
Comparison of the Effects between Tannins Extracted from Different Natural Plants on Growth Performance, Antioxidant Capacity, Immunity, and Intestinal Flora of Broiler Chickens

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Treatment
2.2. Growth Performance
2.3. Sample Collection
2.4. Measurement of Antioxidant Activity, Immune Factors and Liver Function
2.5. Measurement of Intestinal Tight Junction Protein
2.6. Histological Examination
2.7. Gut Microbiome Analysis
2.8. Statistical Analysis
3. Results
3.1. Growth Performance
3.2. Diarrhea Rate and Cecum Length
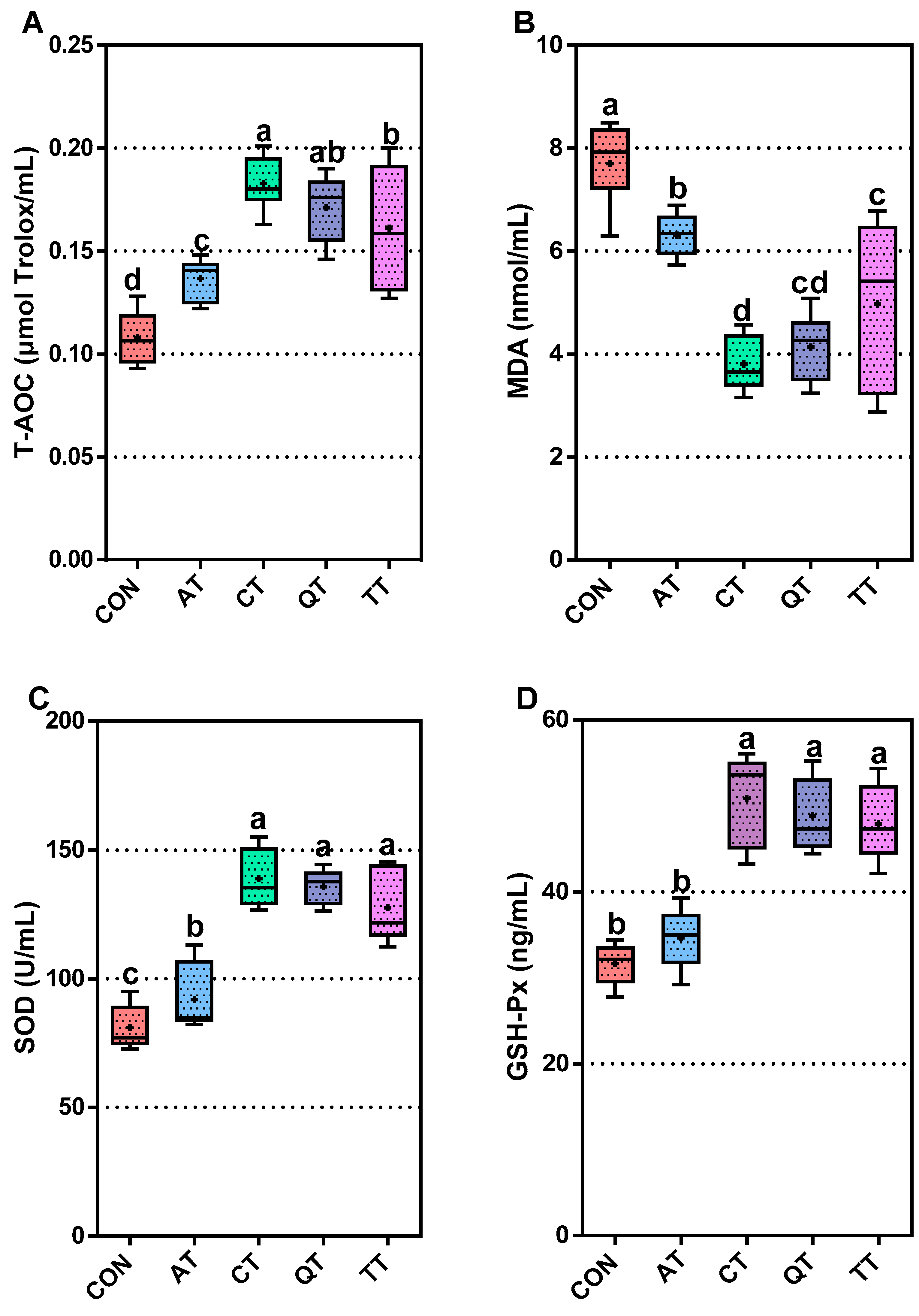
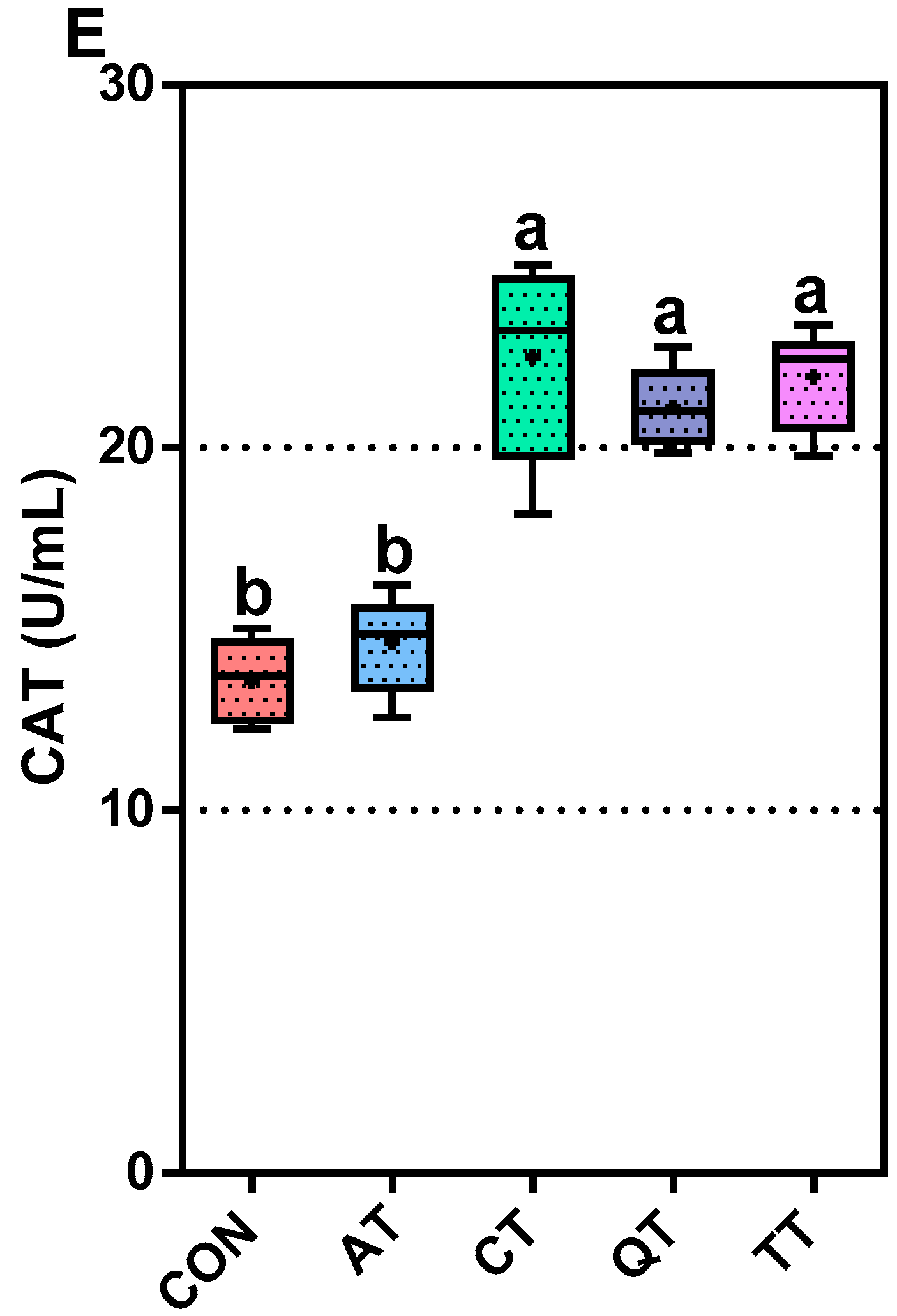
3.3. Comparative Assessment of Antioxidant Capacity
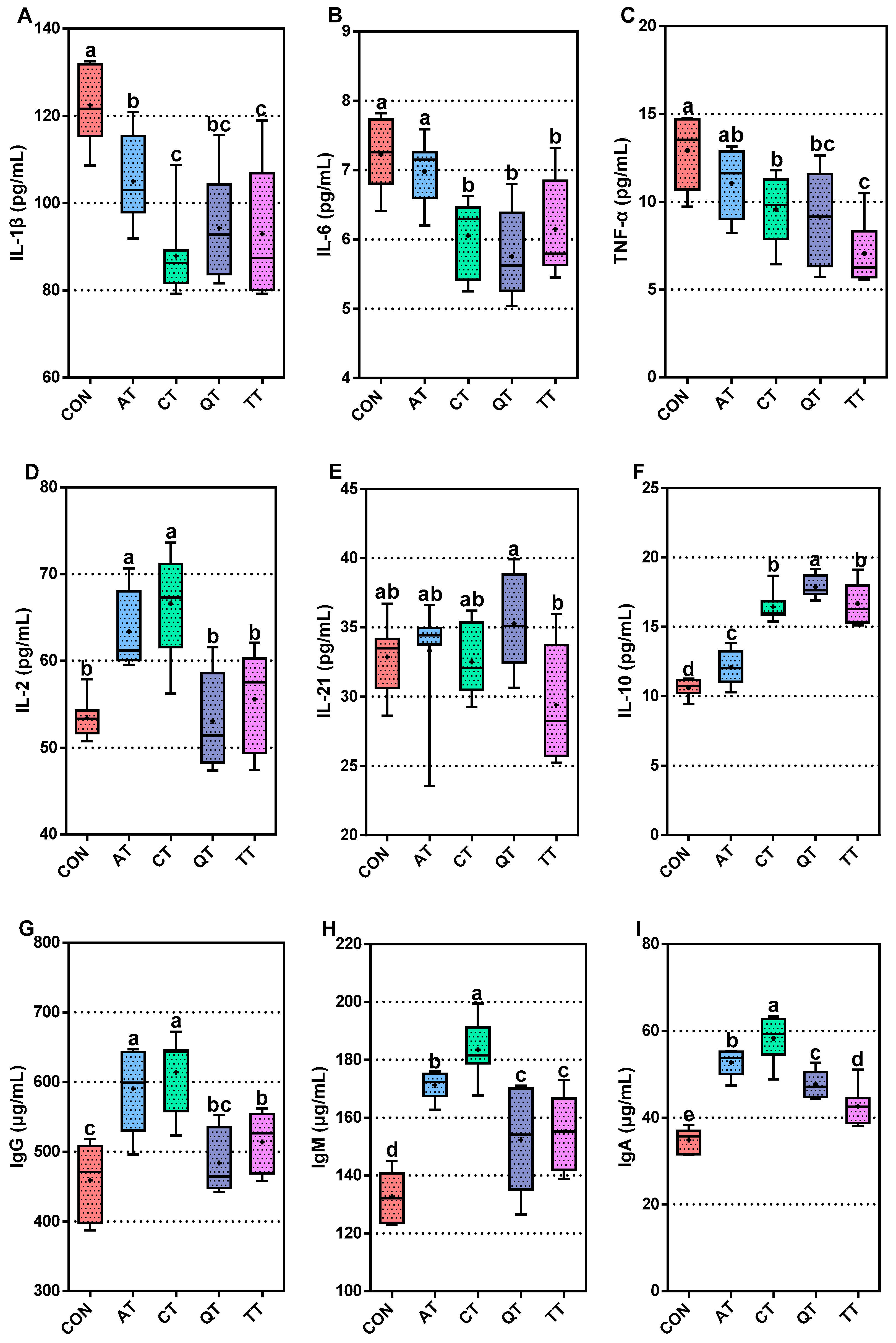
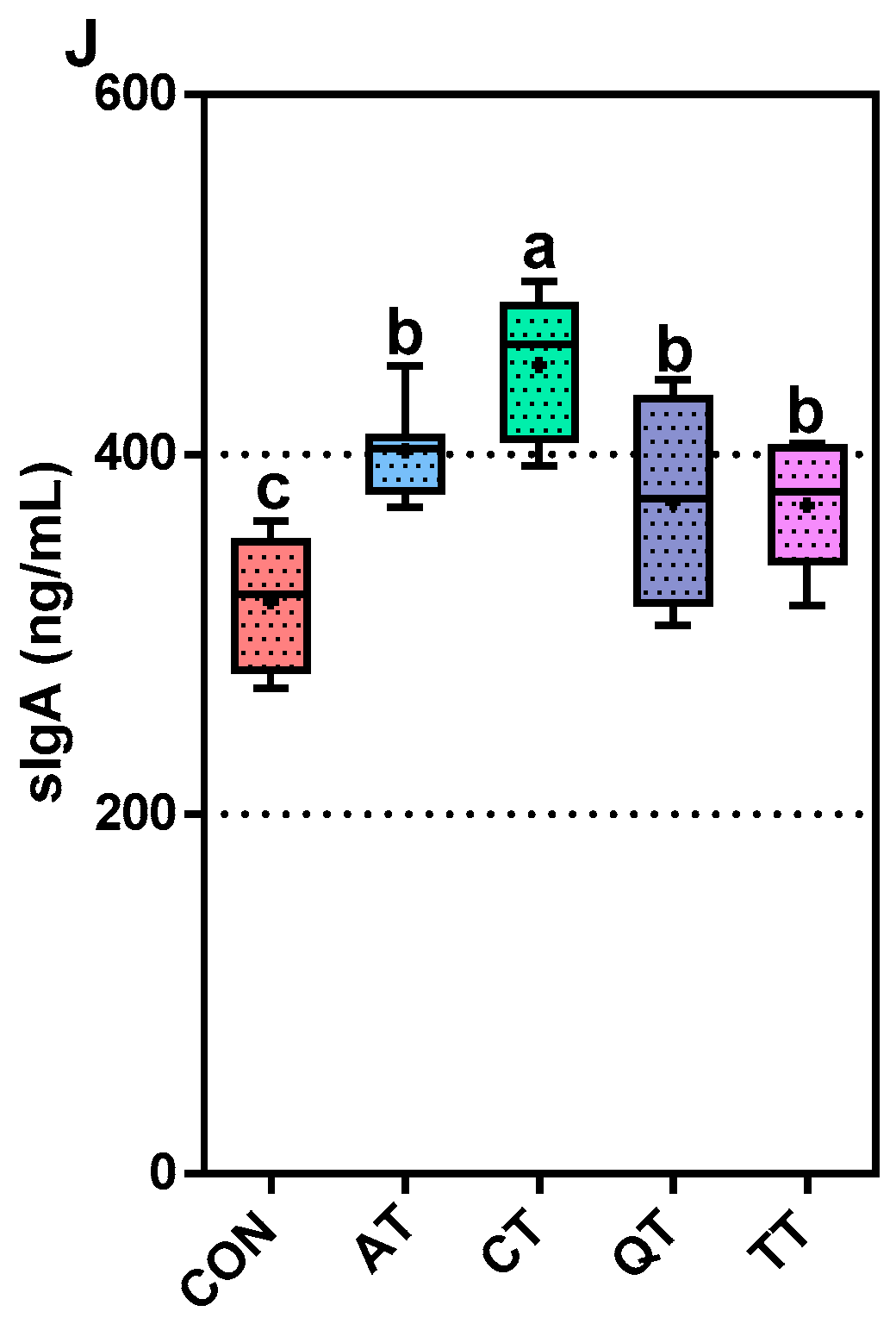
3.4. Comparative Assessment of Immunity
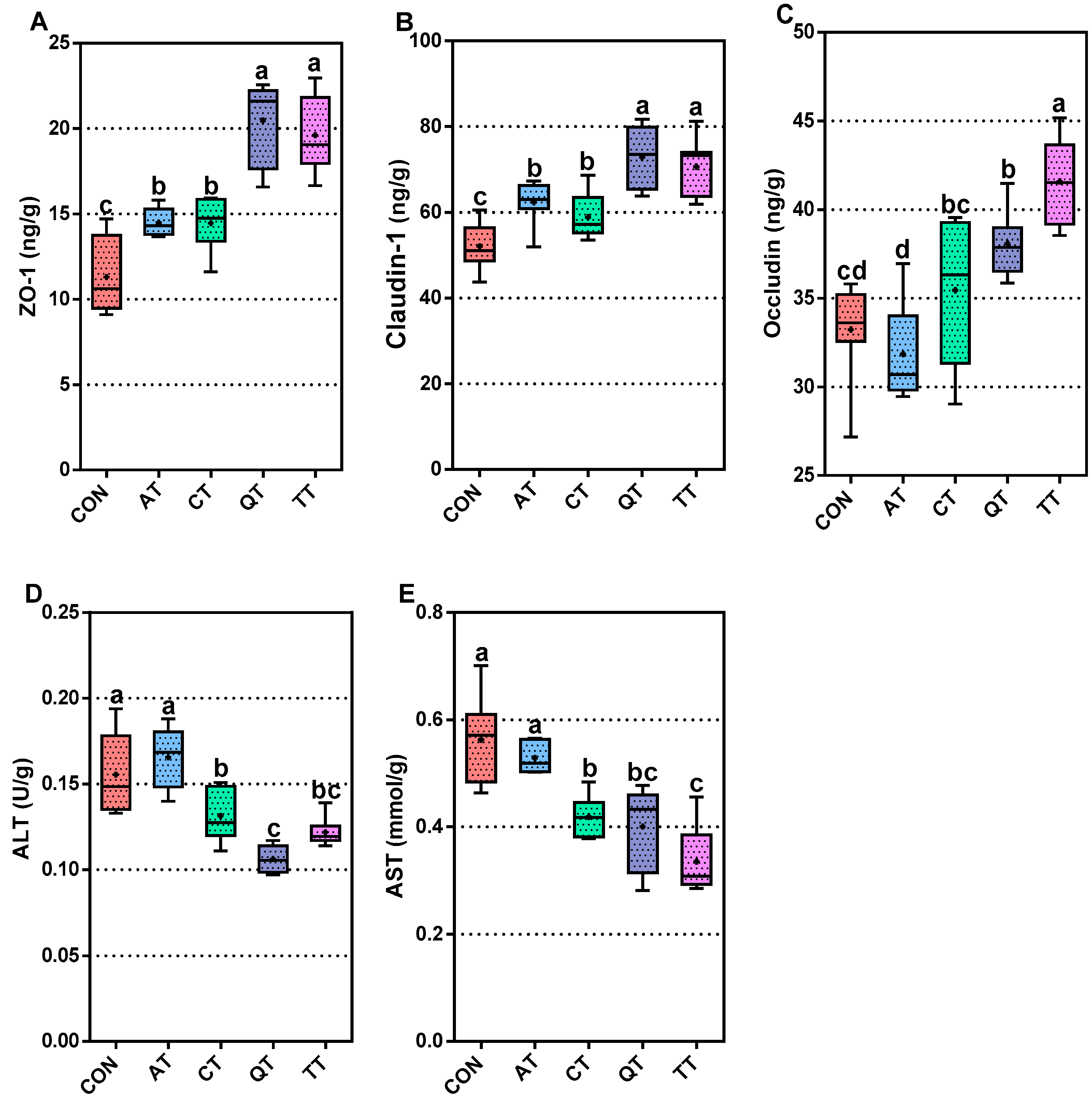
3.5. Intestinal Mucosal Barrier and Liver Function Evaluation
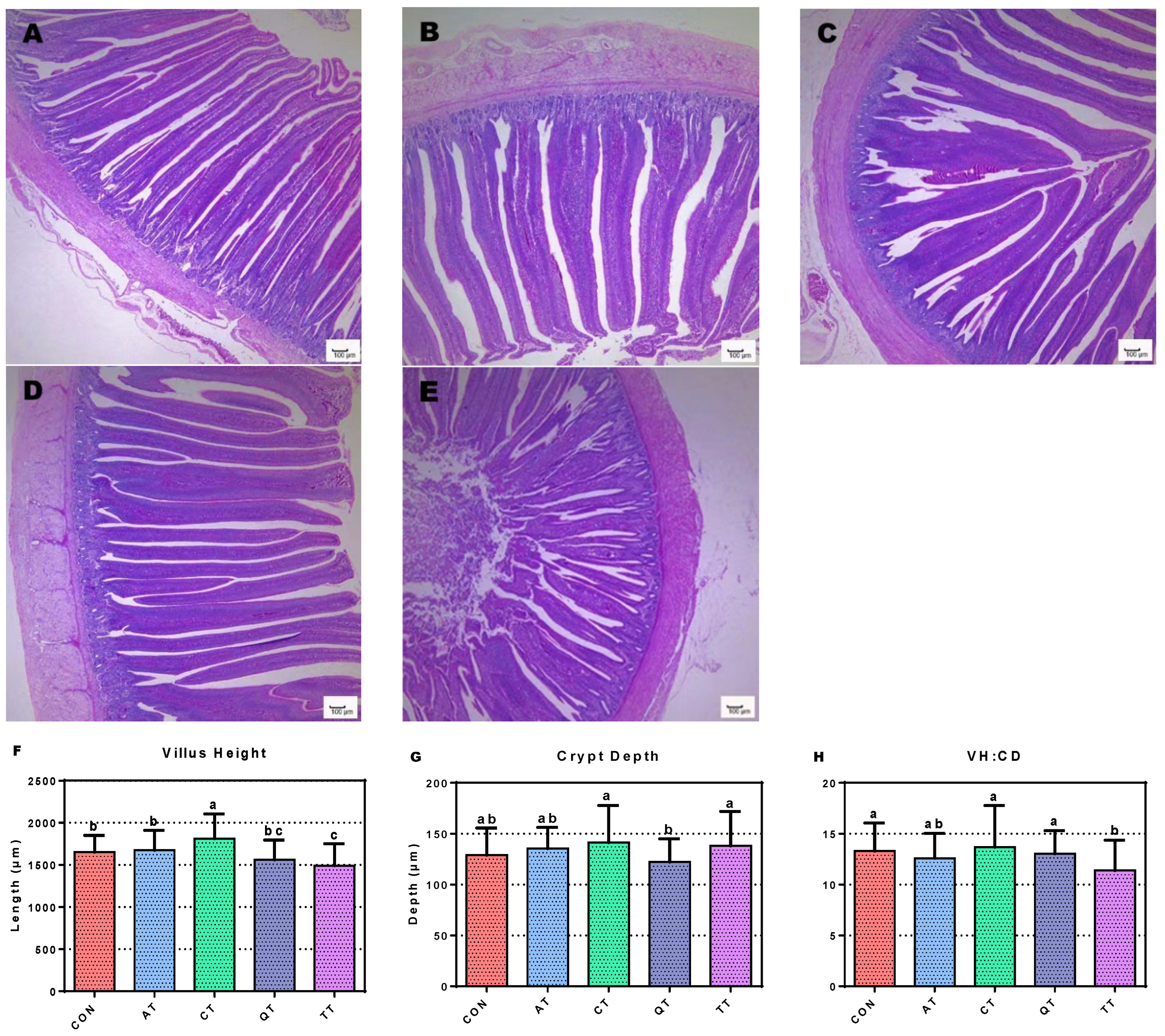
3.6. Assessment of Small Intestine Morphology
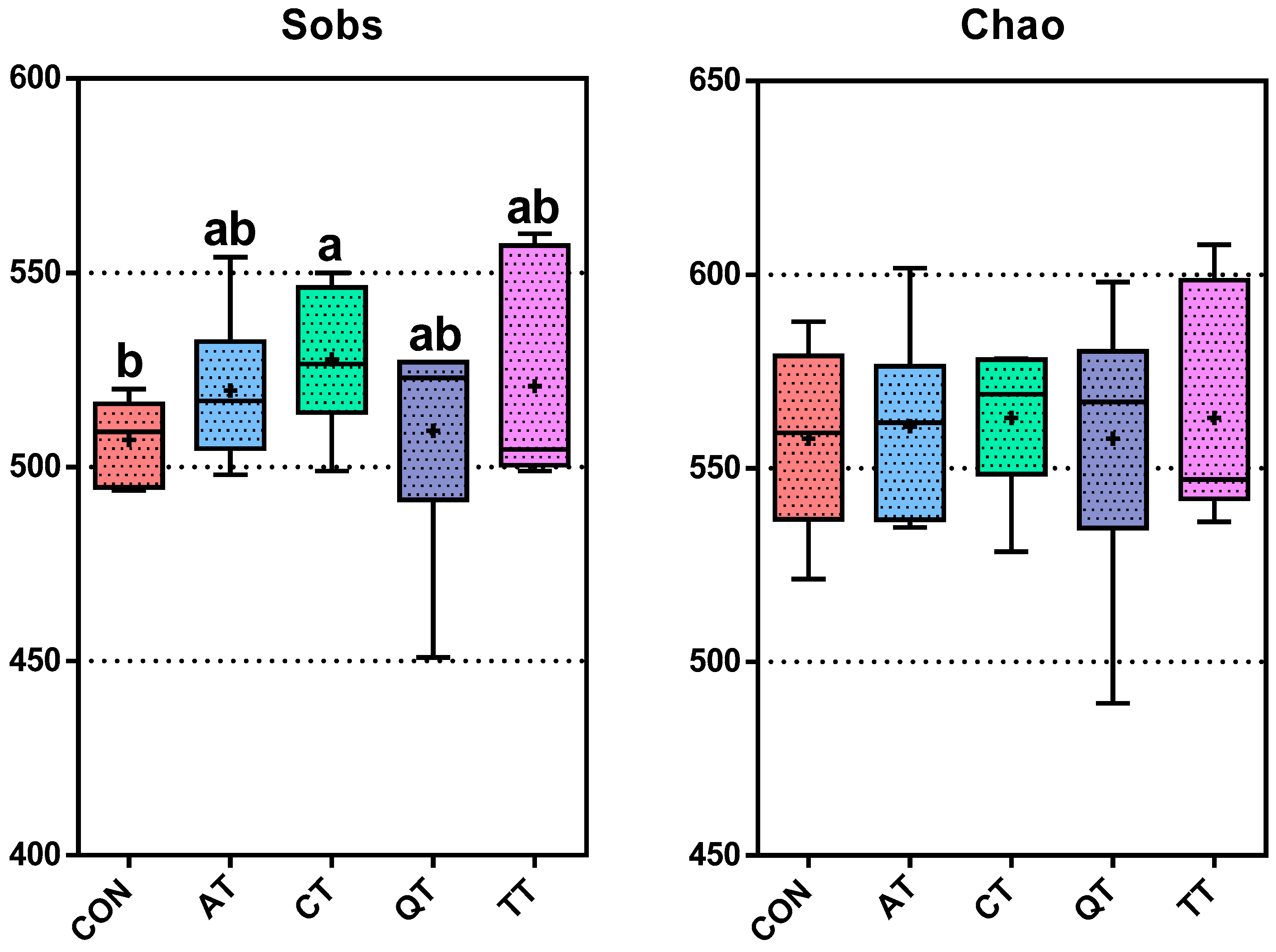
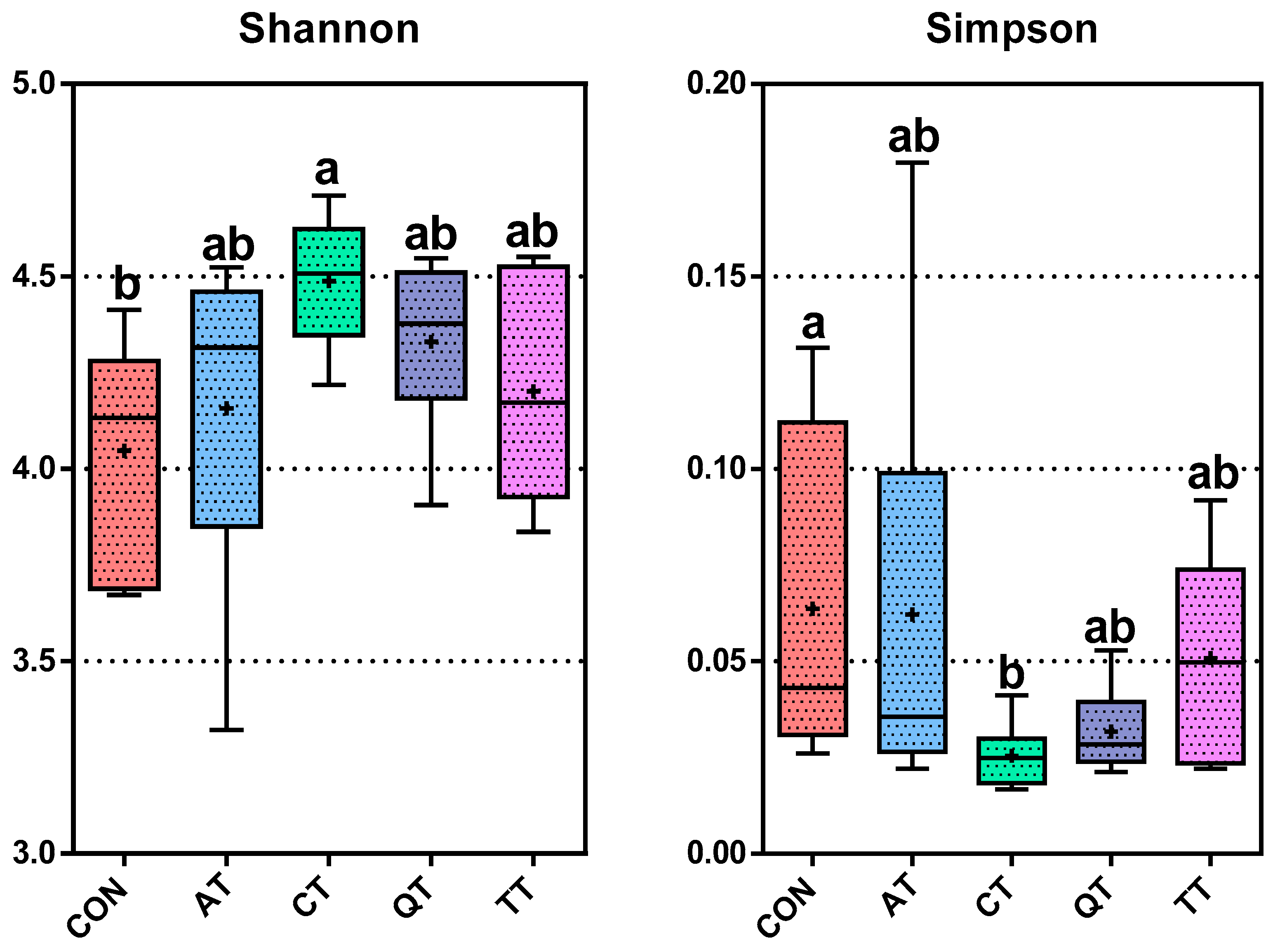
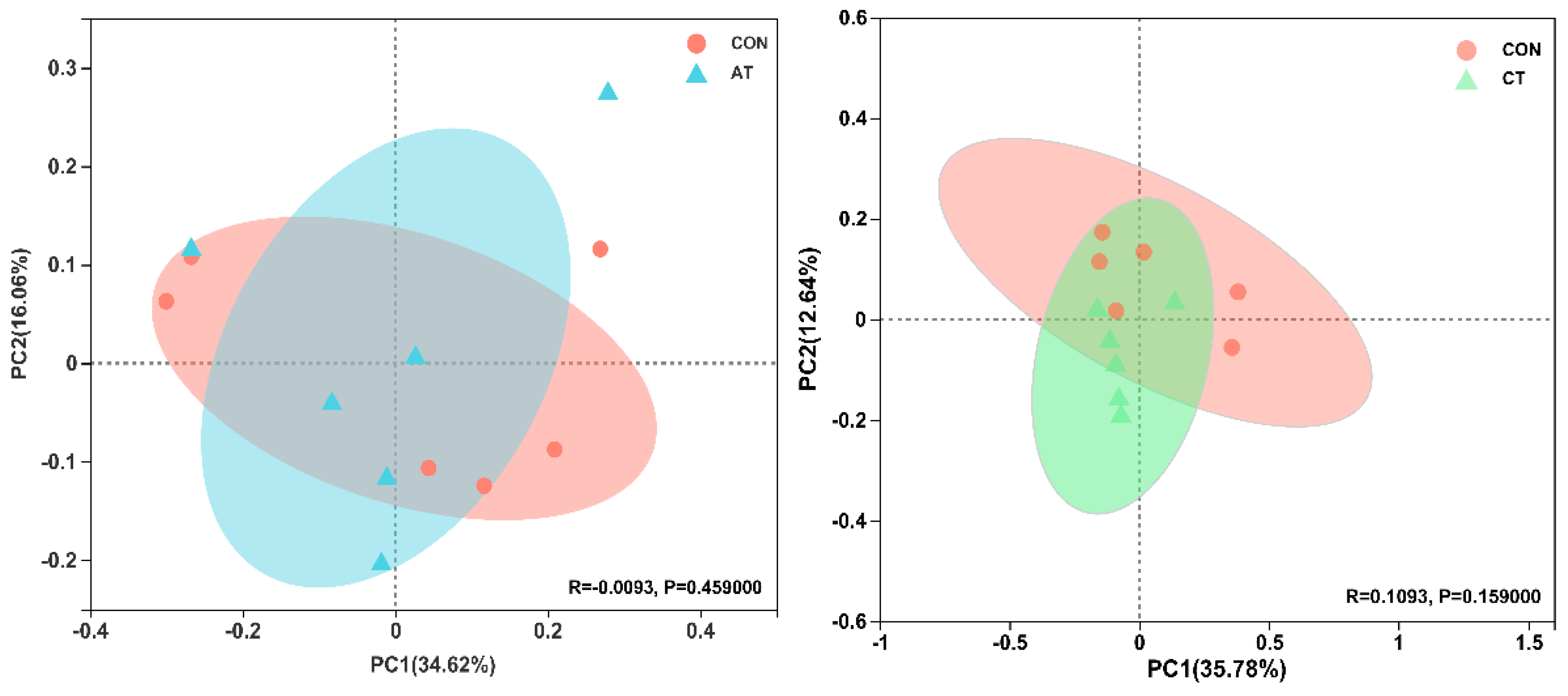
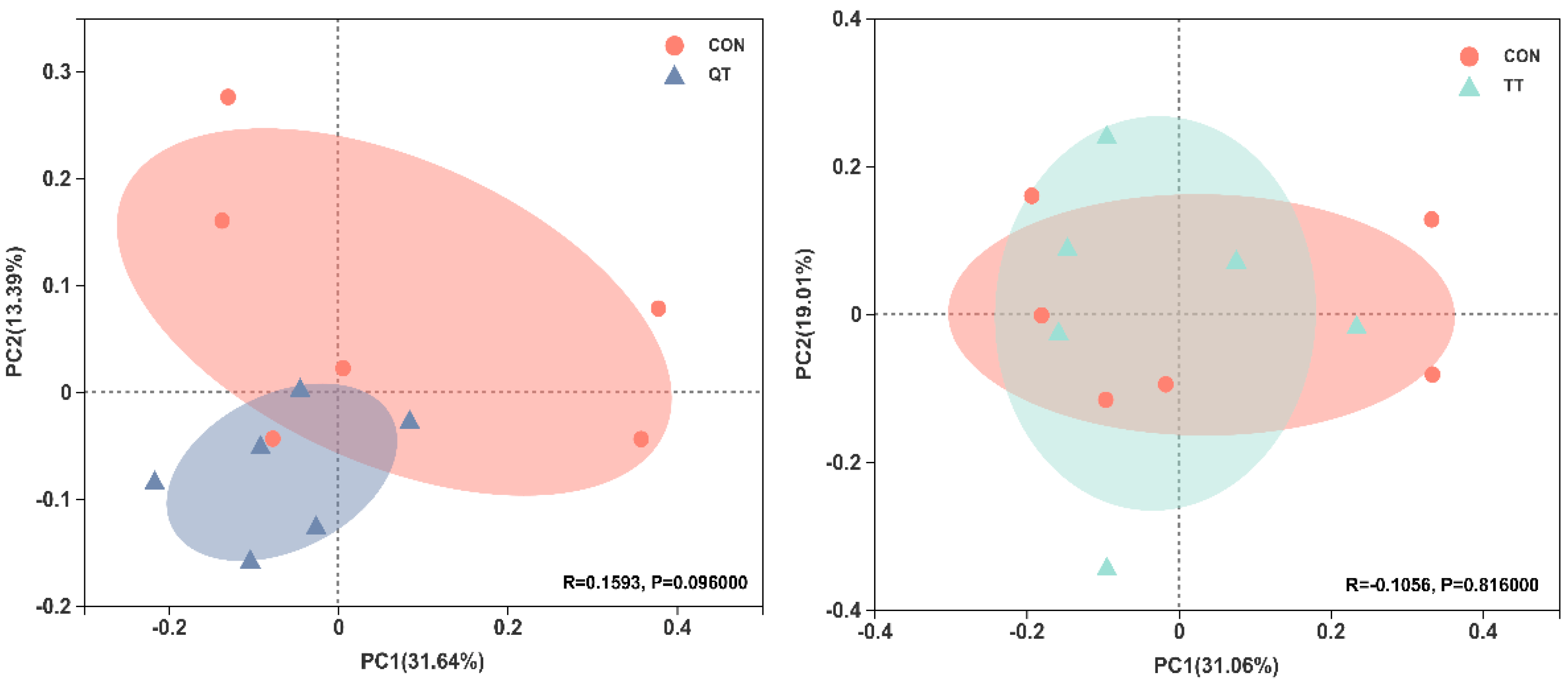
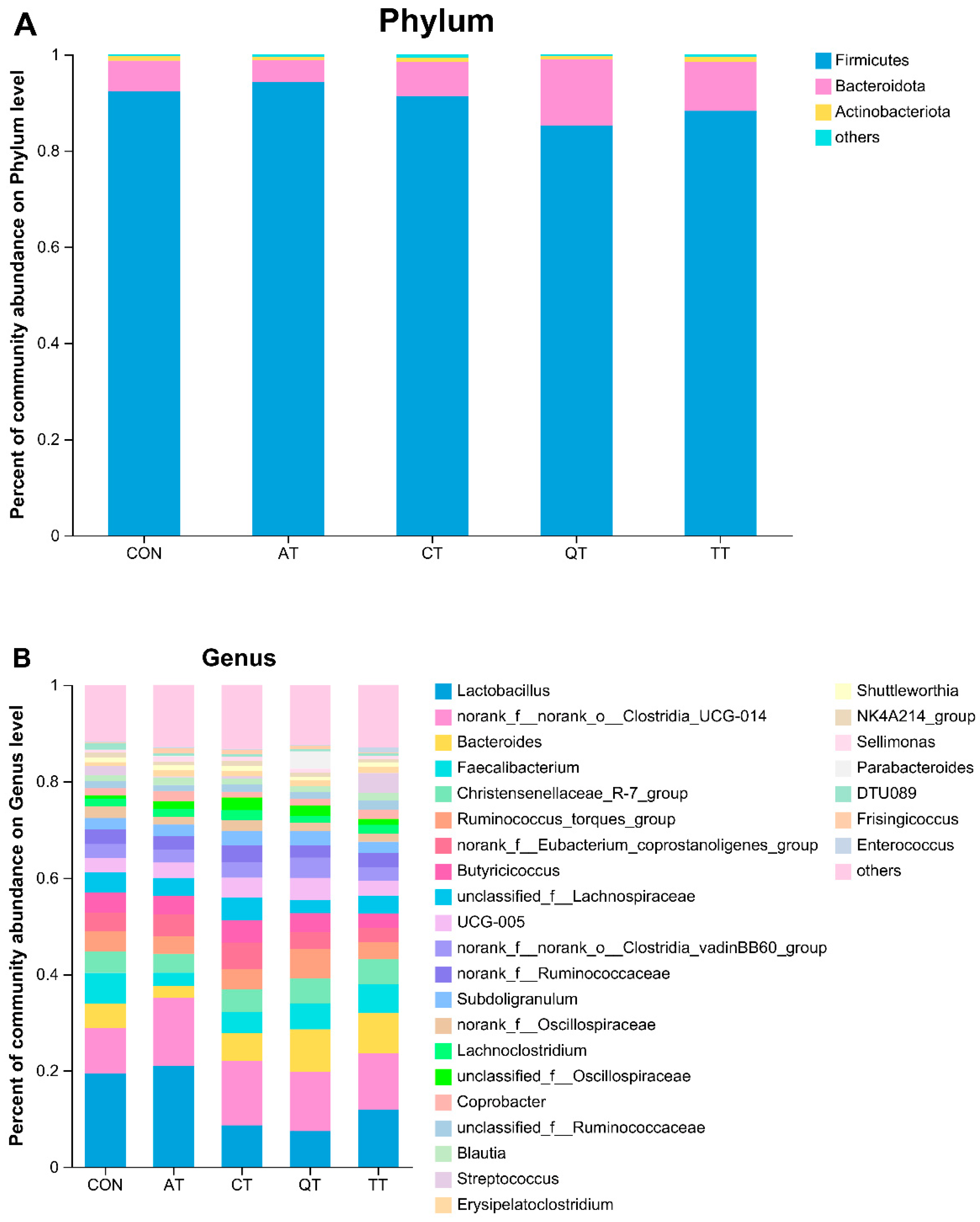
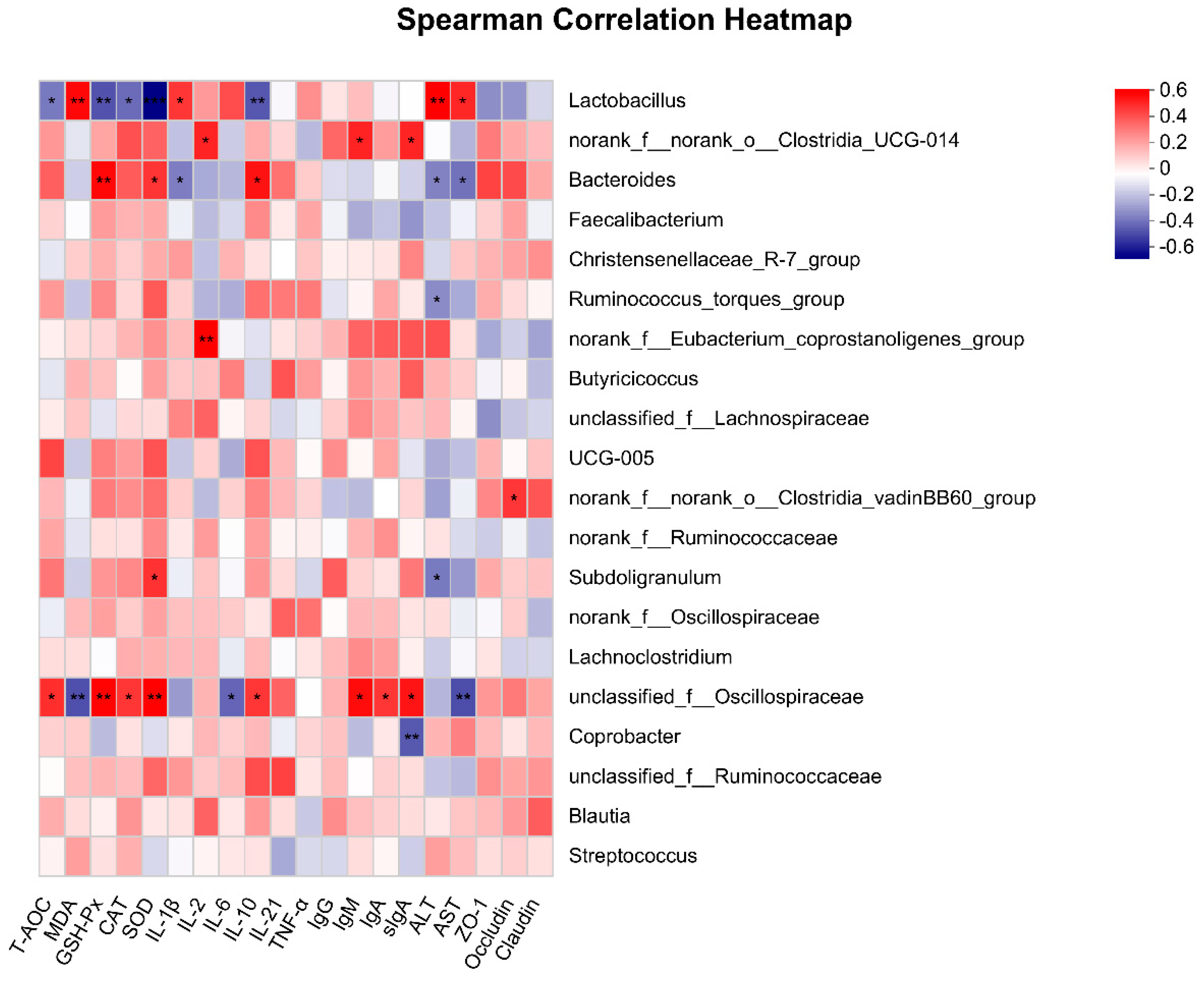
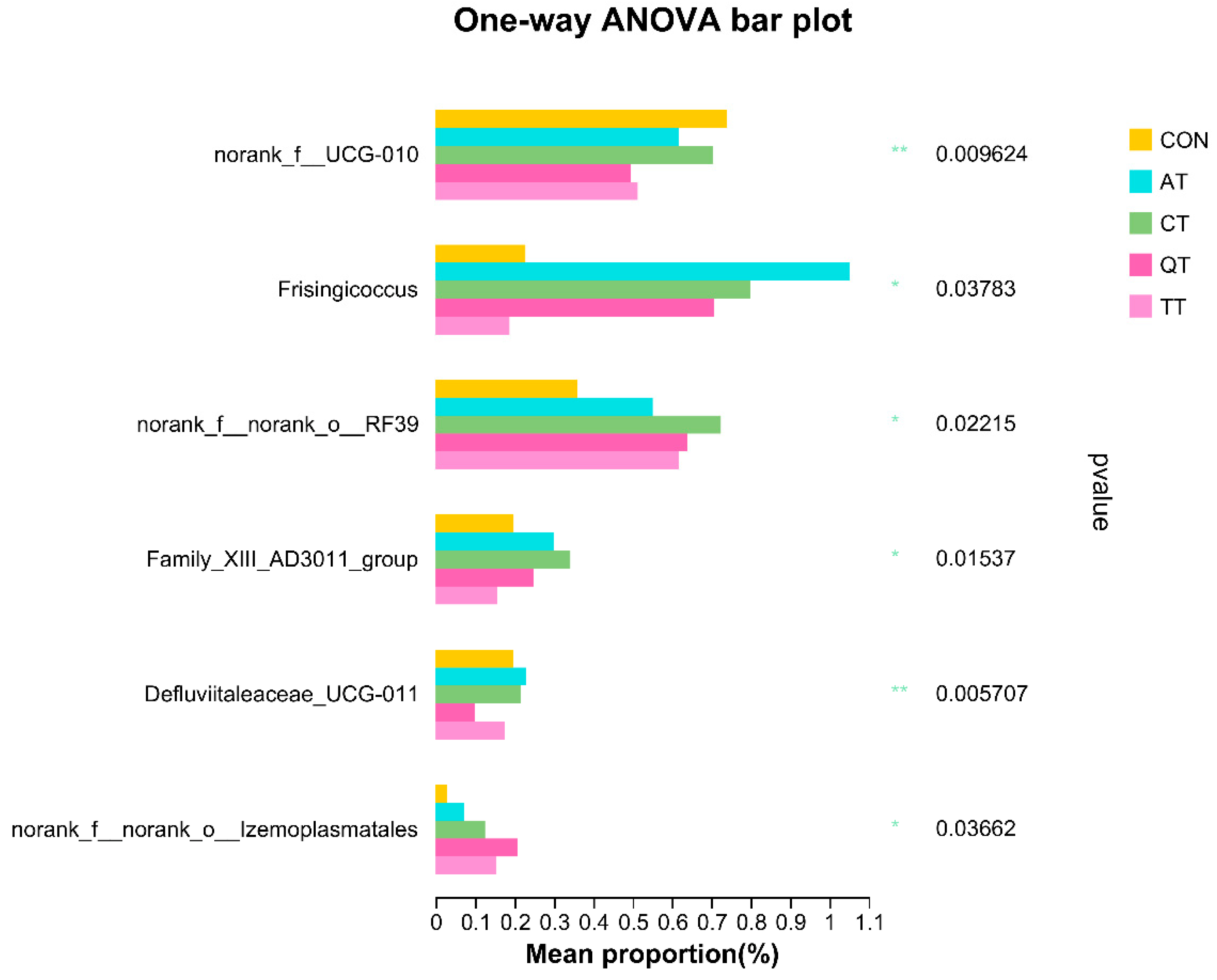
3.7. Effects of Different Tannins on Intestinal Flora in Healthy Subjects
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Abd El-Hack, M.E.; El-Saadony, M.T.; Elbestawy, A.R.; Gado, A.R.; Nader, M.M.; Saad, A.M.; El-Tahan, A.M.; Taha, A.E.; Salem, H.M.; El-Tarabily, K.A. Hot red pepper powder as a safe alternative to antibiotics in organic poultry feed: An updated review. Poult. Sci. 2022, 101, 101684. [Google Scholar] [CrossRef] [PubMed]
- Silveira, R.F.; Roque-Borda, C.A.; Vicente, E.F. Antimicrobial peptides as a feed additive alternative to animal production, food safety and public health implications: An overview. Anim. Nutr. 2021, 7, 896–904. [Google Scholar] [CrossRef] [PubMed]
- Low, C.X.; Tan, L.T.H.; Ab Mutalib, N.S.; Pusparajah, P.; Goh, B.H.; Chan, K.G.; Letchumanan, V.; Lee, L.H. Unveiling the impact of antibiotics and alternative methods for animal husbandry: A review. Antibiotics 2021, 10, 578. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Q.; Sun, P.; Zhang, B.; Kong, L.; Xiao, C.; Song, Z. Progress on gut health maintenance and antibiotic alternatives in broiler chicken production. Front. Nutr. 2021, 8, 692839. [Google Scholar] [CrossRef] [PubMed]
- Righi, F.; Pitino, R.; Manuelian, C.; Simoni, M.; Quarantelli, A.; De Marchi, M.; Tsiplakou, E. Plant Feed Additives as Natural Alternatives to the Use of Synthetic Antioxidant Vitamins on Poultry Performances, Health, and Oxidative Status: A Review of the Literature in the Last 20 Years. Antioxidants 2021, 10, 659. [Google Scholar] [CrossRef] [PubMed]
- Ali, A.; Ponnampalam, E.N.; Pushpakumara, G.; Cottrell, J.J.; Suleria, H.A.; Dunshea, F.R. Cinnamon: A natural feed additive for poultry health and production—A review. Animals 2021, 11, 2026. [Google Scholar] [CrossRef]
- Mahfuz, S.; Shang, Q.; Piao, X. Phenolic compounds as natural feed additives in poultry and swine diets: A review. J. Anim. Sci. Biotechnol. 2021, 12, 48. [Google Scholar] [CrossRef]
- Grochowski, D.M.; Skalicka-Wozniak, K.; Orhan, I.E.; Xiao, J.; Locatelli, M.; Piwowarski, J.P.; Granica, S.; Tomczyk, M. A comprehensive review of agrimoniin. Ann. N. Y. Acad. Sci. 2017, 1401, 166–180. [Google Scholar] [CrossRef]
