

Anti-Osteoarthritic Effects of Prunella Vulgaris and Gentiana Lutea In Vitro and In Vivo
 , , , , and
, , , , and 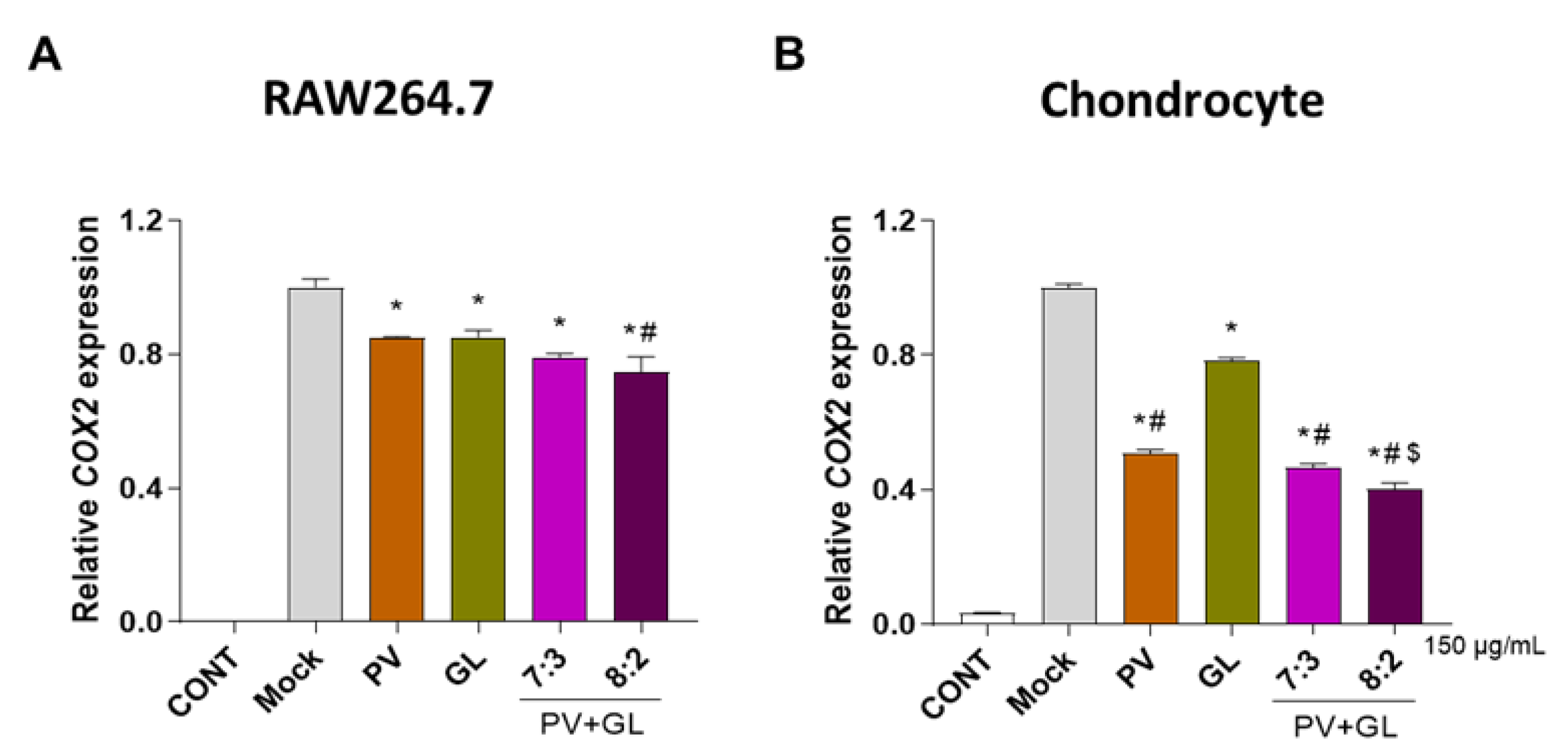{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Extraction and Confirmation of Standards Compounds from PV and GL Extract
2.2. Cell Culture and Primary Chondrocyte Isolation
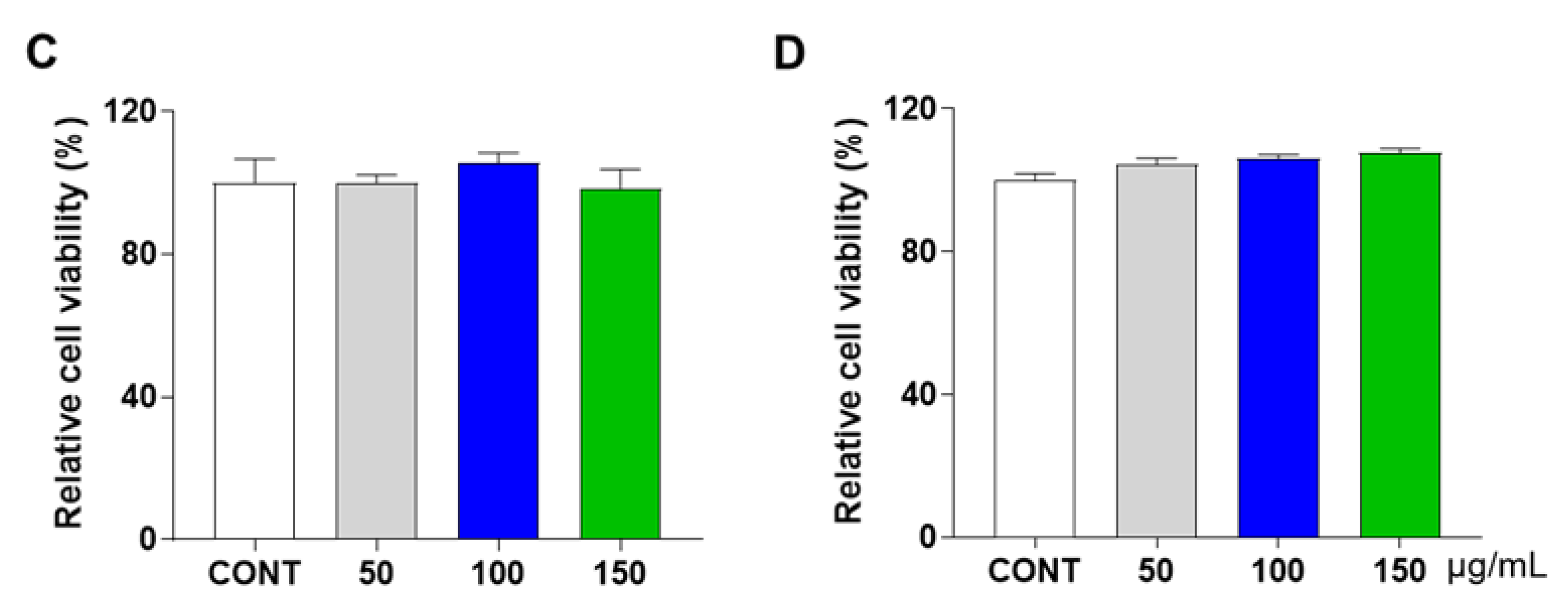
2.3. Water-Soluble Tetrazolium Salt (WST) Assay
2.4. Reverse Transcription-Polymerase Chain Reaction (RT-PCR) and Quantitative Reverse-Transcription PCR (qRT-PCR)
2.5. Western Blot Analysis
2.6. Destabilization of Medial Meniscus (DMM) Mouse Model and Micro-CT Analysis
2.7. Histology
2.8. NO and Plasma Cytokine Analysis
2.9. Statistical Analysis
3. Results
3.1. PV and GL Mixtures Inhibit COX2 mRNA Expression
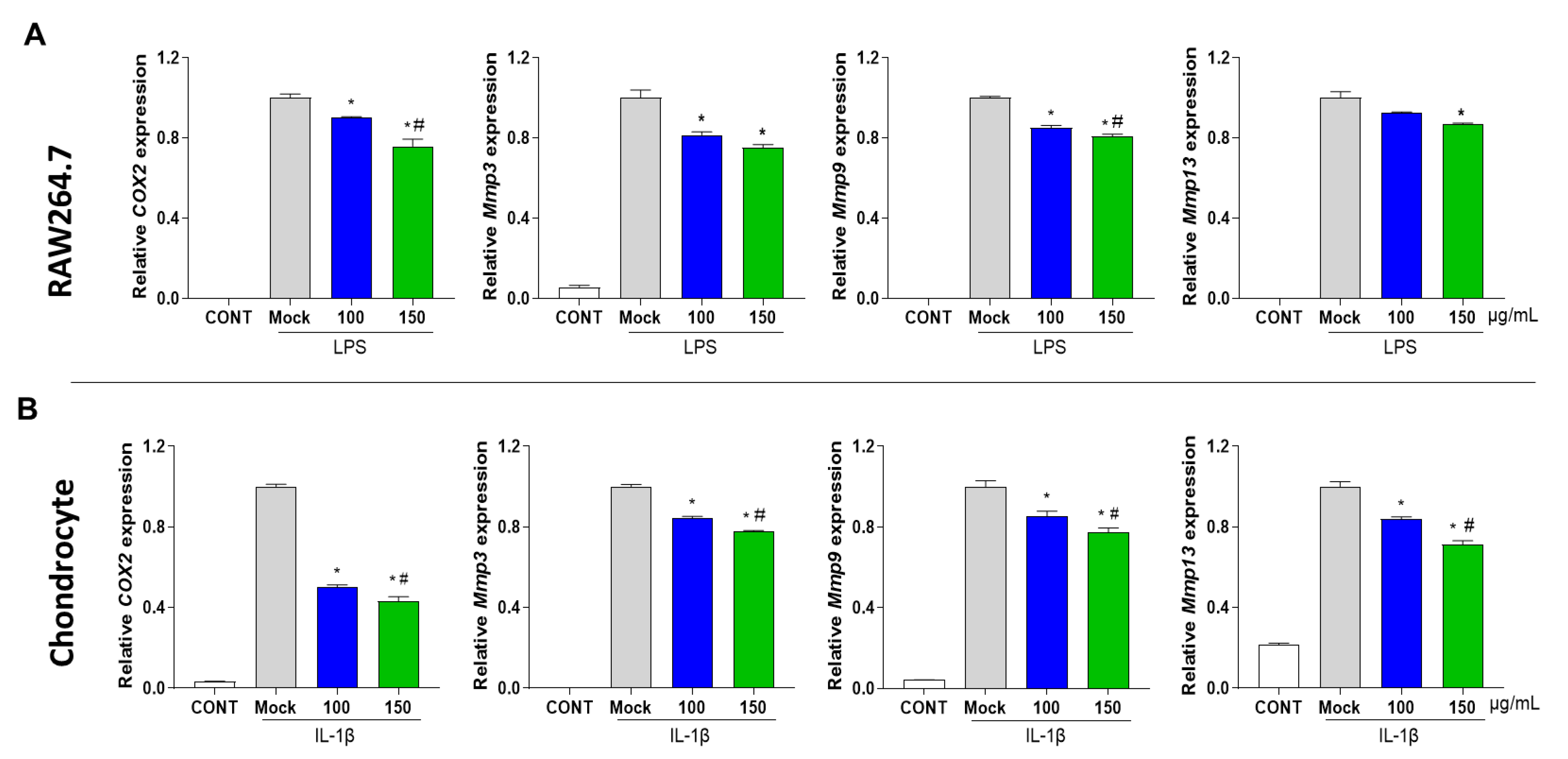
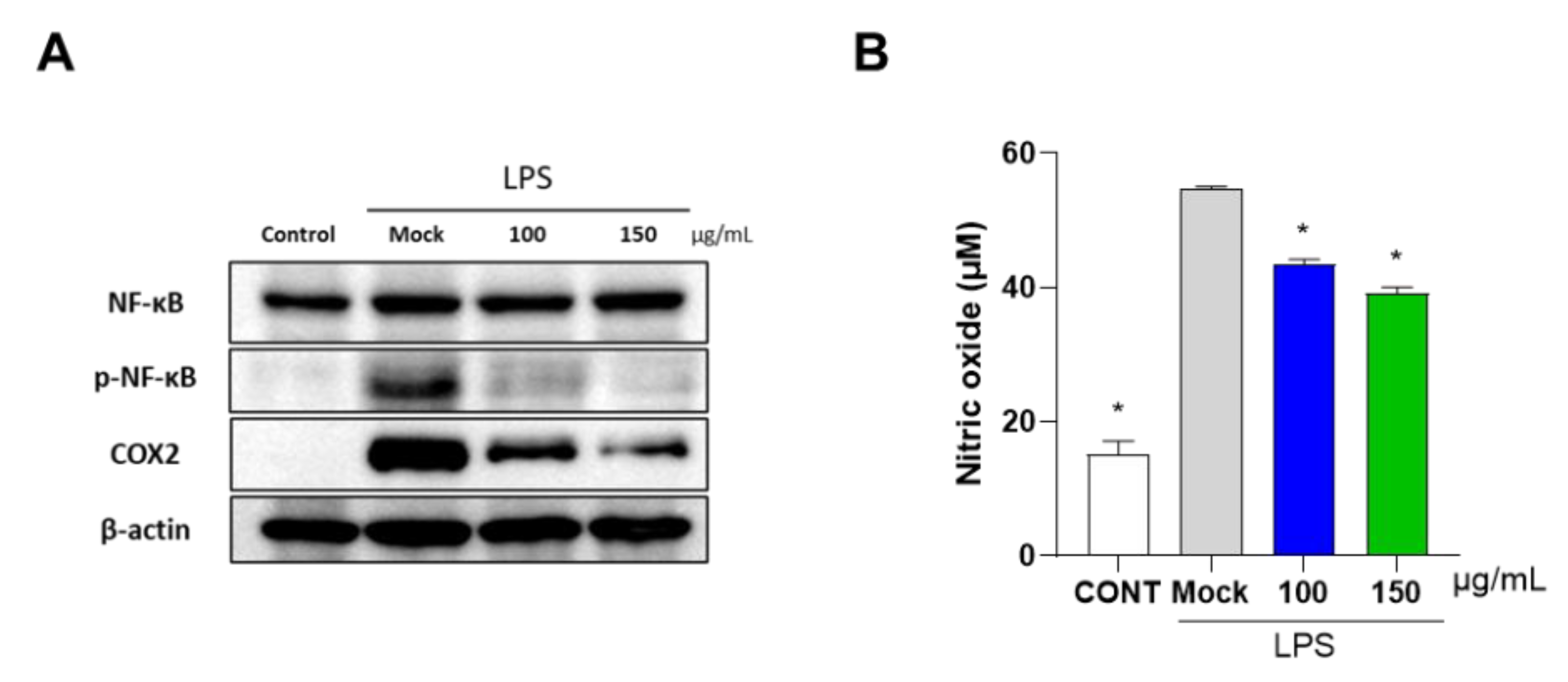
3.2. PG Decreases Inflammation in Mouse Primary Chondrocytes and RAW 264.7 Cells
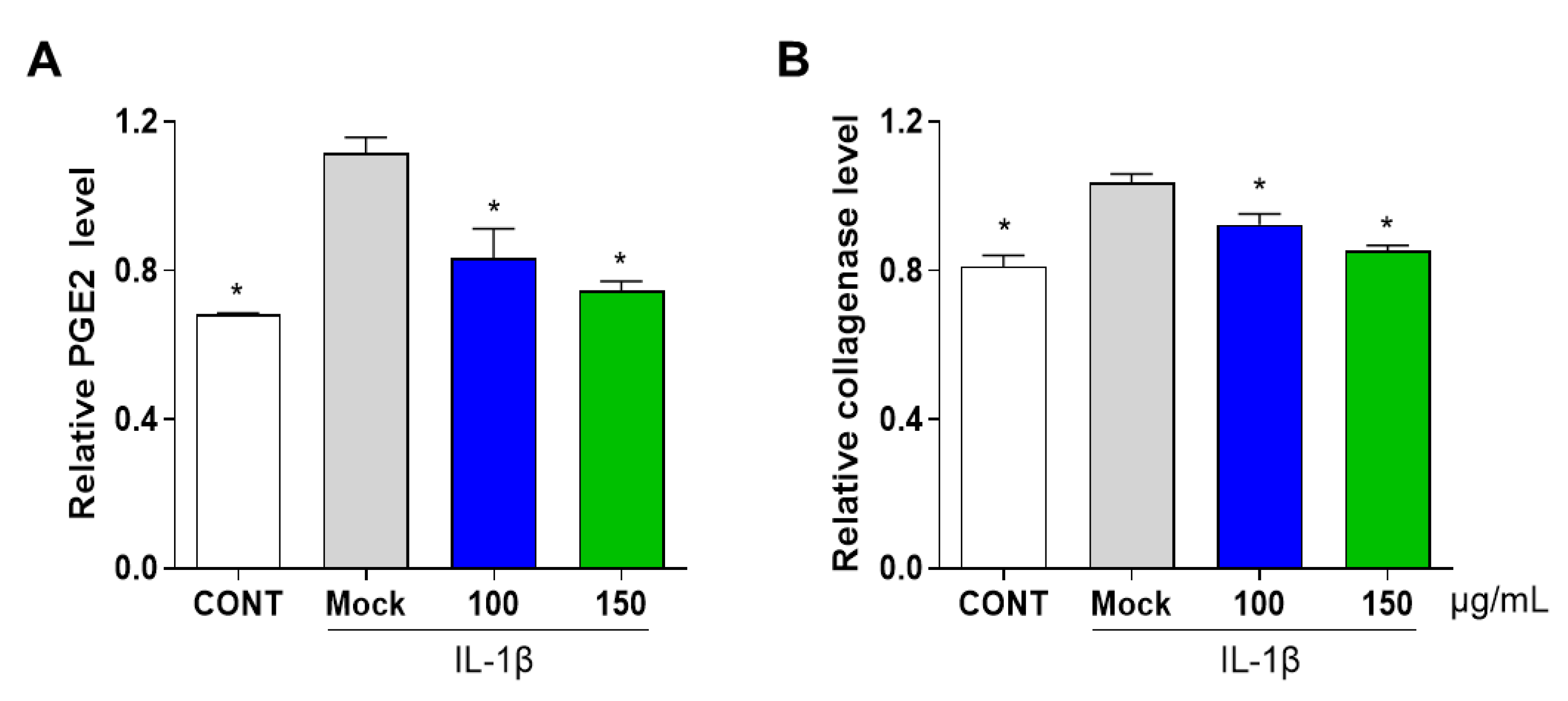
3.3. PG Inhibits Secretion of Prostaglandin E2 (PGE2) and Collagenase in IL-1β-Induced Primary Chondrocyte Inflammation
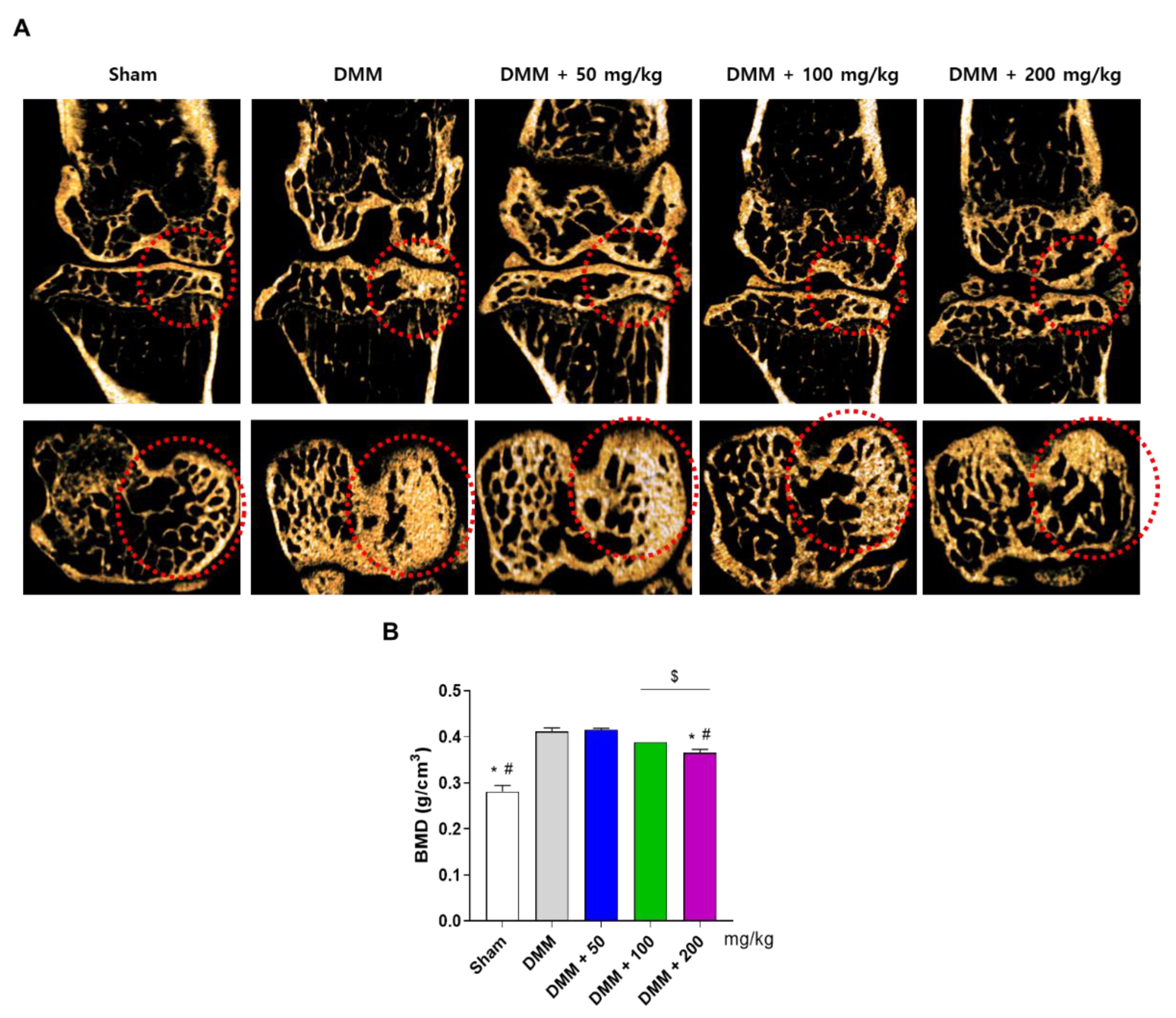
3.4. PG Prevents Subchondral Sclerosis in the DMM-Induced OA Mouse Model
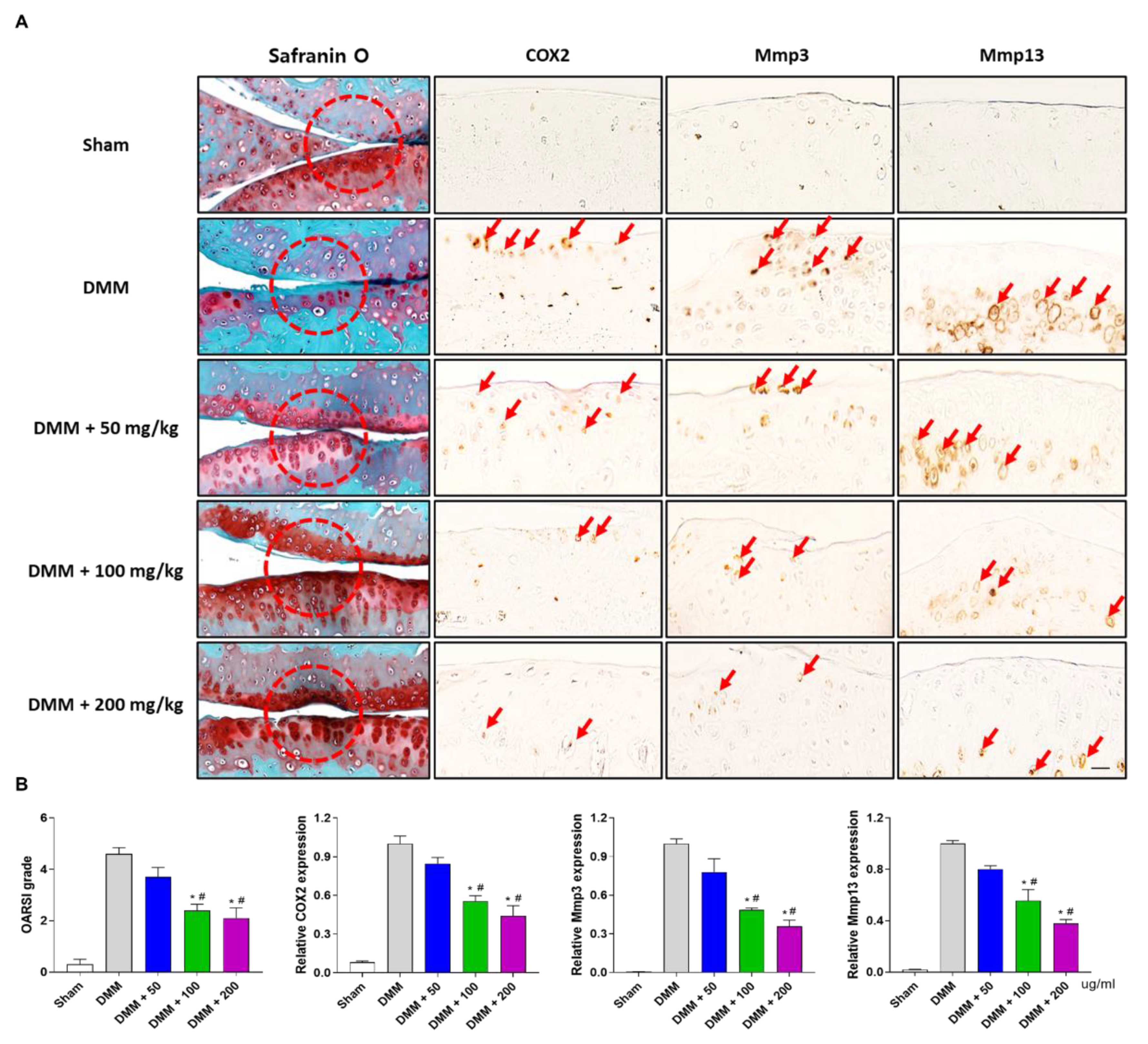
3.5. PG Reduces Cartilage Destruction in DMM-Induced OA Mouse Model
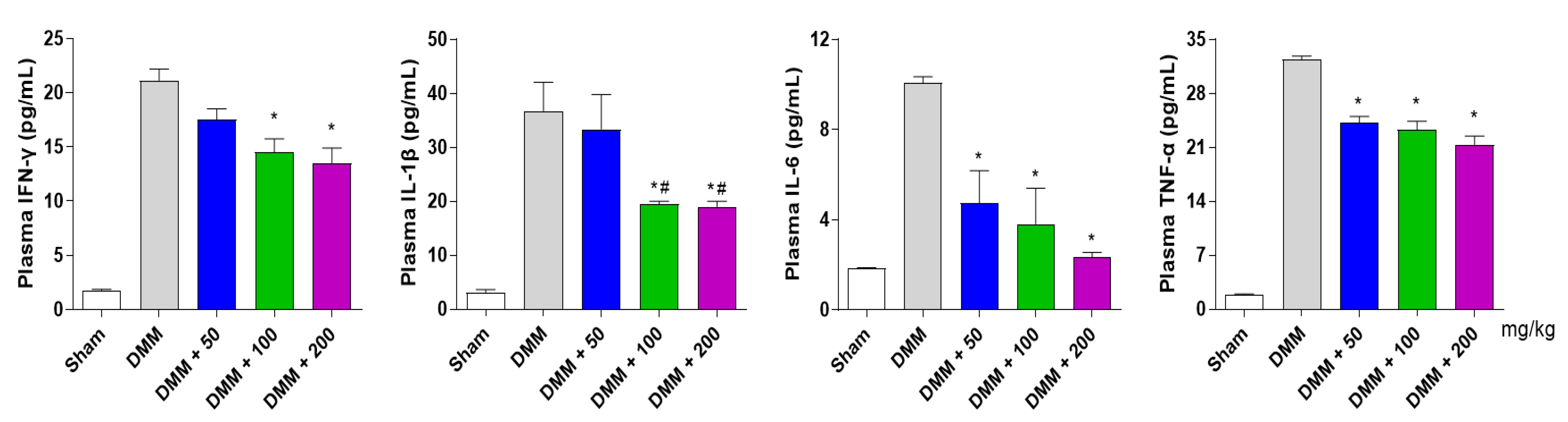
3.6. PG Decreases Inflammatory Cytokines in the DMM-Induced OA Mouse Model
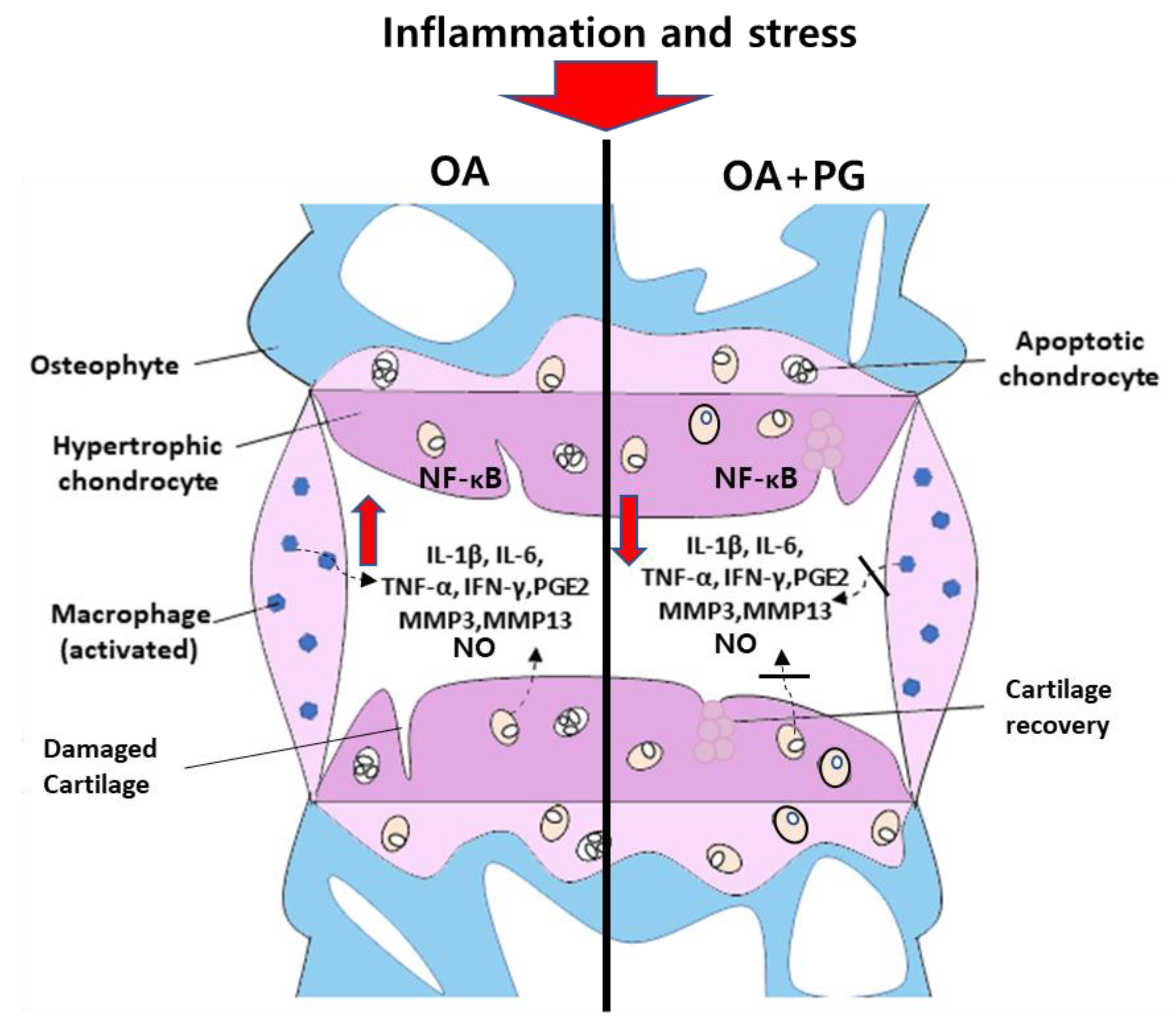
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mora, J.C.; Przkora, R.; Cruz-Almeida, Y. Knee osteoarthritis: Pathophysiology and current treatment modalities. J. Pain Res. 2018, 11, 2189–2196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, L. Osteoarthritis of the knee. N. Engl. J. Med. 2021, 384, 51–59. [Google Scholar] [CrossRef] [PubMed]
- Mobasheri, A.; Batt, M. An update on the pathophysiology of osteoarthritis. Ann. Phys. Rehabil. Med. 2016, 59, 333–339. [Google Scholar] [CrossRef] [PubMed]
