
AD−1 Small Molecule Improves Learning and Memory Function in Scopolamine-Induced Amnesic Mice Model through Regulation of CREB/BDNF and NF-κB/MAPK Signaling Pathway

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
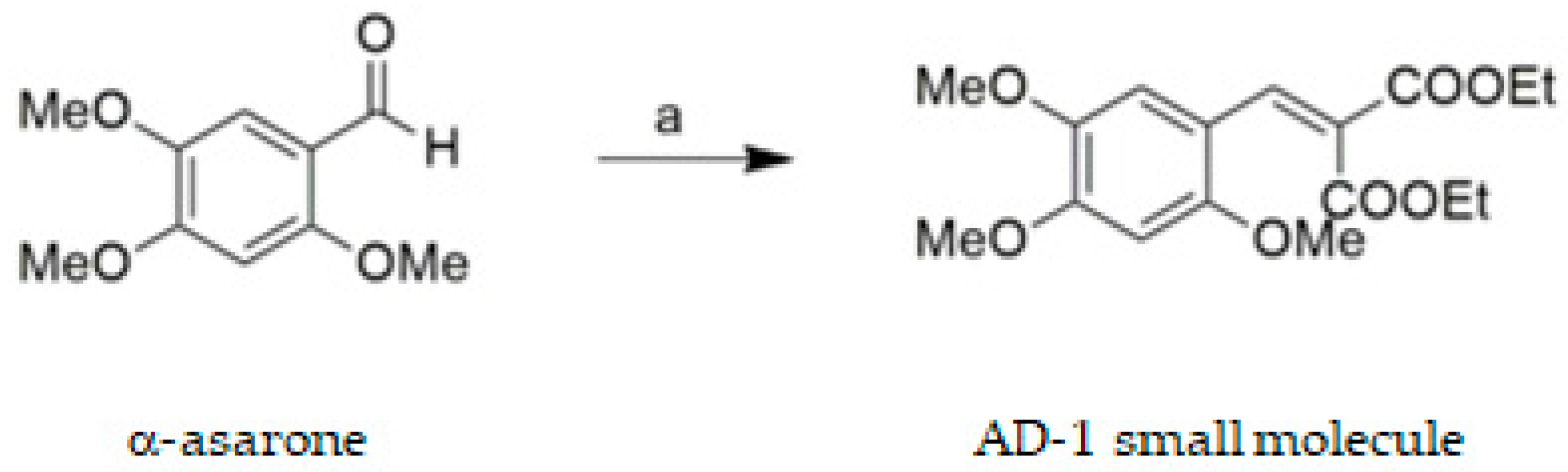
2.2. Synthesis of AD−1 Small Molecule
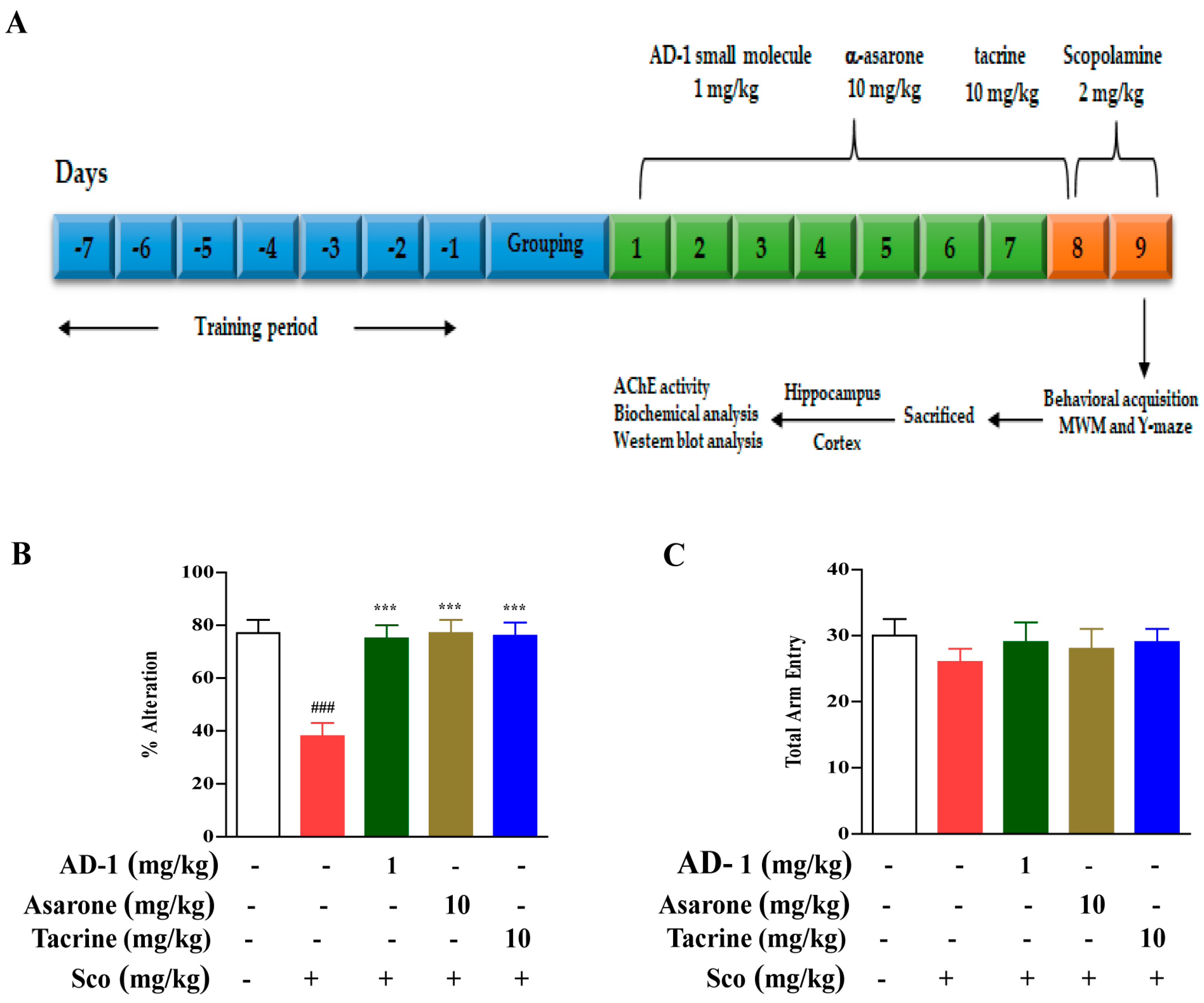
2.3. Animals Handling and Treatment
2.4. Y-Maze Test
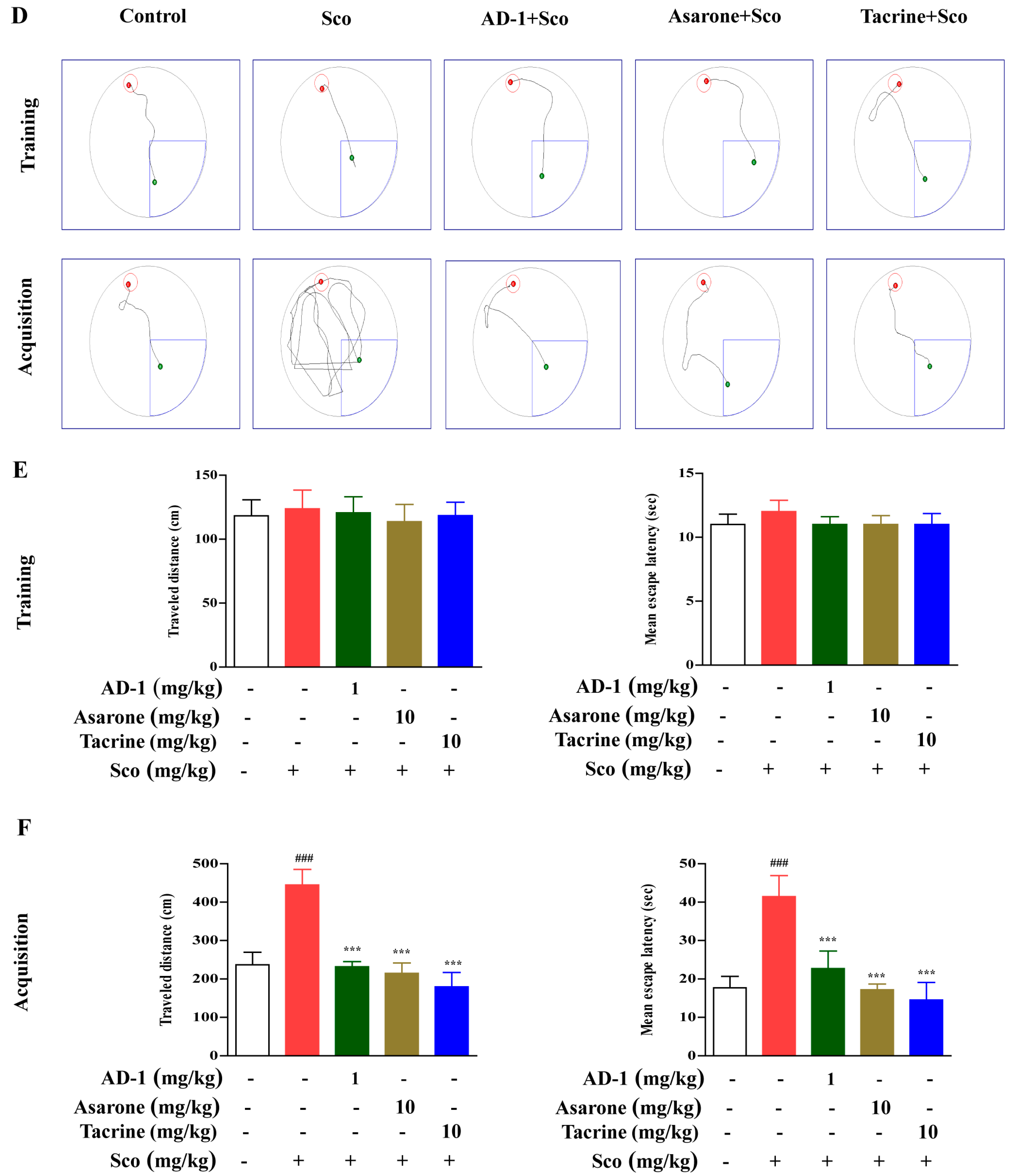
2.5. Morris Water Maze Test
2.6. Tissue Preparation
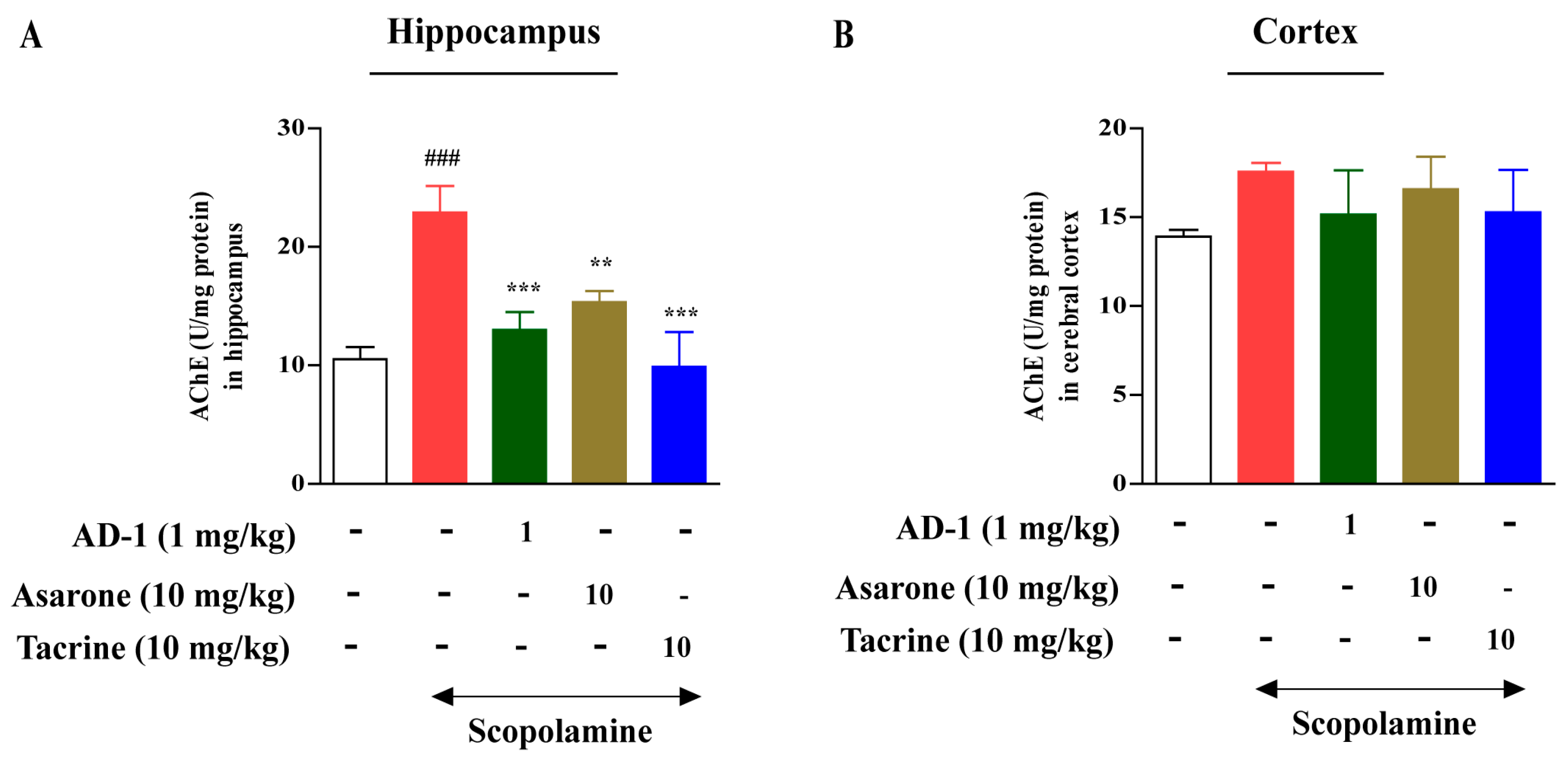
2.7. Measurement of AChE Activity
2.8. Measurement of Lipid Peroxidation, GSH, SOD, Catalase, and GPx Activities
2.9. Western Blotting
2.10. Genetic Toxicity Test
2.11. Statistical Analysis
3. Results
3.1. Genotoxicity Evaluation of AD−1 Small Molecule
3.2. Effect of AD−1 Small Molecule on Cognitive and memory impairments in the Scopolamine-Treated Mice
3.3. Effect of AD−1 Small Molecule on AChE Activity in the Hippocampus and Cortex of Scopolamine-Treated Mice
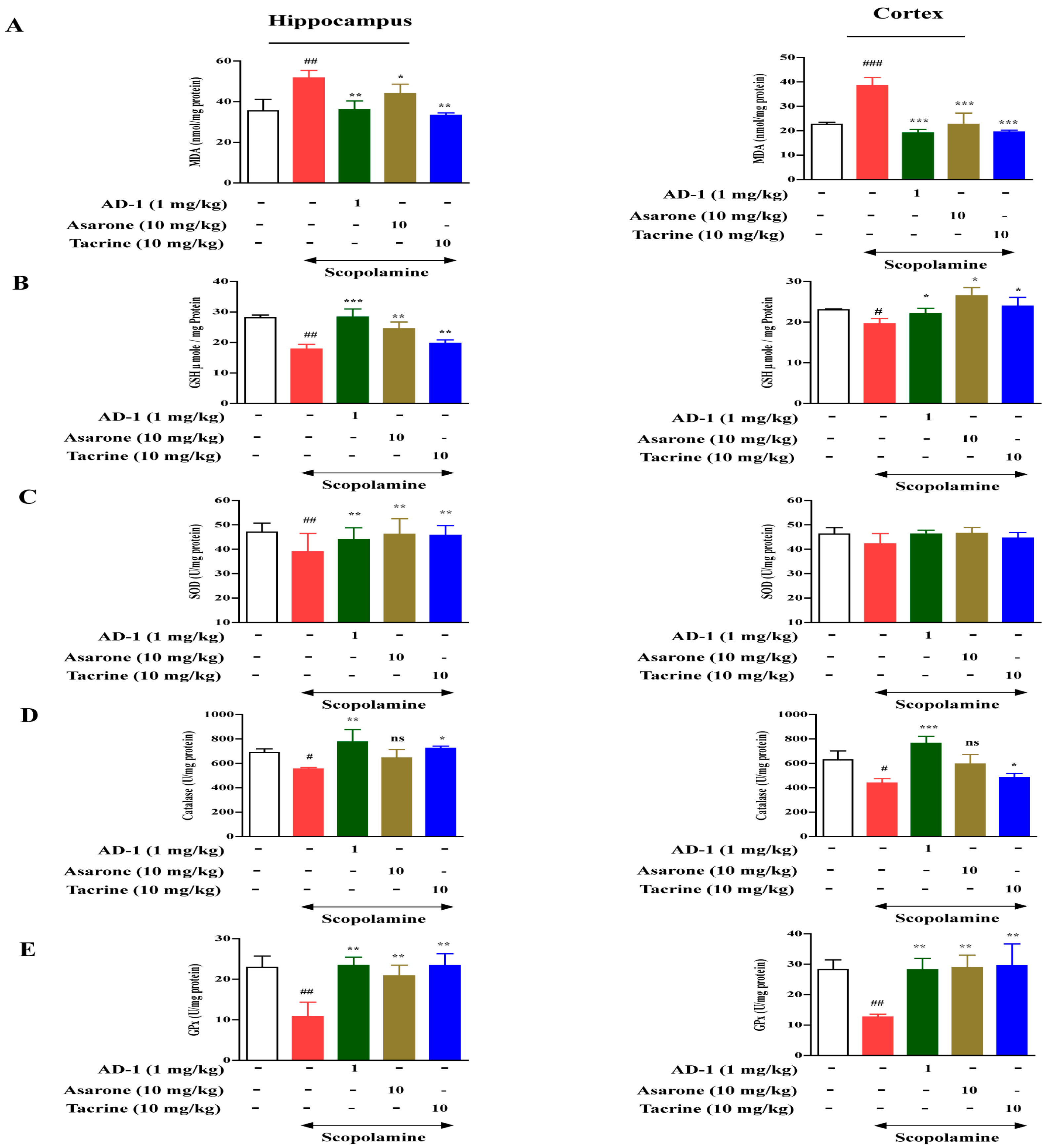
3.4. Effect of AD−1 small molecule on Lipid Peroxidation and Antioxidant Biomarkers in the Hippocampus and Cortex of Scopolamine-Treated Mice
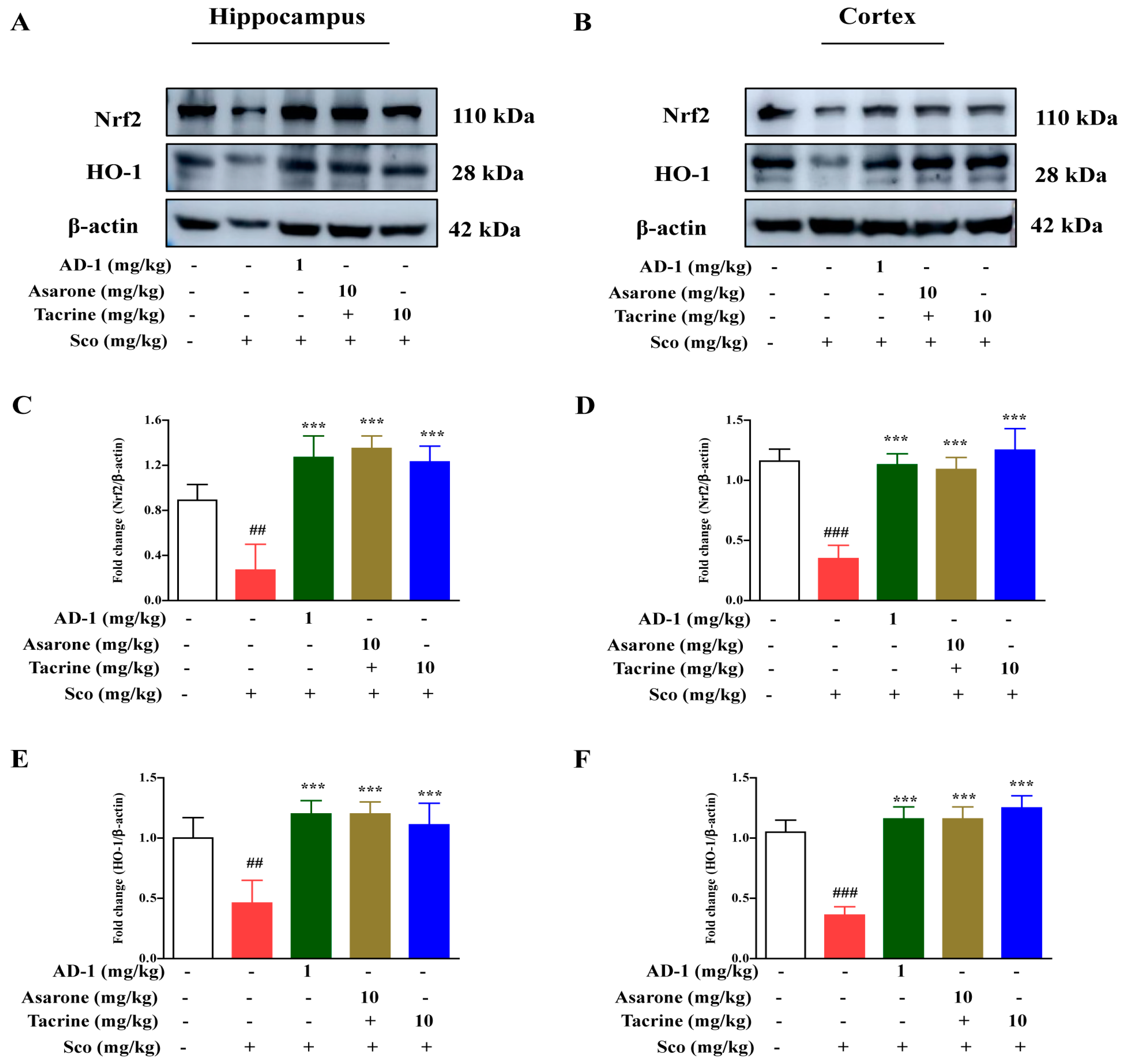
3.5. Effect of AD−1 Small Molecule on Nrf2 and HO-1 Expression in the Hippocampus and Cortex of Scopolamine-Treated Mice
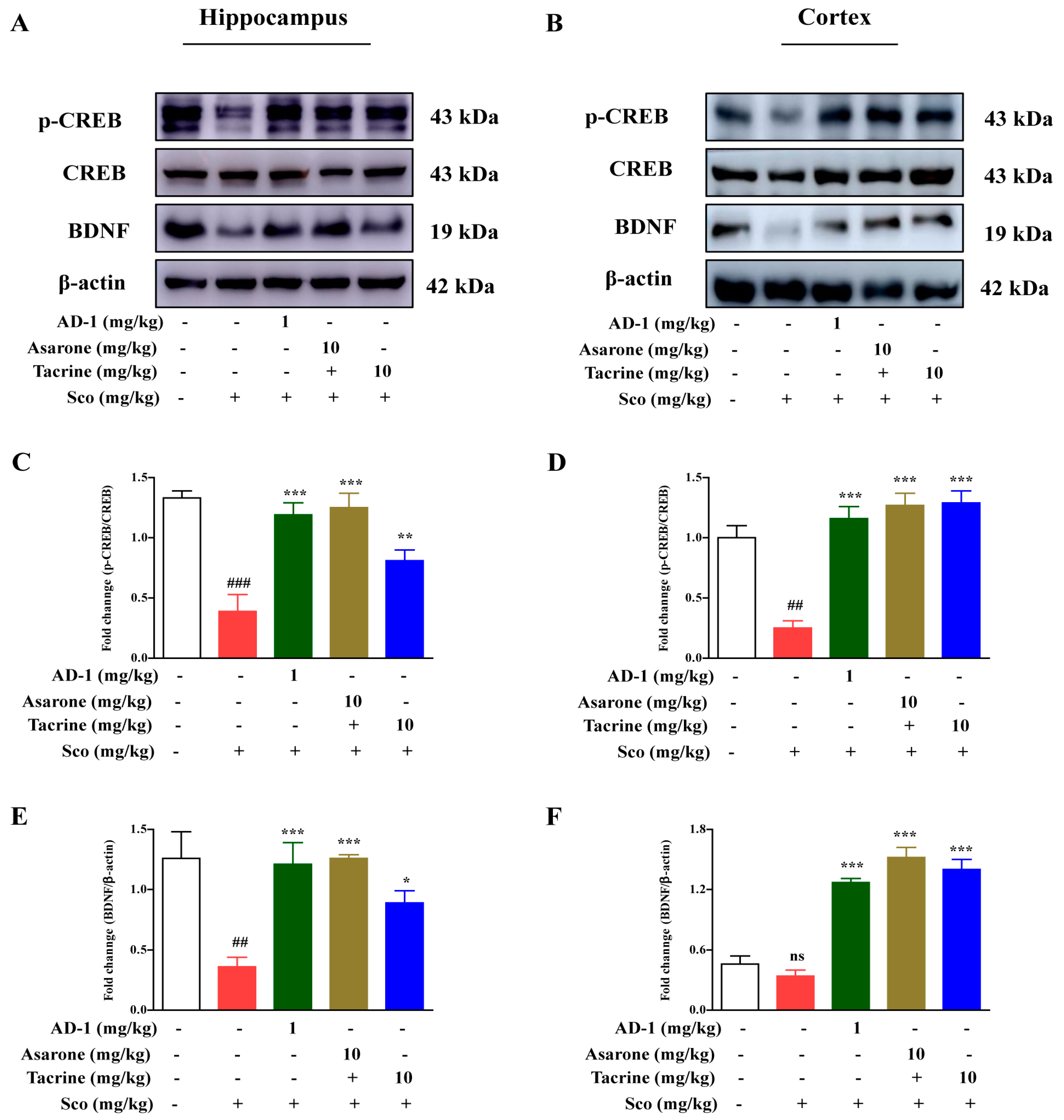
3.6. Effect of AD−1 Small Molecule on BDNF and CREB Expression in the Hippocampus and Cortex of Scopolamine-Treated Mice
