
Chronic High-Altitude Hypoxia Alters Iron and Nitric Oxide Homeostasis in Fetal and Maternal Sheep Blood and Aorta
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Animals
2.2. Sample Collection and Preparation
2.3. Analytical Methods
2.4. Statistics
3. Results
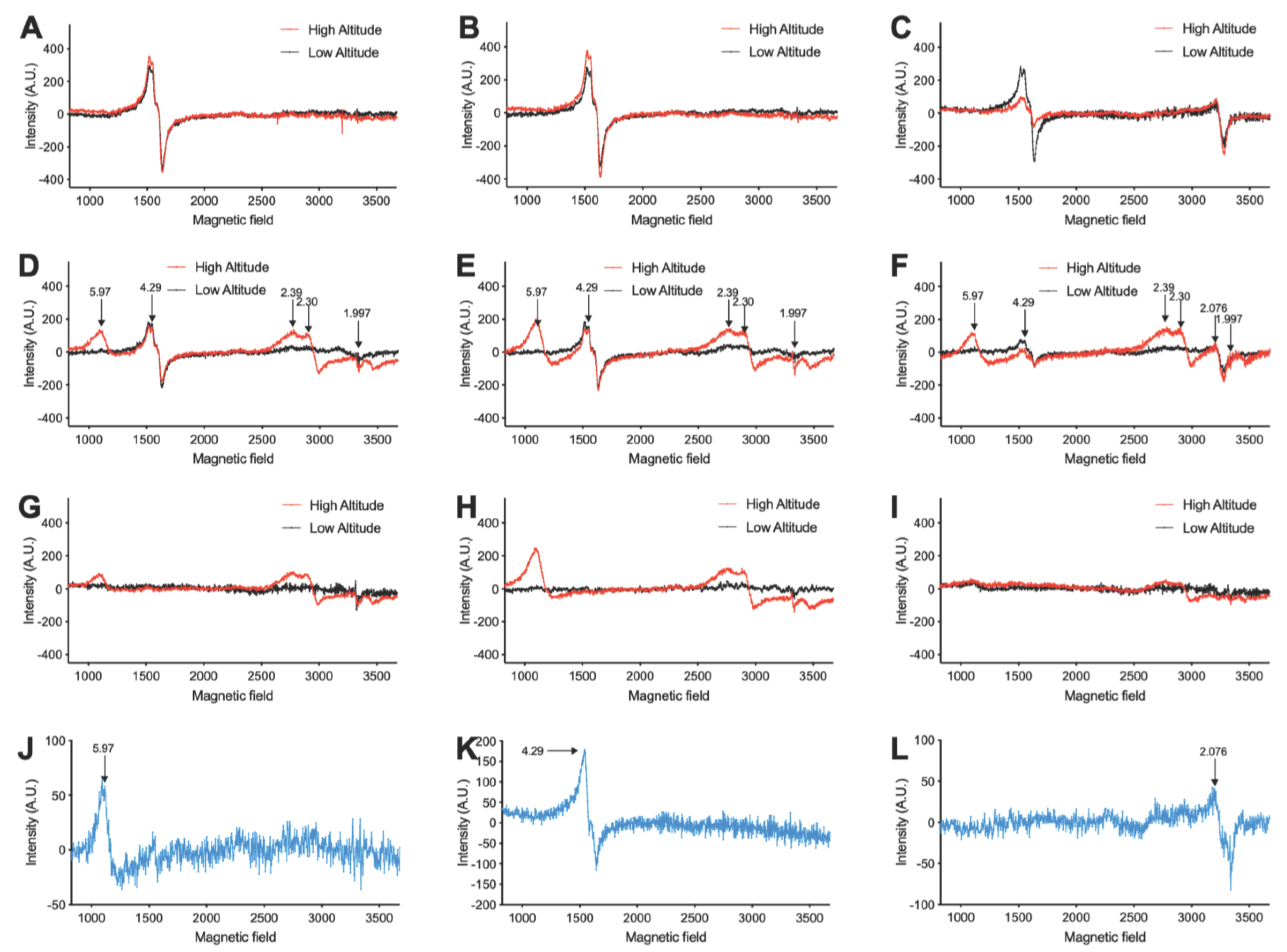
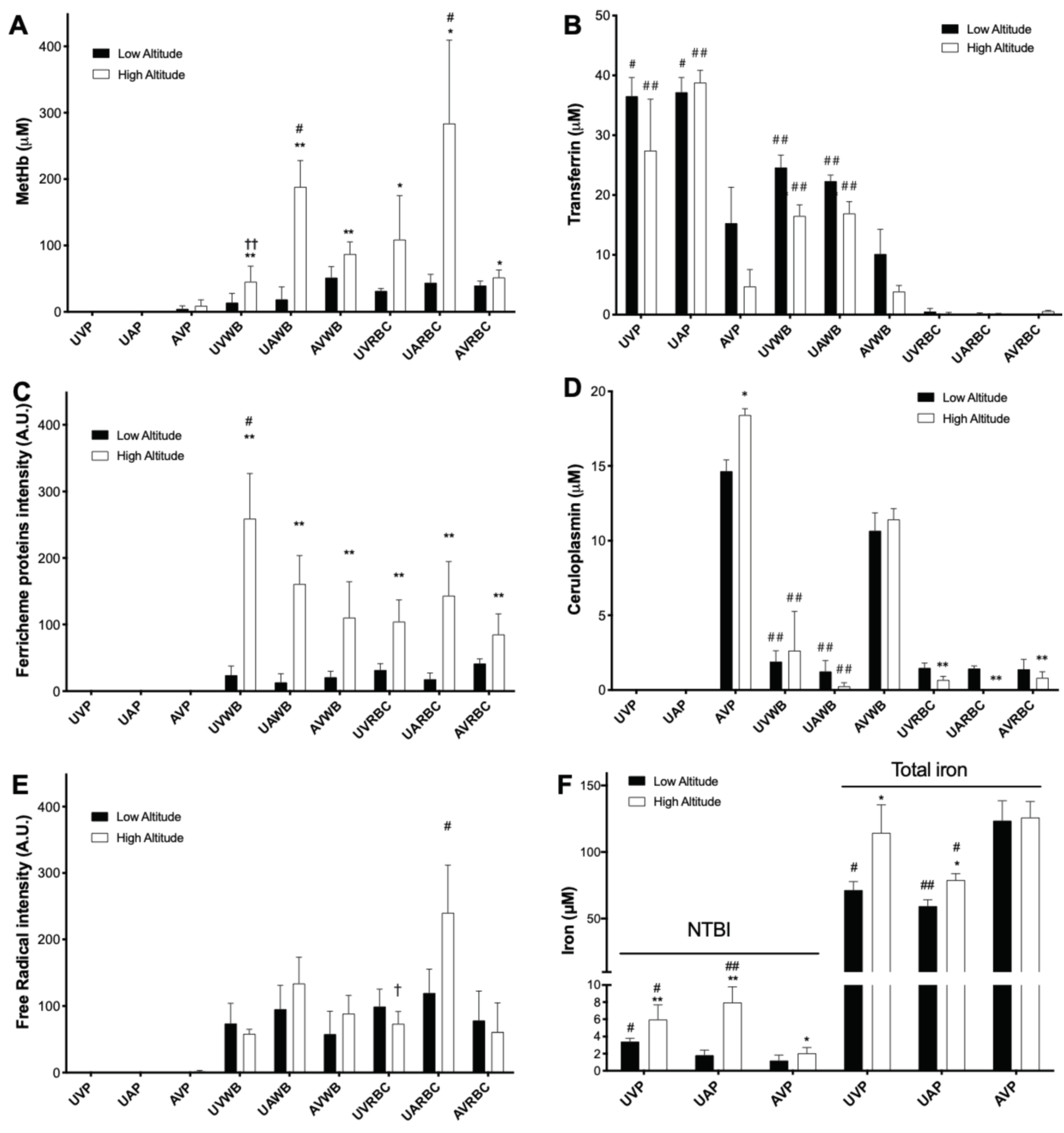
3.1. High Altitude-Induced Alterations of Metalloproteins, Free Radicals, and Iron Levels in Fetal and Maternal Sheep Blood
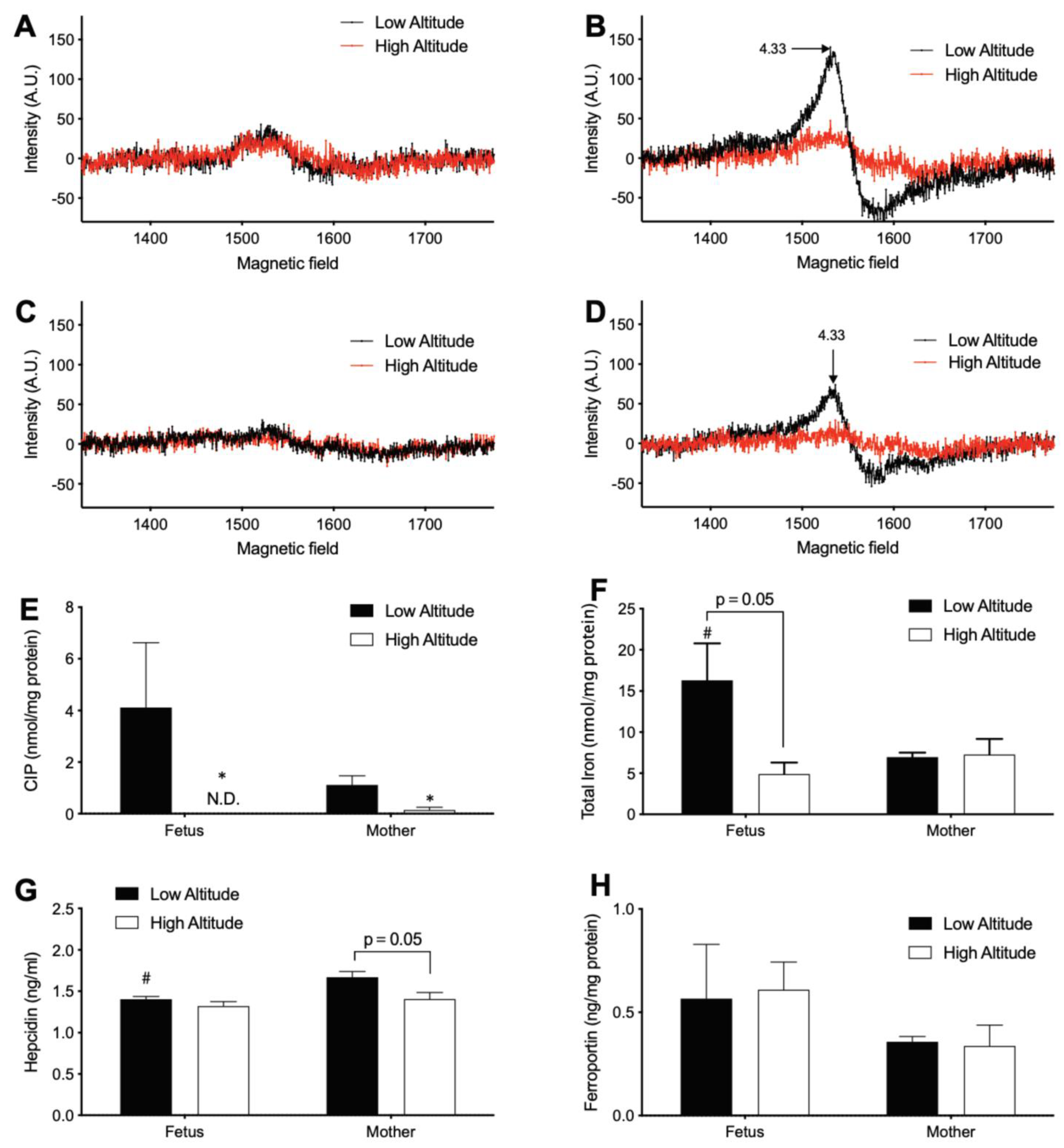
3.2. High Altitude Decreases Iron in Fetal and Maternal Aorta
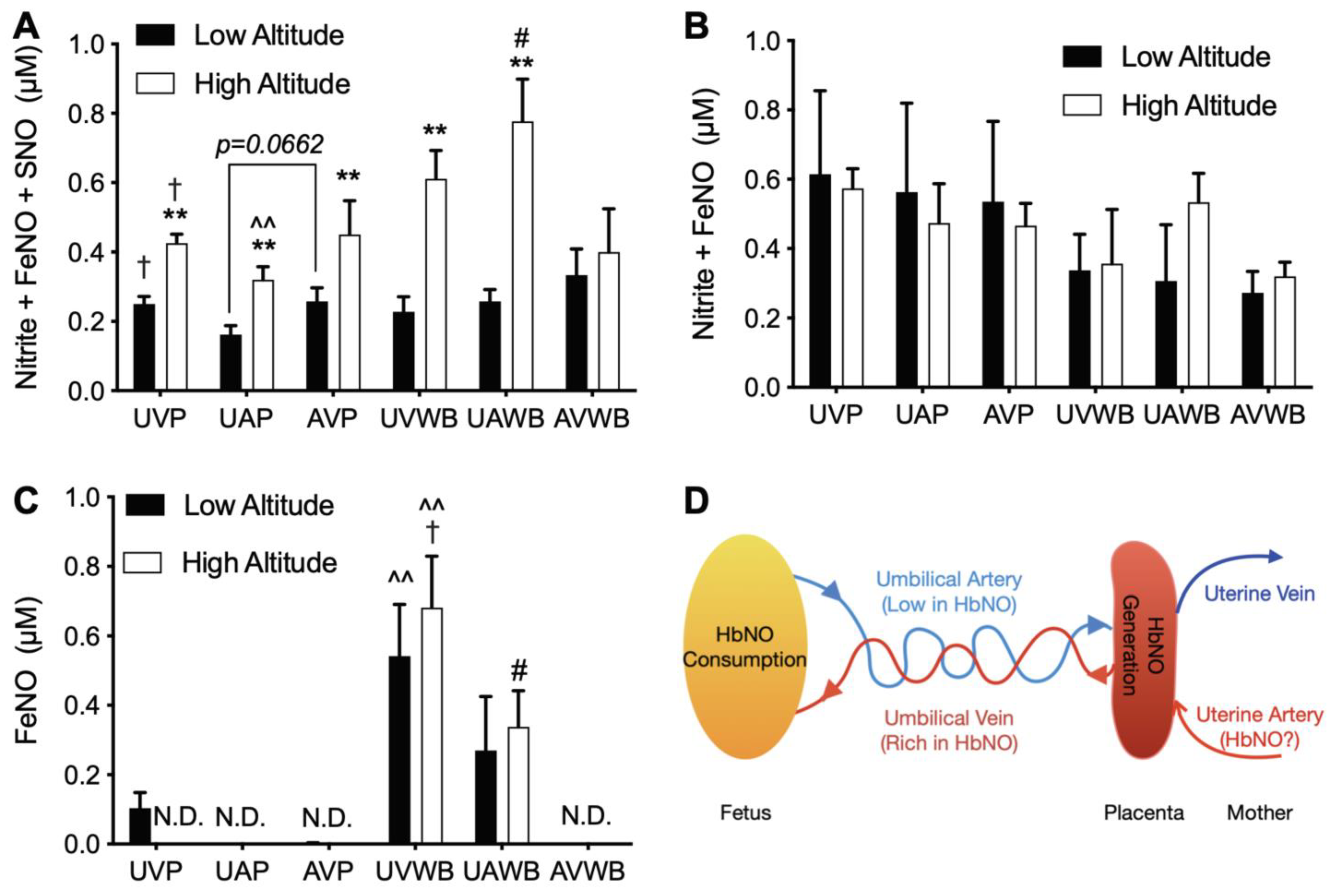
3.3. High Altitude Increases NOx in Fetal and Maternal Blood
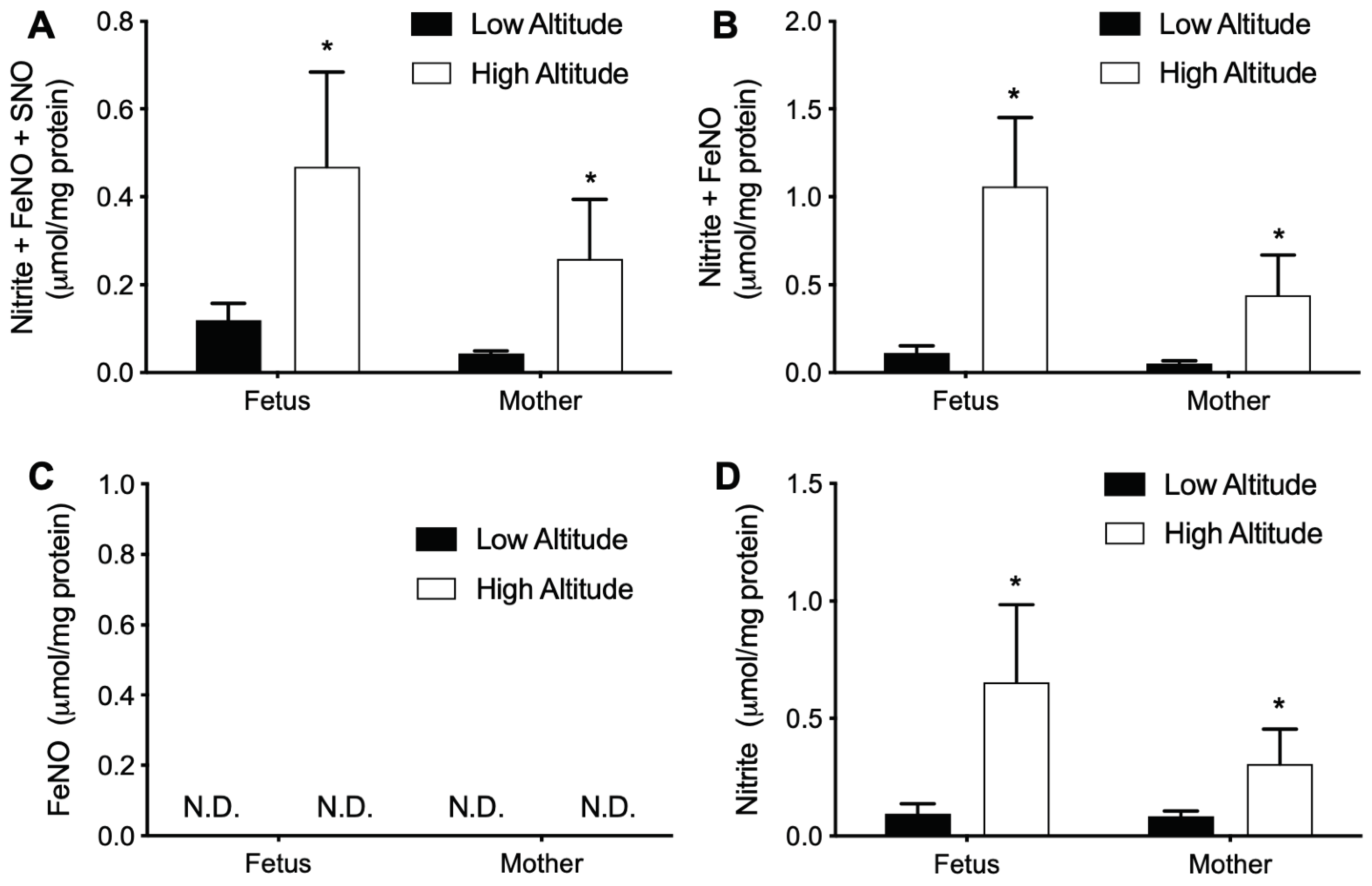
3.4. High Altitude Increases NOx in Fetal and Maternal Aorta
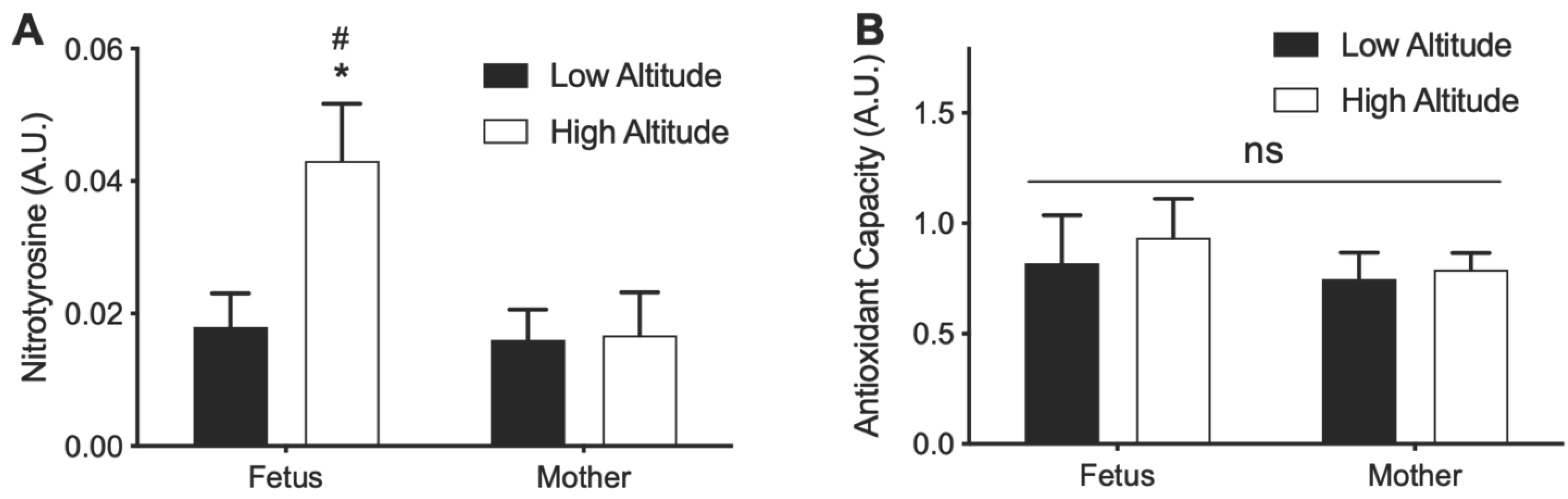
3.5. Effects of High Altitude on Nitrotyrosine and Antioxidant Capacity in Aorta
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Ducsay, C.A.; Goyal, R.; Pearce, W.J.; Wilson, S.; Hu, X.-Q.; Zhang, L. Gestational Hypoxia and Developmental Plasticity. Physiol. Rev. 2018, 98, 1241–1334. [Google Scholar] [CrossRef]
- Zamudio, S. High-Altitude Hypoxia and Preeclampsia. Front. Biosci. 2007, 12, 2967–2977. [Google Scholar] [CrossRef]
- Tremblay, J.C.; Ainslie, P.N. Global and Country-Level Estimates of Human Population at High Altitude. Proc. Natl. Acad. Sci. USA 2021, 118, e2102463118. [Google Scholar] [CrossRef]
- Galaris, D.; Barbouti, A.; Pantopoulos, K. Iron Homeostasis and Oxidative Stress: An Intimate Relationship. Biochim. Biophys. Acta Mol. Cell Res. 2019, 1866, 118535. [Google Scholar] [CrossRef]
- Kruszewski, M. Labile Iron Pool: The Main Determinant of Cellular Response to Oxidative Stress. Mutat. Res. 2003, 531, 81–92. [Google Scholar] [CrossRef]
- Epsztejn, S.; Glickstein, H.; Picard, V.; Slotki, I.N.; Breuer, W.; Beaumont, C.; Cabantchik, Z.I. H-Ferritin Subunit Overexpression in Erythroid Cells Reduces the Oxidative Stress Response and Induces Multidrug Resistance Properties. Blood 1999, 94, 3593–3603. [Google Scholar] [CrossRef]
- Chepelev, N.L.; Willmore, W.G. Regulation of Iron Pathways in Response to Hypoxia. Free Radic. Biol. Med. 2011, 50, 645–666. [Google Scholar] [CrossRef]
