
Oxidative and Inflammatory Imbalance in Placenta and Kidney of sFlt1-Induced Early-Onset Preeclampsia Rat Model

 ,
,  ,
,  ,
,  , and
, and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Design
2.2. Preeclampsia Induction
2.3. Animal Characteristics
2.4. Sample Harvesting and Tissue Homogenization
2.5. Determination of sFlt1, PlGF and NO Levels
2.6. Histological Staining and Superoxide Anion Levels
2.7. NADPH Oxidase Activity
2.8. Antioxidant Enzyme Activities
2.9. Real-Time PCR
2.10. Western Blotting
2.11. Statistical Analyses
3. Results
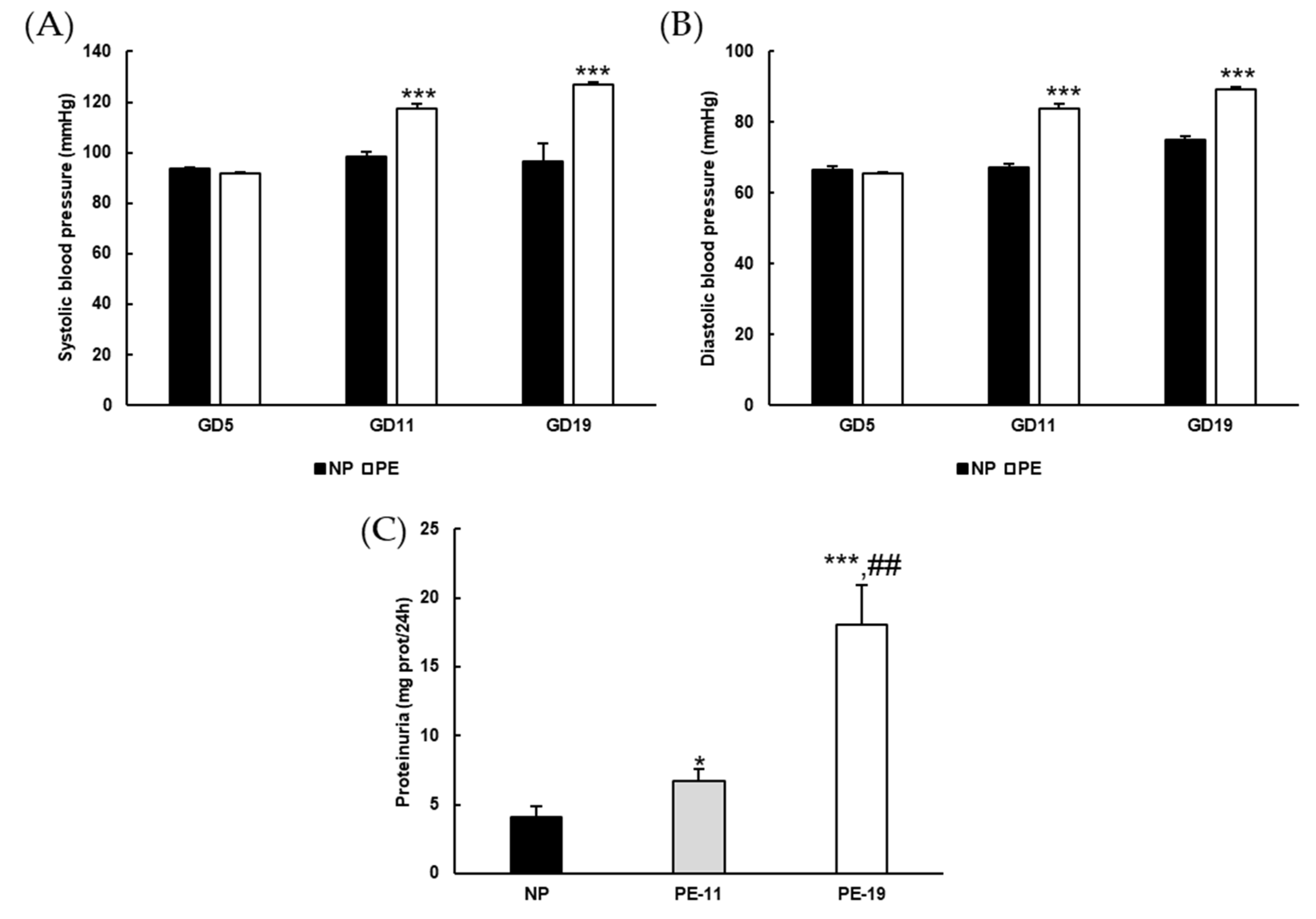
3.1. General Characteristics of the sFlt1-Dependent Animal Model of Preeclampsia
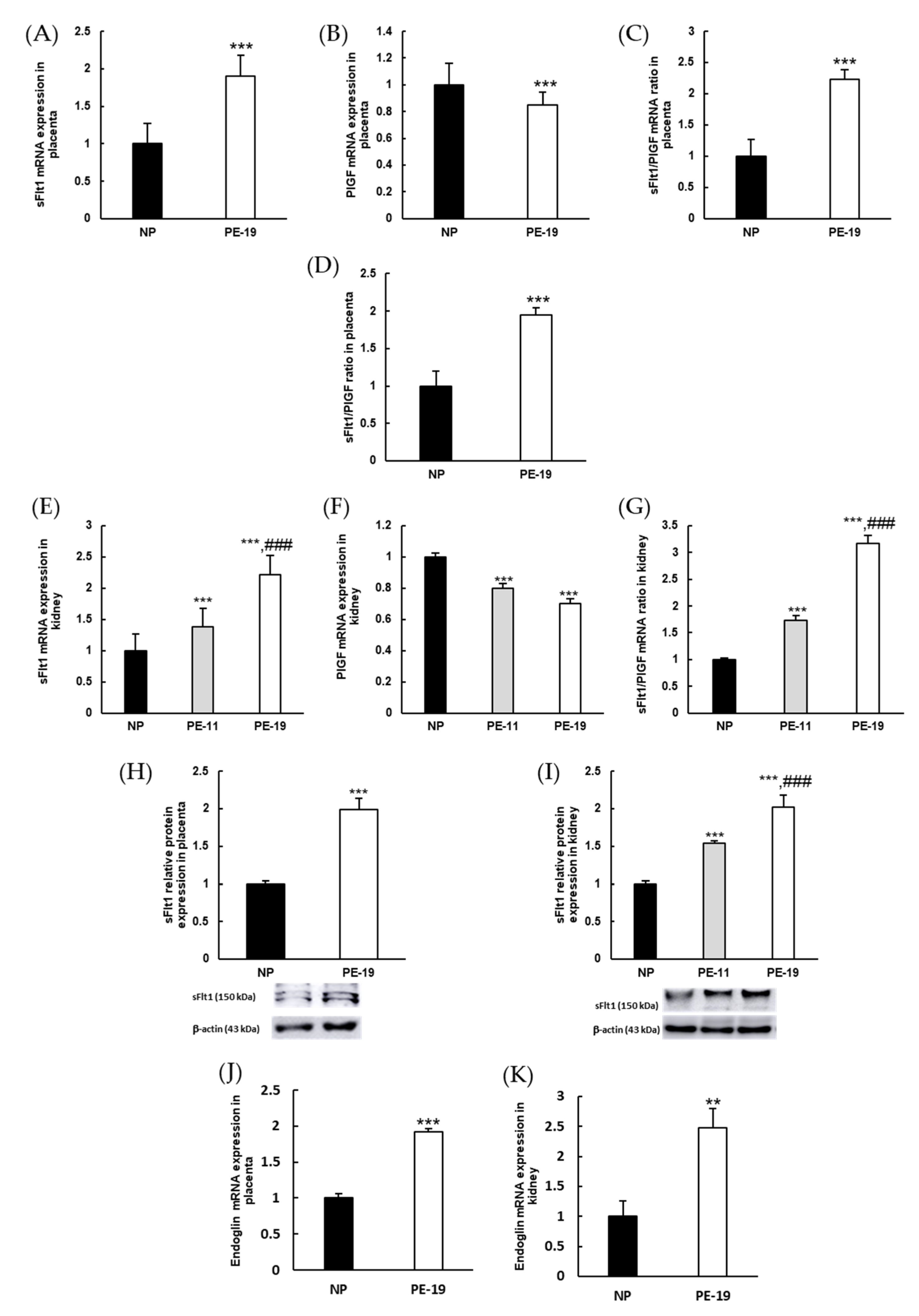
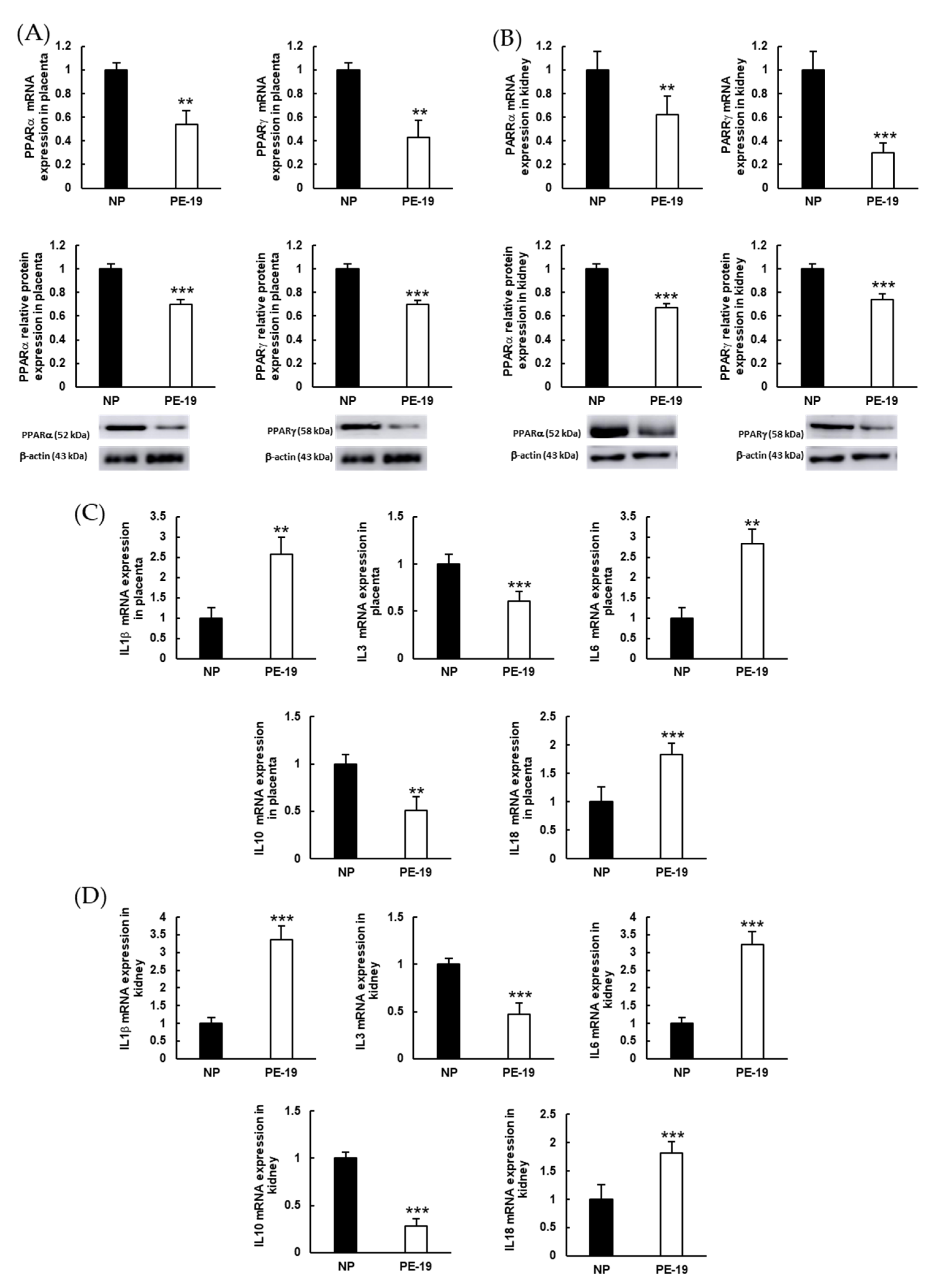
3.2. Molecular Characterization of the sFlt1-Based Preeclampsia Animal Model
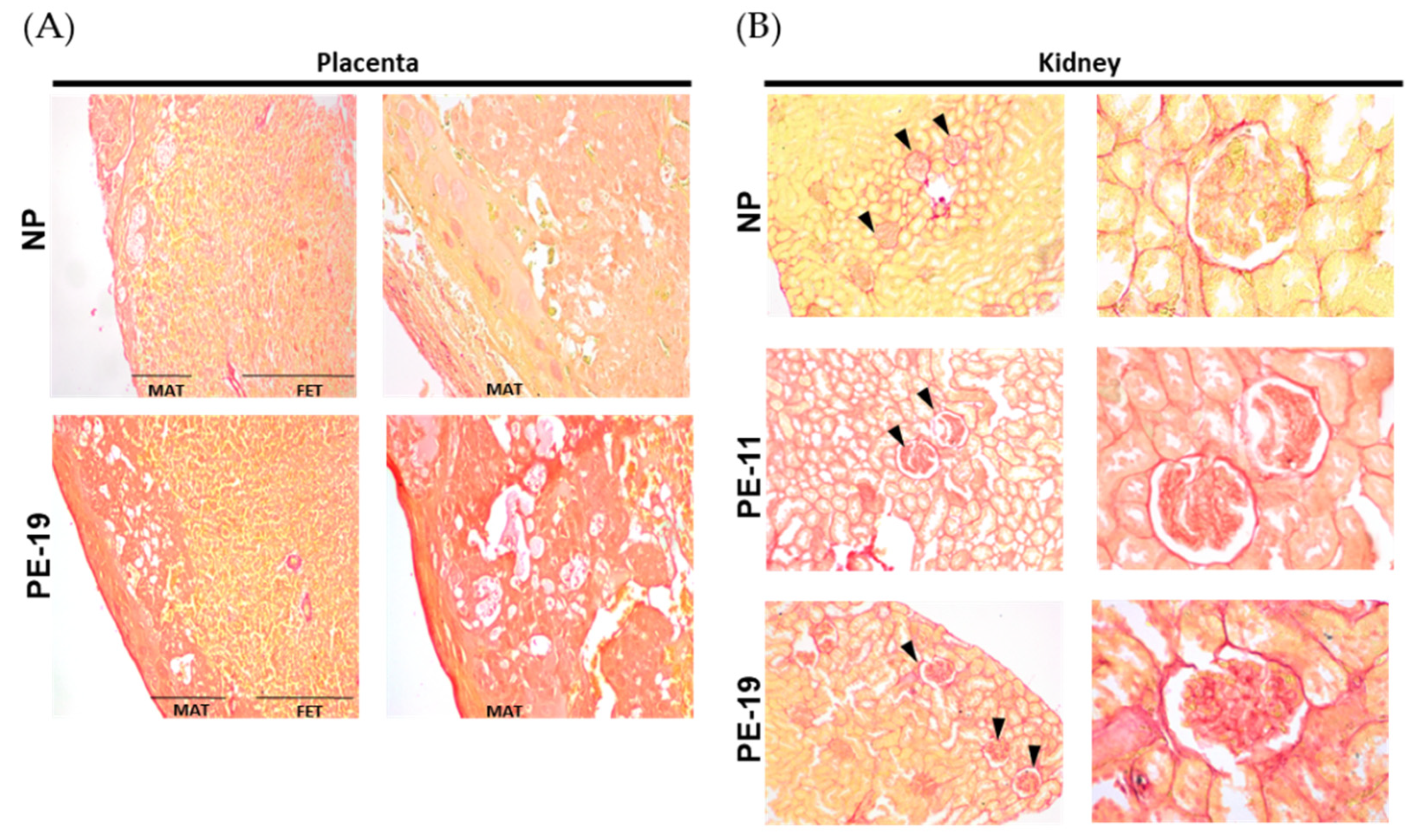
3.3. Collagen Deposition in the Placenta and Kidney
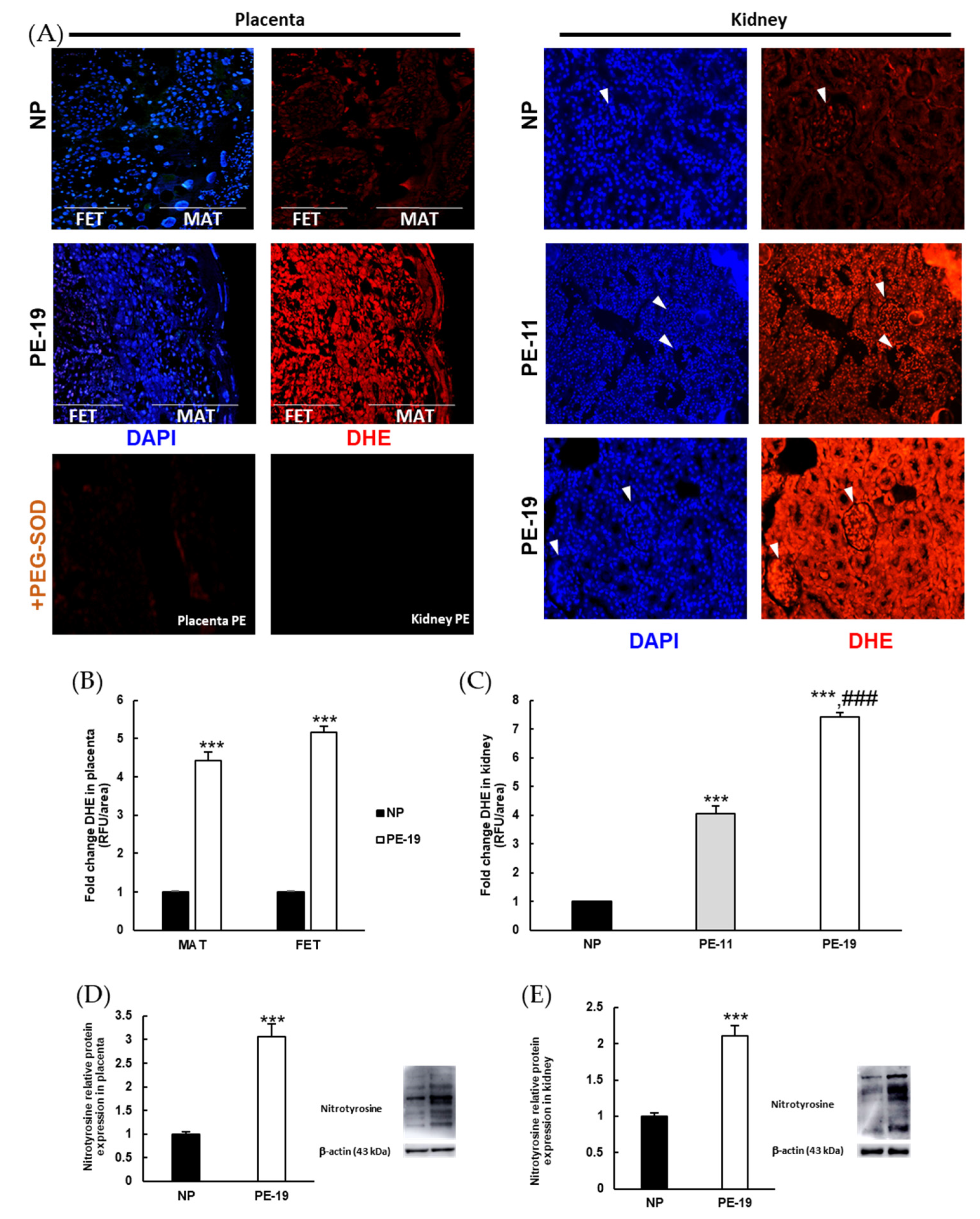
3.4. Preeclampsia-Induced O2●− Overproduction and Nitrosylation of Proteins in Placental and Renal Tissues
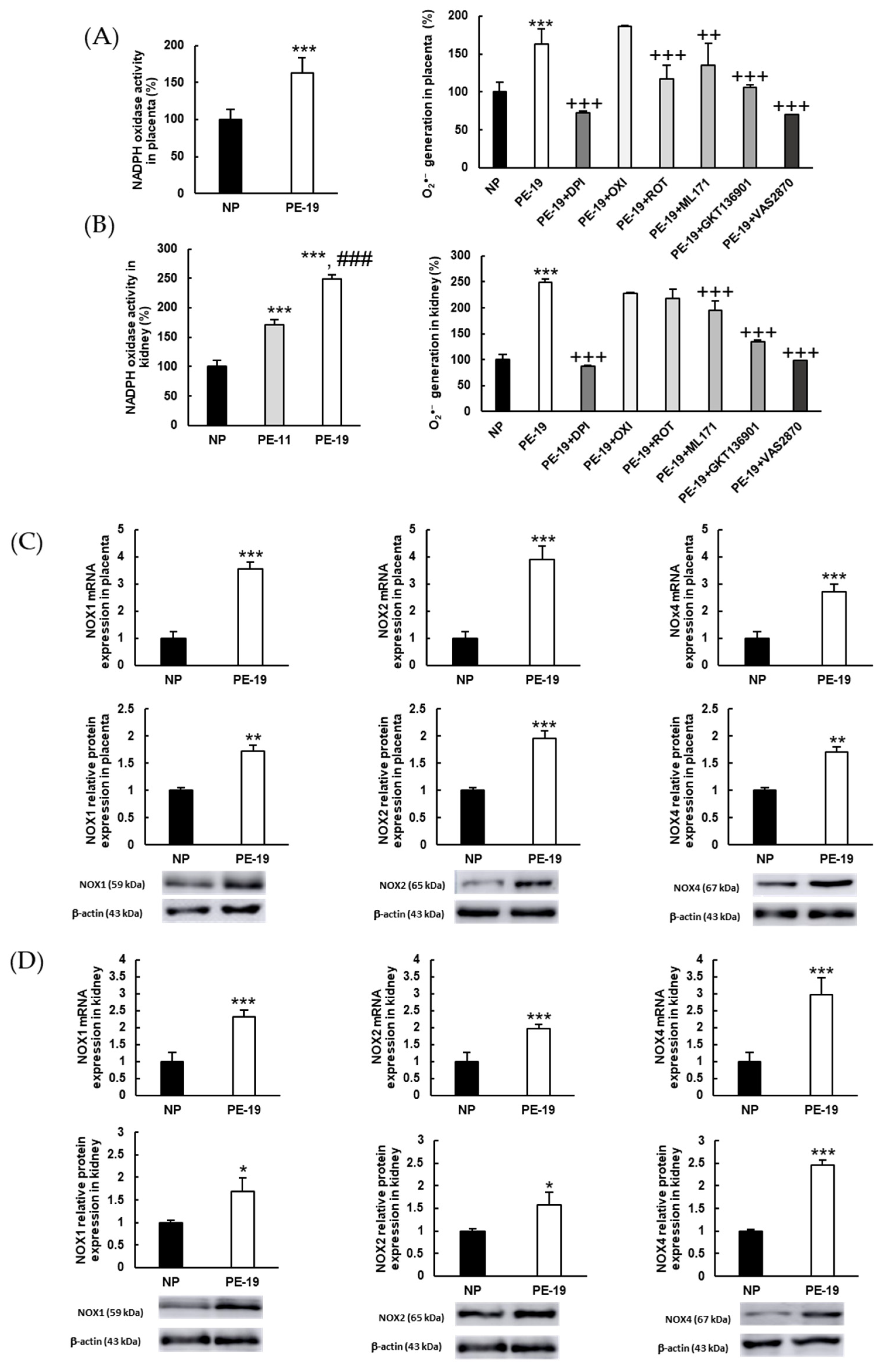
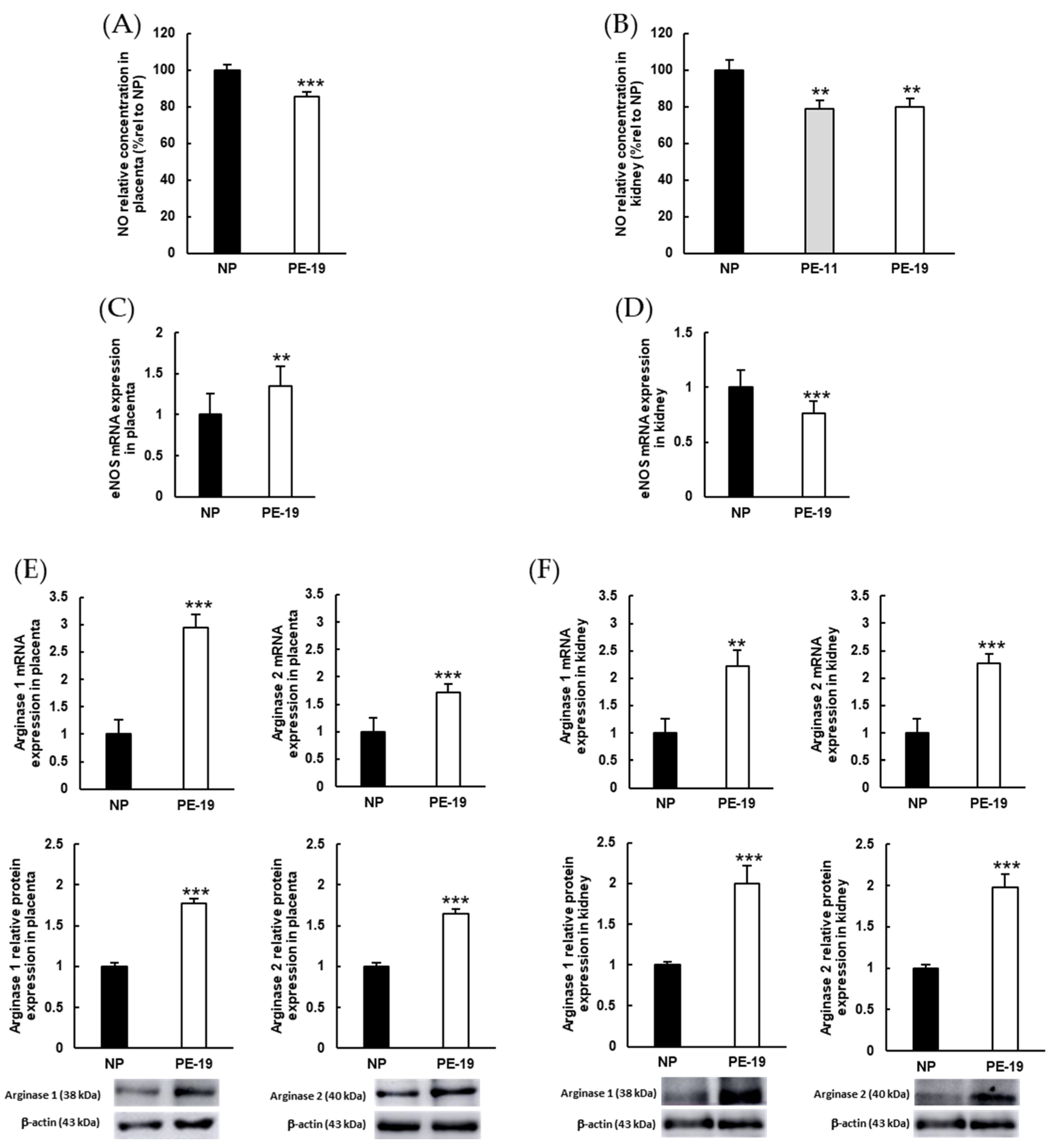
3.5. NOX Activation and Alterations in NO Metabolism Are Involved in the Oxidative Imbalance Present in the sFlt1-Dependent Model of Preeclampsia
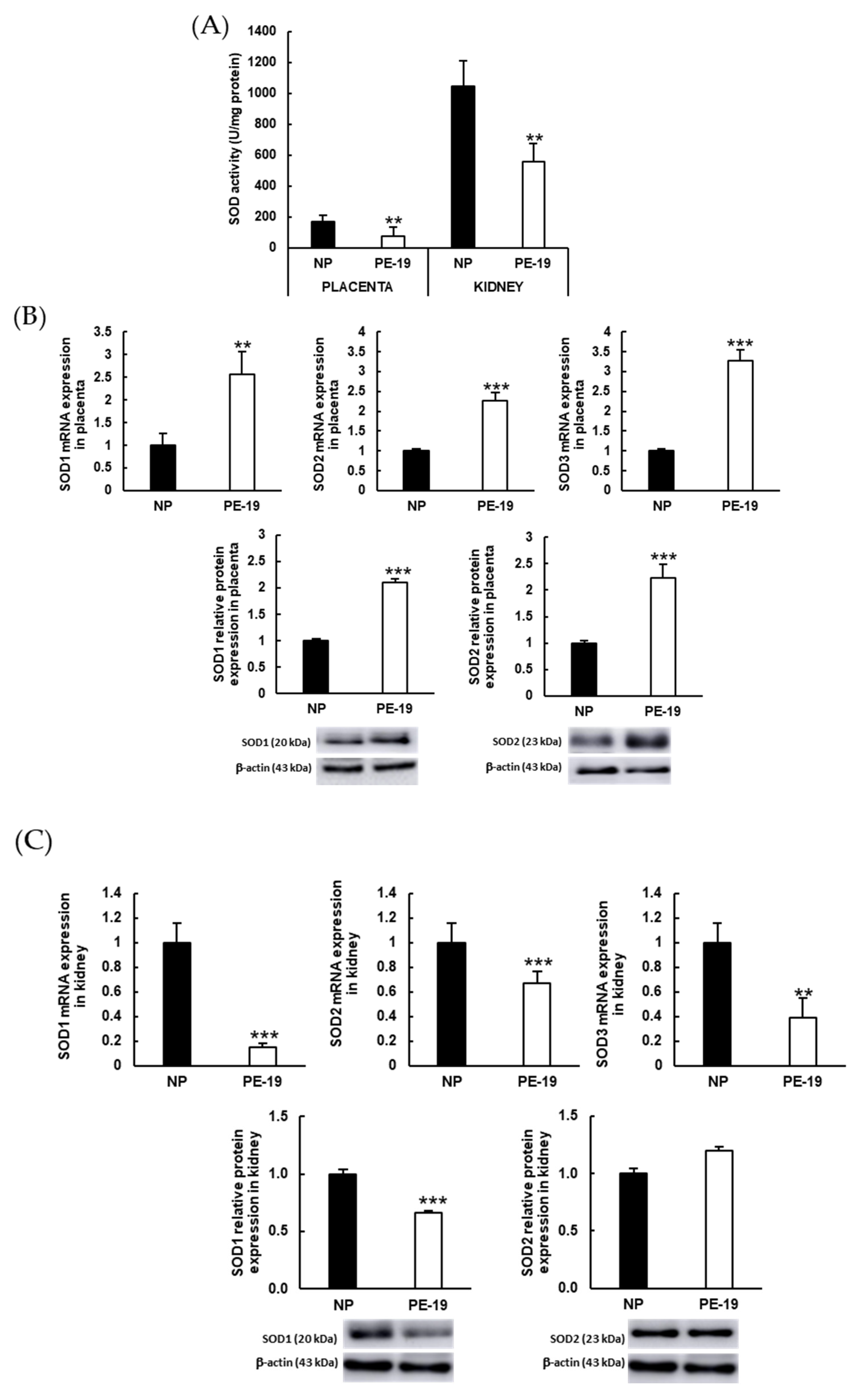
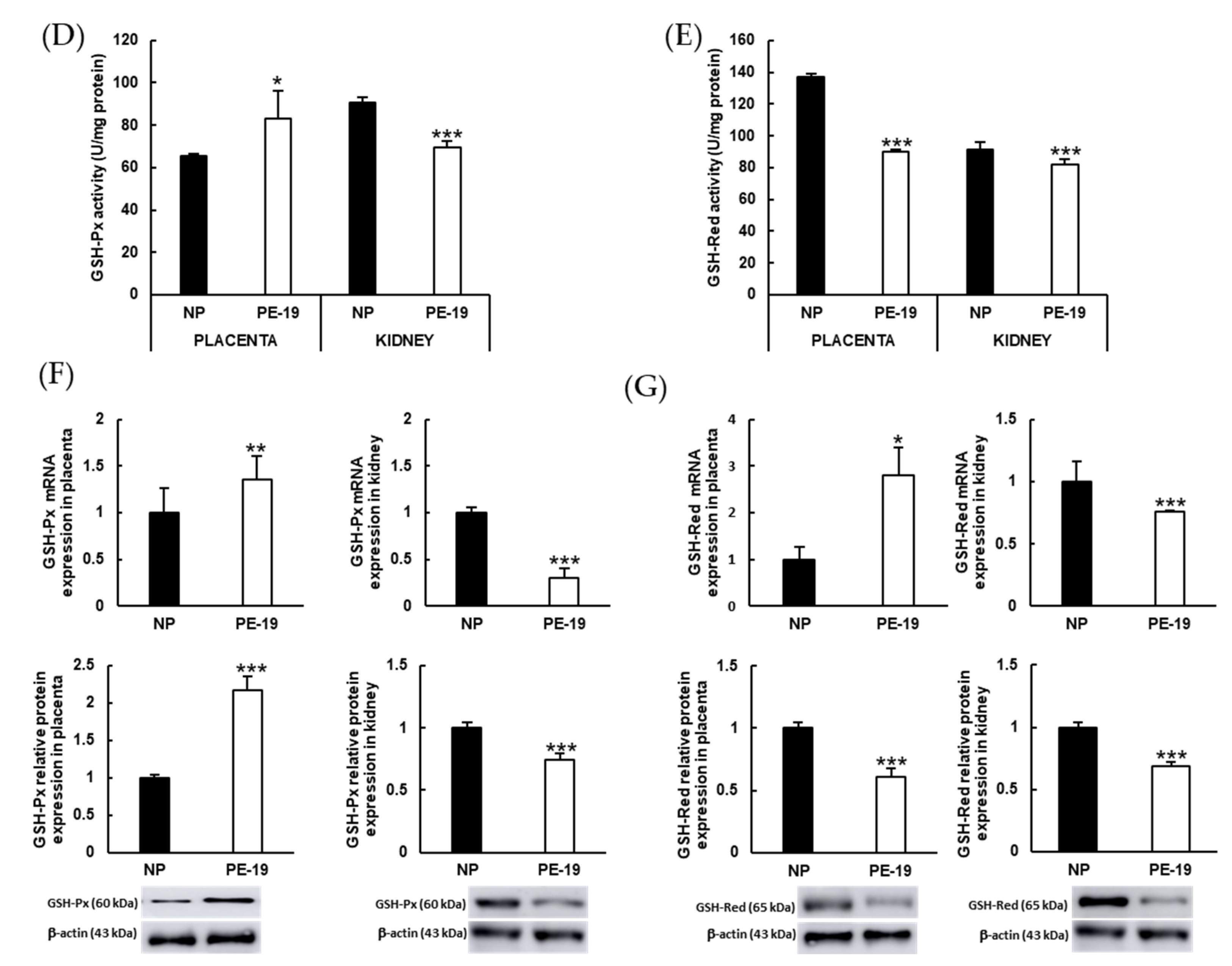
3.6. Preeclampsia Modifies Enzymatic Antioxidant Defense Systems
3.7. Inflammation Biomarkers in the sFlt1-dependent Model of Preeclampsia
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tamás, P.; Kovács, K.; Várnagy, Á.; Farkas, B.; Wami, G.A.; Bódis, J. Preeclampsia subtypes: Clinical aspects regarding pathogenesis, signs, and management with special attention to diuretic administration. Eur. J. Obstet. Gynecol. Reprod. Biol. 2022, 274, 175–181. [Google Scholar] [CrossRef] [PubMed]
- Brown, M.A.; Magee, L.A.; Kenny, L.C.; Karumanchi, S.A.; McCarthy, F.P.; Saito, S.; Hall, D.R.; Warren, C.E.; Adoyi, G.; Ishaku, S. Hypertensive Disorders of Pregnancy. Hypertension 2018, 72, 24–43. [Google Scholar] [CrossRef] [PubMed]
- Helmo, F.R.; Lopes, A.M.M.; Carneiro, A.C.D.M.; Campos, C.G.; Silva, P.B.; dos Reis Monteiro, M.L.G.; Rocha, L.P.; dos Reis, M.A.; Etchebehere, R.M.; Machado, J.R.; et al. Angiogenic and antiangiogenic factors in preeclampsia. Pathol.-Res. Pract. 2018, 214, 7–14. [Google Scholar] [CrossRef] [PubMed]
- Mate, A.; Reyes-Goya, C.; Santana-Garrido, Á.; Vázquez, C.M. Lifestyle, Maternal Nutrition and Healthy Pregnancy. Curr. Vasc. Pharmacol. 2020, 19, 132–140. [Google Scholar] [CrossRef] [PubMed]
- Mate, A.; Reyes-Goya, C.; Santana-Garrido, Á.; Sobrevia, L.; Vázquez, C.M. Impact of maternal nutrition in viral infections during pregnancy. Biochim. Et Biophys. Acta BBA-Mol. Basis Dis. 2021, 1867, 166231. [Google Scholar] [CrossRef] [PubMed]
- Calimag-Loyola, A.P.P.; Lerma, E.V. Renal complications during pregnancy: In the hypertension spectrum. Dis.-A-Mon. 2019, 65, 25–44. [Google Scholar] [CrossRef] [PubMed]
- Alese, M.O.; Moodley, J.; Naicker, T. Preeclampsia and HELLP syndrome, the role of the liver. J. Matern. Neonatal Med. 2021, 34, 117–123. [Google Scholar] [CrossRef]