- Xia, B.; Di, C.; Zhang, J.; Hu, S.; Jin, H.; Tong, P. Osteoarthritis pathogenesis: A review of molecular mechanisms. Calcif. Tissue Int. 2014, 95, 495–505. [Google Scholar] [CrossRef] [Green Version]
- Jenei-Lanzl, Z.; Meurer, A.; Zaucke, F. Interleukin-1β signaling in osteoarthritis-chondrocytes in focus. Cell. Signal. 2019, 53, 212–223. [Google Scholar] [CrossRef]
- Loeser, R.F.; Erickson, E.A.; Long, D.L. Mitogen-activated protein kinases as therapeutic targets in osteoarthritis. Curr. Opin. Rheumatol. 2008, 20, 581–586. [Google Scholar] [CrossRef]
- Li, X.; Wu, D.; Hu, Z.; Xuan, J.; Ding, X.; Zheng, G.; Feng, Z.; Ni, W.; Wu, A. The protective effect of ligustilide in osteoarthritis: An in vitro and in vivo study. Cell. Physiol. Biochem. 2018, 48, 2583–2595. [Google Scholar] [CrossRef]
- Montaseri, A.; Busch, F.; Mobasheri, A.; Buhrmann, C.; Aldinger, C.; Rad, J.S.; Shakibaei, M. IGF-1 and PDGF-bb suppress IL-1β-induced cartilage degradation through down-regulation of NF-κB signaling: Involvement of Src/PI-3K/AKT pathway. PLoS ONE 2011, 6, e28663. [Google Scholar] [CrossRef] [Green Version]
- Buhrmann, C.; Brockmueller, A.; Mueller, A.L.; Shayan, P.; Shakibaei, M. Curcumin attenuates environment-derived osteoarthritis by Sox9/NF-kB Signaling Axis. Int. J. Mol. Sci. 2021, 22, 7645. [Google Scholar] [CrossRef]
- Hunter, D.J.; Bierma-Zeinstra, S. Osteoarthritis. Lancet 2019, 393, 1745–1759. [Google Scholar] [CrossRef]
- Robinson, W.H.; Lepus, C.M.; Wang, Q.; Raghu, H.; Mao, R.; Lindstrom, T.M.; Sokolove, J. Low-grade inflammation as a key mediator of the pathogenesis of osteoarthritis. Nat. Rev. Rheumatol. 2016, 12, 580–592. [Google Scholar] [CrossRef] [PubMed]
- Katz, J.N.; Arant, K.R.; Loeser, R.F. Diagnosis and treatment of hip and knee osteoarthritis: A review. JAMA 2021, 325, 568–578. [Google Scholar] [CrossRef] [PubMed]
