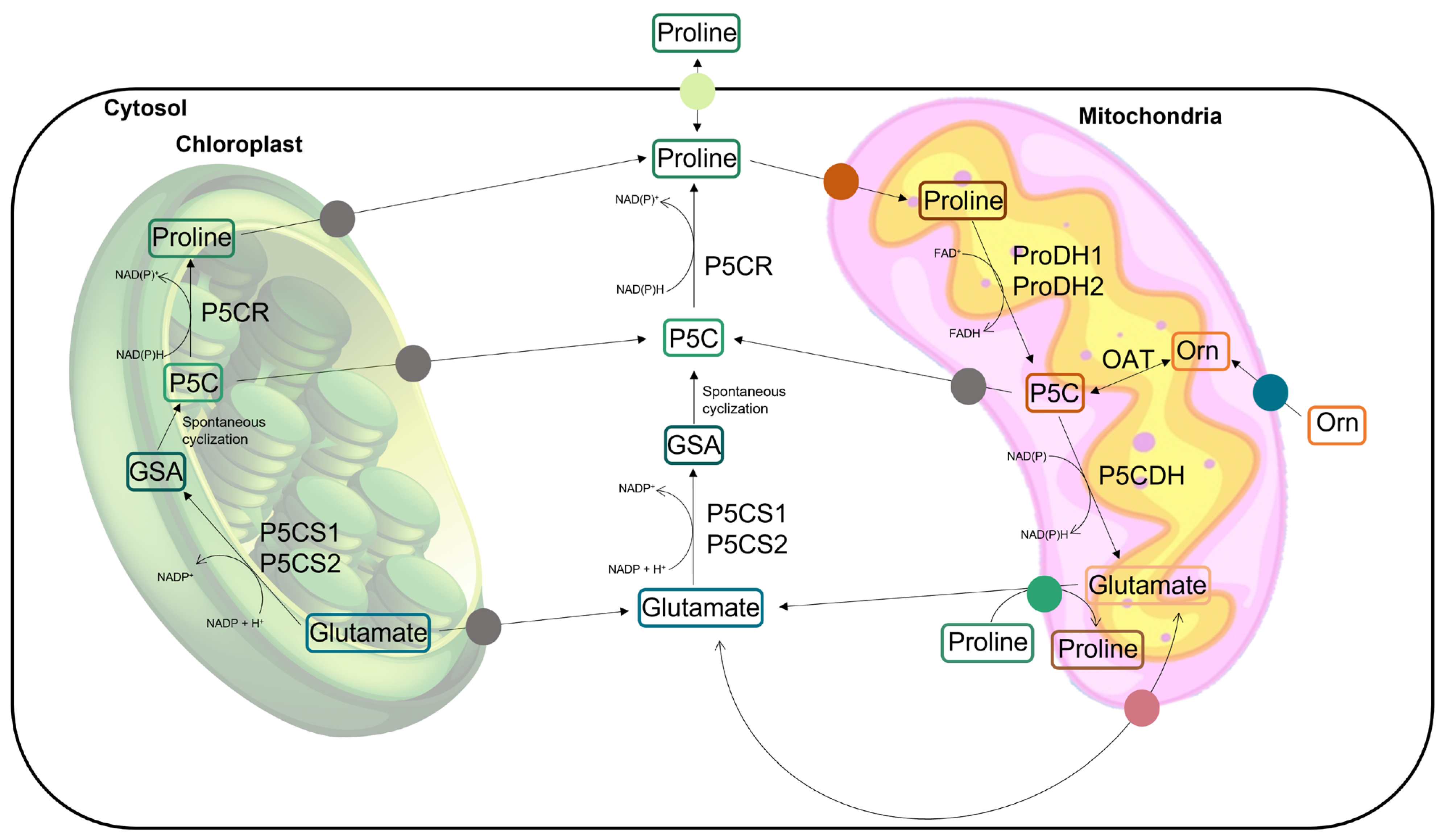{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Proline: Functions and Metabolism