- Facca, T.A.; Kirsztajn, G.M.; Sass, N. Pré-eclâmpsia (indicador de doença renal crônica): Da gênese aos riscos futuros. J. Bras. Nefrol. 2012, 34, 87–93. [Google Scholar] [CrossRef]
- Auger, N.; Fraser, W.D.; Schnitzer, M.; Leduc, L.; Healy-Profitós, J.; Paradis, G. Recurrent pre-eclampsia and subsequent cardiovascular risk. Heart 2017, 103, 235–243. [Google Scholar] [CrossRef]
- Hammer, E.S.; Cipolla, M.J. Cerebrovascular Dysfunction in Preeclamptic Pregnancies. Curr. Hypertens. Rep. 2015, 17, 64. [Google Scholar] [CrossRef]
- Gatford, K.L.; Andraweera, P.H.; Roberts, C.T.; Care, A.S. Animal Models of Preeclampsia: Causes, Consequences, and Interventions. Hypertension 2020, 75, 1363–1381. [Google Scholar] [CrossRef]
- Staff, A.C. The two-stage placental model of preeclampsia: An update. J. Reprod. Immunol. 2019, 134, 1–10. [Google Scholar] [CrossRef]
- Chiarello, D.I.; Abad, C.; Rojas, D.; Toledo, F.; Vázquez, C.M.; Mate, A.; Sobrevia, L.; Marín, R. Oxidative stress: Normal pregnancy versus preeclampsia. Biochim. Biophys. Acta-Mol. Basis Dis. 2020, 1866, 165354. [Google Scholar] [CrossRef]
- Matsubara, K.; Higaki, T.; Matsubara, Y.; Nawa, A. Nitric Oxide and Reactive Oxygen Species in the Pathogenesis of Preeclampsia. Int. J. Mol. Sci. 2015, 16, 4600–4614. [Google Scholar] [CrossRef]
- Salsoso, R.; Guzmán-Gutiérrez, E.; Sáez, T.; Bugueño, K.; Ramírez, M.A.; Farías, M.; Pardo, F.; Leiva, A.; Sanhueza, C.; Mate, A.; et al. Insulin restores l-arginine transport requiring adenosine receptors activation in umbilical vein endothelium from late-onset preeclampsia. Placenta 2015, 36, 287–296. [Google Scholar] [CrossRef]
- Raijmakers, M.T.M.; Peters, W.H.M.; Steegers, E.A.P.; Poston, L. NAD(P)H oxidase associated superoxide production in human placenta from normotensive and pre-eclamptic women. Placenta 2004, 25, 85–89. [Google Scholar] [CrossRef]
- Hernandez, I.; Fournier, T.; Chissey, A.; Therond, P.; Slama, A.; Beaudeux, J.L.; Zerrad-Saadi, A. NADPH oxidase is the major source of placental superoxide in early pregnancy: Association with MAPK pathway activation. Sci. Rep. 2019, 9, 13962. [Google Scholar] [CrossRef]
- Nguyen, M.V.C.; Zhang, L.; Lhomme, S.; Mouz, N.; Lenormand, J.L.; Lardy, B.; Morel, F. Recombinant Nox4 cytosolic domain produced by a cell or cell-free base systems exhibits constitutive diaphorase activity. Biochem. Biophys. Res. Commun. 2012, 419, 453–458. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Santana-Garrido, Á.; Reyes-Goya, C.; Fernández-Bobadilla, C.; Blanca, A.J.; André, H.; Mate, A.; Vázquez, C.M. NADPH oxidase-induced oxidative stress in the eyes of hypertensive rats. Mol. Vis. 2021, 27, 161–178. [Google Scholar]
- Santana-Garrido, Á.; Reyes-Goya, C.; Pérez-Camino, M.C.; André, H.; Mate, A.; Vázquez, C.M. Retinoprotective Effect of Wild Olive (Acebuche) Oil-Enriched Diet against Ocular Oxidative Stress Induced by Arterial Hypertension. Antioxidants 2020, 9, 885. [Google Scholar] [CrossRef] [PubMed]
- Zambrano, S.; Blanca, A.J.; Ruiz-Armenta, M.V.; Miguel-Carrasco, J.L.; Arévalo, M.; Vázquez, M.J.; Mate, A.; Vázquez, C.M. L-Carnitine protects against arterial hypertension-related cardiac fibrosis through modulation of PPAR-γ expression. Biochem. Pharmacol. 2013, 85, 937–944. [Google Scholar] [CrossRef] [PubMed]