- Crofford, L.J. Use of NSAIDs in treating patients with arthritis. Arthritis Res. Ther. 2013, 15, S2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.; Meng, Q. Global research trends of herbal medicine for pain in three decades (1990–2019): A bibliometric analysis. J. Pain Res. 2021, 14, 1611–1626. [Google Scholar] [CrossRef] [PubMed]
- Panickar, K.S. Beneficial effects of herbs, spices and medicinal plants on the metabolic syndrome, brain and cognitive function. Cent. Nerv. Syst. Agents Med. Chem. 2013, 13, 13–29. [Google Scholar] [CrossRef] [PubMed]
- Li, X.Z.; Zhang, S.N. Recent advance in treatment of osteoarthritis by bioactive components from herbal medicine. Chin. Med. 2020, 15, 80. [Google Scholar] [CrossRef]
- Wong, R.H.X.; Evans, H.M.; Howe, P.R.C. Resveratrol supplementation reduces pain experience by postmenopausal women. Menopause 2017, 24, 916–922. [Google Scholar] [CrossRef]
- Lopresti, A.L.; Smith, S.J.; Jackson-Michel, S.; Fairchild, T. An investigation into the effects of a curcumin extract (Curcugen®) on osteoarthritis pain of the knee: A randomised, double-blind, placebo-controlled study. Nutrients 2021, 14, 41. [Google Scholar] [CrossRef]
- Shakibaei, M.; Mobasheri, A.; Buhrmann, C. Curcumin synergizes with resveratrol to stimulate the MAPK signaling pathway in human articular chondrocytes in vitro. Genes Nutr. 2011, 6, 171–179. [Google Scholar] [CrossRef] [Green Version]
- Pak, P.J.; Lee, D.G.; Sung, J.H.; Jung, S.H.; Han, T.Y.; Park, S.H.; Chung, N. Synergistic effect of the herbal mixture C5E on gemcitabine treatment in PANC-1 cells. Mol. Med. Rep. 2021, 23, 315. [Google Scholar] [CrossRef]
- Yousaf, M.; Razmovski-Naumovski, V.; Zubair, M.; Chang, D.; Zhou, X. Synergistic effects of natural product combinations in protecting the endothelium against cardiovascular risk factors. J. Evid. Based Integr. Med. 2022, 27, 2515690x221113327. [Google Scholar] [CrossRef]
- Paun, G.; Neagu, E.; Albu, C.; Savin, S.; Radu, G.L. In vitro evaluation of antidiabetic and anti-inflammatory activities of polyphenolic-rich extracts from Anchusa officinalis and Melilotus officinalis. ACS Omega 2020, 5, 13014–13022. [Google Scholar] [CrossRef] [PubMed]