- Min, B.R.; Solaiman, S.; Waldrip, H.M.; Parker, D.; Todd, R.W.; Brauer, D. Dietary mitigation of enteric methane emissions from ruminants: A review of plant tannin mitigation options. Anim. Nutr. 2020, 6, 231–246. [Google Scholar] [CrossRef]
- Huang, Q.; Liu, X.; Zhao, G.; Hu, T.; Wang, Y. Potential and challenges of tannins as an alternative to in-feed antibiotics for farm animal production. Anim. Nutr. 2018, 4, 137–150. [Google Scholar] [CrossRef]
- Shirmohammadli, Y.; Efhamisisi, D.; Pizzi, A. Tannins as a sustainable raw material for green chemistry: A review. Ind. Crop. Prod. 2018, 126, 316–332. [Google Scholar] [CrossRef]
- Aboagye, I.A.; Beauchemin, K.A. Potential of molecular weight and structure of tannins to reduce methane emissions from ruminants: A review. Animals 2019, 9, 856. [Google Scholar] [CrossRef]
- Fraga-Corral, M.; Otero, P.; Echave, J.; Garcia-Oliveira, P.; Carpena, M.; Jarboui, A.; Nunez-Estevez, B.; Simal-Gandara, J.; Prieto, M.A. By-products of agri-food industry as tannin-rich sources: A review of tannins’ biological activities and their potential for valorization. Foods 2021, 10, 137. [Google Scholar] [CrossRef]
- Tang, Q.; Xu, E. Research progress on biological functions of tannins and their application in livestock and poultry breeding. China Anim. Husb. 2021, 57, 33–39. [Google Scholar]
- Chen, L.; Bao, X.; Guo, G.; Huo, W.; Xu, Q.; Wang, C.; Li, Q.; Liu, Q. Effects of hydrolysable tannin with or without condensed tannin on alfalfa silage fermentation characteristics and in vitro ruminal methane production, fermentation patterns, and microbiota. Animals 2021, 11, 1967. [Google Scholar] [CrossRef]
- Girard, M.; Hu, D.; Pradervand, N.; Neuenschwander, S.; Bee, G. Chestnut extract but not sodium salicylate decreases the severity of diarrhea and enterotoxigenic Escherichia coli F4 shedding in artificially infected piglets. PLoS ONE 2020, 15, e0214267. [Google Scholar] [CrossRef]
- Sun, J. Effects of Tannins on the Intestinal Barrier and Microbial Community in the Cecum of Eimeria Tenella-Infected Chicks. Bachelor’s Thesis, Guangxi University, Nanning, China, 2020. [Google Scholar]
- Liu, H.S.; Mahfuz, S.U.; Wu, D.; Shang, Q.H.; Piao, X.S. Effect of chestnut wood extract on performance, meat quality, antioxidant status, immune function, and cholesterol metabolism in broilers. Poult. Sci. 2020, 99, 4488–4495. [Google Scholar] [CrossRef]
- Wang, K.; Yan, Q.; Ren, A.; Zheng, M.; Zhang, P.; Tan, Z.; Zhou, C. Novel linkages between bacterial composition of hindgut and host metabolic responses to sara induced by high-paddy diet in young goats. Front. Vet. Sci. 2022, 8, 791482. [Google Scholar] [CrossRef]
- He, B.; Zhu, R.; Yang, H.; Lu, Q.; Wang, W.; Song, L.; Sun, X.; Zhang, G.; Li, S.; Yang, J.; et al. Assessing the impact of data preprocessing on analyzing next generation sequencing data. Front. Bioeng. Biotechnol. 2020, 8, 817. [Google Scholar] [CrossRef]
- Wang, K.; Peng, X.; Yang, A.; Huang, Y.; Tan, Y.; Qian, Y.; Lv, F.; Si, H. Effects of diets with different protein levels on lipid metabolism and gut microbes in the host of different genders. Front. Nutr. 2022, 9, 940217. [Google Scholar] [CrossRef]
- Valenti, B.; Natalello, A.; Vasta, V.; Campidonico, L.; Roscini, V.; Mattioli, S.; Pauselli, M.; Priolo, A.; Lanza, M.; Luciano, G. Effect of different dietary tannin extracts on lamb growth performances and meat oxidative stability: Comparison between mimosa, chestnut and tara. Animal 2019, 13, 435–443. [Google Scholar] [CrossRef] [PubMed]