3. How Is Proline Homeostasis Affected by Stress?
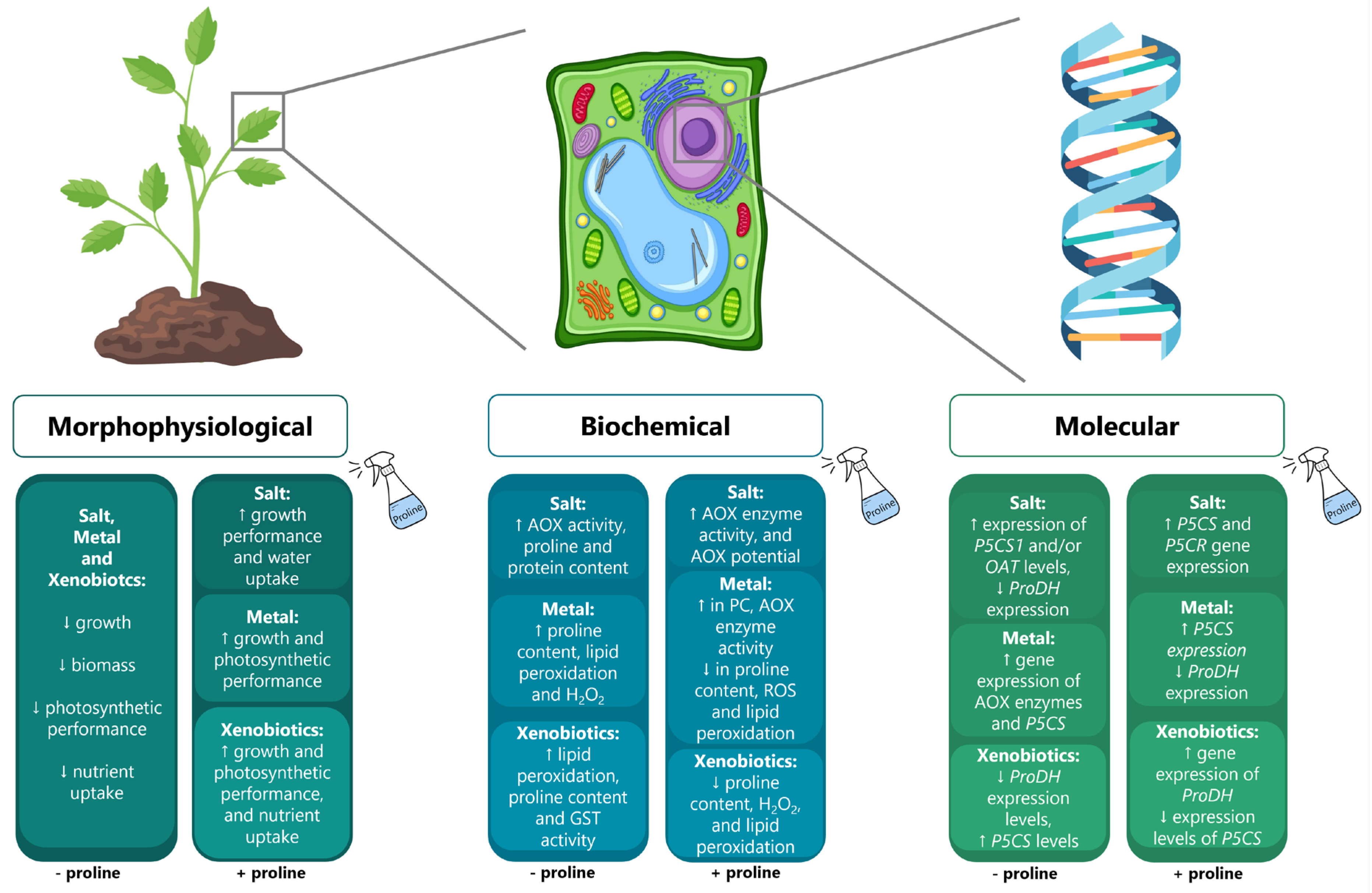
4. The Involvement of Proline in Plants Facing Issues of Soil Degradation
4.1. Salinity Stress
4.1.1. Role of Proline in Salt-Stressed Plants
4.1.2. Proline Functions in Salt-Stressed Plants—Increased Tolerance or Higher Susceptibility?
4.1.3. Exogenous Application of Proline to Mitigate Salt Stress
4.1.4. Modulation of Endogenous Proline Levels in Response to Salt Stress—Genetic Engineering Approaches
4.2. Metal-Induced Phytotoxicity
4.2.1. Proline Homeostasis in Response to Metal Stress
4.2.2. Exogenous Application of Proline to Mitigate Metal-Induced Stress
4.3. Exposure to Xenobiotics
4.3.1. Xenobiotic Exposure and Proline Metabolism—Is There a Connection?
4.3.2. Impacts of Pharmaceuticals-Induced Stress on Proline Endogenous Levels
4.3.3. Impacts of Pesticide-Induced Stress on Proline Endogenous Levels
4.3.4. Xenobiotic Exposure vs. Mitigation Strategies—Is There Room for Proline Action?
5. Concluding Remarks and Future Perspectives
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Korres, N.E.; Norsworthy, J.K.; Tehranchian, P.; Gitsopoulos, T.K.; Loka, D.A.; Oosterhuis, D.M.; Gealy, D.R.; Moss, S.R.; Burgos, N.R.; Miller, M.R.; et al. Cultivars to Face Climate Change Effects on Crops and Weeds: A Review. Agron. Sustain. Dev. 2016, 36, 12. [Google Scholar] [CrossRef] [Green Version]
- Becklin, K.M.; Anderson, J.T.; Gerhart, L.M.; Wadgymar, S.M.; Wessinger, C.A.; Ward, J.K. Examining Plant Physiological Responses to Climate Change through an Evolutionary Lens. Plant Physiol. 2016, 172, 635–649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- FAO. FAO Strategy on Climate Change; FAO: Rome, Italy, 2017. [Google Scholar]
- FAO. FAO’s Work on Climate Change. In Proceedings of the United Nations Climate Change Conference 2018, Food and Agriculture Organization of the United Nations, Katowice, Poland, 2–14 December 2018. [Google Scholar]
- Prăvălie, R.; Patriche, C.; Borrelli, P.; Panagos, P.; Roșca, B.; Dumitraşcu, M.; Nita, I.A.; Săvulescu, I.; Birsan, M.V.; Bandoc, G. Arable Lands under the Pressure of Multiple Land Degradation Processes. A Global Perspective. Environ. Res. 2021, 194, 110697. [Google Scholar] [CrossRef] [PubMed]
- Mukhopadhyay, R.; Sarkar, B.; Jat, H.S.; Sharma, P.C.; Bolan, N.S. Soil Salinity under Climate Change: Challenges for Sustainable Agriculture and Food Security. J. Environ. Manag. 2021, 280, 111736. [Google Scholar] [CrossRef] [PubMed]
- Talukder, B.; Ganguli, N.; Matthew, R.; van Loon, G.W.; Hipel, K.W.; Orbinski, J. Climate Change-Triggered Land Degradation and Planetary Health: A Review. Land Degrad. Dev. 2021, 32, 4509–4522. [Google Scholar] [CrossRef]
- Nguyen, H.C.; Lin, K.H.; Ho, S.L.; Chiang, C.M.; Yang, C.M. Enhancing the Abiotic Stress Tolerance of Plants: From Chemical Treatment to Biotechnological Approaches. Physiol. Plant 2018, 164, 452–466. [Google Scholar] [CrossRef]
- Hossain, A.; Skalicky, M.; Brestic, M.; Maitra, S.; Alam, M.A.; Syed, M.A.; Hossain, J.; Sarkar, S.; Saha, S.; Bhadra, P.; et al. Consequences and Mitigation Strategies of Abiotic Stresses in Wheat (Triticum aestivum L.) under the Changing Climate. Agronomy 2021, 11, 241. [Google Scholar] [CrossRef]
- Hosseinifard, M.; Stefaniak, S.; Javid, M.G.; Soltani, E.; Wojtyla, Ł.; Garnczarska, M. Contribution of Exogenous Proline to Abiotic Stresses Tolerance in Plants: A Review. Int. J. Mol. Sci. 2022, 23, 5186. [Google Scholar] [CrossRef]
- Fleming, T.R.; Fleming, C.C.; Levy, C.C.B.; Repiso, C.; Hennequart, F.; Nolasco, J.B.; Liu, F. Biostimulants Enhance Growth and Drought Tolerance in Arabidopsis thaliana and Exhibit Chemical Priming Action. Ann. Appl. Biol. 2019, 174, 153–165. [Google Scholar] [CrossRef]
- Hayat, S.; Hayat, Q.; Alyemeni, M.N.; Wani, A.S.; Pichtel, J.; Ahmad, A. Role of Proline under Changing Environments: A Review. Plant Signal. Behav. 2012, 7, 1456–1466. [Google Scholar] [CrossRef] [Green Version]
- Dar, M.I.; Naikoo, M.I.; Rehman, F.; Naushin, F.; Khan, F.A. Proline Accumulation in Plants: Roles in Stress Tolerance and Plant Development. In Osmolytes and Plants Acclimation to Changing Environment: Emerging Omics Technologies; Springer: New Delhi, India, 2015; pp. 155–166. ISBN 9788132226161. [Google Scholar]
- Aslam, M. Specific Role of Proline Against Heavy Metals Toxicity in Plants. Int. J. Pure Appl. Biosci. 2017, 5, 27–34. [Google Scholar] [CrossRef]
- Ghosh, U.K.; Islam, M.N.; Siddiqui, M.N.; Cao, X.; Khan, M.A.R. Proline, a Multifaceted Signalling Molecule in Plant Responses to Abiotic Stress: Understanding the Physiological Mechanisms. Plant Biol. 2022, 24, 227–239. [Google Scholar] [CrossRef]
- Sharma, S.S.; Dietz, K. The Significance of Amino Acids and Amino Acid-Derived Molecules in Plant Responses and Adaptation to Heavy Metal Stress. J. Exp. Bot. 2006, 57, 711–726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- EL-Bauome, H.A.; Abdeldaym, E.A.; Abd El-Hady, M.A.M.; Darwish, D.B.E.; Alsubeie, M.S.; El-Mogy, M.M.; Basahi, M.A.; Al-Qahtani, S.M.; Al-Harbi, N.A.; Alzuaibr, F.M.; et al. Exogenous Proline, Methionine, and Melatonin Stimulate Growth, Quality, and Drought Tolerance in Cauliflower Plants. Agriculture 2022, 12, 1301. [Google Scholar] [CrossRef]
- Verslues, P.E.; Sharma, S. Proline Metabolism and Its Implications for Plant-Environment Interaction. Arab. Book 2010, 8, e0140. [Google Scholar] [CrossRef] [Green Version]
- Szabados, L.; Savouré, A. Proline: A Multifunctional Amino Acid. Trends Plant Sci. 2010, 15, 89–97. [Google Scholar] [CrossRef]
- Alvarez, M.E.; Savouré, A.; Szabados, L. Proline Metabolism as Regulatory Hub. Trends Plant Sci. 2022, 27, 39–55. [Google Scholar] [CrossRef]
- Williamson, M.P. The Structure and Function of Proline-Rich Regions in Proteins. Biochem. J. 1994, 297, 249. [Google Scholar] [CrossRef] [Green Version]
- Mattioli, R.; Costantino, P.; Trovato, M. Proline Accumulation in Plants: Not Only Stress. Plant Signal. Behav. 2009, 4, 1016–1018. [Google Scholar] [CrossRef]
- Funck, D.; Baumgarten, L.; Stift, M.; von Wirén, N.; Schönemann, L. Differential Contribution of P5CS Isoforms to Stress Tolerance in Arabidopsis. Front. Plant Sci. 2020, 11, 1483. [Google Scholar] [CrossRef]
- Meena, M.; Divyanshu, K.; Kumar, S.; Swapnil, P.; Zehra, A.; Shukla, V.; Yadav, M.; Upadhyay, R.S. Regulation of L-Proline Biosynthesis, Signal Transduction, Transport, Accumulation and Its Vital Role in Plants during Variable Environmental Conditions. Heliyon 2019, 5, e02952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ribarits, A.; Abdullaev, A.; Tashpulatov, A.; Richter, A.; Heberle-Bors, E.; Touraev, A. Two Tobacco Proline Dehydrogenases Are Differentially Regulated and Play a Role in Early Plant Development. Planta 2007, 225, 1313–1324. [Google Scholar] [CrossRef] [PubMed]
- Szepesi, Á.; Szőllősi, R. Mechanism of Proline Biosynthesis and Role of Proline Metabolism Enzymes Under Environmental Stress in Plants. In Plant Metabolites and Regulation under Environmental Stress; Academic Press: Cambridge, MA, USA, 2018; pp. 337–353. ISBN 9780128126899. [Google Scholar]
- Kavi Kishor, P.B.; Sreenivasulu, N. Is Proline Accumulation per Se Correlated with Stress Tolerance or Is Proline Homeostasis a More Critical Issue? Plant Cell Environ. 2014, 37, 300–311. [Google Scholar] [CrossRef] [PubMed]
- Rejeb, K.B.; Abdelly, C.; Savouré, A. How Reactive Oxygen Species and Proline Face Stress Together. Plant Physiol. Biochem. 2014, 80, 278–284. [Google Scholar] [CrossRef]
- Heuer, B. Role of Proline in Plant Response to Drought and Salinity. In Handbook of Plant and Crop Stress, 3rd ed.; CRC Press: Boca Raton, FL, USA, 2016; pp. 213–238. ISBN 9781439813997. [Google Scholar]
- Verbruggen, N.; Hermans, C. Proline Accumulation in Plants: A Review. Amino Acids 2008, 35, 753–759. [Google Scholar] [CrossRef]
- Liang, X.; Zhang, L.; Natarajan, S.K.; Becker, D.F. Proline Mechanisms of Stress Survival. Antioxid. Redox Signal. 2013, 19, 998–1011. [Google Scholar] [CrossRef] [Green Version]
- Kavi Kishor, P.B.; Kumari, P.H.; Sunita, M.S.L.; Sreenivasulu, N. Role of Proline in Cell Wall Synthesis and Plant Development and Its Implications in Plant Ontogeny. Front. Plant Sci. 2015, 6, 544. [Google Scholar] [CrossRef] [Green Version]
- Hare, P.D.; Cress, W.A. Metabolic Implications of Stress-Induced Proline Accumulation in Plants; Kluwer Academic Publishers: Alphen am Rhein, The Netherlands, 1997; Volume 21. [Google Scholar]
- Kaur, G.; Asthir, B. Proline: A Key Player in Plant Abiotic Stress Tolerance. Biol. Plant 2015, 59, 609–619. [Google Scholar] [CrossRef]
- Shahid, S.A.; Zaman, M.; Heng, L. Soil Salinity: Historical Perspectives and a World Overview of the Problem. Guidel. Salin. Assess. Mitig. Adapt. Using Nucl. Relat. Tech. 2018, 2018, 43–53. [Google Scholar] [CrossRef] [Green Version]
- Hossain, M.S. Present Scenario of Global Salt Affected Soils, Its Management and Importance of Salinity Research. Int. Res. J. Biol. Sci. 2019, 1, 1–3. Available online: https://www.researchgate.net/profile/Md-Hossain-72/publication/334773002_Present_Scenario_of_Global_Salt_Affected_Soils_its_Management_and_Importance_of_Salinity_Research/links/5d7150a292851cacdb23c8d9/Present-Scenario-of-Global-Salt-Affected-Soils-its-Management-and-Importance-of-Salinity-Research.pdf (accessed on 2 February 2023).
