
The Aftermath of Long-Term Cigarette Smoking on Telomere Length and Mitochondrial DNA Copy Number in Human Cumulus Cells Prior to In Vitro Fertilization—A Pilot Study

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethical Approval
2.2. Source of Cumulus Cells and Patients’ Selection Criteria
2.3. Ovarian Stimulation Protocols
2.4. Cumulus Cells Isolation
2.5. DNA, RNA and Protein Extraction
2.6. Telomere Length and Mitochondrial DNA Copy Number Measurement
2.7. Telomere Gene Expression Analysis
2.8. MicroRNA Target Analysis
2.9. Protein Target Analysis
3. Results
3.1. Patients’ Characteristics
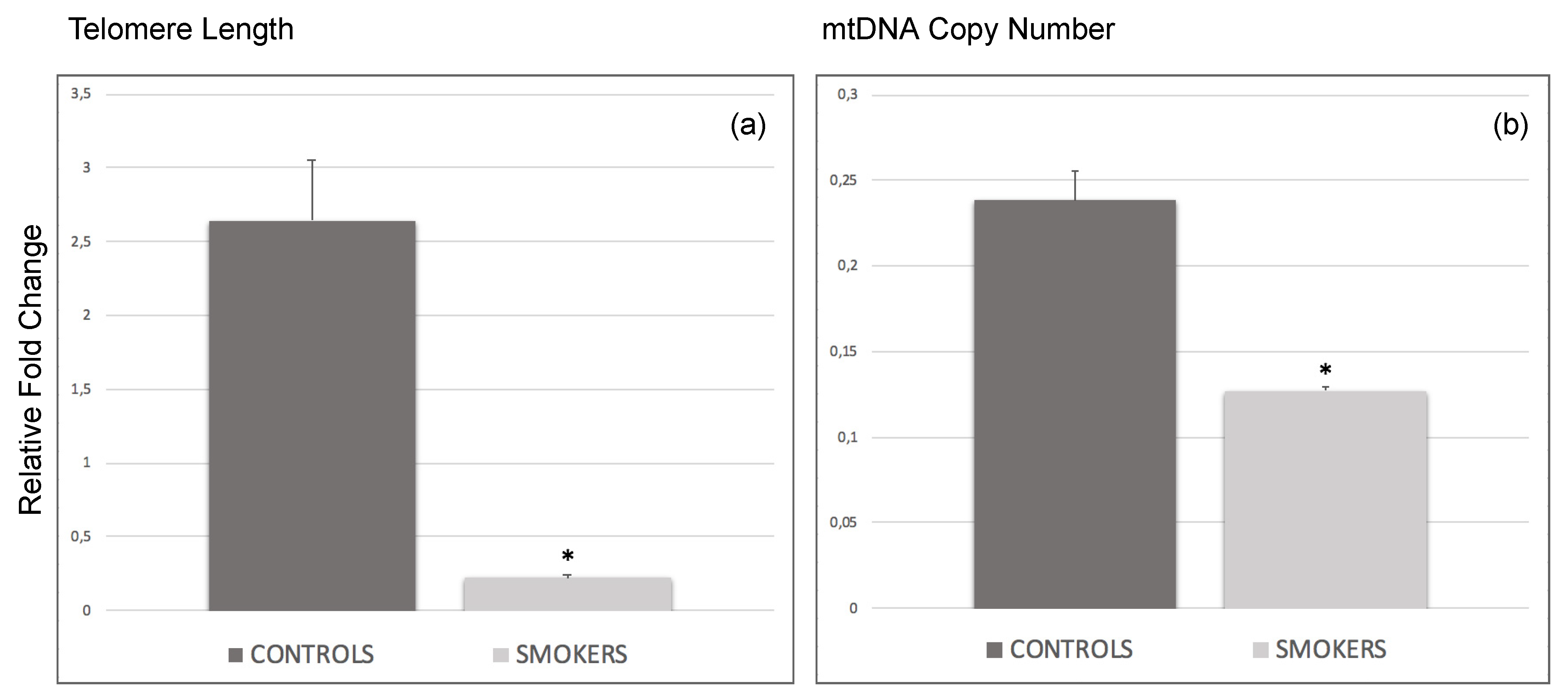
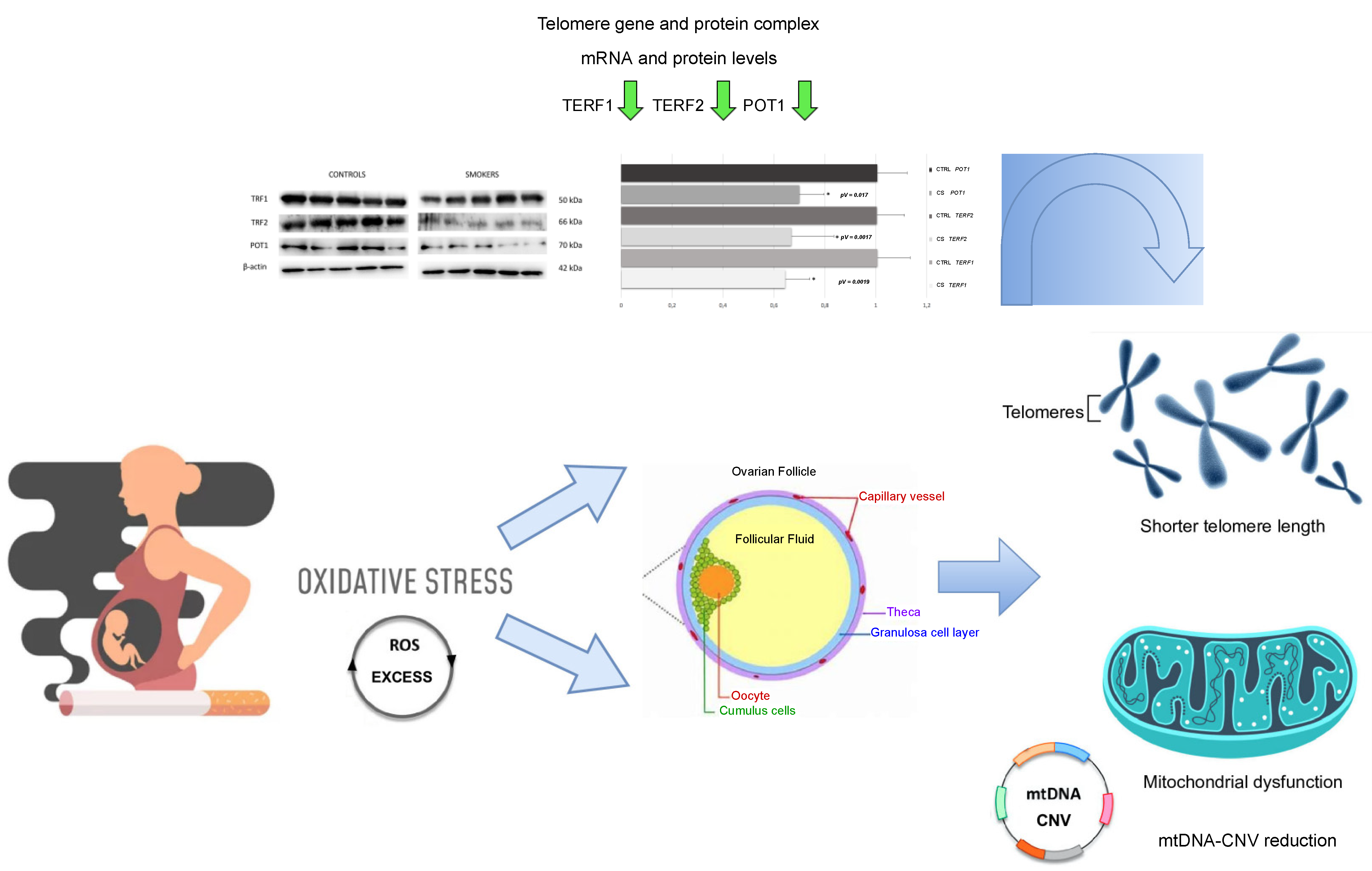
3.2. Decrease of Average Telomere Length and mtDNA Copy Number in Smokers’ CCs
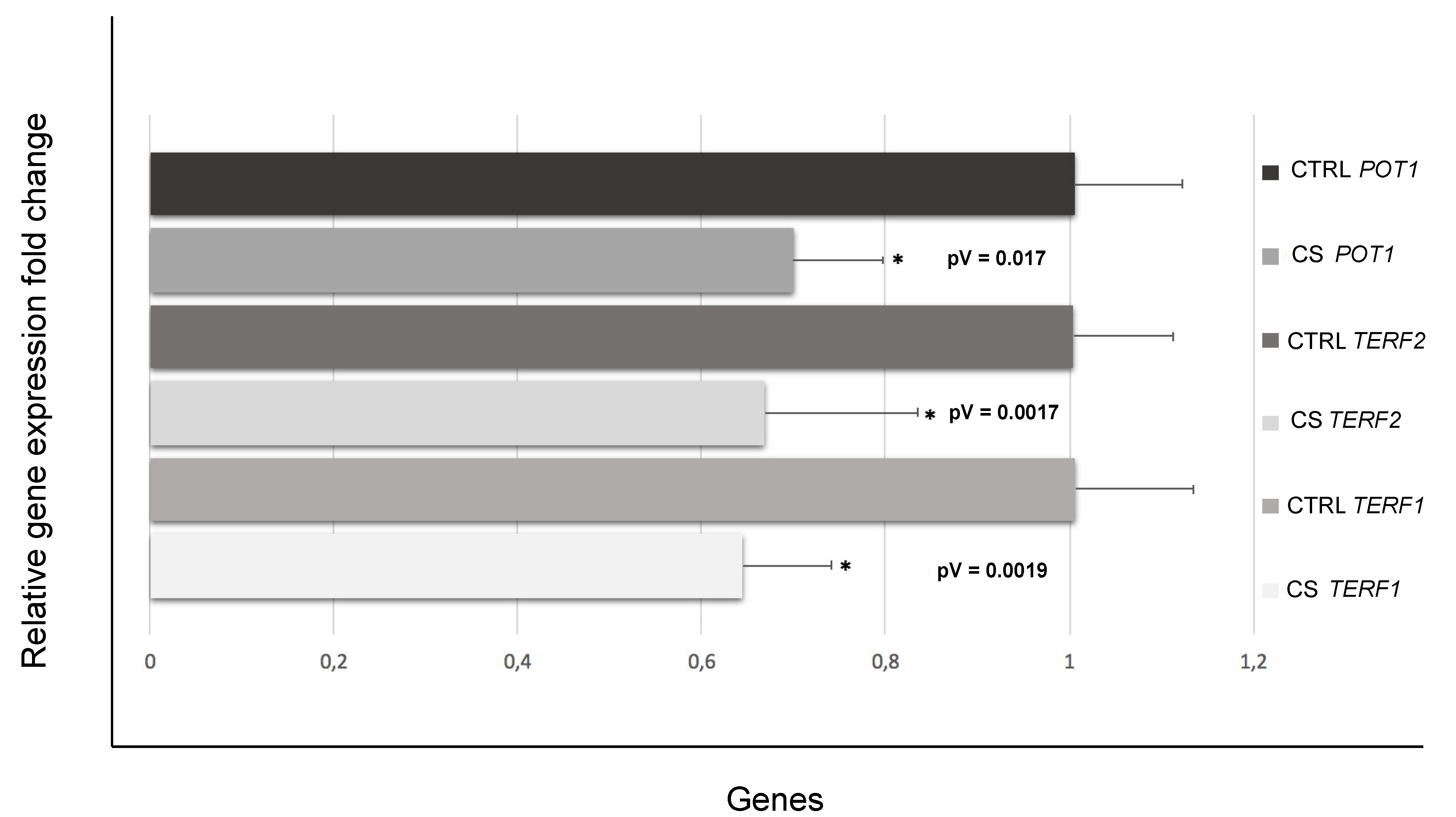
3.3. Down-Regulation of Genes Involved in Telomere Length and Protection
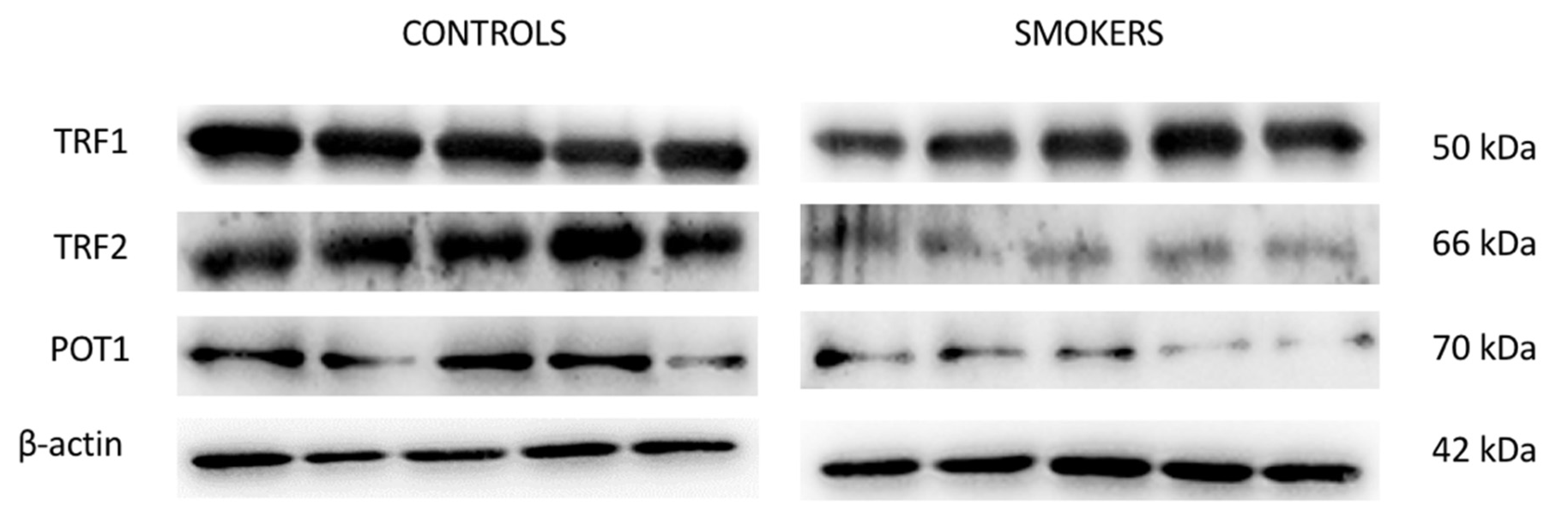
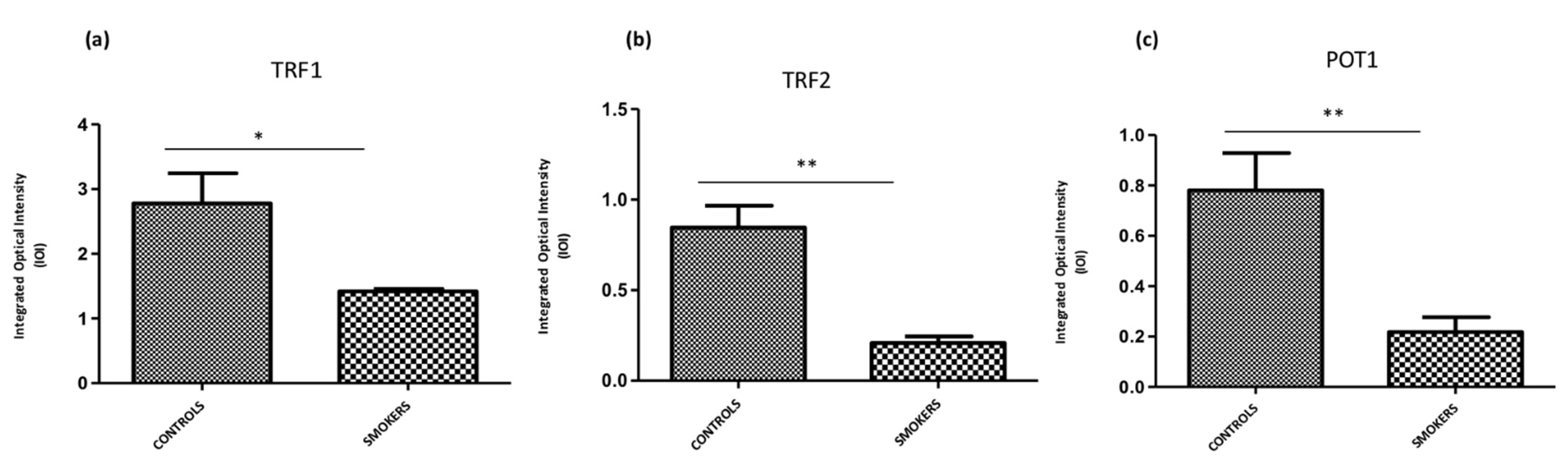
3.4. Expression of TERF1, TERF2 and POT1 Protein Levels
3.5. Expression Profiling of miR-155, miR-23a and miR-185
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- James, P.; Bah, A.J.; Kabba, J.; Dalinjong, P.; Kassim, S.A.; Dalinjong, P.A. Prevalence and Correlates of Current Tobacco use and Non-user Susceptibility to using Tobacco Product Among School-going Adolescents in 22 African Countries: A secondary analysis of the 2013–2018 global youth tobacco surveys. Arch. Public Health 2022, 80, 121. [Google Scholar] [CrossRef] [PubMed]
- Tsai, P.C.; Glastonbury, C.A.; Eliot, M.N.; Bollepalli, S.; Yet, I.; Castillo-Fernandez, J.E.; Carnero-Montoro, E.; Hardiman, T.; Martin, T.C.; Vickers, A.; et al. Smoking induces coordinated DNA methylation and gene expression changes in adipose tissue with consequences for metabolic health. Clin. Epigenetics 2018, 10, 126. [Google Scholar] [CrossRef] [PubMed]