- Peshavariya, H.M.; Dusting, G.J.; Selemidis, S. Analysis of dihydroethidium fluorescence for the detection of intracellular and extracellular superoxide produced by NADPH oxidase. Free Radic. Res. 2007, 41, 699–712. [Google Scholar] [CrossRef] [PubMed]
- Santana-Garrido, Á.; Reyes-Goya, C.; André, H.; Aramburu, Ó.; Mate, A.; Vázquez, C.M. Sunitinib-induced oxidative imbalance and retinotoxic effects in rats. Life Sci. 2020, 257, 118072. [Google Scholar] [CrossRef]
- Santana-Garrido, Á.; Reyes-Goya, C.; Milla-Navarro, S.; de la Villa, P.; André, H.; Vázquez, C.M.; Mate, A. Anti-Inflammatory Action of Dietary Wild Olive (Acebuche) Oil in the Retina of Hypertensive Mice. Foods 2021, 10, 1993. [Google Scholar] [CrossRef]
- Bridges, J.P.; Gilbert, J.S.; Colson, D.; Gilbert, S.A.; Dukes, M.P.; Ryan, M.J.; Granger, J.P. Oxidative Stress Contributes to Soluble Fms-Like Tyrosine Kinase-1 Induced Vascular Dysfunction in Pregnant Rats. Am. J. Hypertens. 2009, 22, 564–568. [Google Scholar] [CrossRef]
- Grases-Pintó, B.; Torres-Castro, P.; Abril-Gil, M.; Castell, M.; Rodríguez-Lagunas, M.J.; Pérez-Cano, F.J.; Franch, À. A Preterm Rat Model for Immunonutritional Studies. Nutrients 2019, 11, 999. [Google Scholar] [CrossRef]
- Zuo, J.; Jiang, Z. Melatonin attenuates hypertension and oxidative stress in a rat model of L-NAME-induced gestational hypertension. Vasc. Med. 2020, 25, 295–301. [Google Scholar] [CrossRef]
- Maynard, S.E.; Min, J.-Y.; Merchan, J.; Lim, K.-H.; Li, J.; Mondal, S.; Libermann, T.A.; Morgan, J.P.; Sellke, F.W.; Stillman, I.E.; et al. Excess placental soluble fms-like tyrosine kinase 1 (sFlt1) may contribute to endothelial dysfunction, hypertension, and proteinuria in preeclampsia. J. Clin. Investig. 2003, 111, 649–658. [Google Scholar] [CrossRef]
- Caillon, H.; Tardif, C.; Dumontet, E.; Winer, N.; Masson, D. Evaluation of sFlt-1/PlGF ratio for predicting and improving clinical management of pre-eclampsia: Experience in a specialized perinatal care center. Ann. Lab. Med. 2018, 38, 95–101. [Google Scholar] [CrossRef]
- Agunanne, E.E.; Uddin, M.N.; Horvat, D.; Puschett, J.B. Contribution of angiogenic factors in a rat model of preeclampsia. Am. J. Nephrol. 2010, 32, 332–339. [Google Scholar] [CrossRef]
- Chen, X.; Huang, J.; Lv, Y.; Chen, Y.; Rao, J. Crocin exhibits an antihypertensive effect in a rat model of gestational hypertension and activates the Nrf-2/HO-1 signaling pathway. Hypertens. Res. 2021, 44, 642–650. [Google Scholar] [CrossRef]
- Cim, N.; Kurdoglu, M.; Ege, S.; Yoruk, I.; Yaman, G.; Yildizhan, R. An analysis on the roles of angiogenesis-related factors including serum vitamin D, soluble endoglin (sEng), soluble fms-like tyrosine kinase 1 (sFlt1), and vascular endothelial growth factor (VEGF) in the diagnosis and severity of late-onset preeclampsia. J. Matern. Neonatal Med. 2017, 30, 1602–1607. [Google Scholar] [CrossRef]
- Venkatesha, S.; Toporsian, M.; Lam, C.; Hanai, J.I.; Mammoto, T.; Kim, Y.M.; Bdolah, Y.; Lim, K.H.; Yuan, H.T.; Libermann, T.A.; et al. Soluble endoglin contributes to the pathogenesis of preeclampsia. Nat. Med. 2006, 12, 642–649. [Google Scholar] [CrossRef]