- Yin, L.; Li, J.; Wang, H.; Yi, Z.; Wang, L.; Zhang, S.; Li, X.; Wang, Q.; Li, J.; Yang, H.; et al. Effects of vitamin B6 on the growth performance, intestinal morphology, and gene expression in weaned piglets that are fed a low-protein diet. J. Anim. Sci. 2020, 98, skaa022. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Ren, A.; Zheng, M.; Jiao, J.; Yan, Q.; Zhou, C.; Tan, Z. Diet with a high proportion of rice alters profiles and potential function of digesta-associated microbiota in the ileum of goats. Animals 2020, 10, 1261. [Google Scholar] [CrossRef] [PubMed]
- Ma, W.; Wei, S.; Peng, W.; Sun, T.; Huang, J.; Yu, R.; Zhang, B.; Li, W. Antioxidant effect of polygonatum sibiricum polysaccharides in D-galactose-induced heart aging Mice. Biomed. Res. Int. 2021, 2021, 6688855. [Google Scholar] [CrossRef]
- Deng, Q.; Shao, Y.; Wang, Q.; Li, J.; Li, Y.; Ding, X.; Huang, P.; Yin, J.; Yang, H.; Yin, Y. Effects and interaction of dietary electrolyte balance and citric acid on growth performance, intestinal histomorphology, digestive enzyme activity and nutrient transporters expression of weaned piglets. J. Anim. Physiol. Anim. Nutr. Berl. 2021, 105, 272–285. [Google Scholar] [CrossRef]
- Wang, H.; Hu, C.; Cheng, C.; Cui, J.; Ji, Y.; Hao, X.; Li, Q.; Ren, W.; Deng, B.; Yin, Y.; et al. Unraveling the association of fecal microbiota and oxidative stress with stillbirth rate of sows. Theriogenology 2019, 136, 131–137. [Google Scholar] [CrossRef]
- Wang, K.; Yang, A.; Peng, X.; Lv, F.; Wang, Y.; Cui, Y.; Wang, Y.; Zhou, J.; Si, H. Linkages of various calcium sources on immune performance, diarrhea rate, intestinal barrier, and post-gut microbial structure and function in piglets. Front. Nutr. 2022, 9, 921773. [Google Scholar]
- Patra, A.K.; Saxena, J. Exploitation of dietary tannins to improve rumen metabolism and ruminant nutrition. J. Sci. Food. Agric. 2011, 91, 24–37. [Google Scholar] [CrossRef]
- McNabb, W.C.; Waghorn, G.C.; Barry, T.N.; Shelton, I.D. The effect of condensed tannins in Lotus pedunculatus on the digestion and metabolism of methionine, cystine and inorganic sulphur in sheep. Br. J. Nutr. 1993, 70, 647–661. [Google Scholar] [CrossRef]
- Redondo, L.M.; Chacana, P.A.; Dominguez, J.E.; Fernandez, M.M. Perspectives in the use of tannins as alternative to antimicrobial growth promoter factors in poultry. Front. Microbiol. 2014, 5, 118. [Google Scholar] [CrossRef]
- Dalle Zotte, A.; Cullere, M.; Tasoniero, G.; Gerencser, Z.; Szendro, Z.; Novelli, E.; Matics, Z. Supplementing growing rabbit diets with chestnut hydrolyzable tannins: Effect on meat quality and oxidative status, nutrient digestibilities, and content of tannin metabolites. Meat Sci. 2018, 146, 101–108. [Google Scholar] [CrossRef]
- Schiavone, A.; Guo, K.; Tassone, S.; Gasco, L.; Hernandez, E.; Denti, R.; Zoccarato, I. Effects of a natural extract of chestnut wood on digestibility, performance traits, and nitrogen balance of broiler chicks. Poult. Sci. 2008, 87, 521–527. [Google Scholar] [CrossRef]
- Lorenz, M.M.; Hayot Carbonero, C.; Smith, L.; Uden, P. In vitro protein degradation of 38 sainfoin accessions and its relationship to tannin content by different assays. J. Agric. Food Chem. 2012, 60, 5071–5075. [Google Scholar] [CrossRef]
- Hassan, Z.M.; Manyelo, T.G.; Selaledi, L.; Mabelebele, M. The effects of tannins in monogastric animals with special reference to alternative feed ingredients. Molecules 2020, 25, 4680. [Google Scholar] [CrossRef]