- De Angelis, C.; Nardone, A.; Garifalos, F.; Pivonello, C.; Sansone, A.; Conforti, A.; Di Dato, C.; Sirico, F.; Alviggi, C.; Isidori, A.; et al. Smoke, alcohol and drug addiction and female fertility. Reprod. Biol. Endocrinol. 2020, 18, 21. [Google Scholar] [CrossRef] [PubMed]
- Kurti, A.N.; Bunn, J.Y.; Villanti, A.C.; Stanton, C.A.; Redner, R.; Lopez, A.A.; Gaalema, D.E.; Doogan, N.J.; Cepeda-Benito, A.; Roberts, M.E.; et al. Patterns of Single and Multiple Tobacco Product Use Among US Women of Reproductive Age. Nicotine Tob Res. 2018, 20 (Suppl. 1), S71–S80. [Google Scholar] [CrossRef]
- Do, E.K.; Nicksic, N.E.; Clifford, J.S.; Hayes, A.; Fuemmeler, B.F. Perceived harms of and exposure to tobacco use and current tobacco use among reproductive-aged women from the PATH study. Women Health 2020, 60, 1040–1051. [Google Scholar] [CrossRef]
- Budani, M.C.; D’Aurora, M.; Stuppia, L.; Gatta, V.; Tiboni, G.M. Whole-body exposure to cigarette smoke alters oocyte miRNAs expression in C57BL/6 mice. Mol. Reprod. Dev. 2019, 86, 1741–1757. [Google Scholar] [CrossRef]
- Konstantinidou, F.; Stuppia, L.; Gatta, V. Looking Inside the World of Granulosa Cells: The Noxious Effects of Cigarette Smoke. Biomedicines 2020, 8, 309. [Google Scholar] [CrossRef]
- Kidder, G.M.; Vanderhyden, B.C. Bidirectional communication between oocytes and follicle cells: Ensuring oocyte developmental competence. Can. J. Physiol. Pharmacol. 2010, 88, 399–413. [Google Scholar] [CrossRef]
- Gannon, A.M.; Staämpfli, M.R.; Foster, W.G. Cigarette smoke exposure leads to follicle loss via an alternative ovarian cell death pathway in a mouse model. Toxicol. Sci. 2012, 125, 274–284. [Google Scholar] [CrossRef]