- Vogtmann, R.; Heupel, J.; Herse, F.; Matin, M.; Hagmann, H.; Bendix, I.; Kräker, K.; Dechend, R.; Winterhager, E.; Kimmig, R.; et al. Circulating Maternal sFLT1 (Soluble fms-Like Tyrosine Kinase-1) Is Sufficient to Impair Spiral Arterial Remodeling in a Preeclampsia Mouse Model. Hypertension 2021, 78, 1067–1079. [Google Scholar] [CrossRef]
- Schoots, M.H.; Gordijn, S.J.; Scherjon, S.A.; van Goor, H.; Hillebrands, J.L. Oxidative stress in placental pathology. Placenta 2018, 69, 153–161. [Google Scholar] [CrossRef]
- Moulin, B.; Hertig, A.; Rondeau, E. Kidney and Preeclampsia. Ann. Fr. Anesth. Reanim. 2010, 29, e83–e90. [Google Scholar] [CrossRef]
- Sudo, M.; Yoshita, K.; Ito, Y.; Imai, N.; Iino, N.; Narita, I. Histopathological features of kidney and renal prognosis in patients with preeclampsia. Pregnancy Hypertens. 2021, 25, 75–80. [Google Scholar] [CrossRef]
- Li, W.; Mata, K.M.; Mazzuca, M.Q.; Khalil, R.A. Altered matrix metalloproteinase-2 and -9 expression/activity links placental ischemia and anti-angiogenic sFlt-1 to uteroplacental and vascular remodeling and collagen deposition in hypertensive pregnancy. Biochem. Pharmacol. 2014, 89, 370–385. [Google Scholar] [CrossRef]
- Feng, Y.; Chen, X.; Wang, H.; Chen, X.; Lan, Z.; Li, P.; Cao, Y.; Liu, M.; Lv, J.; Chen, Y.; et al. Collagen I Induces Preeclampsia-Like Symptoms by Suppressing Proliferation and Invasion of Trophoblasts. Front. Endocrinol. 2021, 12, 664766. [Google Scholar] [CrossRef]
- Jiang, R.; Teng, Y.; Huang, Y.; Gu, J.; Ma, L.; Li, M.; Zhou, Y. Preeclampsia serum-induced collagen I expression and intracellular calcium levels in arterial smooth muscle cells are mediated by the PLC-γ1 pathway. Exp. Mol. Med. 2014, 46, e115. [Google Scholar] [CrossRef]
- Tam, K.B.T.; Lamarca, B.; Arany, M.; Cockrell, K.; Fournier, L.; Murphy, S.; Martin, J.N.; Granger, J.P. Role of reactive oxygen species during hypertension in response to chronic antiangiogenic factor (sFlt-1) excess in pregnant rats. Am. J. Hypertens. 2011, 24, 110–113. [Google Scholar] [CrossRef]
- Vangrieken, P.; Al-Nasiry, S.; Bast, A.; Leermakers, P.A.; Tulen, C.B.M.; Schiffers, P.M.H.; van Schooten, F.J.; Remels, A.H.V. Placental Mitochondrial Abnormalities in Preeclampsia. Reprod. Sci. 2021, 28, 2186–2199. [Google Scholar] [CrossRef]
- Cui, X.-L.; Brockman, D.; Campos, B.; Myatt, L. Expression of NADPH Oxidase Isoform 1 (Nox1) in Human Placenta: Involvement in Preeclampsia. Placenta 2006, 27, 422–431. [Google Scholar] [CrossRef]
- Chen, J.; Gao, Q.; Jiang, L.; Feng, X.; Zhu, X.; Fan, X.; Mao, C.; Xu, Z. The NOX2-derived reactive oxygen species damaged endothelial nitric oxide system via suppressed BKCa/SKCa in preeclampsia. Hypertens. Res. 2017, 40, 457–464. [Google Scholar] [CrossRef]
- Choi, S.; Kim, J.A.; Na, H.Y.; Kim, J.E.; Park, S.; Han, K.H.; Kim, Y.J.; Suh, S.H. NADPH oxidase 2-derived superoxide downregulates endothelial K Ca3.1 in preeclampsia. Free Radic. Biol. Med. 2013, 57, 10–21. [Google Scholar] [CrossRef]