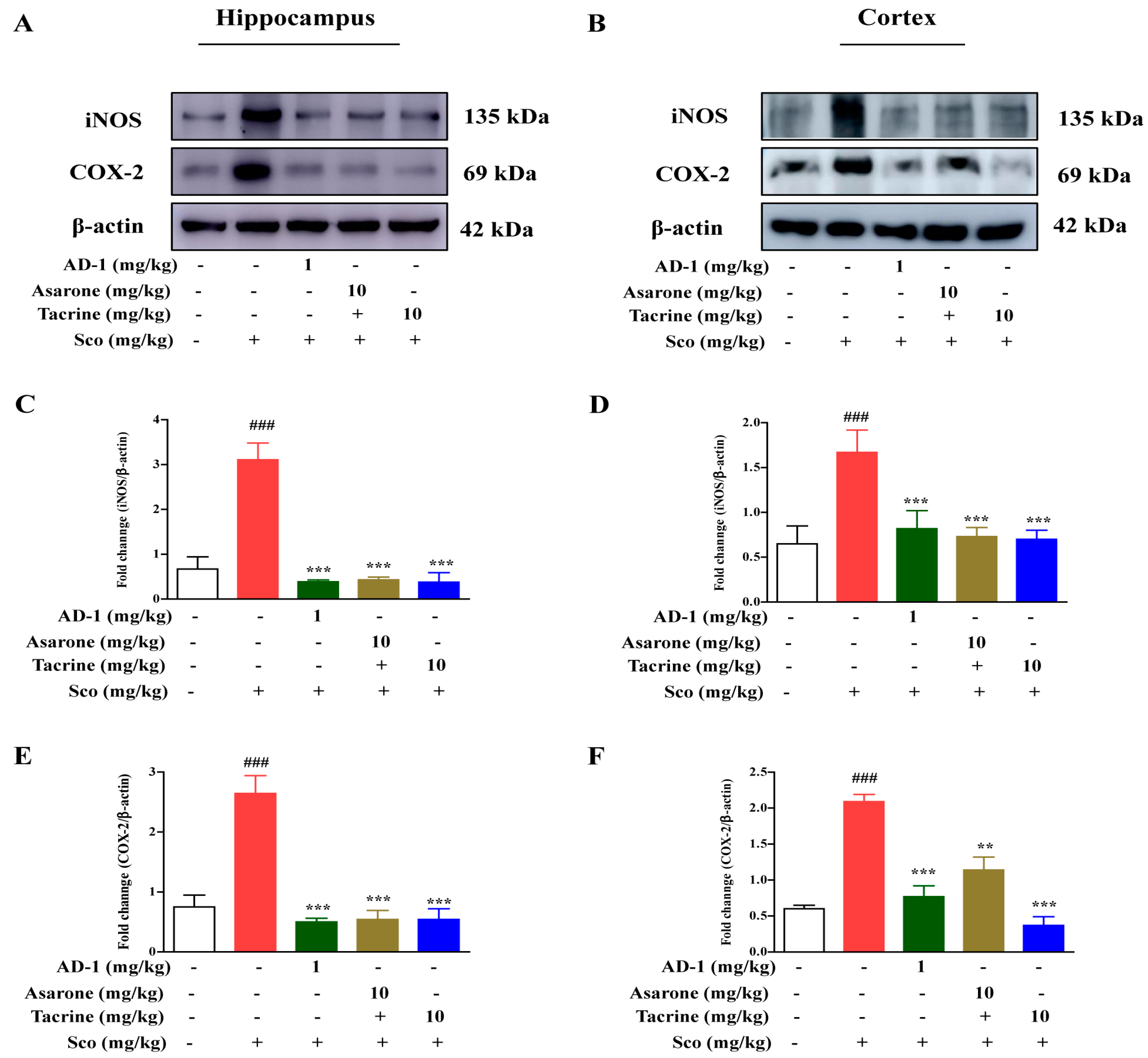
3.7. Effect of AD−1 small molecule on iNOS and COX-2 expression in the Hippocampus and Cortex of Scopolamine-Treated Mice
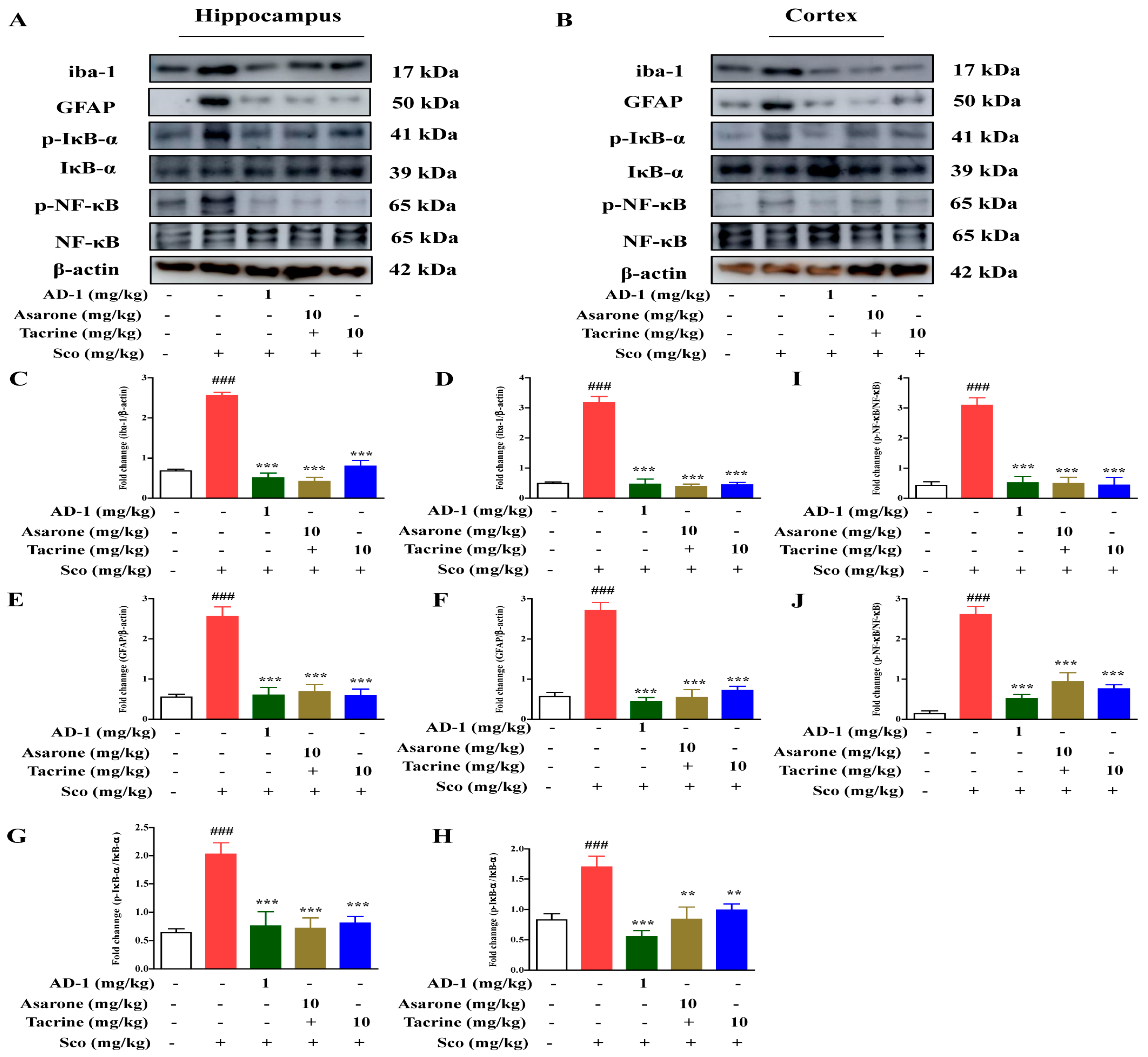
3.8. Effect of AD−1 small molecule on Glial Activation and NF-κB-Mediated Neuroinflammation in the Hippocampus and Cortex of Scopolamine-Treated Mice
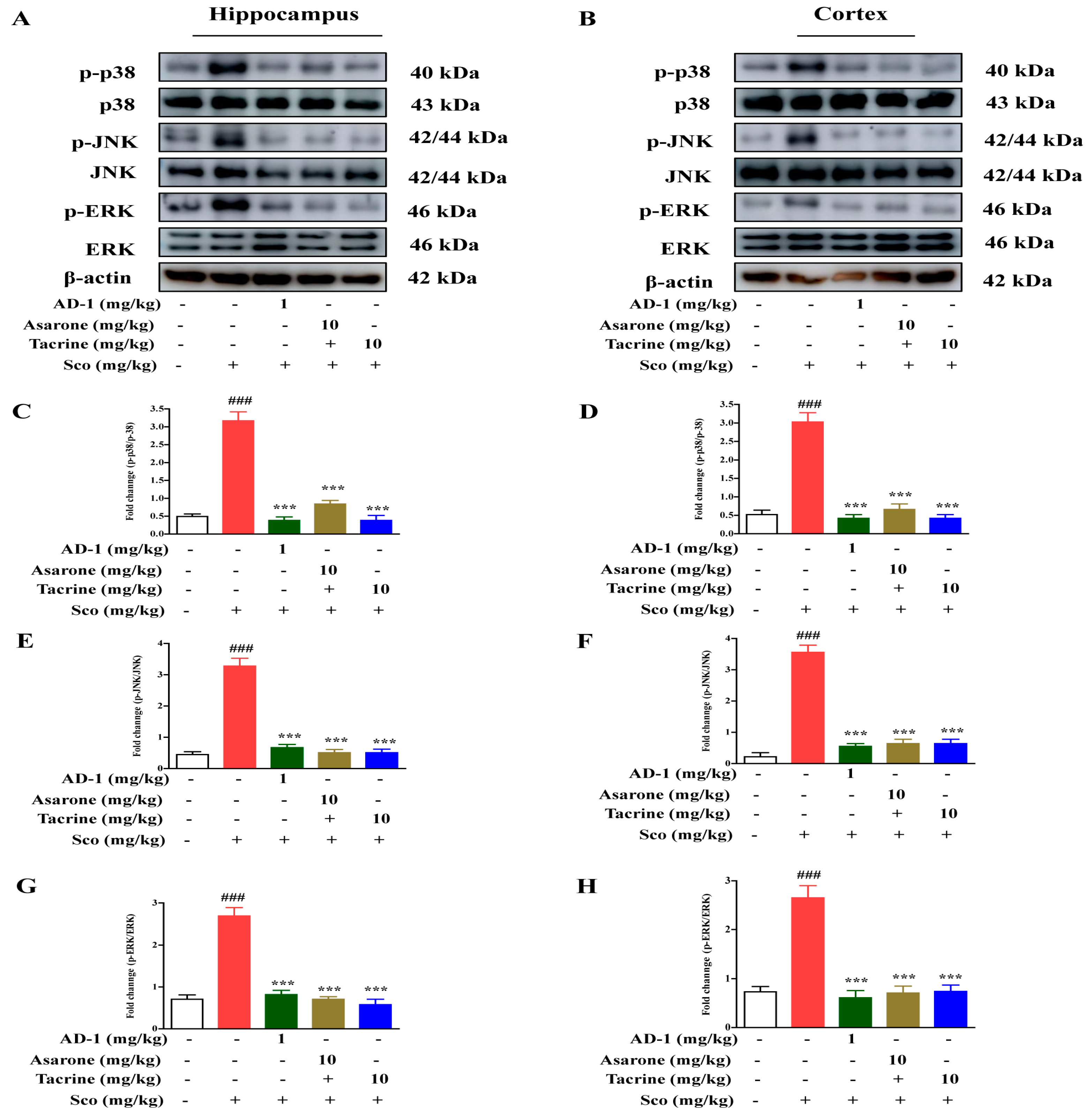
3.9. Effect of AD−1 Small Molecule on MAPK Signaling in the Hippocampus and Cortex of Scopolamine-Treated Mice
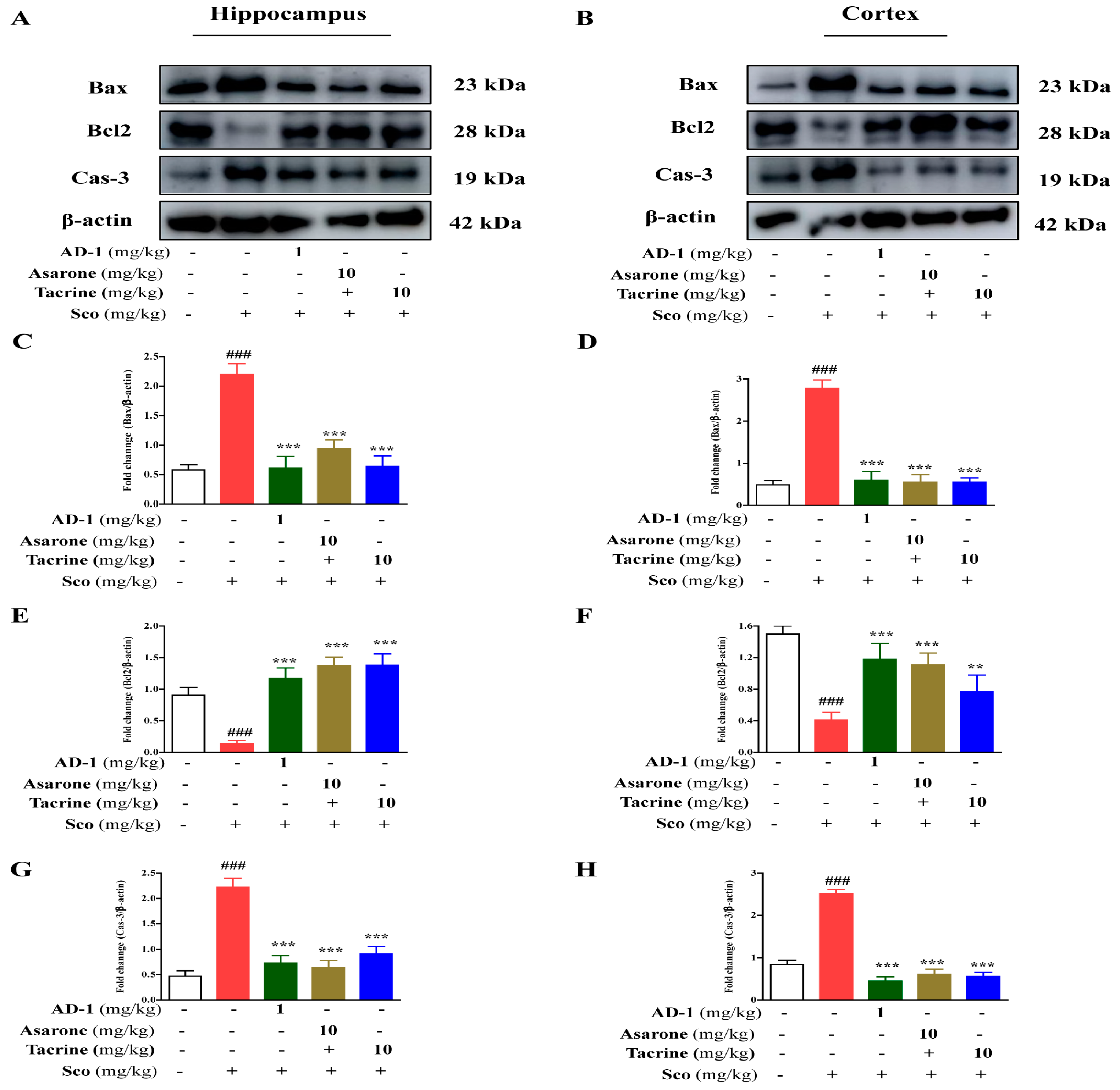
3.10. Effect of AD−1 Small Molecule on Neuronal Apoptosis in the Hippocampus and Cortex of Scopolamine-Treated Mice
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wan, T.; Wang, Z.; Luo, Y.; Zhang, Y.; He, W.; Mei, Y.; Xue, J.; Li, M.; Pan, H.; Li, W.; et al. FA-97, a New Synthetic Caffeic Acid Phenethyl Ester Derivative, Protects against Oxidative Stress-Mediated Neuronal Cell Apoptosis and Scopolamine-Induced Cognitive Impairment by Activating Nrf2/HO-1 Signaling. Oxid. Med. Cell. Longev. 2019, 2019, 8239642. [Google Scholar] [CrossRef] [PubMed]
- Karthivashan, G.; Kweon, M.H.; Park, S.Y.; Kim, J.S.; Kim, D.H.; Ganesan, P.; Choi, D.K. Cognitive-enhancing and ameliorative effects of acanthoside B in a scopolamine-induced amnesic mouse model through regulation of oxidative/inflammatory/cholinergic systems and activation of the TrkB/CREB/BDNF pathway. Food Chem. Toxicol. 2019, 129, 444–457. [Google Scholar] [CrossRef]