- Parihar, P.; Singh, S.; Singh, R.; Singh, V.P.; Prasad, S.M. Effect of Salinity Stress on Plants and Its Tolerance Strategies: A Review. Environ. Sci. Pollut. Res. 2015, 22, 4056–4075. [Google Scholar] [CrossRef] [PubMed]
- Ami, K.; Planchais, S.; Cabassa, C.; Guivarc’h, A.; Very, A.A.; Khelifi, M.; Djebbar, R.; Abrous-Belbachir, O.; Carol, P. Different Proline Responses of Two Algerian Durum Wheat Cultivars to in Vitro Salt Stress. Acta Physiol. Plant 2020, 42, 21. [Google Scholar] [CrossRef]
- ben Ahmed, C.; ben Rouina, B.; Sensoy, S.; Boukhriss, M.; ben Abdullah, F. Saline Water Irrigation Effects on Antioxidant Defense System and Proline Accumulation in Leaves and Roots of Field-Grown Olive. J. Agric. Food Chem. 2009, 57, 11484–11490. [Google Scholar] [CrossRef]
- Hnilickova, H.; Kraus, K.; Vachova, P.; Hnilicka, F. Salinity Stress Affects Photosynthesis, Malondialdehyde Formation, and Proline Content in Portulaca oleracea L. Plants 2021, 10, 845. [Google Scholar] [CrossRef]
- Izadi, M.H.; Rabbani, J.; Emam, Y.; Pessarakli, M.; Tahmasebi, A. Effects of Salinity Stress on Physiological Performance of Various Wheat and Barley Cultivars. J. Plant Nutr. 2014, 37, 520–531. [Google Scholar] [CrossRef]
- Moghaddam, M.; Farhadi, N.; Panjtandoust, M.; Ghanati, F. Seed Germination, Antioxidant Enzymes Activity and Proline Content in Medicinal Plant Tagetes minuta under Salinity Stress. Plant Biosyst. Int. J. Deal. All Asp. Plant Biol. 2019, 154, 835–842. [Google Scholar] [CrossRef]
- Shin, Y.K.; Bhandari, S.R.; Cho, M.C.; Lee, J.G. Evaluation of Chlorophyll Fluorescence Parameters and Proline Content in Tomato Seedlings Grown under Different Salt Stress Conditions. Hortic. Environ. Biotechnol. 2020, 61, 433–443. [Google Scholar] [CrossRef]
- Sousa, B.; Rodrigues, F.; Soares, C.; Martins, M.; Azenha, M.; Lino-Neto, T.; Santos, C.; Cunha, A.; Fidalgo, F. Impact of Combined Heat and Salt Stresses on Tomato Plants—Insights into Nutrient Uptake and Redox Homeostasis. Antioxidants 2022, 11, 478. [Google Scholar] [CrossRef] [PubMed]
- de la Torre-González, A.; Montesinos-Pereira, D.; Blasco, B.; Ruiz, J.M. Influence of the Proline Metabolism and Glycine Betaine on Tolerance to Salt Stress in Tomato (Solanum lycopersicum L.) Commercial Genotypes. J. Plant Physiol. 2018, 231, 329–336. [Google Scholar] [CrossRef]
- Goharrizi, K.J.; Baghizadeh, A.; Afroushteh, M.; Amirmahani, F.; Kermani, S.G. Effects of Salinity Stress on Proline Content and Expression of Δ1-Pyrroline-5-Carboxylate Synthase and Vacuolar-Type H+ Subunit E Genes in Wheat. Plant Genet. Resour. 2020, 18, 334–342. [Google Scholar] [CrossRef]
- Arabbeigi, M.; Arzani, A.; Majidi, M.M. Expression Profiles of P5CS and DREB2 Genes under Salt Stress in Aegilops cylindrica. Russ. J. Plant Physiol. 2019, 66, 583–590. [Google Scholar] [CrossRef]
- Singh, M.; Singh, V.P.; Prasad, S.M. Responses of Photosynthesis, Nitrogen and Proline Metabolism to Salinity Stress in Solanum lycopersicum under Different Levels of Nitrogen Supplementation. Plant Physiol. Biochem. 2016, 109, 72–83. [Google Scholar] [CrossRef]
- Singh, M.; Singh, V.P.; Prasad, S.M. Nitrogen Modifies NaCl Toxicity in Eggplant Seedlings: Assessment of Chlorophyll a Fluorescence, Antioxidative Response and Proline Metabolism. Biocatal. Agric. Biotechnol. 2016, 7, 76–86. [Google Scholar] [CrossRef]
- Zhu, Y.; Jiang, X.; Zhang, J.; He, Y.; Zhu, X.; Zhou, X.; Gong, H.; Yin, J.; Liu, Y. Silicon Confers Cucumber Resistance to Salinity Stress through Regulation of Proline and Cytokinins. Plant Physiol. Biochem. 2020, 156, 209–220. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Tang, X.; Wang, H.; Shao, H.B. Proline Accumulation and Metabolism-Related Genes Expression Profiles in Kosteletzkya virginica Seedlings under Salt Stress. Front. Plant Sci. 2015, 6, 792. [Google Scholar] [CrossRef] [Green Version]
- Yusuf, M.; Fariduddin, Q.; Khan, T.A.; Hayat, S. Epibrassinolide Reverses the Stress Generated by Combination of Excess Aluminum and Salt in Two Wheat Cultivars through Altered Proline Metabolism and Antioxidants. S. Afr. J. Bot. 2017, 112, 391–398. [Google Scholar] [CrossRef]
- Wang, K.; Liu, Y.; Dong, K.; Dong, J.; Kang, J.; Yang, Q.; Zhou, H.; Sun, Y. The Effect of NaCl on Proline Metabolism in Saussurea amara Seedlings. Afr. J. Biotechnol. 2011, 10, 2886–2893. [Google Scholar] [CrossRef]
- Bagdi, D.L.; Shaw, B.B.; Purohit, G.K. Real Time PCR Expression Analysis of Gene Encoding P5CS Enzyme and Proline Metabolism under NaCl Salinity in Rice. J. Environ. Biol. 2015, 36, 955–961. [Google Scholar]
- Xue, X.; Liu, A.; Hua, X. Proline Accumulation and Transcriptional Regulation of Proline Biothesynthesis and Degradation in Brassica napus. BMB Rep. 2009, 42, 28–34. [Google Scholar] [CrossRef] [Green Version]
- Naliwajski, M.; Skłodowska, M. The Relationship between the Antioxidant System and Proline Metabolism in the Leaves of Cucumber Plants Acclimated to Salt Stress. Cells 2021, 10, 609. [Google Scholar] [CrossRef]
- Mansour, M.M.F.; Ali, E.F. Evaluation of Proline Functions in Saline Conditions. Phytochemistry 2017, 140, 52–68. [Google Scholar] [CrossRef] [PubMed]
- Forlani, G.; Bertazzini, M.; Cagnano, G. Stress-Driven Increase in Proline Levels, and Not Proline Levels Themselves, Correlates with the Ability to Withstand Excess Salt in a Group of 17 Italian Rice Genotypes. Plant Biol. 2019, 21, 336–342. [Google Scholar] [CrossRef] [PubMed]
- Gharsallah, C.; Fakhfakh, H.; Grubb, D.; Gorsane, F. Effect of Salt Stress on Ion Concentration, Proline Content, Antioxidant Enzyme Activities and Gene Expression in Tomato Cultivars. AoB Plants 2016, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ashrafi, E.; Razmjoo, J.; Zahedi, M.; Pessarakli, M. Selecting Alfalfa Cultivars for Salt Tolerance Based on Some Physiochemical Traits. Agron. J. 2014, 106, 1758–1764. [Google Scholar] [CrossRef]
- Badran, A.E.; Elsherebeny, E.A.M.; Salama, Y.A. Performance of Some Alfalfa Cultivars under Salinity Stress Conditions. J. Agric. Sci. 2015, 7, 281. [Google Scholar] [CrossRef]
- Wang, X.S.; Han, J.G. Changes of Proline Content, Activity, and Active Isoforms of Antioxidative Enzymes in Two Alfalfa Cultivars Under Salt Stress. Agric. Sci. China 2009, 8, 431–440. [Google Scholar] [CrossRef]
- Carpici, E.B.; Celik, N.; Bayram, G. The Effects of Salt Stress on the Growth, Biochemical Parameter and Mineral Element Content of Some Maize (Zea mays L.) Cultivars. Afr. J. Biotechnol. 2010, 9, 6937–6942. [Google Scholar] [CrossRef]
- Ghoulam, C.; Foursy, A.; Fares, K. Effects of Salt Stress on Growth, Inorganic Ions and Proline Accumulation in Relation to Osmotic Adjustment in Five Sugar Beet Cultivars. Environ. Exp. Bot. 2002, 47, 39–50. [Google Scholar] [CrossRef]
- Wu, G.Q.; Liang, N.; Feng, R.J.; Zhang, J.J. Evaluation of Salinity Tolerance in Seedlings of Sugar Beet (Beta vulgaris L.) Cultivars Using Proline, Soluble Sugars and Cation Accumulation Criteria. Acta Physiol. Plant 2013, 35, 2665–2674. [Google Scholar] [CrossRef]
- Poustini, K.; Siosemardeh, A.; Ranjbar, M. Proline Accumulation as a Response to Salt Stress in 30 Wheat (Triticum aestivum L.) Cultivars Differing in Salt Tolerance. Genet. Resour. Crop Evol. 2007, 54, 925–934. [Google Scholar] [CrossRef]
- Kanawapee, N.; Sanitchon, J.; Lontom, W.; Threerakulpisut, P. Evaluation of Salt Tolerance at the Seedling Stage in Rice Genotypes by Growth Performance, Ion Accumulation, Proline and Chlorophyll Content. Plant Soil 2012, 358, 235–249. [Google Scholar] [CrossRef]
- Lutts, S.; Kinet, J.M.; Bouharmont, J. Effects of Various Salts and of Mannitol on Ion and Proline Accumulation in Relation to Osmotic Adjustment in Rice (Oryza sativa L.) Callus Cultures. J. Plant Physiol. 1996, 149, 186–195. [Google Scholar] [CrossRef]
- Ashraf, M.A.; Ashraf, M.; Shahbaz, M. Growth Stage-Based Modulation in Antioxidant Defense System and Proline Accumulation in Two Hexaploid Wheat (Triticum aestivum L.) Cultivars Differing in Salinity Tolerance. Flora Morphol. Distrib. Funct. Ecol. Plants 2012, 207, 388–397. [Google Scholar] [CrossRef]