- Abu-Ouf, N.M.; Jan, M.M. The Impact of Maternal Iron Deficiency and Iron Deficiency Anemia on Child’s Health. Saudi Med. J. 2015, 36, 146–149. [Google Scholar] [CrossRef]
- Haig, D. Cooperation and Conflict in Human Pregnancy. Curr. Biol. 2019, 29, R455–R458. [Google Scholar] [CrossRef]
- Liu, T.; Mukosera, G.T.; Blood, A.B. The Role of Gasotransmitters in Neonatal Physiology. Nitric Oxide 2020, 95, 29–44. [Google Scholar] [CrossRef]
- Liu, T.; Schroeder, H.; Power, G.G.; Blood, A.B. A Physiologically Relevant Role for NO Stored in Vascular Smooth Muscle Cells: A Novel Theory of Vascular NO Signaling. Redox Biol. 2022, 53, 102327. [Google Scholar] [CrossRef] [PubMed]
- Thomas, D.D.; Liu, X.; Kantrow, S.P.; Lancaster, J.R. The Biological Lifetime of Nitric Oxide: Implications for the Perivascular Dynamics of NO and O2. Proc. Natl. Acad. Sci. USA 2001, 98, 355–360. [Google Scholar] [CrossRef] [PubMed]
- Lauer, T.; Preik, M.; Rassaf, T.; Strauer, B.E.; Deussen, A.; Feelisch, M.; Kelm, M. Plasma Nitrite rather than Nitrate Reflects Regional Endothelial Nitric Oxide Synthase Activity but Lacks Intrinsic Vasodilator Action. Proc. Natl. Acad. Sci. USA 2001, 98, 12814–12819. [Google Scholar] [CrossRef]
- Liu, T.; Zhang, M.; Terry, M.H.; Schroeder, H.; Wilson, S.M.; Power, G.G.; Li, Q.; Tipple, T.E.; Borchardt, D.; Blood, A.B. Hemodynamic Effects of Glutathione-Liganded Binuclear Dinitrosyl Iron Complex: Evidence for Nitroxyl Generation and Modulation by Plasma Albumin. Mol. Pharmacol. 2018, 93, 427–437. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Schroeder, H.J.; Wilson, S.M.; Terry, M.H.; Romero, M.; Longo, L.D.; Power, G.G.; Blood, A.B. Local and Systemic Vasodilatory Effects of Low Molecular Weight S-Nitrosothiols. Free Radic. Biol. Med. 2016, 91, 215–223. [Google Scholar] [CrossRef] [PubMed]
- Mukosera, G.T.; Clark, T.C.; Ngo, L.; Liu, T.; Schroeder, H.; Power, G.G.; Yellon, S.M.; Parast, M.M.; Blood, A.B. Nitric Oxide Metabolism in the Human Placenta during Aberrant Maternal Inflammation. J. Physiol. 2020, 598, 2223–2241. [Google Scholar] [CrossRef]
- Hickok, J.R.; Sahni, S.; Shen, H.; Arvind, A.; Antoniou, C.; Fung, L.W.M.; Thomas, D.D. Dinitrosyliron Complexes Are the Most Abundant Nitric Oxide-Derived Cellular Adduct: Biological Parameters of Assembly and Disappearance. Free Radic. Biol. Med. 2011, 51, 1558–1566. [Google Scholar] [CrossRef]