- Bajo, R.; Pusil, S.; López, M.E.; Canuet, L.; Pereda, E.; Osipova, D.; Maestú, F.; Pekkonen, E. Scopolamine effects on functional brain connectivity: A pharmacological model of Alzheimer’s disease. Sci. Rep. 2015, 5, 9748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piermartiri, T.C.B.; Figueiredo, C.P.; Rial, D.; Duarte, F.S.; Bezerra, S.C.; Mancini, G.; de Bem, A.F.; Prediger, R.D.S.; Tasca, C.I. Atorvastatin prevents hippocampal cell death, neuroinflammation and oxidative stress following amyloid-β(1-40) administration in mice: Evidence for dissociation between cognitive deficits and neuronal damage. Exp. Neurol. 2010, 226, 274–284. [Google Scholar] [CrossRef]
- Sharma, K. Cholinesterase inhibitors as Alzheimer’s therapeutics (Review). Mol. Med. Rep. 2019, 20, 1479–1487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reddy, P.H.; Oliver, D.M.A. Amyloid Beta and Phosphorylated Tau-Induced Defective Autophagy and Mitophagy in Alzheimer’s Disease. Cells 2019, 8, 488. [Google Scholar] [CrossRef] [Green Version]
- Cheon, S.Y.; Koo, B.N.; Kim, S.Y.; Kam, E.H.; Nam, J.; Kim, E.J. Scopolamine promotes neuroinflammation and delirium-like neuropsychiatric disorder in mice. Sci. Rep. 2021, 11, 8376. [Google Scholar] [CrossRef]
- Wong-Guerra, M.; Jiménez-Martin, J.; Pardo-Andreu, G.L.; Fonseca-Fonseca, L.A.; Souza, D.O.; de Assis, A.M.; Ramirez-Sanchez, J.; del Valle, R.M.S.; Nuñez-Figueredo, Y. Mitochondrial involvement in memory impairment induced by scopolamine in rats. Neurol. Res. 2017, 39, 649–659. [Google Scholar] [CrossRef]