- Mašek, T.; Starčević, K. The influence of the addition of thymol, tannic acid or gallic acid to broiler diet on growth performance, serum malondialdehyde value and cecal fermentation. Yeast 2014, 5, 1–8. [Google Scholar] [CrossRef]
- Kubena, L.F.; Byrd, J.A.; Young, C.R.; Corrier, D.E. Effects of tannic acid on cecal volatile fatty acids and susceptibility to Salmonella typhimurium colonization in broiler chicks. Poult. Sci. 2001, 80, 1293–1298. [Google Scholar] [CrossRef]
- Marzo, F.; Urdaneta, E.; Santidrian, S. Liver proteolytic activity in tannic acid-fed birds. Poult. Sci. 2002, 81, 92–94. [Google Scholar] [CrossRef]
- Buyse, K.; Delezie, E.; Goethals, L.; Van Noten, N.; Ducatelle, R.; Janssens, G.; Lourenco, M. Chestnut tannins in broiler diets: Performance, nutrient digestibility, and meat quality. Poult. Sci. 2021, 100, 101479. [Google Scholar] [CrossRef]
- Jamroz, D.; Wiliczkiewicz, A.; Skorupińska, J.; Orda, J.; Kuryszko, J.; Tschirch, H. Effect of sweet chestnut tannin (SCT) on the performance, microbial status of intestine and histological characteristics of intestine wall in chickens. Br. Poult. Sci. 2009, 50, 687–699. [Google Scholar] [CrossRef]
- Tonda, R.M.; Rubach, J.K.; Lumpkins, B.S.; Mathis, G.F.; Poss, M.J. Effects of tannic acid extract on performance and intestinal health of broiler chickens following coccidiosis vaccination and/or a mixed-species Eimeria challenge. Poult. Sci. 2018, 97, 3031–3042. [Google Scholar] [CrossRef]
- Makkar, H.P.S. Effects and fate of tannins in ruminant animals, adaptation to tannins, and strategies to overcome detrimental effects of feeding tannin-rich feeds. Small Rumin. Res. 2003, 49, 241–256. [Google Scholar] [CrossRef]
- Mueller-Harvey, I. Unravelling the conundrum of tannins in animal nutrition and health. J. Sci. Food Agric. 2006, 86, 2010–2037. [Google Scholar] [CrossRef]
- Hussain, T.; Tan, B.; Yin, Y.; Blachier, F.; Tossou, M.C.; Rahu, N. Oxidative Stress and Inflammation: What polyphenols can do for Us. Oxid. Med. Cell. Longev. 2016, 2016, 7432797. [Google Scholar] [CrossRef] [PubMed]
- Samuel, K.G.; Wang, J.; Yue, H.Y.; Wu, S.G.; Zhang, H.J.; Duan, Z.Y.; Qi, G.H. Effects of dietary gallic acid supplementation on performance, antioxidant status, and jejunum intestinal morphology in broiler chicks. Poult. Sci. 2017, 96, 2768–2775. [Google Scholar]
- Dong, S.; Li, H.; Gasco, L.; Xiong, Y.; Guo, K.J.; Zoccarato, I. Antioxidative activity of the polyphenols from the involucres of Castanea mollissima Blume and their mitigating effects on heat stress. Poult. Sci. 2015, 94, 1096–1104. [Google Scholar] [CrossRef]
- De Hoyos-Martínez, P.L.; Merle, J.; Labidi, J. Tannins extraction: A key point for their valorization and cleaner production. Clean. Prod. 2019, 206, 1138–1155. [Google Scholar] [CrossRef]
- Sieniawska, E. Activities of tannins—From in vitro studies to clinical trials. Nat. Prod. Commun. 2015, 10, 1877–1884. [Google Scholar] [CrossRef]
- Kitada, S.; Leone, M.; Sareth, S.; Zhai, D.; Reed, J.C.; Pellecchia, M. Discovery, characterization, and structure-activity relationships studies of proapoptotic polyphenols targeting B-cell lymphocyte/leukemia-2 proteins. J. Med. Chem. 2003, 46, 4259–4264. [Google Scholar] [CrossRef]
- Cao, G.; Sofic, E.; Prior, R.L. Antioxidant and prooxidant behavior of flavonoids: Structure-activity relationships. Free Radic. Biol. Med. 1997, 22, 749–760. [Google Scholar] [CrossRef]