- Lok, H.C.; Sahni, S.; Jansson, P.J.; Kovacevic, Z.; Hawkins, C.L.; Richardson, D.R. A Nitric Oxide Storage and Transport System That Protects Activated Macrophages from Endogenous Nitric Oxide Cytotoxicity. J. Biol. Chem. 2016, 291, 27042–27061. [Google Scholar] [CrossRef]
- Thomas, D.D. Breathing New Life into Nitric Oxide Signaling: A Brief Overview of the Interplay between Oxygen and Nitric Oxide. Redox Biol. 2015, 5, 225–233. [Google Scholar] [CrossRef]
- Kim-Shapiro, D.B.; Gladwin, M.T. Mechanisms of Nitrite Bioactivation. Nitric Oxide 2014, 38, 58–68. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Xiao, D.; Bouslough, D. Long-term High-altitude Hypoxia Increases Plasma Nitrate Levels in Pregnant Ewes and Their Fetuses. Am. J. Obstet. Gynecol. 1998, 179, 1594–1598. [Google Scholar] [CrossRef]
- Erzurum, S.C.; Ghosh, S.; Janocha, A.J.; Xu, W.; Bauer, S.; Bryan, N.S.; Tejero, J.; Hemann, C.; Hille, R.; Stuehr, D.J.; et al. Higher Blood Flow and Circulating NO Products Offset High-Altitude Hypoxia among Tibetans. Proc. Natl. Acad. Sci. USA 2007, 104, 17593–17598. [Google Scholar] [CrossRef]
- Feelisch, M. Enhanced Nitric Oxide Production Is a Universal Response to Hypoxic Stress. Natl. Sci. Rev. 2018, 5, 532–533. [Google Scholar] [CrossRef]
- He, Y.; Qi, X.; Ouzhuluobu, O.; Liu, S.; Li, J.; Zhang, H.; Baimakangzhuo, B.; Bai, C.; Zheng, W.; Guo, Y.; et al. Blunted Nitric Oxide Regulation in Tibetans under High-Altitude Hypoxia. Natl. Sci. Rev. 2018, 5, 516–529. [Google Scholar] [CrossRef]
- Julian, C.G.; Galan, H.L.; Wilson, M.J.; DeSilva, W.; Cioffi-Ragan, D.; Schwartz, J.; Moore, L.G. Lower Uterine Artery Blood Flow and Higher Endothelin Relative to Nitric Oxide Metabolite Levels Are Associated with Reductions in Birth Weight at High Altitude. Am. J. Physiol.-Regul. Integr. Comp. Physiol. 2008, 295, R906–R915. [Google Scholar] [CrossRef] [PubMed]
- Blood, A.B.; Liu, T.; Mukosera, G.; Hanson, S.F.; Terry, M.H.; Schroeder, H.; Power, G.G. Evidence for Placental-Derived Iron-Nitrosyls in the Circulation of the Fetal Lamb and against a Role for Nitrite in Mediating the Cardiovascular Transition at Birth. Am. J. Physiol.-Regul. Integr. Comp. Physiol. 2020, 319, R401–R411. [Google Scholar] [CrossRef]