- Konstantinidou, F.; Budani, M.C.; Sarra, A.; Stuppia, L.; Tiboni, G.M.; Gatta, V. Impact of Cigarette Smoking on the Expression of Oxidative Stress-Related Genes in Cumulus Cells Retrieved from Healthy Women Undergoing IVF. Int. J. Mol. Sci. 2021, 22, 13147. [Google Scholar] [CrossRef]
- Fetterman, J.L.; Sammy, M.J.; Ballinger, S.W. Mitochondrial toxicity of tobacco smoke and air pollution. Toxicology 2017, 391, 18–33. [Google Scholar] [CrossRef] [PubMed]
- O’Sullivan, R.J.; Karlseder, J. Telomeres: Protecting chromosomes against genome instability. Nat. Rev. Mol. Cell Biol. 2010, 11, 171–181. [Google Scholar] [CrossRef] [PubMed]
- Vaiserman, A.; Krasnienkov, D. Telomere Length as a Marker of Biological Age: State-of-the-Art, Open Issues, and Future Perspectives. Front. Genet. 2021, 11, 630186. [Google Scholar] [CrossRef] [PubMed]
- Astuti, Y.; Wardhana, A.; Watkins, J.; Wulaningsih, W.; PILAR Research Network. Cigarette smoking and telomere length: A systematic review of 84 studies and meta-analysis. Environ. Res. 2017, 158, 480–489. [Google Scholar] [CrossRef]
- Di Nottia, M.; Verrigni, D.; Torraco, A.; Rizza, T.; Bertini, E.; Carrozzo, R. Mitochondrial Dynamics: Molecular Mechanisms, Related Primary Mitochondrial Disorders and Therapeutic Approaches. Genes 2021, 12, 247. [Google Scholar] [CrossRef]
- Hu, L.; Yao, X.; Shen, Y. Altered mitochondrial DNA copy number contributes to human cancer risk: Evidence from an updated meta-analysis. Sci. Rep. 2016, 6, 35859. [Google Scholar] [CrossRef]
- Jafari, A.; Rajabi, A.; Gholian-Aval, M.; Peyman, N.; Mahdizadeh, M.; Tehrani, H. National, regional, and global prevalence of cigarette smoking among women/females in the general population: A systematic review and meta-analysis. Environ. Health Prev. Med. 2021, 26, 5. [Google Scholar] [CrossRef]