- Patle, D.; Vyas, M.; Khatik, G.L. A review on natural products and herbs used in the management of diabetes. Curr. Diabetes Rev. 2021, 17, 186–197. [Google Scholar] [CrossRef] [PubMed]
- Cheng, L.; Wang, F.; Cao, Y.; Cai, G.; Wei, Q.; Shi, S.; Guo, Y. Screening of potent alpha-glucosidase inhibitory and antioxidant polyphenols in Prunella vulgaris L. by bioreaction-HPLC-quadrupole-time-of-flight-MS/MS and in silico analysis. J. Sep. Sci. 2022, 45, 3393–3403. [Google Scholar] [CrossRef] [PubMed]
- Huang, N.; Hauck, C.; Yum, M.Y.; Rizshsky, L.; Widrlechner, M.P.; McCoy, J.A.; Murphy, P.A.; Dixon, P.M.; Nikolau, B.J.; Birt, D.F. Rosmarinic acid in Prunella vulgaris ethanol extract inhibits lipopolysaccharide-induced prostaglandin E2 and nitric oxide in RAW 264.7 mouse macrophages. J. Agric. Food Chem. 2009, 57, 10579–10589. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joksic, G.; Radak, D.; Sudar-Milovanovic, E.; Obradovic, M.; Radovanovic, J.; Isenovic, E.R. Effects of Gentiana lutea root on vascular diseases. Curr. Vasc. Pharmacol. 2021, 19, 359–369. [Google Scholar] [CrossRef] [PubMed]
- Kesavan, R.; Chandel, S.; Upadhyay, S.; Bendre, R.; Ganugula, R.; Potunuru, U.R.; Giri, H.; Sahu, G.; Kumar, P.U.; Reddy, G.B.; et al. Gentiana lutea exerts anti-atherosclerotic effects by preventing endothelial inflammation and smooth muscle cell migration. Nutr. Metab. Cardiovasc. Dis. 2016, 26, 293–301. [Google Scholar] [CrossRef]
- Pan, Y.; Zhao, Y.L.; Zhang, J.; Li, W.Y.; Wang, Y.Z. Phytochemistry and pharmacological activities of the genus Gentiana (Gentianaceae). Chem. Biodivers. 2016, 13, 107–150. [Google Scholar] [CrossRef]
- Park, E.; Lee, C.G.; Kim, J.; Yeo, S.; Kim, J.A.; Choi, C.W.; Jeong, S.Y. Antiobesity effects of Gentiana lutea extract on 3T3-L1 preadipocytes and a high-fat diet-induced mouse model. Molecules 2020, 25, 2453. [Google Scholar] [CrossRef]