- Mukosera, G.T.; Liu, T.; Ishtiaq Ahmed, A.S.; Li, Q.; Sheng, M.H.-C.; Tipple, T.E.; Baylink, D.J.; Power, G.G.; Blood, A.B. Detection of Dinitrosyl Iron Complexes by Ozone-Based Chemiluminescence. Nitric Oxide 2018, 79, 57–67. [Google Scholar] [CrossRef]
- Goyal, R.; Longo, L.D. Metabolic Profiles in Ovine Carotid Arteries with Developmental Maturation and Long-Term Hypoxia. PLoS ONE 2015, 10, e0130739. [Google Scholar] [CrossRef]
- Moser, J.C.; Rawal, M.; Wagner, B.A.; Du, J.; Cullen, J.J.; Buettner, G.R. Pharmacological Ascorbate and Ionizing Radiation (IR) Increase Labile Iron in Pancreatic Cancer. Redox Biol. 2013, 2, 22–27. [Google Scholar] [CrossRef]
- Kleinhans, F.W.; Kline, S.C.; Dugan, W.M., Jr.; Williams, J.G. Comparison of Electron Paramagnetic Resonance and Atomic Absorption Serum Copper Measurements in Human Normal Control and Cancer Patients. Cancer Res. 1983, 43, 3447–3450. [Google Scholar]
- Burlaka, A.P.; Vovk, A.V.; Burlaka, A.A.; Gafurov, M.R.; Iskhakova, K.B.; Lukin, S.N. Rectal Cancer: Redox State of Venous Blood and Tissues of Blood Vessels from Electron Paramagnetic Resonance and Its Correlation with the Five-Year Survival. Biomed Res. Int. 2018, 2018, 4848652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yurtaeva, S.V.; Efimov, V.N.; Silkin, N.I.; Rodionov, A.A.; Burmistrov, M.V.; Panov, A.V.; Moroshek, A.A. Magnetic Resonance of Ferritin Crystalline Particles in Tumor Tissue. Appl. Magn. Reson. 2012, 42, 299–311. [Google Scholar] [CrossRef]
- Komosinska-Vassev, K.; Olczyk, P.; Kasperczyk, J.; Pilawa, B.; Krzyminiewski, R.; Dobosz, B.; Ramos, P.; Stojko, J.; Stojko, M.; Ivanova, D.; et al. EPR Spectroscopic Examination of Different Types of Paramagnetic Centers in the Blood in the Course of Burn Healing. Oxid. Med. Cell. Longev. 2019, 2019, 7506274. [Google Scholar] [CrossRef] [PubMed]
- Vrancken, K.; Schroeder, H.J.; Longo, L.D.; Power, G.G.; Blood, A.B. Role of Ceruloplasmin in Nitric Oxide Metabolism in Plasma of Humans and Sheep: A Comparison of Adults and Fetuses. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2013, 305, R1401–R1410. [Google Scholar] [CrossRef] [PubMed]
- Ramos, D.; Mar, D.; Ishida, M.; Vargas, R.; Gaite, M.; Montgomery, A.; Linder, M.C. Mechanism of Copper Uptake from Blood Plasma Ceruloplasmin by Mammalian Cells. PLoS ONE 2016, 11, e0149516. [Google Scholar] [CrossRef]