- Sekher Pannala, A.; Chan, T.S.; O’Brien, P.J.; Rice-Evans, C.A. Flavonoid B-ring chemistry and antioxidant activity: Fast reaction kinetics. Biochem. Biophys. Res. Commun. 2001, 282, 1161–1168. [Google Scholar] [CrossRef]
- Yahfoufi, N.; Alsadi, N.; Jambi, M.; Matar, C. The Immunomodulatory and anti-inflammatory role of polyphenols. Nutrients 2018, 10, 1618. [Google Scholar] [CrossRef]
- Mishra, A.; Sharma, A.K.; Kumar, S.; Saxena, A.K.; Pandey, A.K. Bauhinia variegata leaf extracts exhibit considerable antibacterial, antioxidant, and anticancer activities. Biomed. Res. Int. 2013, 2013, 915436. [Google Scholar]
- Li, J.; Yin, L.; Wang, L.; Li, J.; Huang, P.; Yang, H.; Yin, Y. Effects of vitamin B6 on growth, diarrhea rate, intestinal morphology, function, and inflammatory factors expression in a high-protein diet fed to weaned piglets. J. Anim. Sci. 2019, 97, 4865–4874. [Google Scholar]
- Cao, G.; Zeng, X.; Liu, J.; Yan, F.; Xiang, Z.; Wang, Y.; Tao, F.; Yang, C. Change of serum metabolome and cecal microflora in broiler chickens supplemented with grape seed extracts. Front. Immunol. 2020, 11, 610934. [Google Scholar]
- Huo, S.; Zhang, J.; Fan, J.; Wang, X.; Wu, F.; Zuo, Y.; Zhong, F. Co-Expression of Chicken IL-2 and IL-7 Enhances the Immunogenicity and Protective Efficacy of a VP2-Expressing DNA Vaccine against IBDV in Chickens. Viruses 2019, 11, 476. [Google Scholar] [CrossRef]
- Abbas, A.K.; Trotta, E.; Simeonov, D.R.; Marson, A.; Bluestone, J.A. Revisiting IL-2: Biology and therapeutic prospects. Sci. Immunol. 2018, 3, eaat1482. [Google Scholar]
- Rothwell, L.; Hu, T.; Wu, Z.; Kaiser, P. Chicken interleukin-21 is costimulatory for T cells and blocks maturation of dendritic cells. Dev. Comp. Immunol. 2012, 36, 475–482. [Google Scholar]
- Amer, S.A.; Al-Khalaifah, H.S.; Gouda, A.; Osman, A.; Goda, N.; Mohammed, H.A.; Darwish, M.; Hassan, A.M.; Mohamed, S. Potential Effects of Anthocyanin-Rich Roselle (Hibiscus sabdariffa L.) Extract on the Growth, Intestinal Histomorphology, Blood Biochemical Parameters, and the Immune Status of Broiler Chickens. Antioxidants 2022, 11, 544. [Google Scholar]
- Liu, H.; Hu, J.; Mahfuz, S.; Piao, X. Effects of Hydrolysable Tannins as Zinc Oxide Substitutes on Antioxidant Status, Immune Function, Intestinal Morphology, and Digestive Enzyme Activities in Weaned Piglets. Animals 2020, 10, 757. [Google Scholar] [CrossRef]
- Cornick, S.; Tawiah, A.; Chadee, K. Roles and regulation of the mucus barrier in the gut. Tissue Barriers 2015, 3, e982426. [Google Scholar] [CrossRef]
- Iqbal, Z.; Kamran, Z.; Sultan, J.I.; Ali, A.; Ahmad, S.; Shahzad, M.I.; Ahsan, U.; Ashraf, S.; Sohail, M.U. Replacement effect of vitamin E with grape polyphenols on antioxidant status, immune, and organs histopathological responses in broilers from 1- to 35-d age. J. Appl. Poult. Res. 2015, 24, 127–134. [Google Scholar] [CrossRef]
- Wu, J.; He, C.; Bu, J.; Luo, Y.; Yang, S.; Ye, C.; Yu, S.; He, B.; Yin, Y.; Yang, X. Betaine attenuates LPS-induced downregulation of Occludin and Claudin-1 and restores intestinal barrier function. BMC Vet. Res. 2020, 16, 75. [Google Scholar] [CrossRef] [PubMed]
- Song, P.; Zhang, R.; Wang, X.; He, P.; Tan, L.; Ma, X. Dietary grape-seed procyanidins decreased postweaning diarrhea by modulating intestinal permeability and suppressing oxidative stress in rats. J. Agric. Food Chem. 2011, 59, 6227–6232. [Google Scholar] [CrossRef] [PubMed]