- Zhao, L.; Ye, J.; Wu, G.T.; Peng, X.J.; Xia, P.F.; Ren, Y. Gentiopicroside prevents interleukin-1β induced inflammation response in rat articular chondrocyte. J. Ethnopharmacol. 2015, 172, 100–107. [Google Scholar] [CrossRef]
- Park, E.; Lee, C.G.; Yun, S.H.; Hwang, S.; Jeon, H.; Kim, J.; Yeo, S.; Jeong, H.; Yun, S.H.; Jeong, S.Y. Ameliorative effects of loganin on arthritis in chondrocytes and destabilization of the medial meniscus-induced animal model. Pharmaceuticals 2021, 14, 135. [Google Scholar] [CrossRef]
- Glasson, S.S.; Chambers, M.G.; Van Den Berg, W.B.; Little, C.B. The OARSI histopathology initiative-recommendations for histological assessments of osteoarthritis in the mouse. Osteoarthr. Cartil. 2010, 18 (Suppl. 3), S17–S23. [Google Scholar] [CrossRef] [Green Version]
- Xu, F.; Zhao, L.J.; Liao, T.; Li, Z.C.; Wang, L.L.; Lin, P.Y.; Jiang, R.; Wei, Q.J. Ononin ameliorates inflammation and cartilage degradation in rat chondrocytes with IL-1β-induced osteoarthritis by downregulating the MAPK and NF-κB pathways. BMC Complement. Med. Ther. 2022, 22, 25. [Google Scholar] [CrossRef]
- Nakayama, M.; Niki, Y.; Kawasaki, T.; Takeda, Y.; Horiuchi, K.; Sasaki, A.; Okada, Y.; Umezawa, K.; Ikegami, H.; Toyama, Y.; et al. Enhanced susceptibility to lipopolysaccharide-induced arthritis and endotoxin shock in interleukin-32α transgenic mice through induction of tumor necrosis factor alpha. Arthritis Res. Ther. 2012, 14, R120. [Google Scholar] [CrossRef] [Green Version]
- Hwang, Y.J.; Lee, E.J.; Kim, H.R.; Hwang, K.A. NF-κB-targeted anti-inflammatory activity of Prunella vulgaris var. lilacina in macrophages RAW 264.7. Int. J. Mol. Sci. 2013, 14, 21489–21503. [Google Scholar] [CrossRef] [Green Version]
- Ju, Z.; Li, M.; Xu, J.; Howell, D.C.; Li, Z.; Chen, F.E. Recent development on COX-2 inhibitors as promising anti-inflammatory agents: The past 10 years. Acta Pharm. Sin. B 2022, 12, 2790–2807. [Google Scholar] [CrossRef]