- Ali, S.G.; Rab, A.; Khan, N.U.; Nawab, K. Enhanced Proline Synthesis May Determine Resistance to Salt Stress in Tomato Cultivars. Pak. J. Bot. 2011, 43, 2707–2710. [Google Scholar]
- Abdelhamid, M.T.; Rady, M.M.; Osman, A.S.; Abdalla, M.A. Exogenous Application of Proline Alleviates Salt-Induced Oxidative Stress in Phaseolus vulgaris L. Plants. J. Hortic. Sci. Biotechnol. 2015, 88, 439–446. [Google Scholar] [CrossRef]
- Bhusan, D.; Das, D.K.; Hossain, M.; Murata, Y.; Hoque, M.A. Improvement of Salt Tolerance in Rice (“Oryza sativa” L.) by Increasing Antioxidant Defense Systems Using Exogenous Application of Proline. Aust. J. Crop Sci. 2016, 10, 50–56. [Google Scholar]
- de Freitas, P.A.F.; de Carvalho, H.H.; Costa, J.H.; Miranda, R.D.S.; Saraiva, K.D.D.C.; de Oliveira, F.D.B.; Coelho, D.G.; Prisco, J.T.; Gomes-Filho, E. Salt Acclimation in Sorghum Plants by Exogenous Proline: Physiological and Biochemical Changes and Regulation of Proline Metabolism. Plant Cell Rep. 2019, 38, 403–416. [Google Scholar] [CrossRef] [PubMed]
- de Freitas, P.A.F.; de Souza Miranda, R.; Marques, E.C.; Prisco, J.T.; Gomes-Filho, E. Salt Tolerance Induced by Exogenous Proline in Maize Is Related to Low Oxidative Damage and Favorable Ionic Homeostasis. J. Plant Growth Regul. 2018, 37, 911–924. [Google Scholar] [CrossRef]
- Heuer, B. Influence of Exogenous Application of Proline and Glycinebetaine on Growth of Salt-Stressed Tomato Plants. Plant Sci. 2003, 165, 693–699. [Google Scholar] [CrossRef]
- Kaya, C.; Tuna, A.L.; Ashraf, M.; Altunlu, H. Improved Salt Tolerance of Melon (Cucumis melo L.) by the Addition of Proline and Potassium Nitrate. Environ. Exp. Bot. 2007, 60, 397–403. [Google Scholar] [CrossRef]
- Mahboob, W.; Khan, M.; Bot, M.S.; Pak, J. Induction of salt tolerance in wheat (Triticum aestivum L.) seedlings through exogenous application of proline. Pak. J. Bot. 2016, 48, 861–867. [Google Scholar]
- Medeiros, M.J.L.; de Silva, M.M.A.; Granja, M.M.C.; de Souza, E.S.G.; Camara, T.; Willadino, L. Effect of Exogenous Proline in Two Sugarcane Genotypes Grown in Vitro under Salt Stress. Acta Biológica Colomb. 2015, 20, 57–63. [Google Scholar] [CrossRef]
- Wu, G.-Q.; Du, Y.-Y.; Wu, G.-Q.; Feng, R.-J.; Li, S.-J.; Du, Y.-Y. Exogenous Application of Proline Alleviates Salt-Induced Toxicity in Sainfoin Seedlings. J. Anim. Plant Sci. 2017, 27, 246–251. [Google Scholar]
- Messedi, D.; Farhani, F.; Hamed, K.; Trabelsi, N.; Bot, R.K.-P.J. Highlighting the Mechanisms by Which Proline Can Confer Tolerance to Salt Stress in Cakile maritima. Pak. J. Bot. 2016, 48, 417–427. [Google Scholar]
- Huang, Y.; Bie, Z.; Liu, Z.; Zhen, A.; Wang, W. Protective Role of Proline against Salt Stress Is Partially Related to the Improvement of Water Status and Peroxidase Enzyme Activity in Cucumber. Soil Sci. Plant Nutr. 2010, 55, 698–704. [Google Scholar] [CrossRef] [Green Version]
- ben Ahmed, C.; Magdich, S.; ben Rouina, B.; Sensoy, S.; Boukhris, M.; ben Abdullah, F. Exogenous Proline Effects on Water Relations and Ions Contents in Leaves and Roots of Young Olive. Amino Acids 2011, 40, 565–573. [Google Scholar] [CrossRef] [PubMed]
- Shahid, M.A.; Balal, R.M.; Pervez, M.A.; Abbas, T.; Aqeel, M.A.; Javaid, M.M.; Garcia-Sanchez, F. Exogenous Proline and Proline-Enriched Lolium perenne Leaf Extract Protects against Phytotoxic Effects of Nickel and Salinity in Pisum Sativum by Altering Polyamine Metabolism in Leaves. Turk. J. Bot. 2014, 38, 914–926. [Google Scholar] [CrossRef] [Green Version]
- Hasanuzzaman, M.; Alam, M.M.; Rahman, A.; Hasanuzzaman, M.; Nahar, K.; Fujita, M. Exogenous Proline and Glycine Betaine Mediated Upregulation of Antioxidant Defense and Glyoxalase Systems Provides Better Protection against Salt-Induced Oxidative Stress in Two Rice (Oryza sativa L.) Varieties. Biomed. Res. Int. 2014, 2014, 757219. [Google Scholar] [CrossRef] [Green Version]
- Wani, A.S.; Ahmad, A.; Hayat, S.; Tahir, I. Is Foliar Spray of Proline Sufficient for Mitigation of Salt Stress in Brassica juncea Cultivars? Environ. Sci. Pollut. Res. 2016, 23, 13413–13423. [Google Scholar] [CrossRef]
- Wani, A.S.; Ahmad, A.; Hayat, S.; Tahir, I. Epibrassinolide and Proline Alleviate the Photosynthetic and Yield Inhibition under Salt Stress by Acting on Antioxidant System in Mustard. Plant Physiol. Biochem. 2019, 135, 385–394. [Google Scholar] [CrossRef]
- Zheng, J.L.; Zhao, L.Y.; Wu, C.W.; Shen, B.; Zhu, A.Y. Exogenous Proline Reduces NaCl-Induced Damage by Mediating Ionic and Osmotic Adjustment and Enhancing Antioxidant Defense in Eurya emarginata. Acta Physiol. Plant 2015, 37, 181. [Google Scholar] [CrossRef]
- Reza, S.; Heidari, R.; Zare, S.; Norastehnia, A. Antioxidant Response of Two Salt-Stressed Barley Varieties in the Presence or Absence of Exogenous Proline. Gen. Appl. Plant Physiol. 2006, 32, 233–251. [Google Scholar]
- Hossain, M.A.; Fujita, M. Evidence for a Role of Exogenous Glycinebetaine and Proline in Antioxidant Defense and Methylglyoxal Detoxification Systems in Mung Bean Seedlings under Salt Stress. Physiol. Mol. Biol. Plants 2010, 16, 19–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agami, R.A. Applications of Ascorbic Acid or Proline Increase Resistance to Salt Stress in Barley Seedlings. Biol. Plant 2014, 58, 341–347. [Google Scholar] [CrossRef]
- ben Ahmed, C.; ben Rouina, B.; Sensoy, S.; Boukhriss, M.; ben Abdullah, F. Exogenous Proline Effects on Photosynthetic Performance and Antioxidant Defense System of Young Olive Tree. J. Agric. Food Chem. 2010, 58, 4216–4222. [Google Scholar] [CrossRef]
- Alam, R.; Das, D.K.; Islam, M.R.; Murata, Y.; Hoque, M.A. Exogenous Proline Enhances Nutrient Uptake and Confers Tolerance to Salt Stress in Maize (Zea mays L.). Progress. Agric. 2016, 27, 409–417. [Google Scholar] [CrossRef] [Green Version]
- el Sabagh, A.; Sorour, S.; Ragab, A.; Saneoka, H.; Islam, M.S.; Sabagh, A. The Effect of Exogenous Application of Proline and Glycine Betaine on the Nodule Activity of Soybean Under Saline Condition. J. Agric. Biotechnol. 2017, 2, 300–310. [Google Scholar] [CrossRef]
- Guerzoni, J.T.S.; Belintani, N.G.; Moreira, R.M.P.; Hoshino, A.A.; Domingues, D.S.; Filho, J.C.B.; Vieira, L.G.E. Stress-Induced Δ1-Pyrroline-5-Carboxylate Synthetase (P5CS) Gene Confers Tolerance to Salt Stress in Transgenic Sugarcane. Acta Physiol. Plant 2014, 36, 2309–2319. [Google Scholar] [CrossRef]
- Guan, C.; Cui, X.; Liu, H.Y.; Li, X.; Li, M.Q.; Zhang, Y.W. Proline Biosynthesis Enzyme Genes Confer Salt Tolerance to Switchgrass (Panicum virgatum L.) in Cooperation With Polyamines Metabolism. Front. Plant Sci. 2020, 11, 46. [Google Scholar] [CrossRef]
- Reddy, P.S.; Jogeswar, G.; Rasineni, G.K.; Maheswari, M.; Reddy, A.R.; Varshney, R.K.; Kavi Kishor, P.B. Proline Over-Accumulation Alleviates Salt Stress and Protects Photosynthetic and Antioxidant Enzyme Activities in Transgenic Sorghum [Sorghum bicolor (L.) Moench]. Plant Physiol. Biochem. 2015, 94, 104–113. [Google Scholar] [CrossRef] [Green Version]
- Soares, C.; Carvalho, M.E.A.; Azevedo, R.A.; Fidalgo, F. Plants Facing Oxidative Challenges—A Little Help from the Antioxidant Networks. Environ. Exp. Bot. 2019, 161, 4–25. [Google Scholar] [CrossRef]
- Zarattini, M.; Forlani, G. Toward Unveiling the Mechanisms for Transcriptional Regulation of Proline Biosynthesis in the Plant Cell Response to Biotic and Abiotic Stress Conditions. Front. Plant Sci. 2017, 8, 927. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, W.; Wang, N.; Yang, J.; Guo, H.; Liu, Z.; Zheng, X.; Li, S.; Xiang, F. The Salt-Induced Transcription Factor GmMYB84 Confers Salinity Tolerance in Soybean. Plant Sci. 2020, 291, 110326. [Google Scholar] [CrossRef] [PubMed]
- Zhu, D.; Hou, L.; Xiao, P.; Guo, Y.; Deyholos, M.K.; Liu, X. VvWRKY30, a Grape WRKY Transcription Factor, Plays a Positive Regulatory Role under Salinity Stress. Plant Sci. 2019, 280, 132–142. [Google Scholar] [CrossRef]