- Koonyosying, P.; Uthaipibull, C.; Fucharoen, S.; Koumoutsea, E.V.; Porter, J.B.; Srichairatanakool, S. Decrement in Cellular Iron and Reactive Oxygen Species, and Improvement of Insulin Secretion in a Pancreatic Cell Line Using Green Tea Extract. Pancreas 2019, 48, 636–643. [Google Scholar] [CrossRef]
- Krzyminiewski, R.; Kruczyński, Z.; Dobosz, B.; Zając, A.; Mackiewicz, A.; Leporowska, E.; Folwaczna, S. EPR Study of Iron Ion Complexes in Human Blood. Appl. Magn. Reson. 2011, 40, 321–330. [Google Scholar] [CrossRef]
- Svistunenko, D.A.; Patel, R.P.; Voloshchenko, S.V.; Wilson, M.T. The Globin-Based Free Radical of Ferryl Hemoglobin Is Detected in Normal Human Blood. J. Biol. Chem. 1997, 272, 7114–7121. [Google Scholar] [CrossRef]
- McDonald, E.A.; Gundogan, F.; Olveda, R.M.; Bartnikas, T.B.; Kurtis, J.D.; Friedman, J.F. Iron Transport across the Human Placenta Is Regulated by Hepcidin. Pediatric Res. 2020, 1–7. [Google Scholar] [CrossRef]
- Ward, D.M.; Kaplan, J. Ferroportin-Mediated Iron Transport: Expression and Regulation. Biochim. Biophys. Acta 2012, 1823, 1426–1433. [Google Scholar] [CrossRef]
- Grant, I.D.; Giussani, D.A.; Aiken, C.E. Fetal Growth and Spontaneous Preterm Birth in High-Altitude Pregnancy: A Systematic Review, Meta-Analysis, and Meta-Regression. Int. J. Gynaecol. Obstet. 2022, 157, 221–229. [Google Scholar] [CrossRef] [PubMed]
- Cao, C.; Fleming, M.D. The Placenta: The Forgotten Essential Organ of Iron Transport. Nutr. Rev. 2016, 74, 421–431. [Google Scholar] [CrossRef]
- Chandra, D.; Dhingra, B.; Seth, T.; Mishra, P.; Bansal, D.; Mahapatra, M.; Pati, H.P. Congenital Hypotransferrinemia, an Unusual Cause of Iron Deficiency Anemia: Report of Two Cases. Indian J. Hematol. Blood Transfus. 2017, 33, 402–404. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Ginzburg, Y.Z. Crosstalk between Iron Metabolism and Erythropoiesis. Adv. Hematol. 2010, 2010, 605435. [Google Scholar] [CrossRef] [PubMed]
- Kakhlon, O.; Breuer, W.; Munnich, A.; Cabantchik, Z.I. Iron Redistribution as a Therapeutic Strategy for Treating Diseases of Localized Iron Accumulation. Can. J. Physiol. Pharmacol. 2010, 88, 187–196. [Google Scholar] [CrossRef]