- Upadhyay, P.; Sadhu, A.; Singh, P.K.; Agrawal, A.; Ilango, K.; Purohit, S.; Dubey, G.P. Revalidation of the neuroprotective effects of a United States patented polyherbal formulation on scopolamine induced learning and memory impairment in rats. Biomed. Pharmacother. 2018, 97, 1046–1052. [Google Scholar] [CrossRef]
- Antonova, E.; Parslow, D.; Brammer, M.; Simmons, A.; Williams, S.; Dawson, G.R.; Morris, R. Scopolamine disrupts hippocampal activity during allocentric spatial memory in humans: An fMRI study using a virtual reality analogue of the Morris Water Maze. J. Psychopharmacol. 2011, 25, 1256–1265. [Google Scholar] [CrossRef]
- Pachauri, S.D.; Tota, S.; Khandelwal, K.; Verma, P.R.P.; Nath, C.; Hanif, K.; Shukla, R.; Saxena, J.K.; Dwivedi, A.K. Protective effect of fruits of Morinda citrifolia L. on scopolamine induced memory impairment in mice: A behavioral, biochemical and cerebral blood flow study. J. Ethnopharmacol. 2012, 139, 34–41. [Google Scholar] [CrossRef] [PubMed]
- Budzynska, B.; Boguszewska-Czubara, A.; Kruk-Slomka, M.; Skalicka-Wozniak, K.; Michalak, A.; Musik, I.; Biala, G. Effects of imperatorin on scopolamine-induced cognitive impairment and oxidative stress in mice. Psychopharmacology 2015, 232, 931–942. [Google Scholar] [CrossRef]