- Gao, Y.F.; Liu, J.K.; Yang, F.M.; Zhang, G.Y.; Wang, D.; Zhang, L.; Ou, Y.B.; Yao, Y.A. The WRKY Transcription Factor WRKY8 Promotes Resistance to Pathogen Infection and Mediates Drought and Salt Stress Tolerance in Solanum lycopersicum. Physiol. Plant 2020, 168, 98–117. [Google Scholar] [CrossRef]
- Liang, J.; Zheng, J.; Wu, Z.; Wang, H. Strawberry FaNAC2 Enhances Tolerance to Abiotic Stress by Regulating Proline Metabolism. Plants 2020, 9, 1417. [Google Scholar] [CrossRef]
- Alshareef, N.O.; Wang, J.Y.; Ali, S.; Al-Babili, S.; Tester, M.; Schmöckel, S.M. Overexpression of the NAC Transcription Factor JUNGBRUNNEN1 (JUB1) Increases Salinity Tolerance in Tomato. Plant Physiol. Biochem. 2019, 140, 113–121. [Google Scholar] [CrossRef] [PubMed]
- Verma, D.; Jalmi, S.K.; Bhagat, P.K.; Verma, N.; Sinha, A.K. A BHLH Transcription Factor, MYC2, Imparts Salt Intolerance by Regulating Proline Biosynthesis in Arabidopsis. FEBS J. 2020, 287, 2560–2576. [Google Scholar] [CrossRef]
- Zhang, W.; Yang, G.; Mu, D.; Li, H.; Zang, D.; Xu, H.; Zou, X.; Wang, Y. An Ethylene-Responsive Factor BpERF11 Negatively Modulates Salt and Osmotic Tolerance in Betula platyphylla. Sci. Rep. 2016, 6, 23085. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Hou, Y.; Li, M.; Zhang, F.; Yi, F.; Kang, J.; Yang, Q.; Long, R. Overexpression of an ABA-Inducible Homeodomain-Leucine Zipper I Gene MsHB7 Confers Salt Stress Sensitivity to Alfalfa. Ind. Crops Prod. 2022, 177, 114463. [Google Scholar] [CrossRef]
- Noor, I.; Sohail, H.; Sun, J.; Nawaz, M.A.; Li, G.; Hasanuzzaman, M.; Liu, J. Heavy Metal and Metalloid Toxicity in Horticultural Plants: Tolerance Mechanism and Remediation Strategies. Chemosphere 2022, 303, 135196. [Google Scholar] [CrossRef]
- Handique, G.K.; Handique, A.K. Proline Accumulation in Lemongrass (Cymbopogon flexuosus Stapf.) Due to Heavy Metal Stress. J. Environ. Biol. 2009, 30, 299–302. [Google Scholar]
- Zengin, F.K.; Munzuroglu, O. Effects of Some Heavy Metals on Content of Chlorophyll, Proline and Some Antioxidant Chemicals in Bean (Phaseolus vulgaris L.) Seedlings. Acta Biol. Crac. Ser. Bot. 2005, 47, 157–164. [Google Scholar]
- Bhagyawant, S.S.; Narvekar, D.T.; Gupta, N.; Bhadkaria, A.; Koul, K.K.; Srivastava, N. Variations in the Antioxidant and Free Radical Scavenging under Induced Heavy Metal Stress Expressed as Proline Content in Chickpea. Physiol. Mol. Biol. Plants 2019, 25, 683–696. [Google Scholar] [CrossRef] [PubMed]
- Ameen, N.; Amjad, M.; Murtaza, B.; Abbas, G.; Shahid, M.; Imran, M.; Naeem, M.A.; Niazi, N.K. Biogeochemical Behavior of Nickel under Different Abiotic Stresses: Toxicity and Detoxification Mechanisms in Plants. Environ. Sci. Pollut. Res. 2019, 26, 10496–10514. [Google Scholar] [CrossRef] [PubMed]
- Ghori, N.H.; Ghori, T.; Hayat, M.Q.; Imadi, S.R.; Gul, A.; Altay, V.; Ozturk, M. Heavy Metal Stress and Responses in Plants. Int. J. Environ. Sci. Technol. 2019, 16, 1807–1828. [Google Scholar] [CrossRef]
- Gajewska, E.; Skłodowska, M. Differential Biochemical Responses of Wheat Shoots and Roots to Nickel Stress: Antioxidative Reactions and Proline Accumulation. Plant Growth Regul. 2008, 54, 179–188. [Google Scholar] [CrossRef]
- Hadi, F.; Ali, N.; Fuller, M.P. Molybdenum (Mo) Increases Endogenous Phenolics, Proline and Photosynthetic Pigments and the Phytoremediation Potential of the Industrially Important Plant Ricinus communis L. for Removal of Cadmium from Contaminated Soil. Environ. Sci. Pollut. Res. 2016, 23, 20408–20430. [Google Scholar] [CrossRef] [PubMed]
- Chaturvedi, R.; Favas, P.; Pratas, J.; Varun, M.; Paul, M.S. EDTA-Assisted Metal Uptake in Raphanus sativus L. and Brassica oleracea L.: Assessment of Toxicity and Food Safety. Bull. Environ. Contam. Toxicol. 2019, 103, 490–495. [Google Scholar] [CrossRef] [PubMed]
- Soares, C.; de Sousa, A.; Pinto, A.; Azenha, M.; Teixeira, J.; Azevedo, R.A.; Fidalgo, F. Effect of 24-Epibrassinolide on ROS Content, Antioxidant System, Lipid Peroxidation and Ni Uptake in Solanum nigrum L. under Ni Stress. Environ. Exp. Bot. 2016, 122, 115–125. [Google Scholar] [CrossRef]
- Siddiqui, M.H.; Al-Whaibi, M.H.; Basalah, M.O. Interactive Effect of Calcium and Gibberellin on Nickel Tolerance in Relation to Antioxidant Systems in Triticum aestivum L. Protoplasma 2011, 248, 503–511. [Google Scholar] [CrossRef] [PubMed]
- Sirhindi, G.; Mir, M.A.; Abd-Allah, E.F.; Ahmad, P.; Gucel, S. Jasmonic Acid Modulates the Physio-Biochemical Attributes, Antioxidant Enzyme Activity, and Gene Expression in Glycine max under Nickel Toxicity. Front. Plant Sci. 2016, 7, 591. [Google Scholar] [CrossRef] [Green Version]
- Hayat, S.; Hayat, Q.; Alyemeni, M.N.; Ahmad, A. Proline Enhances Antioxidative Enzyme Activity, Photosynthesis and Yield of Cicer arietinum L. Exposed to Cadmium Stress. Acta Bot. Croat. 2013, 72, 323–335. [Google Scholar] [CrossRef] [Green Version]
- Zouari, M.; ben Hassena, A.; Trabelsi, L.; ben Rouina, B.; Decou, R.; Labrousse, P. Exogenous Proline-Mediated Abiotic Stress Tolerance in Plants: Possible Mechanisms. Osmoprotectant Mediat. Abiotic Stress Toler. Plants 2019, 2019, 99–121. [Google Scholar] [CrossRef]
- Rizwan, M.; Imtiaz, M.; Dai, Z.; Mehmood, S.; Adeel, M.; Liu, J.; Tu, S. Nickel Stressed Responses of Rice in Ni Subcellular Distribution, Antioxidant Production, and Osmolyte Accumulation. Environ. Sci. Pollut. Res. 2017, 24, 20587–20598. [Google Scholar] [CrossRef] [PubMed]
- Spormann, S.; Soares, C.; Martins, V.; Azenha, M.; Gerós, H.; Fidalgo, F. Early Activation of Antioxidant Responses in Ni-Stressed Tomato Cultivars Determines Their Resilience Under Co-Exposure to Drought. J. Plant Growth Regul. 2022, 42, 877–891. [Google Scholar] [CrossRef]
- Tiwari, K.K.; Singh, N.K.; Rai, U.N. Chromium Phytotoxicity in Radish (Raphanus sativus): Effects on Metabolism and Nutrient Uptake. Bull. Environ. Contam. Toxicol. 2013, 91, 339–344. [Google Scholar] [CrossRef]
- Cia, M.C.; Guimarães, A.C.R.; Medici, L.O.; Chabregas, S.M.; Azevedo, R.A. Antioxidant Responses to Water Deficit by Drought-Tolerant and -Sensitive Sugarcane Varieties. Ann. Appl. Biol. 2012, 161, 313–324. [Google Scholar] [CrossRef]
- ur Rehman, M.Z.; Rizwan, M.; Ali, S.; Ok, Y.S.; Ishaque, W.; Saifullah; Nawaz, M.F.; Akmal, F.; Waqar, M. Remediation of Heavy Metal Contaminated Soils by Using Solanum nigrum: A Review. Ecotoxicol. Environ. Saf. 2017, 143, 236–248. [Google Scholar] [CrossRef]
- Nedjimi, B.; Daoud, Y. Cadmium Accumulation in Atriplex halimus Subsp. Schweinfurthii and Its Influence on Growth, Proline, Root Hydraulic Conductivity and Nutrient Uptake. Flora Morphol. Distrib. Funct. Ecol. Plants 2009, 204, 316–324. [Google Scholar] [CrossRef]
- Sun, R.L.; Zhou, Q.X.; Sun, F.H.; Jin, C.X. Antioxidative Defense and Proline/Phytochelatin Accumulation in a Newly Discovered Cd-Hyperaccumulator, Solanum nigrum L. Environ. Exp. Bot. 2007, 60, 468–476. [Google Scholar] [CrossRef]
- Matsunami, M.; Toyofuku, K.; Kimura, N.; Plants, A.O. Osmotic Stress Leads to Significant Changes in Rice Root Metabolic Profiles between Tolerant and Sensitive Genotypes. Plants 2020, 9, 1503. [Google Scholar] [CrossRef]
- Schat, H.; Sharma, S.S.; Schat, R.V.; Sharma, H.; Vooijs, S.S. Heavy Metal-Induced Accumulation of Free Proline in a Metal-Tolerant and a Nontolerant Ecotype of Silene vuigaris. Physiol. Plant. 1997, 101, 477–482. [Google Scholar] [CrossRef]
- Sofy, M.R.; Seleiman, M.F.; Alhammad, B.A.; Alharbi, B.M.; Mohamed, H.I. Minimizing Adverse Effects of Pb on Maize Plants by Combined Treatment with Jasmonic, Salicylic Acids and Proline. Agronomy 2020, 10, 699. [Google Scholar] [CrossRef]
- Noreen, S.; Akhter, M.S.; Yaamin, T.; Arfan, M. The Ameliorative Effects of Exogenously Applied Proline on Physiological and Biochemical Parameters of Wheat (Triticum aestivum L.) Crop under Copper Stress Condition. J. Plant Interact. 2018, 13, 221–230. [Google Scholar] [CrossRef] [Green Version]