- Sacitharan, P.K. Ageing and osteoarthritis. Subcell. Biochem. 2019, 91, 123–159. [Google Scholar] [CrossRef]
- Kolasinski, S.L.; Neogi, T.; Hochberg, M.C.; Oatis, C.; Guyatt, G.; Block, J.; Callahan, L.; Copenhaver, C.; Dodge, C.; Felson, D.; et al. 2019 American College of Rheumatology/Arthritis Foundation guideline for the management of osteoarthritis of the hand, hip, and knee. Arthritis Rheumatol. 2020, 72, 220–233. [Google Scholar] [CrossRef]
- Bannuru, R.R.; Osani, M.C.; Vaysbrot, E.E.; Arden, N.K.; Bennell, K.; Bierma-Zeinstra, S.M.A.; Kraus, V.B.; Lohmander, L.S.; Abbott, J.H.; Bhandari, M.; et al. OARSI guidelines for the non-surgical management of knee, hip, and polyarticular osteoarthritis. Osteoarthr. Cartil. 2019, 27, 1578–1589. [Google Scholar] [CrossRef] [Green Version]
- Loeser, R.F.; Goldring, S.R.; Scanzello, C.R.; Goldring, M.B. Osteoarthritis: A disease of the joint as an organ. Arthritis Rheumatol. 2012, 64, 1697–1707. [Google Scholar] [CrossRef]
- Yunus, M.H.M.; Nordin, A.; Kamal, H. Pathophysiological perspective of osteoarthritis. Medicina 2020, 56, 614. [Google Scholar] [CrossRef]
- Wu, Y.; Wang, Z.; Lin, Z.; Fu, X.; Zhan, J.; Yu, K. Salvianolic acid A has anti-osteoarthritis effect in vitro and in vivo. Front. Pharmacol. 2020, 11, 682. [Google Scholar] [CrossRef]
- Park, J.Y.; Pillinger, M.H.; Abramson, S.B. Prostaglandin E2 synthesis and secretion: The role of PGE2 synthases. Clin. Immunol. 2006, 119, 229–240. [Google Scholar] [CrossRef]
- Dejica, V.M.; Mort, J.S.; Laverty, S.; Antoniou, J.; Zukor, D.J.; Tanzer, M.; Poole, A.R. Increased type II collagen cleavage by cathepsin K and collagenase activities with aging and osteoarthritis in human articular cartilage. Arthritis Res. Ther. 2012, 14, R113. [Google Scholar] [CrossRef] [Green Version]
- Shimpo, H.; Sakai, T.; Kondo, S.; Mishima, S.; Yoda, M.; Hiraiwa, H.; Ishiguro, N. Regulation of prostaglandin E(2) synthesis in cells derived from chondrocytes of patients with osteoarthritis. J. Orthop. Sci. 2009, 14, 611–617. [Google Scholar] [CrossRef]