- Subedi, L.; Cho, K.; Park, Y.U.; Choi, H.J.; Kim, S.Y. Sulforaphane-Enriched Broccoli Sprouts Pretreated by Pulsed Electric Fields Reduces Neuroinflammation and Ameliorates Scopolamine-Induced Amnesia in Mouse Brain through Its Antioxidant Ability via Nrf2-HO-1 Activation. Oxid. Med. Cell. Longev. 2019, 2019, 35492740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, C.D.; Yoo, J.; Hwang, S.Y.; Cho, S.Y.; Kim, M.; Jang, H.; No, K.O.; Shin, J.C.; Kim, J.H.; Lee, G. Bee Venom Activates the Nrf2/HO-1 and TrkB/CREB/BDNF Pathways in Neuronal Cell Responses against Oxidative Stress Induced by Aβ1-42. Int. J. Mol. Sci. 2022, 23, 1193. [Google Scholar] [CrossRef]
- Zhu, X.; Castellani, R.J.; Takeda, A.; Nunomura, A.; Atwood, C.S.; Perry, G.; Smith, M.A. Differential activation of neuronal ERK, JNK/SAPK and p38 in Alzheimer disease: The “two hit” hypothesis. Mech. Ageing Dev. 2001, 123, 39–46. [Google Scholar] [CrossRef] [PubMed]
- Giese, K.P.; Mizuno, K. The roles of protein kinases in learning and memory. Learn. Mem. 2013, 20, 540–552. [Google Scholar] [CrossRef] [Green Version]
- Venkatesan, K. Anti-amnesic and anti-cholinesterase activities of α-asarone against scopolamine-induced memory impairments in rats. Eur. Rev. Med. Pharmacol. Sci. 2022, 26, 6344–6350. [Google Scholar]
- Zeng, L.; Zhang, D.; Liu, Q.; Zhang, J.; Mu, K.; Gao, X.; Zhang, K.; Li, H.; Wang, Q.; Zheng, Y.; et al. Alpha-asarone Improves Cognitive Function of APP/PS1 Mice and Reducing Aβ42, P-tau and Neuroinflammation, and Promoting Neuron Survival in the Hippocampus. Neuroscience 2021, 458, 141–152. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.W.; Koppula, S.; Kumar, H.; Park, J.Y.; Kim, I.W.; More, S.V.; Kim, I.S.; Han, S.D.; Kim, S.K.; Yoon, S.H.; et al. α-Asarone attenuates microglia-mediated neuroinflammation by inhibiting NF kappa B activation and mitigates MPTP-induced behavioral deficits in a mouse model of Parkinson’s disease. Neuropharmacology 2015, 97, 46–57. [Google Scholar] [CrossRef]
- Jo, S.H.; Kang, T.B.; Koppula, S.; Cho, D.Y.; Kim, J.S.; Kim, I.S.; Choi, D.K. Mitigating Effect of Lindera obtusiloba Blume Extract on Neuroinflammation in Microglial Cells and Scopolamine-Induced Amnesia in Mice. Molecules 2021, 26, 2870. [Google Scholar] [CrossRef]
- Karthivashan, G.; Park, S.Y.; Kweon, M.H.; Kim, J.; Haque, M.E.; Cho, D.Y.; Kim, I.S.; Cho, E.A.; Ganesan, P.; Choi, D.K. Ameliorative potential of desalted Salicornia europaea L. extract in multifaceted Alzheimer’s-like scopolamine-induced amnesic mice model. Sci. Rep. 2018, 8, 7174. [Google Scholar] [CrossRef] [PubMed]
- Khurana, K.; Kumar, M.; Bansal, N. Lacidipine Prevents Scopolamine-Induced Memory Impairment by Reducing Brain Oxido-nitrosative Stress in Mice. Neurotox. Res. 2021, 39, 1087–1102. [Google Scholar] [CrossRef]
- Saura, C.A.; Cardinaux, J.R. Emerging Roles of CREB-Regulated Transcription Coactivators in Brain Physiology and Pathology. Trends Neurosci. 2017, 40, 720–733. [Google Scholar] [CrossRef] [Green Version]