- Zouari, M.; Elloumi, N.; ben Ahmed, C.; Delmail, D.; ben Rouina, B.; ben Abdallah, F.; Labrousse, P. Exogenous Proline Enhances Growth, Mineral Uptake, Antioxidant Defense, and Reduces Cadmium-Induced Oxidative Damage in Young Date Palm (Phoenix dactylifera L.). Ecol. Eng. 2016, 86, 202–209. [Google Scholar] [CrossRef]
- Wang, F.; Zeng, B.; Sun, Z.; Zhu, C. Relationship between Proline and Hg2+-Induced Oxidative Stress in a Tolerant Rice Mutant. Arch. Environ. Contam. Toxicol. 2009, 56, 723–731. [Google Scholar] [CrossRef]
- Chen, C.T.; Chen, T.H.; Lo, K.F.; Chiu, C.Y. Effects of Proline on Copper Transport in Rice Seedlings under Excess Copper Stress. Plant Sci. 2004, 166, 103–111. [Google Scholar] [CrossRef]
- Chandrakar, V.; Parkhey, S.; Dubey, A.; Keshavkant, S. Modulation in Arsenic-Induced Lipid Catabolism in Glycine max Using Proline, 24-Epibrassinolide and Diphenylene Iodonium. Biologia 2017, 72, 292–299. [Google Scholar] [CrossRef]
- Khatun, M.; Matsushima, D.; Rhaman, M.S.; Okuma, E.; Nakamura, T.; Nakamura, Y.; Munemasa, S.; Murata, Y. Exogenous Proline Enhances Antioxidant Enzyme Activities but Does Not Mitigate Growth Inhibition by Selenate Stress in Tobacco BY-2 Cells. Biosci. Biotechnol. Biochem. 2020, 84, 2281–2292. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Yin, H.; Li, X. Protective Effects of Proline against Cadmium Toxicity in Micropropagated Hyperaccumulator, Solanum nigrum L. Plant Cell Rep. 2009, 28, 325–333. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.Z.; Lin, Y.J.; Fan, W.J.; Lu, M.R. The Role of Exogenous Proline in Amelioration of Lipid Peroxidation in Rice Seedlings Exposed to Cr(VI). Int. Biodeterior. Biodegrad. 2017, 123, 106–112. [Google Scholar] [CrossRef]
- Wang, Y.; Tan, P.; Chang, L.; Yue, Z.; Zeng, C.; Li, M.; Liu, Z.; Dong, X.; Yan, M. Exogenous Proline Mitigates Toxic Effects of Cadmium via the Decrease of Cadmium Accumulation and Reestablishment of Redox Homeostasis in Brassica juncea. BMC Plant Biol. 2022, 22, 182. [Google Scholar] [CrossRef] [PubMed]
- Nedjimi, B. Phytoremediation: A Sustainable Environmental Technology for Heavy Metals Decontamination. SN Appl. Sci. 2021, 3, 286. [Google Scholar] [CrossRef]
- Ullah, R.; Hadi, F.; Ahmad, S.; Jan, A.U.; Rongliang, Q. Phytoremediation of Lead and Chromium Contaminated Soil Improves with the Endogenous Phenolics and Proline Production in Parthenium, Cannabis, Euphorbia, and Rumex Species. Water Air Soil Pollut. 2019, 230, 40. [Google Scholar] [CrossRef]
- Balal, R.M.; Shahid, M.A.; Javaid, M.M.; Anjum, M.A.; Ali, H.H.; Mattson, N.S.; Garcia-Sanchez, F. Foliar Treatment with Lolium perenne (Poaceae) Leaf Extract Alleviates Salinity and Nickel-Induced Growth Inhibition in Pea. Rev. Bras. De Bot. 2016, 39, 453–463. [Google Scholar] [CrossRef]
- Singh, M.; Pratap Singh, V.; Dubey, G.; Mohan Prasad, S. Exogenous Proline Application Ameliorates Toxic Effects of Arsenate in Solanum melongena L. Seedlings. Ecotoxicol. Environ. Saf. 2015, 117, 164–173. [Google Scholar] [CrossRef]
- Aggarwal, M.; Sharma, S.; Kaur, N.; Pathania, D.; Bhandhari, K.; Kaushal, N.; Kaur, R.; Singh, K.; Srivastava, A.; Nayyar, H. Exogenous Proline Application Reduces Phytotoxic Effects of Selenium by Minimising Oxidative Stress and Improves Growth in Bean (Phaseolus vulgaris L.) Seedlings. Biol. Trace Elem. Res. 2011, 140, 354–367. [Google Scholar] [CrossRef]
- Hayat, K.; Khan, J.; Khan, A.; Ullah, S.; Ali, S.; Salahudin; Fu, Y. Ameliorative Effects of Exogenous Proline on Photosynthetic Attributes, Nutrients Uptake, and Oxidative Stresses under Cadmium in Pigeon Pea (Cajanus cajan L.). Plants 2021, 10, 796. [Google Scholar] [CrossRef]
- Zouari, M.; Elloumi, N.; Labrousse, P.; ben Rouina, B.; ben Abdallah, F.; ben Ahmed, C. Olive Trees Response to Lead Stress: Exogenous Proline Provided Better Tolerance than Glycine Betaine. S. Afr. J. Bot. 2018, 118, 158–165. [Google Scholar] [CrossRef]
- Adejumo, S.A.; Awoyemi, V.; Togun, A.O. Exogenous Proline and Hormone in Combination with Compost Improves Growth and Tolerance of Maize under Heavy Metal Stress. Plants Environ. 2020, 2, 40–53. [Google Scholar] [CrossRef]
- Kapoor, D.; Kavani, K.; Rattan, A.; Landi, M.; Sharma, A. Ameliorative Role of Pre-Sowing Proline Treatment in Coriandrum sativum L. Seedlings under Mercury Toxicity. Phyton Int. J. Exp. Bot. 2021, 90, 489–501. [Google Scholar] [CrossRef]
- Shahbaz, A.K.; Adnan Ramzani, P.M.; Saeed, R.; Turan, V.; Iqbal, M.; Lewińska, K.; Abbas, F.; Saqib, M.; Tauqeer, H.M.; Iqbal, M.; et al. Effects of Biochar and Zeolite Soil Amendments with Foliar Proline Spray on Nickel Immobilization, Nutritional Quality and Nickel Concentrations in Wheat. Ecotoxicol. Environ. Saf. 2019, 173, 182–191. [Google Scholar] [CrossRef] [PubMed]
- Rady, M.M.; Elrys, A.S.; Abo El-Maati, M.F.; Desoky, E.S.M. Interplaying Roles of Silicon and Proline Effectively Improve Salt and Cadmium Stress Tolerance in Phaseolus vulgaris Plant. Plant Physiol. Biochem. 2019, 139, 558–568. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.; Gupta, V.K.; Kumar, A.; Sharma, B. Synergistic Effects of Heavy Metals and Pesticides in Living Systems. Front. Chem. 2017, 5, 70. [Google Scholar] [CrossRef]
- Godheja, J.; Shekhar, S.K.; Siddiqui, S.A. Xenobiotic Compounds Present in Soil and Water: A Review on Remediation Strategies. J. Environ. Anal. Toxicol. 2016, 6, 1000392. [Google Scholar] [CrossRef]
- Duarte, R.M.B.O.; Matos, J.T.V.; Senesi, N. Organic Pollutants in Soils. Soil Pollut. Monit. Remediat. 2018, 103–126. [Google Scholar] [CrossRef]
- Soares, C.; Branco-Neves, S.; de Sousa, A.; Pereira, R.; Fidalgo, F. Ecotoxicological Relevance of Nano-NiO and Acetaminophen to Hordeum vulgare L.: Combining Standardized Procedures and Physiological Endpoints. Chemosphere 2016, 165, 442–452. [Google Scholar] [CrossRef]
- Soares, C.; Pereira, R.; Spormann, S.; Fidalgo, F. Is Soil Contamination by a Glyphosate Commercial Formulation Truly Harmless to Non-Target Plants?—Evaluation of Oxidative Damage and Antioxidant Responses in Tomato. Environ. Pollut. 2019, 247, 256–265. [Google Scholar] [CrossRef]
- Soares, C.; Pereira, R.; Martins, M.; Tamagnini, P.; Serôdio, J.; Moutinho-Pereira, J.; Cunha, A.; Fidalgo, F. Glyphosate-Dependent Effects on Photosynthesis of Solanum lycopersicum L.—An Ecophysiological, Ultrastructural and Molecular Approach. J. Hazard. Mater. 2020, 398, 122871. [Google Scholar] [CrossRef]
- Martins, M.; Sousa, B.; Lopes, J.; Soares, C.; Machado, J.; Carvalho, S.; Fidalgo, F.; Teixeira, J. Diclofenac Shifts the Role of Root Glutamine Synthetase and Glutamate Dehydrogenase for Maintaining Nitrogen Assimilation and Proline Production at the Expense of Shoot Carbon Reserves in Solanum lycopersicum L. Environ. Sci. Pollut. Res. 2020, 27, 29130–29142. [Google Scholar] [CrossRef] [PubMed]
- Sousa, B.; Lopes, J.; Leal, A.; Martins, M.; Soares, C.; Valente, I.M.; Rodrigues, J.A.; Fidalgo, F.; Teixeira, J. Response of Solanum lycopersicum L. to Diclofenac—Impacts on the Plant’s Antioxidant Mechanisms. Environ. Pollut. 2020, 258, 113762. [Google Scholar] [CrossRef]
- Branco-Neves, S.; Soares, C.; de Sousa, A.; Martins, V.; Azenha, M.; Gerós, H.; Fidalgo, F. An Efficient Antioxidant System and Heavy Metal Exclusion from Leaves Make Solanum cheesmaniae More Tolerant to Cu than Its Cultivated Counterpart. Food Energy Secur. 2017, 6, 123–133. [Google Scholar] [CrossRef]
- Gomes, M.P.; le Manac’h, S.G.; Moingt, M.; Smedbol, E.; Paquet, S.; Labrecque, M.; Lucotte, M.; Juneau, P. Impact of Phosphate on Glyphosate Uptake and Toxicity in Willow. J. Hazard. Mater. 2016, 304, 269–279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartha, B.; Huber, C.; Harpaintner, R.; Schröder, P. Effects of Acetaminophen in Brassica juncea L. Czern.: Investigation of Uptake, Translocation, Detoxification, and the Induced Defense Pathways. Environ. Sci. Pollut. Res. 2010, 17, 1553–1562. [Google Scholar] [CrossRef]