- Dechend, R.; Viedt, C.; Müller, D.N.; Ugele, B.; Brandes, R.P.; Wallukat, G.; Park, J.-K.; Janke, J.; Barta, P.; Theuer, J.; et al. AT 1 Receptor Agonistic Antibodies From Preeclamptic Patients Stimulate NADPH Oxidase. Circulation 2003, 107, 1632–1639. [Google Scholar] [CrossRef]
- Osol, G.; Ko, N.L.; Mandalà, M. Altered Endothelial Nitric Oxide Signaling as a Paradigm for Maternal Vascular Maladaptation in Preeclampsia. Curr. Hypertens. Rep. 2017, 19, 82. [Google Scholar] [CrossRef]
- Schmid, M.; Sollwedel, A.; Thuere, C.; Wafula, P.O.; Zenclussen, M.L.; Müller, D.N.; Gratze, P.; Woiciechowsky, C.; Volk, H.-D.; Zenclussen, A.C. Murine Pre-Eclampsia Induced by Unspecific Activation of the Immune System Correlates with Alterations in the eNOS and AT1 Receptor Expression in the Kidneys and Placenta. Placenta 2007, 28, 688–700. [Google Scholar] [CrossRef]
- Salsoso, R.; Mate, A.; Toledo, F.; Vázquez, C.M.; Sobrevia, L. Insulin requires A2B adenosine receptors to modulate the L-arginine/nitric oxide signalling in the human fetoplacental vascular endothelium from late-onset preeclampsia. Biochim. Biophys. Acta-Mol. Basis Dis. 2021, 1867, 165993. [Google Scholar] [CrossRef]
- Danielle, L.M.; Michell, D.L.; Andrews, K.L.; Chin-Dusting, J.P.F. Endothelial dysfunction in hypertension: The role of arginase. Front. Biosci.-Sch. 2011, 1, 946–960. [Google Scholar] [CrossRef]
- Murphy, S.R.; LaMarca, B.; Cockrell, K.; Arany, M.; Granger, J.P. L-arginine supplementation abolishes the blood pressure and endothelin response to chronic increases in plasma sFlt-1 in pregnant rats. Am. J. Physiol.-Regul. Integr. Comp. Physiol. 2012, 302, 259–263. [Google Scholar] [CrossRef] [PubMed]
- Shu, W.; Li, H.; Gong, H.; Zhang, M.; Niu, X.; Ma, Y.; Zhang, X.; Cai, W.; Yang, G.; Wei, M.; et al. Evaluation of blood vessel injury, oxidative stress and circulating inflammatory factors in an L-NAME-induced preeclampsia-like rat model. Exp. Ther. Med. 2018, 16, 585–594. [Google Scholar] [CrossRef] [PubMed]
- Guerby, P.; Tasta, O.; Swiader, A.; Pont, F.; Bujold, E.; Parant, O.; Vayssiere, C.; Salvayre, R.; Negre-Salvayre, A. Role of oxidative stress in the dysfunction of the placental endothelial nitric oxide synthase in preeclampsia. Redox Biol. 2021, 40, 101861. [Google Scholar] [CrossRef]
- Shaamash, A.H.; Elsonosy, E.D.; Zakhari, M.M.; Radwan, S.H.; El-Dien, H.M. Placental nitric oxide synthase (NOS) activity and nitric oxide (NO) production in normal pregnancy, pre-eclampsia and eclampsia. Int. J. Gynecol. Obstet. 2001, 72, 127–133. [Google Scholar] [CrossRef]
- Zhao, S.; Gu, X.; Groome, L.J.; Wang, Y. Decreased Nephrin and GLEPP-1, But Increased VEGF, Flt-1, and Nitrotyrosine, Expressions in Kidney Tissue Sections From Women With Preeclampsia. Reprod. Sci. 2009, 16, 970–979. [Google Scholar] [CrossRef]
- Saif, J.; Ahmad, S.; Rezai, H.; Litvinova, K.; Sparatore, A.; Alzahrani, F.A.; Wang, K.; Ahmed, A. Hydrogen sulfide releasing molecule MZe786 inhibits soluble Flt-1 and prevents preeclampsia in a refined RUPP mouse model. Redox Biol. 2021, 38, 101814. [Google Scholar] [CrossRef]
- Bosco, C.; González, J.; Gutiérrez, R.; Parra-Cordero, M.; Barja, P.; Rodrigo, R. Oxidative damage to pre-eclamptic placenta: Immunohistochemical expression of VEGF, nitrotyrosine residues and von Willebrand factor. J. Matern. Neonatal Med. 2012, 25, 2339–2345. [Google Scholar] [CrossRef]