- Wu, S.; Li, X.; Meng, S.; Fung, T.; Chan, A.T.; Liang, G.; Giovannucci, E.; De Vivo, I.; Lee, J.H.; Nan, H. Fruit and vegetable consumption, cigarette smoke, and leukocyte mitochondrial DNA copy number. Am. J. Clin. Nutr. 2019, 109, 424–432. [Google Scholar] [CrossRef]
- Lee, H.C.; Wei, Y.H. Mitochondrial biogenesis and mitochondrial DNA maintenance of mammalian cells under oxidative stress. Int. J. Biochem. Cell Biol. 2005, 37, 822–834. [Google Scholar] [CrossRef]
- Yu, M. Generation, function and diagnostic value of mitochondrial DNA copy number alterations in human cancers. Life Sci. 2011, 89, 65–71. [Google Scholar] [CrossRef]
- Westbrook, D.G.; Anderson, P.G.; Pinkerton, K.E.; Ballinger, S.W. Perinatal tobacco smoke exposure increases vascular oxidative stress and mitochondrial damage in non-human primates. Cardiovasc. Toxicol. 2010, 10, 216–226. [Google Scholar] [CrossRef] [PubMed]
- Lingner, J.; Cooper, J.; Cech, T. Telomerase and DNA end replication: No longer a lagging strand problem? Science 1995, 269, 1533–1534. [Google Scholar] [CrossRef] [PubMed]
- Srinivas, N.; Rachakonda, S.; Kumar, R. Telomeres and Telomere Length: A General Overview. Cancers 2020, 12, 558. [Google Scholar] [CrossRef] [PubMed]
- Reichert, S.; Stier, A. Does oxidative stress shorten telomeres in vivo? A review. Biol. Lett. 2017, 13, 20170463. [Google Scholar] [CrossRef] [PubMed]
- Fouquerel, E.; Lormand, J.; Bose, A.; Lee, H.T.; Kim, G.S.; Li, J.; Sobol, R.W.; Freudenthal, B.D.; Myong, S.; Opresko, P.L. Oxidative guanine base damage regulates human telomerase activity. Nat. Struct. Mol. Biol. 2016, 23, 1092–1100. [Google Scholar] [CrossRef]