- Soares, C.; Fernandes, B.; Paiva, C.; Nogueira, V.; Cachada, A.; Fidalgo, F.; Pereira, R. Ecotoxicological Relevance of Glyphosate and Flazasulfuron to Soil Habitat and Retention Functions—Single vs. Combined Exposures. J. Hazard. Mater. 2023, 442, 130128. [Google Scholar] [CrossRef]
- Singh, S.; Kumar, V.; Datta, S.; Wani, A.B.; Dhanjal, D.S.; Romero, R.; Singh, J. Glyphosate Uptake, Translocation, Resistance Emergence in Crops, Analytical Monitoring, Toxicity and Degradation: A Review. Environ. Chem. Lett. 2020, 18, 663–702. [Google Scholar] [CrossRef]
- Sun, C.; Dudley, S.; Trumble, J.; Gan, J. Pharmaceutical and Personal Care Products-Induced Stress Symptoms and Detoxification Mechanisms in Cucumber Plants. Environ. Pollut. 2018, 234, 39–47. [Google Scholar] [CrossRef] [Green Version]
- Gomes, M.P.; Smedbol, E.; Chalifour, A.; Hénault-Ethier, L.; Labrecque, M.; Lepage, L.; Lucotte, M.; Juneau, P. Alteration of Plant Physiology by Glyphosate and Its By-Product Aminomethylphosphonic Acid: An Overview. J. Exp. Bot. 2014, 65, 4691–4703. [Google Scholar] [CrossRef] [Green Version]
- Gomes, M.P.; le Manac’h, S.G.; Hénault-Ethier, L.; Labrecque, M.; Lucotte, M.; Juneau, P. Glyphosate-Dependent Inhibition of Photosynthesis in Willow. Front. Plant Sci. 2017, 8, 207. [Google Scholar] [CrossRef] [Green Version]
- Soares, C.; Nadais, P.; Sousa, B.; Pinto, E.; Ferreira, I.M.P.L.V.O.; Pereira, R.; Fidalgo, F. Silicon Improves the Redox Homeostasis to Alleviate Glyphosate Toxicity in Tomato Plants—Are Nanomaterials Relevant? Antioxidants 2021, 10, 1320. [Google Scholar] [CrossRef] [PubMed]
- Soares, C.; Rodrigues, F.; Sousa, B.; Pinto, E.; Ferreira, I.M.P.L.V.O.; Pereira, R.; Fidalgo, F. Foliar Application of Sodium Nitroprusside Boosts Solanum lycopersicum L. Tolerance to Glyphosate by Preventing Redox Disorders and Stimulating Herbicide Detoxification Pathways. Plants 2021, 10, 1862. [Google Scholar] [CrossRef]
- Stuchlíková, L.R.; Skálová, L.; Szotáková, B.; Syslová, E.; Vokřál, I.; Vaněk, T.; Podlipná, R. Biotransformation of Flubendazole and Fenbendazole and Their Effects in the Ribwort Plantain (Plantago lanceolata). Ecotoxicol. Environ. Saf. 2018, 147, 681–687. [Google Scholar] [CrossRef] [PubMed]
- Navrátilová, M.; Raisová Stuchlíková, L.; Skálová, L.; Szotáková, B.; Langhansová, L.; Podlipná, R. Pharmaceuticals in Environment: The Effect of Ivermectin on Ribwort Plantain (Plantago lanceolata L.). Environ. Sci. Pollut. Res. 2020, 27, 31202–31210. [Google Scholar] [CrossRef]
- Christou, A.; Antoniou, C.; Christodoulou, C.; Hapeshi, E.; Stavrou, I.; Michael, C.; Fatta-Kassinos, D.; Fotopoulos, V. Stress-Related Phenomena and Detoxification Mechanisms Induced by Common Pharmaceuticals in Alfalfa (Medicago sativa L.) Plants. Sci. Total Environ. 2016, 557–558, 652–664. [Google Scholar] [CrossRef]
- Pawłowska, B.; Telesiński, A.; Biczak, R. Effect of Diclofenac and Naproxen and Their Mixture on Spring Barley Seedlings and Heterocypris incongruens. Environ. Toxicol. Pharmacol. 2021, 88, 103746. [Google Scholar] [CrossRef]
- Soares, C.; Branco-Neves, S.; de Sousa, A.; Teixeira, J.; Pereira, R.; Fidalgo, F. Can Nano-SiO2 Reduce the Phytotoxicity of Acetaminophen?—A Physiological, Biochemical and Molecular Approach. Environ. Pollut. 2018, 241, 900–911. [Google Scholar] [CrossRef] [PubMed]
- Nunes, B.; Pinto, G.; Martins, L.; Gonçalves, F.; Antunes, S.C. Biochemical and Standard Toxic Effects of Acetaminophen on the Macrophyte Species Lemna minor and Lemna gibba. Environ. Sci. Pollut. Res. 2014, 21, 10815–10822. [Google Scholar] [CrossRef]
- Liu, Y.; Pang, Y.; Yang, L.; Ning, S.; Wang, D.; Wu, Z. Responses of Hydrocharis dubia (Bl.) Backer and Trapa Bispinosa Roxb. to Tetracycline Exposure. Ecotoxicol. Environ. Saf. 2020, 202, 110890. [Google Scholar] [CrossRef]
- Wu, G.L.; Cui, J.; Tao, L.; Yang, H. Fluroxypyr Triggers Oxidative Damage by Producing Superoxide and Hydrogen Peroxide in Rice (Oryza sativa). Ecotoxicology 2010, 19, 124–132. [Google Scholar] [CrossRef]
- Moreira, J.T.; Moreira, T.M.; Cunha, J.B.; Azenha, M.; Fidalgo, F.; Teixeira, J. Differential Effects of Acetophenone on Shoots’ and Roots’ Metabolism of Solanum nigrum L. Plants and Implications in Its Phytoremediation. Plant Physiol. Biochem. 2018, 130, 391–398. [Google Scholar] [CrossRef]
- Shakir, S.K.; Irfan, S.; Akhtar, B.; ur Rehman, S.; Daud, M.K.; Taimur, N.; Azizullah, A. Pesticide-Induced Oxidative Stress and Antioxidant Responses in Tomato (Solanum lycopersicum) Seedlings. Ecotoxicology 2018, 27, 919–935. [Google Scholar] [CrossRef] [PubMed]
- Ould Said, C.; Boulahia, K.; Eid, M.A.M.; Rady, M.M.; Djebbar, R.; Abrous-Belbachir, O. Exogenously Used Proline Offers Potent Antioxidative and Osmoprotective Strategies to Re-Balance Growth and Physio-Biochemical Attributes in Herbicide-Stressed Trigonella foenum-graecum. J. Soil Sci. Plant Nutr. 2021, 21, 3254–3268. [Google Scholar] [CrossRef]
- Shopova, E.; Katerova, Z.; Brankova, L.; Dimitrova, L.; Sergiev, I.; Todorova, D.; Talaat, N.B. Modulation of Physiological Stress Response of Triticum aestivum L. to Glyphosate by Brassinosteroid Application. Life 2021, 11, 1156. [Google Scholar] [CrossRef] [PubMed]
- Han, H.; Zhu, B.; Fu, X.; You, S.; Wang, B.; Li, Z.; Zhao, W.; Peng, R.; Yao, Q. Overexpression of D-Amino Acid Oxidase from Bradyrhizobium japonicum, Enhances Resistance to Glyphosate in Arabidopsis thaliana. Plant Cell. Rep. 2015, 34, 2043–2051. [Google Scholar] [CrossRef] [PubMed]
- Spormann, S.; Soares, C.; Fidalgo, F. Salicylic Acid Alleviates Glyphosate-Induced Oxidative Stress in Hordeum vulgare L. J. Environ. Manag. 2019, 241, 226–234. [Google Scholar] [CrossRef]
- Wang, Z.; Jiang, Y.; Peng, X.; Xu, S.; Zhang, H.; Gao, J.; Xi, Z. Exogenous 24-Epibrassinolide Regulates Antioxidant and Pesticide Detoxification Systems in Grapevine after Chlorothalonil Treatment. Plant Growth Regul. 2017, 81, 455–466. [Google Scholar] [CrossRef]
- Hellmann, H.; Funck, D.; Rentsch, D.; Frommer, W.B. Hypersensitivity of an Arabidopsis Sugar Signaling Mutant toward Exogenous Proline Application. Plant Physiol. 2000, 122, 357–368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hashem, A.; Alqarawi, A.A.; Radhakrishnan, R.; Al-Arjani, A.-B.F.; Aldehaish, H.A.; Egamberdieva, D.; Abd_Allah, E.F. Arbuscular Mycorrhizal Fungi Regulate the Oxidative System, Hormones and Ionic Equilibrium to Trigger Salt Stress Tolerance in Cucumis sativus L. Saudi J. Biol. Sci. 2018, 25, 1102–1114. [Google Scholar] [CrossRef]
- Latef, A.A.A.; Akter, A.; Tahjib-Ul-Arif, M. Foliar Application of Auxin or Cytokinin Can Confer Salinity Stress Tolerance in Vicia faba L. Agronomy 2021, 11, 228–233. [Google Scholar] [CrossRef]
- Siddiqui, H.; Yusuf, M.; Faraz, A.; Faizan, M.; Sami, F.; Hayat, S. 24-Epibrassinolide Supplemented with Silicon Enhances the Photosynthetic Efficiency of Brassica juncea under Salt Stress. S. Afr. J. Bot. 2018, 118, 120–128. [Google Scholar] [CrossRef]
- Kaur, H.; Hussain, S.J.; Kaur, G.; Poor, P.; Alamri, S.; Siddiqui, M.H.; Khan, M.I.R. Salicylic Acid Improves Nitrogen Fixation, Growth, Yield and Antioxidant Defence Mechanisms in Chickpea Genotypes Under Salt Stress. J. Plant Growth Regul. 2022, 41, 2034–2047. [Google Scholar] [CrossRef]
- Ahmad, S.; Cui, W.; Kamran, M.; Ahmad, I.; Meng, X.; Wu, X.; Su, W.; Javed, T.; El-Serehy, H.A.; Jia, Z.; et al. Exogenous Application of Melatonin Induces Tolerance to Salt Stress by Improving the Photosynthetic Efficiency and Antioxidant Defense System of Maize Seedling. J. Plant Growth Regul. 2021, 40, 1270–1283. [Google Scholar] [CrossRef]
- Shah, W.H.; Rasool, A.; Tahir, I.; Rehman, R.U. Exogenously Applied Selenium (Se) Mitigates the Impact of Salt Stress in Setaria italica L. and Panicum miliaceum L. Nucleus 2020, 63, 327–339. [Google Scholar] [CrossRef]