- Park, S.H.; Koo, H.J.; Sung, Y.Y.; Kim, H.K. The protective effect of Prunella vulgaris ethanol extract against vascular inflammation in TNF-α-stimulated human aortic smooth muscle cells. BMB Rep. 2013, 46, 352–357. [Google Scholar] [CrossRef] [Green Version]
- Abulhasan, J.; Grey, M. Anatomy and physiology of knee stability. J. Funct. Morphol. Kinesiol. 2017, 2, 34. [Google Scholar] [CrossRef] [Green Version]
- Sophia Fox, A.J.; Bedi, A.; Rodeo, S.A. The basic science of articular cartilage: Structure, composition, and function. Sports Health 2009, 1, 461–468. [Google Scholar] [CrossRef] [Green Version]
- Makris, E.A.; Hadidi, P.; Athanasiou, K.A. The knee meniscus: Structure-function, pathophysiology, current repair techniques, and prospects for regeneration. Biomaterials 2011, 32, 7411–7431. [Google Scholar] [CrossRef] [Green Version]
- Fang, H.; Beier, F. Mouse models of osteoarthritis: Modelling risk factors and assessing outcomes. Nat. Rev. Rheumatol. 2014, 10, 413–421. [Google Scholar] [CrossRef]
- Hardcastle, S.A.; Dieppe, P.; Gregson, C.L.; Davey Smith, G.; Tobias, J.H. Osteoarthritis and bone mineral density: Are strong bones bad for joints? Bonekey Rep. 2015, 4, 624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aho, O.-M.; Finnilä, M.; Thevenot, J.; Saarakkala, S.; Lehenkari, P. Subchondral bone histology and grading in osteoarthritis. PLoS ONE 2017, 12, e0173726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCulloch, K.; Huesa, C.; Dunning, L.; Litherland, G.J.; Van ‘T Hof, R.J.; Lockhart, J.C.; Goodyear, C.S. Accelerated post traumatic osteoarthritis in a dual injury murine model. Osteoarthr. Cartil. 2019, 27, 1800–1810. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munjal, A.; Bapat, S.; Hubbard, D.; Hunter, M.; Kolhe, R.; Fulzele, S. Advances in molecular biomarker for early diagnosis of osteoarthritis. Biomol. Concepts 2019, 10, 111–119. [Google Scholar] [CrossRef]