- Rocca, M.S.; Foresta, C.; Ferlin, A. Telomere length: Lights and shadows on their role in human reproduction. Biol. Reprod. 2018, 100, 305–317. [Google Scholar] [CrossRef]
- Cheng, E.H.; Chen, S.U.; Lee, T.H.; Pai, Y.P.; Huang, L.S.; Huang, C.C.; Lee, M.S. Evaluation of telomere length in cumulus cells as a potential biomarker of oocyte and embryo quality. Hum. Reprod. 2013, 28, 929–936. [Google Scholar] [CrossRef]
- Yu, T.N.; Cheng, E.H.; Tsai, H.N.; Lin, P.Y.; Chen, C.H.; Huang, C.C.; Lee, T.H.; Lee, M.S. Assessment of Telomere Length and Mitochondrial DNA Copy Number in Granulosa Cells as Predictors of Aneuploidy Rate in Young Patients. J. Clin. Med. 2022, 11, 1824. [Google Scholar] [CrossRef]
- Keefe, D.L. Telomeres, Reproductive Aging, and Genomic Instability During Early Development. Reprod. Sci. 2016, 23, 1612–1615. [Google Scholar] [CrossRef]
- Xu, X.; Chen, X.; Zhang, X.; Liu, Y.; Wang, Z.; Wang, P.; Du, Y.; Qin, Y.; Chen, Z.J. Impaired telomere length and telomerase activity in peripheral blood leukocytes and granulosa cells in patients with biochemical primary ovarian insufficiency. Hum. Reprod. 2017, 32, 201–207. [Google Scholar] [CrossRef]
- Pedroso, D.C.C.; Santana, V.P.; Donaires, F.S.; Picinato, M.C.; Giorgenon, R.C.; Santana, B.A.; Pimentel, R.N.; Keefe, D.L.; Calado, R.T.; Ferriani, R.A.; et al. Telomere Length and Telomerase Activity in Immature Oocytes and Cumulus Cells of Women with Polycystic Ovary Syndrome. Reprod Sci. 2020, 27, 1293–1303. [Google Scholar] [CrossRef] [PubMed]
- Morin, S.J.; Tao, X.; Marin, D.; Zhan, Y.; Landis, J.; Bedard, J.; Scott, R.T.; Seli, E. DNA methylation-based age prediction and telomere length in white blood cells and cumulus cells of infertile women with normal or poor response to ovarian stimulation. Aging 2018, 10, 3761–3773. [Google Scholar] [CrossRef] [PubMed]