- Dev, S.; Babitt, J.L. Overview of Iron Metabolism in Health and Disease. Hemodial. Int. 2017, 21 (Suppl. S1), S6–S20. [Google Scholar] [CrossRef]
- Robach, P.; Cairo, G.; Gelfi, C.; Bernuzzi, F.; Pilegaard, H.; Viganò, A.; Santambrogio, P.; Cerretelli, P.; Calbet, J.A.L.; Moutereau, S.; et al. Strong Iron Demand during Hypoxia-Induced Erythropoiesis Is Associated with down-Regulation of Iron-Related Proteins and Myoglobin in Human Skeletal Muscle. Blood 2007, 109, 4724–4731. [Google Scholar] [CrossRef]
- Pena, J.P.; Tomimatsu, T.; Hatran, D.P.; McGill, L.L.; Longo, L.D. Cerebral Blood Flow and Oxygenation in Ovine Fetus: Responses to Superimposed Hypoxia at Both Low and High Altitude. J. Physiol. 2007, 578, 359–370. [Google Scholar] [CrossRef]
- Ganz, T. The Role of Hepcidin in Fetal Iron Homeostasis. Blood 2020, 136, 1474–1475. [Google Scholar] [CrossRef]
- Li, Q.; Li, C.; Mahtani, H.K.; Du, J.; Patel, A.R.; Lancaster, J.R., Jr. Nitrosothiol Formation and Protection against Fenton Chemistry by Nitric Oxide-Induced Dinitrosyliron Complex Formation from Anoxia-Initiated Cellular Chelatable Iron Increase. J. Biol. Chem. 2014, 289, 19917–19927. [Google Scholar] [CrossRef]
- Ganz, T.; Nemeth, E. Regulation of Iron Acquisition and Iron Distribution in Mammals. Biochim. Biophys. Acta 2006, 1763, 690–699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leung, P.S. Increased Duodenal Iron Uptake and Transfer in a Rat Model of Chronic Hypoxia Is Accompanied by Reduced Hepcidin Expression. Gut 2005, 54, 1391–1395. [Google Scholar] [CrossRef] [PubMed]
- Sahni, S.; Hickok, J.R.; Thomas, D.D. Nitric Oxide Reduces Oxidative Stress in Cancer Cells by Forming Dinitrosyliron Complexes. Nitric Oxide 2018, 76, 37–44. [Google Scholar] [CrossRef] [PubMed]
- Kakhlon, O.; Cabantchik, Z.I. The Labile Iron Pool: Characterization, Measurement, and Participation in Cellular processes. Free Radic. Biol. Med. 2002, 33, 1037–1046. [Google Scholar] [CrossRef]
- Herrera, E.A.; Ebensperger, G.; Hernández, I.; Sanhueza, E.M.; Llanos, A.J.; Reyes, R.V. The Role of Nitric Oxide Signaling in Pulmonary Circulation of High- and Low-Altitude Newborn Sheep under Basal and Acute Hypoxic Conditions. Nitric Oxide 2019, 89, 71–80. [Google Scholar] [CrossRef] [PubMed]