- Sutton, E.F.; Gemmel, M.; Powers, R.W. Nitric oxide signaling in pregnancy and preeclampsia. Nitric Oxide 2020, 95, 55–62. [Google Scholar] [CrossRef]
- Huang, J.; Zheng, L.; Wang, F.; Su, Y.; Kong, H.; Xin, H. Mangiferin ameliorates placental oxidative stress and activates PI3K/Akt/mTOR pathway in mouse model of preeclampsia. Arch. Pharm. Res. 2020, 43, 233–241. [Google Scholar] [CrossRef]
- Roland, L.; Beauchemin, D.; Acteau, G.; Fradette, C.; St-Pierre, I.; Bilodeau, J.F. Effects of Labor on Placental Expression of Superoxide Dismutases in Preeclampsia. Placenta 2010, 31, 392–400. [Google Scholar] [CrossRef] [PubMed]
- McCarthy, C.; Kenny, L.C. Therapeutically targeting mitochondrial redox signalling alleviates endothelial dysfunction in preeclampsia. Sci. Rep. 2016, 6, 32683. [Google Scholar] [CrossRef] [PubMed]
- Nuzzo, A.M.; Camm, E.J.; Sferruzzi-Perri, A.N.; Ashmore, T.J.; Yung, H.; Cindrova-Davies, T.; Spiroski, A.-M.; Sutherland, M.R.; Logan, A.; Austin-Williams, S.; et al. Placental Adaptation to Early-Onset Hypoxic Pregnancy and Mitochondria-Targeted Antioxidant Therapy in a Rodent Model. Am. J. Pathol. 2018, 188, 2704–2716. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.-Q.; Zhang, L. Mitochondrial Dysfunction in the Pathogenesis of Preeclampsia. Curr. Hypertens. Rep. 2022, 24, 157–172. [Google Scholar] [CrossRef]
- Kaur, G.; Mishra, S.; Sehgal, A.; Prasad, R. Alterations in lipid peroxidation and antioxidant status in pregnancy with preeclampsia. Mol. Cell. Biochem. 2008, 313, 37–44. [Google Scholar] [CrossRef]
- Zhang, Y.; Gu, Y.; Lewis, D.F.; Wang, Y. Reduced Cellular Glutathione Reductase Activity and Increased Adhesion Molecule Expression in Endothelial Cells Cultured With Maternal Plasma From Women With Preeclampsia. J. Soc. Gynecol. Investig. 2006, 13, 412–417. [Google Scholar] [CrossRef]
- Myatt, L.; Webster, R.P. Vascular biology of preeclampsia. J. Thromb. Haemost. 2009, 7, 375–384. [Google Scholar] [CrossRef]
- Joharapurkar, A.; Patel, V.; Kshirsagar, S.; Patel, M.S.; Savsani, H.; Jain, M. Effect of dual PPAR-α/γ agonist saroglitazar on diabetic retinopathy and oxygen-induced retinopathy. Eur. J. Pharmacol. 2021, 899, 174032. [Google Scholar] [CrossRef]
- Bo, Q.-L.; Chen, Y.-H.; Yu, Z.; Fu, L.; Zhou, Y.; Zhang, G.-B.; Wang, H.; Zhang, Z.-H.; Xu, D.-X. Rosiglitazone pretreatment protects against lipopolysaccharide-induced fetal demise through inhibiting placental inflammation. Mol. Cell. Endocrinol. 2016, 423, 51–59. [Google Scholar] [CrossRef]
- Allam, H.I.G.; Masri, A.A.A. The Potential Therapeutic Role Of Peroxisome Proliferatoractivated Receptors Agonist In Preeclamptic Pregnant Rats. J. Coll. Physicians Surg. Pakistan 2018, 28, 31–35. [Google Scholar] [CrossRef]
- Harmon, A.C.; Cornelius, D.C.; Amaral, L.M.; Faulkner, J.L.; Cunningham, M.W.; Wallace, K.; LaMarca, B. The role of inflammation in the pathology of preeclampsia. Clin. Sci. 2016, 130, 409–419. [Google Scholar] [CrossRef]