- Lara-Molina, E.E.; Franasiak, J.M.; Marin, D.; Tao, X.; Díaz-Gimeno, P.; Florensa, M.; Martin, M.; Seli, E.; Pellicer, A. Cumulus cells have longer telomeres than leukocytes in reproductive-age women. Fertil. Steril. 2020, 113, 217–223. [Google Scholar] [CrossRef] [PubMed]
- Sfeir, A.; de Lange, T. Removal of shelterin reveals the telomere end-protection problem. Science 2012, 336, 593–597. [Google Scholar] [CrossRef] [PubMed]
- de Lange, T. Shelterin: The protein complex that shapes and safeguards human telomeres. Genes Dev. 2005, 19, 2100–2110. [Google Scholar] [CrossRef]
- Iwano, T.; Tachibana, M.; Reth, M.; Shinkai, Y. Importance of TRF1 for functional telomere structure. J. Biol. Chem. 2004, 279, 1442–1448. [Google Scholar] [CrossRef]
- Martínez, P.; Thanasoula, M.; Muñoz, P.; Liao, C.; Tejera, A.; McNees, C.; Flores, J.M.; Fernández-Capetillo, O.; Tarsounas, M.; Blasco, M.A. Increased telomere fragility and fusions resulting from TRF1 deficiency lead to degenerative pathologies and increased cancer in mice. Genes Dev. 2009, 23, 2060–2075. [Google Scholar] [CrossRef]
- Smogorzewska, A.; van Steensel, B.; Bianchi, A.; Oelmann, S.; Schaefer, M.R.; Schnapp, G.; de Lange, T. Control of human telomere length by TRF1 and TRF2. Mol. Cell. Biol. 2000, 20, 1659–1668. [Google Scholar] [CrossRef]
- Pirzada, R.H.; Orun, O.; Erzik, C.; Cagsin, H.; Serakinci, N. Role of TRF2 and TPP1 regulation in idiopathic recurrent pregnancy loss. Int. J. Biol. Macromol. 2019, 127, 306–310. [Google Scholar] [CrossRef]
- Ye, J.; Lenain, C.; Bauwens, S.; Rizzo, A.; Saint-Léger, A.; Poulet, A.; Benarroch, D.; Magdinier, F.; Morere, J.; Amiard, S.; et al. TRF2 and apollo cooperate with topoisomerase 2alpha to protect human telomeres from replicative damage. Cell 2010, 142, 230–242. [Google Scholar] [CrossRef]