- Reyes, R.V.; Díaz, M.; Ebensperger, G.; Herrera, E.A.; Quezada, S.A.; Hernandez, I.; Sanhueza, E.M.; Parer, J.T.; Giussani, D.A.; Llanos, A.J. The Role of Nitric Oxide in the Cardiopulmonary Response to Hypoxia in Highland and Lowland Newborn Llamas. J. Physiol. 2018, 596, 5907–5923. [Google Scholar] [CrossRef]
- Xue, Q.; Ducsay, C.A.; Longo, L.D.; Zhang, L. Effect of Long-Term High-Altitude Hypoxia on Fetal Pulmonary Vascular Contractility. J. Appl. Physiol. 2008, 104, 1786–1792. [Google Scholar] [CrossRef]
- Thompson, L.P.; Dong, Y. Chronic Hypoxia Decreases Endothelial Nitric Oxide Synthase Protein Expression in Fetal Guinea Pig Hearts. J. Soc. Gynecol. Investig. 2005, 12, 388–395. [Google Scholar] [CrossRef]
- Aguan, K.; Murotsuki, J.; Gagnon, R.; Thompson, L.P.; Weiner, C.P. Effect of Chronic Hypoxemia on the Regulation of Nitric-Oxide Synthase in the Fetal Sheep Brain. Brain Res. Dev. Brain Res. 1998, 111, 271–277. [Google Scholar] [CrossRef]
- Cicinelli, E.; Ignarro, L.J.; Schonauer, L.M.; Matteo, M.G.; Galantino, P.; Falco, N. Different Plasma Levels of Nitric Oxide in Arterial and Venous Blood. Clin. Physiol. 1999, 19, 440–442. [Google Scholar] [CrossRef]
- Gladwin, M.T.; Shelhamer, J.H.; Schechter, A.N.; Pease-Fye, M.E.; Waclawiw, M.A.; Panza, J.A.; Ognibene, F.P.; Cannon, R.O. Role of Circulating Nitrite and S -Nitrosohemoglobin in the Regulation of Regional Blood Flow in Humans. Proc. Natl. Acad. Sci. USA 2000, 97, 11482–11487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morgan, M.A.M.; Frasier, L.M.; Stewart, J.C.; Mack, C.M.; Gough, M.S.; Graves, B.T.; Apostolakos, M.J.; Doolin, K.P.; Darling, D.C.; Frampton, M.W.; et al. Artery-to-Vein Differences in Nitric Oxide Metabolites Are Diminished in Sepsis. Crit. Care Med. 2010, 38, 1069–1077. [Google Scholar] [CrossRef] [PubMed]
- Blood, A.B.; Tiso, M.; Verma, S.T.; Lo, J.; Joshi, M.S.; Azarov, I.; Longo, L.D.; Gladwin, M.T.; Kim-Shapiro, D.B.; Power, G.G. Increased Nitrite Reductase Activity of Fetal versus Adult Ovine Hemoglobin. Am. J. Physiol. Heart Circ. Physiol. 2009, 296, H237–H246. [Google Scholar] [CrossRef]
- Doctor, A.; Stamler, J.S. Nitric Oxide Transport in Blood: A Third Gas in the Respiratory Cycle. Compr. Physiol. 2011, 541–568. [Google Scholar]