- Nallathambi, R.; Poulev, A.; Zuk, J.B.; Raskin, I. Proanthocyanidin-Rich Grape Seed Extract Reduces Inflammation and Oxidative Stress and Restores Tight Junction Barrier Function in Caco-2 Colon Cells. Nutrients 2020, 12, 1623. [Google Scholar] [CrossRef] [PubMed]
- Feldman, G.J.; Mullin, J.M.; Ryan, M.P. Occludin: Structure, function and regulation. Adv. Drug Deliv. Rev. 2005, 57, 883–917. [Google Scholar] [CrossRef]
- Liu, W.C.; Guo, Y.; An, L.L.; Zhao, Z.H. Protective effects of dietary betaine on intestinal barrier function and cecal microbial community in indigenous broiler chickens exposed to high temperature environment. Environ. Sci. Pollut. Res. Int. 2021, 28, 10860–10871. [Google Scholar] [CrossRef]
- Hikino, H.; Kiso, Y.; Hatano, T.; Yoshida, T.; Okuda, T. Antihepatotoxic actions of tannins. J. Ethnopharmacol. 1985, 14, 19. [Google Scholar]
- Chen, C.; Wang, Z.; Li, J.; Li, Y.; Huang, P.; Ding, X.; Yin, J.; He, S.; Yang, H.; Yin, Y. Dietary vitamin E affects small intestinal histomorphology, digestive enzyme activity, and the expression of nutrient transporters by inhibiting proliferation of intestinal epithelial cells within jejunum in weaned piglets. J. Anim. Sci. 2019, 97, 1212–1221. [Google Scholar] [CrossRef]
- Wang, J.X.; Peng, K.M. Developmental morphology of the small intestine of African ostrich chicks. Poult. Sci. 2008, 87, 2629–2635. [Google Scholar] [CrossRef]
- Liu, H.W.; Li, K.; Zhao, J.S.; Deng, W. Effects of chestnut tannins on intestinal morphology, barrier function, pro-inflammatory cytokine expression, microflora and antioxidant capacity in heat-stressed broilers. J. Anim. Physiol. Anim. Nutr. Berl. 2018, 102, 717–726. [Google Scholar] [CrossRef]
- Tong, Z.; Lei, F.; Liu, L.; Wang, F.; Guo, A. Effects of Plotytarya strohilacea Sieb. et Zuce Tannin on the Growth Performance, Oxidation Resistance, Intestinal Morphology and Cecal Microbial Composition of Broilers. Front. Vet. Sci. 2021, 8, 806105. [Google Scholar] [CrossRef]
- Wang, K.; Peng, X.; Lv, F.; Zheng, M.; Long, D.; Mao, H.; Si, H.; Zhang, P. Microbiome-Metabolites Analysis Reveals Unhealthy Alterations in the Gut Microbiota but Improved Meat Quality with a High-Rice Diet Challenge in a Small Ruminant Model. Animals 2021, 11, 2306. [Google Scholar] [CrossRef]
- Ji, F.J.; Wang, L.X.; Yang, H.S.; Hu, A.; Yin, Y.L. Review: The roles and functions of glutamine on intestinal health and performance of weaning pigs. Animal 2019, 13, 2727–2735. [Google Scholar] [CrossRef]
- Barnes, E.M.; Mead, G.C.; Barnum, D.A.; Harry, E.G. The intestinal flora of the chicken in the period 2 to 6 weeks of age, with particular reference to the anaerobic bacteria. Br. Poult. Sci. 1972, 13, 311–326. [Google Scholar] [CrossRef]
- Sun, Y.; Liu, Y.; Ai, C.; Song, S.; Chen, X. Caulerpa lentillifera polysaccharides enhance the immunostimulatory activity in immunosuppressed mice in correlation with modulating gut microbiota. Food Funct. 2019, 10, 4315–4329. [Google Scholar] [CrossRef]
- Rehman, A.; Lepage, P.; Nolte, A.; Hellmig, S.; Schreiber, S.; Ott, S.J. Transcriptional activity of the dominant gut mucosal microbiota in chronic inflammatory bowel disease patients. J. Med. Microbiol. 2010, 59, 1114–1122. [Google Scholar] [CrossRef]
- Jandhyala, S.M.; Talukdar, R.; Subramanyam, C.; Vuyyuru, H.; Sasikala, M.; Nageshwar, R.D. Role of the normal gut microbiota. World J. Gastroenterol. 2015, 21, 8787–8803. [Google Scholar] [CrossRef]