- Palm, W.; de Lange, T. How shelterin protects mammalian telomeres. Annu. Rev. Genet. 2008, 42, 301–334. [Google Scholar] [CrossRef] [PubMed]
- Veldman, T.; Etheridge, K.T.; Counter, C.M. Loss of hPot1 function leads to telomere instability and a cut-like phenotype. Curr. Biol. 2004, 14, 2264–2270. [Google Scholar] [CrossRef] [PubMed]
- Escoffier, E.; Rezza, A.; Roborel de Climens, A.; Belleville, A.; Gazzolo, L.; Gilson, E.; Duc Dodon, M. A balanced transcription between telomerase and the telomeric DNA-binding proteins TRF1, TRF2 and Pot1 in resting, activated, HTLV-1-transformed and Tax-expressing human T lymphocytes. Retrovirology 2005, 2, 77. [Google Scholar] [CrossRef] [PubMed]
- Hohensinner, P.J.; Kaun, C.; Buchberger, E.; Ebenbauer, B.; Demyanets, S.; Huk, I.; Eppel, W.; Maurer, G.; Huber, K.; Wojta, J. Age intrinsic loss of telomere protection via TRF1 reduction in endothelial cells. Biochim. Biophys. Acta 2016, 1863, 360–367. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Wu, X. Abnormal function of telomere protein TRF2 induces cell mutation and the effects of environmental tumor-promoting factors (Review). Oncol. Rep. 2021, 46, 184. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Zheng, Y.L.; Harris, C.C. POT1 and TRF2 cooperate to maintain telomeric integrity. Mol. Cell. Biol. 2005, 25, 1070–1080. [Google Scholar] [CrossRef]
- Harries, L.W. MicroRNAs as Mediators of the Ageing Process. Genes 2014, 5, 656–670. [Google Scholar] [CrossRef]
- Bhide, P.; Timlick, E.; Kulkarni, A.; Gudi, A.; Shah, A.; Homburg, R.; Acharya, G. Effect of cigarette smoking on serum anti-Mullerian hormone and antral follicle count in women seeking fertility treatment: A prospective cross-sectional study. BMJ Open. 2022, 12, e049646. [Google Scholar] [CrossRef]