- Hseu, Y.C.; Wu, F.Y.; Wu, J.J.; Chen, J.Y.; Chang, W.H.; Lu, F.J.; Lai, Y.C.; Yang, H.L. Anti-inflammatory potential of Antrodia Camphorata through inhibition of iNOS, COX-2 and cytokines via the NF-kappaB pathway. Int. Immunopharmacol. 2005, 5, 1914–1925. [Google Scholar] [CrossRef] [PubMed]
- Maqbool, A.; Lattke, M.; Wirth, T.; Baumann, B. Sustained, neuron-specific IKK/NF-κB activation generates a selective neuroinflammatory response promoting local neurodegeneration with aging. Mol. Neurodegener. 2013, 8, 40. [Google Scholar] [CrossRef] [Green Version]
- Kim, E.A.; Choi, J.; Han, A.R.; Cho, C.H.; Choi, S.Y.; Ahn, J.Y.; Cho, S.W. 2-Cyclopropylimino-3-methyl-1,3-thiazoline hydrochloride inhibits microglial activation by suppression of nuclear factor-kappa B and mitogen-activated protein kinase signaling. J. Neuroimmune Pharmacol. 2014, 9, 461–467. [Google Scholar] [CrossRef]
- Wagner, E.F.; Nebreda, Á.R. Signal integration by JNK and p38 MAPK pathways in cancer development. Nat. Rev. Cancer 2009, 9, 537–549. [Google Scholar] [CrossRef]
- Cuenda, A.; Rousseau, S. p38 MAP-kinases pathway regulation, function and role in human diseases. Biochim. Biophys. Acta 2007, 1773, 1358–1375. [Google Scholar] [CrossRef] [Green Version]
- Dhanasekaran, D.N.; Reddy, E.P. JNK signaling in apoptosis. Oncogene 2008, 27, 6245–6251. [Google Scholar] [CrossRef] [PubMed] [Green Version]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Chemical Name | Origin | (Q)SAR Model | Conclusion | ICH M7 *** Classification | |
|---|---|---|---|---|---|
| Derek * | Sarah ** | ||||
| 2,4,5-trimethoxy benzaldehyde | Starting material | Inactive | Negative | Negative | Class 5 |