- Fesseha, H.; Demlie, T.; Mathewos, M.; Eshetu, E. Effect of Lactobacillus Species Probiotics on Growth Performance of Dual-Purpose Chicken. Vet. Med. Auckl. 2021, 12, 75–83. [Google Scholar] [CrossRef]
- Khaksefidi, A.; Rahimi, S. Effect of probiotic inclusion in the diet of broiler chickens on performance, feed efficiency and carcass quality. Asian-Australas. J. Anim. Sci. 2005, 18, 1153–1156. [Google Scholar] [CrossRef]
- Durre, P. Physiology and Sporulation in Clostridium. Microbiol. Spectr. 2014, 2, 10–2012. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Wang, H.; Wang, B.; Zhang, B.; Li, W. Effects of manganese and Bacillus subtilis on the reproductive performance, egg quality, antioxidant capacity, and gut microbiota of breeding geese during laying period. Poult. Sci. 2020, 99, 6196–6204. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.K.; Zhang, X.X.; Li, F.D.; Li, C.; Li, G.Z.; Zhang, D.Y.; Song, Q.Z.; Li, X.L.; Zhao, Y.; Wang, W.M. Characterization of the rumen microbiota and its relationship with residual feed intake in sheep. Animal 2021, 15, 100161. [Google Scholar] [CrossRef]
- Weinert-Nelson, J.R.; Biddle, A.S.; Williams, C.A. Fecal microbiome of horses transitioning between warm-season and cool-season grass pasture within integrated rotational grazing systems. Anim. Microbiome 2022, 4, 41. [Google Scholar] [CrossRef] [PubMed]
- Niu, K.; Bai, P.; Yang, B.; Feng, X.; Qiu, F. Asiatic acid alleviates metabolism disorders in ob/ob mice: Mechanistic insights. Food Funct. 2022, 13, 6934–6946. [Google Scholar] [CrossRef] [PubMed]
- Zou, X.Y.; Zhang, M.; Tu, W.J.; Zhang, Q.; Jin, M.L.; Fang, R.D.; Jiang, S. Bacillus subtilis inhibits intestinal inflammation and oxidative stress by regulating gut flora and related metabolites in laying hens. Animal 2022, 16, 100474. [Google Scholar] [CrossRef]
- Wen, C.; Yan, W.; Mai, C.; Duan, Z.; Zheng, J.; Sun, C.; Yang, N. Joint contributions of the gut microbiota and host genetics to feed efficiency in chickens. Microbiome 2021, 9, 126. [Google Scholar] [CrossRef]
- Zhang, J.; Yang, G.; Wen, Y.; Liu, S.; Li, C.; Yang, R.; Li, W. Intestinal microbiota are involved in the immunomodulatory activities of longan polysaccharide. Mol. Nutr. Food Res. 2017, 61, 1700466. [Google Scholar] [CrossRef]
- Trutschel, L.R.; Chadwick, G.L.; Kruger, B.; Blank, J.G.; Brazelton, W.J.; Dart, E.R.; Rowe, A.R. Investigation of microbial metabolisms in an extremely high pH marine-like terrestrial serpentinizing system: Ney Springs. Sci. Total Environ. 2022, 836, 155492. [Google Scholar] [CrossRef]
- Lutsiv, T.; McGinley, J.N.; Neil-McDonald, E.S.; Weir, T.L.; Foster, M.T.; Thompson, H.J. Relandscaping the Gut Microbiota with a Whole Food: Dose-Response Effects to Common Bean. Foods 2022, 11, 1153. [Google Scholar] [CrossRef]
- Li, X.; Yi, Y.; Wu, J.; Yang, Q.; Tan, B.; Chi, S. Effects of Plant-Derived Glycerol Monolaurate (GML) Additive on the Antioxidant Capacity, Anti-Inflammatory Ability, Muscle Nutritional Value, and Intestinal Flora of Hybrid Grouper (Epinephelus fuscoguttatus x Epinephelus lanceolatus). Metabolites 2022, 12, 1089. [Google Scholar] [CrossRef]
- Molino, S.; Lerma-Aguilera, A.; Jiménez-Hernández, N.; Rufián Henares, J.Á.; Francino, M.P. Evaluation of the effects of a short supplementation with tannins on the gut microbiota of healthy subjects. Front. Microbiol. 2022, 13, 1141. [Google Scholar] [CrossRef]