- Hurtado, A.C.; Chiconato, D.A.; Prado, R.d.M.; Sousa Junior, G.d.S.; Gratão, P.L.; Felisberto, G.; Olivera Viciedo, D.; Mathias dos Santos, D.M. Different Methods of Silicon Application Attenuate Salt Stress in Sorghum and Sunflower by Modifying the Antioxidative Defense Mechanism. Ecotoxicol. Environ. Saf. 2020, 203, 110964. [Google Scholar] [CrossRef] [PubMed]
- Sobahan, M.A.; Arias, C.R.; Okuma, E.; Shimoishi, Y.; Nakamura, Y.; Hirai, Y.; Mori, I.C.; Murata, Y. Exogenous Proline and Glycinebetaine Suppress Apoplastic Flow to Reduce Na+ Uptake in Rice Seedlings. Biosci. Biotechnol. Biochem. 2009, 73, 2037–2042. [Google Scholar] [CrossRef] [Green Version]
- Nawaz, K.; Talat, A.; Hussain, K.; Majeed, A. Induction of Salt Tolerance in Two Cultivars of Sorghum (Sorghum bicolor L.) by Exogenous Application of Proline at Seedling Stage. World Appl. Sci. J. 2010, 10, 93–99. [Google Scholar]
- Deivanai, S.; Xavier, R.; Vinod, V.; Timalata, K.; Lim, O.F. Role of Exogenous Proline in Ameliorating Salt Stress at Early Stage in Two Rice Cultivars. J. Stress Physiol. Biochem. 2011, 7, 157–174. [Google Scholar]
- Sakr, M.T.; El-Sarkassy, N.M.; Fuller, M.P. Osmoregulators Proline and Glycine Betaine Counteract Salinity Stress in Canola. Agron. Sustain. Dev. 2012, 32, 747–754. [Google Scholar] [CrossRef] [Green Version]
- Sobahan, M.A.; Akter, N.; Ohno, M.; Okuma, E.; Hirai, Y.; Mori, I.C.; Nakamura, Y.; Murata, Y. Effects of Exogenous Proline and Glycinebetaine on the Salt Tolerance of Rice Cultivars. Biosci. Biotechnol. Biochem 2012, 76, 1568–1570. [Google Scholar] [CrossRef]
- Nounjan, N.; Nghia, P.T.; Theerakulpisut, P. Exogenous Proline and Trehalose Promote Recovery of Rice Seedlings from Salt-Stress and Differentially Modulate Antioxidant Enzymes and Expression of Related Genes. J. Plant Physiol. 2012, 169, 596–604. [Google Scholar] [CrossRef]
- Nounjan, N.; Theerakulpisut, P. Effects of Exogenous Proline and Trehalose on Physiological Responses in Rice Seedlings during Salt-Stress and after Recovery. Plant Soil Environ. 2012, 58, 309–315. [Google Scholar] [CrossRef] [Green Version]
- Hua-long, L.; Han-jing, S.; Jing-guo, W.; Yang, L.; De-tang, Z.; Hong-wei, Z. Effect of Seed Soaking with Exogenous Proline on Seed Germination of Rice Under Salt Stress. J. Northeast. Agric. Univ. (Engl. Ed.) 2014, 21, 1–6. [Google Scholar] [CrossRef]
- El Sabagh, A.; Sorour, S.; Omar, A.E.; Ragab, A.; Islam, M.S.; Barutçular, C.; Ueda, A.; Saneoka, H. Alleviation of Adverse Effects of Salt Stress on Soybean (Glycine max L.) by Using Osmoprotectants and Organic Nutrients. Int. J. Agric. Biosyst. Eng. 2015, 9, 1014–1018. [Google Scholar]
- Teh, C.Y.; Mahmood, M.; Shaharuddin, N.A.; Ho, C.L. In Vitro Rice Shoot Apices as Simple Model to Study the Effect of NaCl and the Potential of Exogenous Proline and Glutathione in Mitigating Salinity Stress. Plant Growth Regul. 2015, 75, 771–781. [Google Scholar] [CrossRef]
- Rady, M.M.; Hemida, K.A. Sequenced Application of Ascorbate-Proline-Glutathione Improves Salt Tolerance in Maize Seedlings. Ecotoxicol. Environ. Saf. 2016, 133, 252–259. [Google Scholar] [CrossRef]
- Sobahan, M.A.; Akter, N.; Murata, Y.; Munemasa, S. Exogenous Proline and Glycinebetaine Mitigate the Detrimental Effect of Salt Stress on Rice Plants. Sci. Eng. Health Stud. 2016, 10, 38–43. [Google Scholar] [CrossRef]
- Singh, M.; Nehal, N.; Sharma, N.; Monika Singh, C.; Singh, A. Effect of Proline on Germination and Seedling Growth of Rice (Oryza sativa L.) under Salt Stress. J. Pharmacogn. Phytochem. 2018, 7, 2449–2452. [Google Scholar]
- Shafiq, F.; Raza, S.H.; Bibi, A.; Khan, I.; Iqbal, M. Influence of Proline Priming on Antioxidative Potential and Ionic Distribution and Its Relationship with Salt Tolerance of Wheat. Cereal Res. Commun. 2018, 46, 287–300. [Google Scholar] [CrossRef]
- Islam, M.M.; Hoque, M.A.; Okuma, E.; Banu, M.N.A.; Shimoishi, Y.; Nakamura, Y.; Murata, Y. Exogenous Proline and Glycinebetaine Increase Antioxidant Enzyme Activities and Confer Tolerance to Cadmium Stress in Cultured Tobacco Cells. J. Plant Physiol. 2009, 166, 1587–1597. [Google Scholar] [CrossRef]
- Min, S.O.N.G.; Wen-jing, X.U.; Xiang-yong, P.E.N.G.; Fan-hua, K.O.N.G. Effects of Exogenous Proline on the Growth of Wheat Seedlings under Cadmium Stress. Yingyong Shengtai Xuebao 2013, 24, 129–134. [Google Scholar]
- Rasheed, R.; Ashraf, M.A.; Hussain, I.; Haider, M.Z.; Kanwal, U.; Iqbal, M. Exogenous Proline and Glycinebetaine Mitigate Cadmium Stress in Two Genetically Different Spring Wheat (Triticum aestivum L.) Cultivars. Rev. Bras. Bot. 2014, 37, 399–406. [Google Scholar] [CrossRef]
- Zouari, M.; ben Ahmed, C.; Elloumi, N.; Bellassoued, K.; Delmail, D.; Labrousse, P.; ben Abdallah, F.; ben Rouina, B. Impact of Proline Application on Cadmium Accumulation, Mineral Nutrition and Enzymatic Antioxidant Defense System of Olea europaea L. Cv Chemlali Exposed to Cadmium Stress. Ecotoxicol. Environ. Saf. 2016, 128, 195–205. [Google Scholar] [CrossRef]
- Alyemeni, M.N.; Hayat, Q.; Hayat, S.; Faizan, M.; Faraz, A. Exogenous Proline Application Enhances the Efficiency of Nitrogen Fixation and Assimilation in Chickpea Plants Exposed to Cadmium. Legume Res. 2016, 39, 221–227. [Google Scholar] [CrossRef] [Green Version]
- Konotop, Y.; Kovalenko, M.; Matušíková, I.; Batsmanova, L.; Taran, N. Proline Application Triggers Temporal Redox Imbalance, but Alleviates Cadmium Stress in Wheat Seedlings. Pak. J. Bot. 2017, 49, 2145–2151. [Google Scholar]
- Singh, V.; Bhatt, I.; Aggarwal, A.; Tripathi, B.N.; Munjal, A.K.; Sharma, V. Proline Improves Copper Tolerance in Chickpea (Cicer arietinum). Protoplasma 2010, 245, 173–181. [Google Scholar] [CrossRef]
- Stetsenko, L.A.; Shevyakova, N.I.; Rakitin, V.Y.; Kuznetsov, V.v. Proline Protects Atropa belladonna Plants against Nickel Salt Toxicity. Russ. J. Plant Physiol. 2011, 58, 275–282. [Google Scholar] [CrossRef]
- Sadeghipour, O. The Effect of Common Bean (Phaseolus vulgaris L.) Seed Pretreatment with Proline and Glycine Betaine on Tolerance to Lead Stress. Environ. Stress. Crop Sci. 2019, 12, 319–329. [Google Scholar] [CrossRef]
- Mishra, S.; Dubey, R.S. Inhibition of Ribonuclease and Protease Activities in Arsenic Exposed Rice Seedlings: Role of Proline as Enzyme Protectant. J. Plant Physiol. 2006, 163, 927–936. [Google Scholar] [CrossRef] [PubMed]
- Ghahremani, B.; Hassannejad, S.; Alizadeh, K.; Eslam, B.P. Salicylic Acid Alleviates Oxidative Stress and Lipid Peroxidation Caused by Clopyralid Herbicide in Indian Mustard Plants. Acta Physiol. Plant 2022, 44, 1–10. [Google Scholar] [CrossRef]
- Soares, C.; Spormann, S.; Fidalgo, F. Salicylic Acid Improves the Performance of the Enzymatic Antioxidant System of Barley Exposed to Glyphosate. Free Radic. Biol. Med. 2018, 120, S157. [Google Scholar] [CrossRef]
- Tripthi, D.K.; Varma, R.K.; Singh, S.; Sachan, M.; Guerriero, G.; Kushwaha, B.K.; Bhardwaj, S.; Ramawat, N.; Sharma, S.; Singh, V.P.; et al. Silicon Tackles Butachlor Toxicity in Rice Seedlings by Regulating Anatomical Characteristics, Ascorbate-Glutathione Cycle, Proline Metabolism and Levels of Nutrients. Sci. Rep. 2020, 10, 1–15. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Spormann, S.; Nadais, P.; Sousa, F.; Pinto, M.; Martins, M.; Sousa, B.; Fidalgo, F.; Soares, C. Accumulation of Proline in Plants under Contaminated Soils—Are We on the Same Page? Antioxidants 2023, 12, 666. https://doi.org/10.3390/antiox12030666
Spormann S, Nadais P, Sousa F, Pinto M, Martins M, Sousa B, Fidalgo F, Soares C. Accumulation of Proline in Plants under Contaminated Soils—Are We on the Same Page? Antioxidants. 2023; 12(3):666. https://doi.org/10.3390/antiox12030666
Chicago/Turabian StyleSpormann, Sofia, Pedro Nadais, Filipa Sousa, Mafalda Pinto, Maria Martins, Bruno Sousa, Fernanda Fidalgo, and Cristiano Soares. 2023. "Accumulation of Proline in Plants under Contaminated Soils—Are We on the Same Page?" Antioxidants 12, no. 3: 666. https://doi.org/10.3390/antiox12030666