- Zheng, L.; Tang, R.; Shi, L.; Zhong, M.; Zhou, Z. Vagus nerve stimulation ameliorates L-NAME-induced preeclampsia-like symptoms in rats through inhibition of the inflammatory response. BMC Pregnancy Childbirth 2021, 21, 177. [Google Scholar] [CrossRef]
- Lin, Z.; Jin, J.; Shan, X. The effects of estradiol on inflammatory and endothelial dysfunction in rats with preeclampsia. Int. J. Mol. Med. 2020, 45, 825–835. [Google Scholar] [CrossRef]
- Zhou, C.C.; Irani, R.A.; Dai, Y.; Blackwell, S.C.; Hicks, M.J.; Ramin, S.M.; Kellems, R.E.; Xia, Y. Autoantibody-Mediated IL-6–Dependent Endothelin-1 Elevation Underlies Pathogenesis in a Mouse Model of Preeclampsia. J. Immunol. 2011, 186, 6024–6034. [Google Scholar] [CrossRef]
- Hayashi, M.; Ueda, Y.; Ohkura, T.; Inaba, N. Interleukin-6 Concentrations in the Placenta and Blood in Normal Pregnancies and Preeclampsia. Horm. Metab. Res. 2005, 37, 419–424. [Google Scholar] [CrossRef]
- Al-Othman, S.; Omu, A.E.; Diejomaoh, F.M.E.; Al-Yatama, M.; Al-Qattan, F. Differential levels of interleukin 6 in maternal and cord sera and placenta in women with pre-eclampsia. Gynecol. Obstet. Investig. 2001, 52, 60–65. [Google Scholar] [CrossRef]
- Roland, L.; Gagné, A.; Bélanger, M.C.; Boutet, M.; Julien, P.; Bilodeau, J.F. Plasma interleukin-18 (IL-18) levels are correlated with antioxidant vitamin coenzyme Q10 in preeclampsia. Acta Obstet. Gynecol. Scand. 2010, 89, 360–366. [Google Scholar] [CrossRef]
- Huang, X.; Huang, H.; Dong, M.; Yao, Q.; Wang, H. Serum and placental interleukin-18 are elevated in preeclampsia. J. Reprod. Immunol. 2005, 65, 77–87. [Google Scholar] [CrossRef]
- Hennessy, A.; Painter, D.M.; Orange, S.; Horvath, J.S. Placental tissue interleukin-10 receptor distribution in pre-eclampsia. Am. J. Reprod. Immunol. 2003, 49, 377–381. [Google Scholar] [CrossRef]
- Orange, S.; Rasko, J.E.; Thompson, J.F.; Vaughan, J.; Olive, E.; Pedler, M.; Horvath, J.S.; Hennessy, A. Interleukin-10 regulates arterial pressure in early primate pregnancy. Cytokine 2005, 29, 176–185. [Google Scholar] [CrossRef]
- Tinsley, J.H.; South, S.; Chiasson, V.L.; Mitchell, B.M. Interleukin-10 reduces inflammation, endothelial dysfunction, and blood pressure in hypertensive pregnant rats. Am. J. Physiol.-Regul. Integr. Comp. Physiol. 2010, 298, 713–719. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer (5’→3’) | Reverse Primer (5’→3’) | Accession Number |
|---|---|---|---|
| sFlt1 | CAAGGGACTCTACACTTGTC | CCGAATAGCGAGCAGATTTC | AF157595.1 |
| PlGF | CAGCCAACATCACTATGCAG | TCCTCTGAGTGGCTGGTTA | NM_053595.2 |
| Endoglin | CCAAGGCTGCCACTTGG | GATGCTGTGGTTGGTAC | NM_001010968.3 |
| Nox1 | GGTTGGGGCTGAACATTTTTC | TCGACACACAGGAATCAGGAT | NM_172203.2 |
| Nox2 | CCCTTTGGTACAGCCAGTGAAGAT | CAATCCCACGTCCCACTAACATCA | AF298656.3 |
| Nox4 | GGATCACAGAAGGTCCCTAGC | AGAAGTTCAGGGCGTTCACC | NM_053524.1 |
| Arginase 1 | ATTTCGGTGGTTTAAGGTAGTCAG | TACAAGACAGGGCTACTTTCAG | NM_017134.3 |
| Arginase 2 | CCTCTTCCTCTTGCCAATCAG | CAGCCTCTTTCCTTTCTCATCAG | U90887.1 |
| SOD1 | AATGTGTCCATTGAAGATCGTGTGA | GCTTCCAGCATTTCCAGTCTTTGTA | NM_017050.1 |
| SOD2 | AGGGCCTGTCCCATGATGTC | AGAAACCCGTTTGCCTCTACTGAA | NM_017051.2 |
| SOD3 | GGGTCTGTCCTGTACTTCACCAGAG | CTGACATGGTCCAGGTGACAGAG | NM_012880.2 |
| GSH-Px | GGAGAATGGCAAGAATGAAGA | CCGCAGGAAGGTAAAGAG | NM_030826.4 |