- Wang, B.; Luo, H. Effects of mulberry leaf silage on antioxidant and immunomodulatory activity and rumen bacterial community of lambs. BMC Microbiol. 2021, 21, 250. [Google Scholar] [CrossRef]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Items | Days 1–14 | Days 15–28 | Days 29–42 |
|---|---|---|---|
| Ingredients, % | |||
| Corn | 56.80 | 57.36 | 58.88 |
| Soybean meal | 36.48 | 34.57 | 33.01 |
| Soybean oil | 1.63 | 3.42 | 3.89 |
| NaCl | 0.53 | 0.53 | 0.53 |
| Limestone | 1.40 | 1.22 | 1.05 |
| CaHPO4 | 2.10 | 1.99 | 1.85 |
| L-Lys•HCl | 0.30 | 0.15 | 0.06 |
| DL-Met | 0.26 | 0.26 | 0.23 |
| Choline chloride | 0.10 | 0.10 | 0.10 |
| Premix 1 | 0.40 | 0.40 | 0.40 |
| Total | 100.00 | 100.00 | 100.00 |
| Nutrient levels 2 | |||
| AME, MJ/kg | 12.89 | 13.50 | 13.94 |
| Crude protein, % | 21.00 | 20.00 | 20.00 |
| Calcium, % | 1.01 | 1.00 | 0.95 |
| Available phosphorus, % | 0.62 | 0.60 | 0.57 |
| Lysine, % | 1.04 | 0.91 | 0.88 |
| Methionine, % | 0.41 | 0.40 | 0.38 |
| Threonine, % | 0.74 | 0.70 | 0.70 |
| Tryptophan, % | 0.22 | 0.19 | 0.18 |
| Age | CON | AT | CT | QT | TT | p-Value |
|---|---|---|---|---|---|---|
| 7 days | 0.163 ± 0.005 | 0.163 ± 0.004 | 0.163 ± 0.005 | 0.163 ± 0.004 | 0.163 ± 0.002 | 1.000 |
| 14 days | 0.504 ± 0.015 | 0.504 ± 0.013 | 0.499 ± 0.011 | 0.505 ± 0.008 | 0.495 ± 0.003 | 0.961 |
| 21 days | 1.054 ± 0.039 | 1.052 ± 0.029 | 1.033 ± 0.030 | 1.063 ± 0.020 | 1.018 ± 0.015 | 0.789 |
| 28 days | 1.790 ± 0.073 | 1.767 ± 0.052 | 1.744 ± 0.059 | 1.790 ± 0.028 | 1.740 ± 0.038 | 0.928 |
| 35 days | 2.619 ± 0.104 | 2.585 ± 0.075 | 2.552 ± 0.095 | 2.523 ± 0.032 | 2.496 ± 0.050 | 0.803 |
| 42 days | 3.370 ± 0.105 | 3.320 ± 0.095 | 3.329 ± 0.118 | 3.252 ± 0.047 | 3.157 ± 0.045 | 0.478 |
| 49 days | 3.923 ± 0.132 | 3.908 ± 0.077 | 3.932 ± 0.125 | 3.837 ± 0.098 | 3.730 ± 0.077 | 0.627 |
| Parameters | CON | AT | CT | QT | TT | p-Value |
|---|---|---|---|---|---|---|
| ADFI (kg) | 0.143 ± 0.004 | 0.142 ± 0.005 | 0.140 ± 0.005 | 0.139 ± 0.001 | 0.139 ± 0.002 | 0.915 |
| ADG (kg) | 0.089 ± 0.003 | 0.089 ± 0.002 | 0.090 ± 0.003 | 0.087 ± 0.002 | 0.084 ± 0.002 | 0.611 |
| FCR | 1.602 ± 0.020 | 1.586 ± 0.031 | 1.564 ± 0.044 | 1.596 ± 0.037 | 1.634 ± 0.016 | 0.632 |
| Parameters | CON | AT | CT | QT | TT | p-Value |
|---|---|---|---|---|---|---|
| DR (%) | 0.30 ± 0.06 | 0.36 ± 0.04 | 0.28 ± 0.04 | 0.25 ± 0.02 | 0.30 ± 0.04 | 0.417 |
| CL (cm) | 17.48 ± 0.61 | 19.12 ± 0.47 | 18.50 ± 0.51 | 17.57 ± 0.57 | 17.77 ± 0.77 | 0.251 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, S.; Wang, K.; Lin, S.; Zhang, Z.; Cheng, M.; Hu, S.; Hu, H.; Xiang, J.; Chen, F.; Li, G.; et al. Comparison of the Effects between Tannins Extracted from Different Natural Plants on Growth Performance, Antioxidant Capacity, Immunity, and Intestinal Flora of Broiler Chickens. Antioxidants 2023, 12, 441. https://doi.org/10.3390/antiox12020441
Liu S, Wang K, Lin S, Zhang Z, Cheng M, Hu S, Hu H, Xiang J, Chen F, Li G, et al. Comparison of the Effects between Tannins Extracted from Different Natural Plants on Growth Performance, Antioxidant Capacity, Immunity, and Intestinal Flora of Broiler Chickens. Antioxidants. 2023; 12(2):441. https://doi.org/10.3390/antiox12020441
Chicago/Turabian StyleLiu, Song, Kaijun Wang, Senzhu Lin, Zhidan Zhang, Ming Cheng, Shanshan Hu, Hongjie Hu, Jun Xiang, Fengming Chen, Gonghe Li, and et al. 2023. "Comparison of the Effects between Tannins Extracted from Different Natural Plants on Growth Performance, Antioxidant Capacity, Immunity, and Intestinal Flora of Broiler Chickens" Antioxidants 12, no. 2: 441. https://doi.org/10.3390/antiox12020441