| Diethylmalonate | Reaction material | Inactive | Negative | Negative | Class 5 |
| AD−1 | - | Inactive | Negative | Negative | Class 5 |
| AD−1 Residual Solvent(ppm) | ||||
|---|---|---|---|---|
| Synthetic solvent | Dichloromethane | Hexane | Ethyl acetate | Toluene |
| USP limit * | 600 | 290 | 5000 | 890 |
| Result | 919 | 1683 | 581 | 240 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Balakrishnan, R.; Park, J.-Y.; Cho, D.-Y.; Ahn, J.-Y.; Yoo, D.-S.; Seol, S.-H.; Yoon, S.-H.; Choi, D.-K. AD−1 Small Molecule Improves Learning and Memory Function in Scopolamine-Induced Amnesic Mice Model through Regulation of CREB/BDNF and NF-κB/MAPK Signaling Pathway. Antioxidants 2023, 12, 648. https://doi.org/10.3390/antiox12030648
Balakrishnan R, Park J-Y, Cho D-Y, Ahn J-Y, Yoo D-S, Seol S-H, Yoon S-H, Choi D-K. AD−1 Small Molecule Improves Learning and Memory Function in Scopolamine-Induced Amnesic Mice Model through Regulation of CREB/BDNF and NF-κB/MAPK Signaling Pathway. Antioxidants. 2023; 12(3):648. https://doi.org/10.3390/antiox12030648
Chicago/Turabian StyleBalakrishnan, Rengasamy, Ju-Young Park, Duk-Yeon Cho, Jae-Yong Ahn, Dong-Sun Yoo, Sang-Ho Seol, Sung-Hwa Yoon, and Dong-Kug Choi. 2023. "AD−1 Small Molecule Improves Learning and Memory Function in Scopolamine-Induced Amnesic Mice Model through Regulation of CREB/BDNF and NF-κB/MAPK Signaling Pathway" Antioxidants 12, no. 3: 648. https://doi.org/10.3390/antiox12030648