- Gladwin, M.T.; Schechter, A.N.; Kim-Shapiro, D.B.; Patel, R.P.; Hogg, N.; Shiva, S.; Cannon, R.O.; Kelm, M.; Wink, D.A.; Espey, M.G.; et al. The Emerging Biology of the Nitrite Anion. Nat. Chem. Biol. 2005, 1, 308–314. [Google Scholar] [CrossRef]
- Mukosera, G.T.; Principe, P.; Mata-Greenwood, E.; Liu, T.; Schroeder, H.; Parast, M.; Blood, A.B. Iron Nitrosyl Complexes Are Formed from Nitrite in the Human Placenta. J. Biol. Chem. 2022, 298, 102078. [Google Scholar] [CrossRef] [PubMed]
- Fazio, F.; Arfuso, F.; Giudice, E.; Giannetto, C.; Piccione, G. Physiological differences between twin and single-born lambs and kids during the first month of life. Arch. Anim. Breed. 2016, 59, 201–207. [Google Scholar] [CrossRef]
- Arfuso, F.; Fazio, F.; Panzera, M.; Giannetto, C.; Di Pietro, S.; Giudice, E.; Piccione, G. Lipid and Lipoprotein Profile Changes in Newborn Calves in Response to the Perinatal Period. Acta Vet.-Beograd. 2017, 67, 25–32. [Google Scholar] [CrossRef]
- Piccione, G.; Rizzo, M.; Arfuso, F.; Giannetto, C.; Di Pietro, S.; Bazzano, M.; Quartuccio, M. Leukocyte modifications during the first month after foaling in mares and their newborn foals. Pol. J. Vet. Sci. 2015, 18, 621–625. [Google Scholar] [CrossRef]
- Arfuso, F.; Giudice, E.; Di Pietro, S.; Quartuccio, M.; Giannetto, C.; Piccione, G. The Dynamics of Serum Lipid and Lipoprotein Profiles in Growing Foals. J. Equine Vet. Sci. 2016, 40, 1–5. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, T.; Zhang, M.; Mourkus, A.; Schroeder, H.; Zhang, L.; Power, G.G.; Blood, A.B. Chronic High-Altitude Hypoxia Alters Iron and Nitric Oxide Homeostasis in Fetal and Maternal Sheep Blood and Aorta. Antioxidants 2022, 11, 1821. https://doi.org/10.3390/antiox11091821
Liu T, Zhang M, Mourkus A, Schroeder H, Zhang L, Power GG, Blood AB. Chronic High-Altitude Hypoxia Alters Iron and Nitric Oxide Homeostasis in Fetal and Maternal Sheep Blood and Aorta. Antioxidants. 2022; 11(9):1821. https://doi.org/10.3390/antiox11091821
Chicago/Turabian StyleLiu, Taiming, Meijuan Zhang, Avoumia Mourkus, Hobe Schroeder, Lubo Zhang, Gordon G. Power, and Arlin B. Blood. 2022. "Chronic High-Altitude Hypoxia Alters Iron and Nitric Oxide Homeostasis in Fetal and Maternal Sheep Blood and Aorta" Antioxidants 11, no. 9: 1821. https://doi.org/10.3390/antiox11091821