- Kapoor, M.; Martel-Pelletier, J.; Lajeunesse, D.; Pelletier, J.-P.; Fahmi, H. Role of proinflammatory cytokines in the pathophysiology of osteoarthritis. Nat. Rev. Rheumatol. 2011, 7, 33–42. [Google Scholar] [CrossRef]
- McNulty, M.A.; Loeser, R.F.; Davey, C.; Callahan, M.F.; Ferguson, C.M.; Carlson, C.S. A comprehensive histological assessment of osteoarthritis lesions in mice. Cartilage 2011, 2, 354–363. [Google Scholar] [CrossRef] [Green Version]
- Jamali, N.; Adib-Hajbaghery, M.; Soleimani, A. The effect of curcumin ointment on knee pain in older adults with osteoarthritis: A randomized placebo trial. BMC Complement. Med. Ther. 2020, 20, 305. [Google Scholar] [CrossRef]
- Thomas, J.V.; Smina, T.P.; Khanna, A.; Kunnumakkara, A.B.; Maliakel, B.; Mohanan, R.; Krishnakumar, I.M. Influence of a low-dose supplementation of curcumagalactomannoside complex (CurQfen) in knee osteoarthritis: A randomized, open-labeled, active-controlled clinical trial. Phytother. Res. 2021, 35, 1443–1455. [Google Scholar] [CrossRef]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, J.; Lee, C.-G.; Hwang, S.; Yun, S.-H.; Uprety, L.P.; Oh, K.-I.; Singh, S.; Yoo, J.; Jeong, H.; Yong, Y.; et al. Anti-Osteoarthritic Effects of Prunella Vulgaris and Gentiana Lutea In Vitro and In Vivo. Antioxidants 2023, 12, 47. https://doi.org/10.3390/antiox12010047
Kim J, Lee C-G, Hwang S, Yun S-H, Uprety LP, Oh K-I, Singh S, Yoo J, Jeong H, Yong Y, et al. Anti-Osteoarthritic Effects of Prunella Vulgaris and Gentiana Lutea In Vitro and In Vivo. Antioxidants. 2023; 12(1):47. https://doi.org/10.3390/antiox12010047
Chicago/Turabian StyleKim, Jeonghyun, Chang-Gun Lee, Seokjin Hwang, Seung-Hee Yun, Laxmi Prasad Uprety, Kang-Il Oh, Shivani Singh, Jisu Yoo, Hyesoo Jeong, Yoonjoong Yong, and et al. 2023. "Anti-Osteoarthritic Effects of Prunella Vulgaris and Gentiana Lutea In Vitro and In Vivo" Antioxidants 12, no. 1: 47. https://doi.org/10.3390/antiox12010047