- Sansone, M.; Zaami, S.; Cetta, L.; Costanzi, F.; Signore, F. Ovotoxicity of smoking and impact on AMH levels: A pilot study. Eur. Rev. Med. Pharmacol. Sci. 2021, 25, 5255–5260. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Non-Smokers (n = 5) | Smokers (n = 5) | p-Value | |
|---|---|---|---|
| Age (years) | 30.8 ± 0.8 | 33.2 ± 2.6 | NS |
| AMH (ng/mL) | 8.6 ± 2.5 | 9.2 ± 8.3 | NS |
| Basal FSH (mIU/mL) | 7.4 ± 3.7 | 6.8 ± 3.7 | NS |
| BMI (kg/m2) | 25.3 ± 4.5 | 24.6 ± 5.4 | NS |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Konstantinidou, F.; Budani, M.C.; Marconi, G.D.; Gonnella, F.; Sarra, A.; Trubiani, O.; Stuppia, L.; Tiboni, G.M.; Gatta, V. The Aftermath of Long-Term Cigarette Smoking on Telomere Length and Mitochondrial DNA Copy Number in Human Cumulus Cells Prior to In Vitro Fertilization—A Pilot Study. Antioxidants 2022, 11, 1841. https://doi.org/10.3390/antiox11091841
Konstantinidou F, Budani MC, Marconi GD, Gonnella F, Sarra A, Trubiani O, Stuppia L, Tiboni GM, Gatta V. The Aftermath of Long-Term Cigarette Smoking on Telomere Length and Mitochondrial DNA Copy Number in Human Cumulus Cells Prior to In Vitro Fertilization—A Pilot Study. Antioxidants. 2022; 11(9):1841. https://doi.org/10.3390/antiox11091841
Chicago/Turabian StyleKonstantinidou, Fani, Maria Cristina Budani, Guya Diletta Marconi, Francesca Gonnella, Annalina Sarra, Oriana Trubiani, Liborio Stuppia, Gian Mario Tiboni, and Valentina Gatta. 2022. "The Aftermath of Long-Term Cigarette Smoking on Telomere Length and Mitochondrial DNA Copy Number in Human Cumulus Cells Prior to In Vitro Fertilization—A Pilot Study" Antioxidants 11, no. 9: 1841. https://doi.org/10.3390/antiox11091841