| GSH-Red | GGAAACTCGCCCATAGACTT | CCAACCACCTTCTCCTCTTT | NM_053906.2 |
| PPARα | ACGATGCTGTCCTCCTTGATG | GCGTCTGACTCGGTCTTCTTG | NM_013196.2 |
| PPARγ | CTGGCCATATTTATAGCTGTCATTATT | AGCAGGTTGTCTTGGATGTCCT | Y12882.2 |
| IL1β | GAGGCTGACAGACCCCAAAAGAT | GCACGAGGCATTTTTGTTGTTCA | NM_031512.2 |
| IL3 | CTTGATGTCCATTGTGTCCTGAG | TCCTGATGCTCTTCCACCAG | NM_031513.2 |
| IL6 | TTCTGCAAGTGCATCATCGT | CTCTGCAAGAGACTTCCATCC | NM_012589.2 |
| IL10 | AGGCCATTCCATCCGGGGTGA | AGGCAGCCCTCAGCTCTCGG | NM_012854.2 |
| IL18 | TCCTTCACAGAGAGGGTCACA | GACAGCCTGTGTTCGAGGAT | NM_019165.2 |
| GAPDH | GCCAAAAGGGTCATCATCTCCGC | GGATGACCTTGCCCACAGCCTTG | NM_017008.4 |
| 1st Antibody | Origin | Dilution | 2nd Antibody | Dilution | Reference |
|---|---|---|---|---|---|
| Anti-VEGFR1 | Rabbit monoclonal | 1:2000 | Goat Anti-Rabbit | 1:4000 | Epitomics-Abcam, Burlingame, CA |
| Anti-NOX1 | Mouse monoclonal | 1:1000 | Goat Anti-Mouse | 1:2000 | SCB 1 |
| Anti-NOX2 | Rabbit monoclonal | 1:9000 | Goat Anti-Rabbit | 1:10,000 | Epitomics-Abcam |
| Anti-NOX4 | Rabbit monoclonal | 1:9000 | Goat Anti-Rabbit | 1:10,000 | Epitomics-Abcam |
| Anti-nitrotyrosine | Mouse Monoclonal | 1:1000 | Goat Anti-Mouse | 1:2000 | SCB |
| Anti-arginase 1 | Mouse monoclonal | 1:1000 | Goat Anti-Mouse | 1:2000 | SCB |
| Anti-arginase 2 | Mouse monoclonal | 1:1000 | Goat Anti-Mouse | 1:2000 | SCB |
| Anti-GSH-Px | Mouse monoclonal | 1:1000 | Goat Anti-Mouse | 1:4000 | SCB |
| Anti-GSH-Red | Rabbit polyclonal | 1:5000 | Goat Anti-Rabbit | 1:8000 | SCB |
| Anti-SOD-1 | Mouse monoclonal | 1:1000 | Goat Anti-Mouse | 1:2000 | SCB |
| Anti-SOD-2 | Mouse monoclonal | 1:1000 | Goat Anti-Mouse | 1:2000 | SCB |
| Anti-PPARα | Mouse monoclonal | 1:2000 | Goat Anti-Mouse | 1:4000 | SCB |
| Anti-PPARγ | Mouse monoclonal | 1:2000 | Goat Anti-Mouse | 1:4000 | SCB |
| Anti-β-Actin | Mouse monoclonal | 1:20,000 | Goat Anti-Mouse | 1:30,000 | SCB |
| Parameters | NP | PE-19 |
|---|---|---|
| Animal weight (g) | 278 ± 3.9 | 267 ± 4.3 |
| Embryo weight (g) | 2.29 ± 0.12 | 1.54 ± 0.10 *** |
| Average no. embryos | 10 | 10 |
| Kidney weight (g) | 0.71 ± 0.02 | 0.75 ± 0.02 |
| Placenta weight (g) | 0.68 ± 0.021 | 0.44 ± 0.066 ** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santana-Garrido, Á.; Reyes-Goya, C.; Espinosa-Martín, P.; Sobrevia, L.; Beltrán, L.M.; Vázquez, C.M.; Mate, A. Oxidative and Inflammatory Imbalance in Placenta and Kidney of sFlt1-Induced Early-Onset Preeclampsia Rat Model. Antioxidants 2022, 11, 1608. https://doi.org/10.3390/antiox11081608
Santana-Garrido Á, Reyes-Goya C, Espinosa-Martín P, Sobrevia L, Beltrán LM, Vázquez CM, Mate A. Oxidative and Inflammatory Imbalance in Placenta and Kidney of sFlt1-Induced Early-Onset Preeclampsia Rat Model. Antioxidants. 2022; 11(8):1608. https://doi.org/10.3390/antiox11081608
Chicago/Turabian StyleSantana-Garrido, Álvaro, Claudia Reyes-Goya, Pablo Espinosa-Martín, Luis Sobrevia, Luis M. Beltrán, Carmen M. Vázquez, and Alfonso Mate. 2022. "Oxidative and Inflammatory Imbalance in Placenta and Kidney of sFlt1-Induced Early-Onset Preeclampsia Rat Model" Antioxidants 11, no. 8: 1608. https://doi.org/10.3390/antiox11081608