
Investigation of the Hydrogen Sulfide Signaling Pathway in Schwann Cells during Peripheral Nerve Degeneration: Multi-Omics Approaches

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Ex Vivo Sciatic Nerve Culture

2.3. Cell Culture and Chemicals
2.4. Proteomic Sample Preparation
2.5. Tandem Mass Tag (TMT)-Based Quantitative Proteomics
2.6. Proteomic Data Analysis

2.7. RNA Preparation and Quantitative Real-Time Polymerase Chain Reaction (qPCR)
2.8. Whole-Genome Profiling
2.9. miRNA Expression Profiling
2.10. Bioinformatics
2.11. Statistical Analysis
3. Results
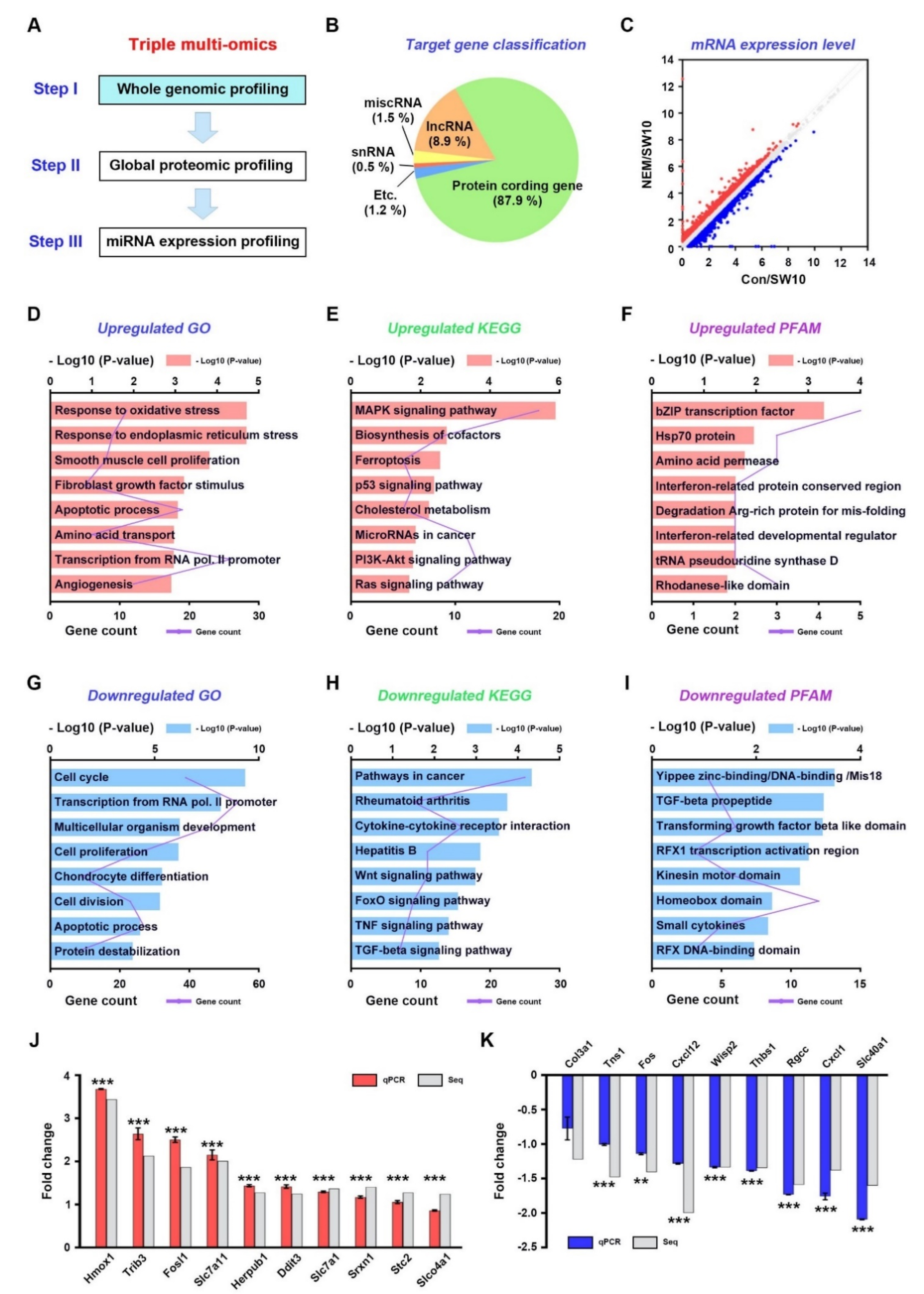
3.1. Whole-Genome Expression Profiling of Dedifferentiated Schwann Cells
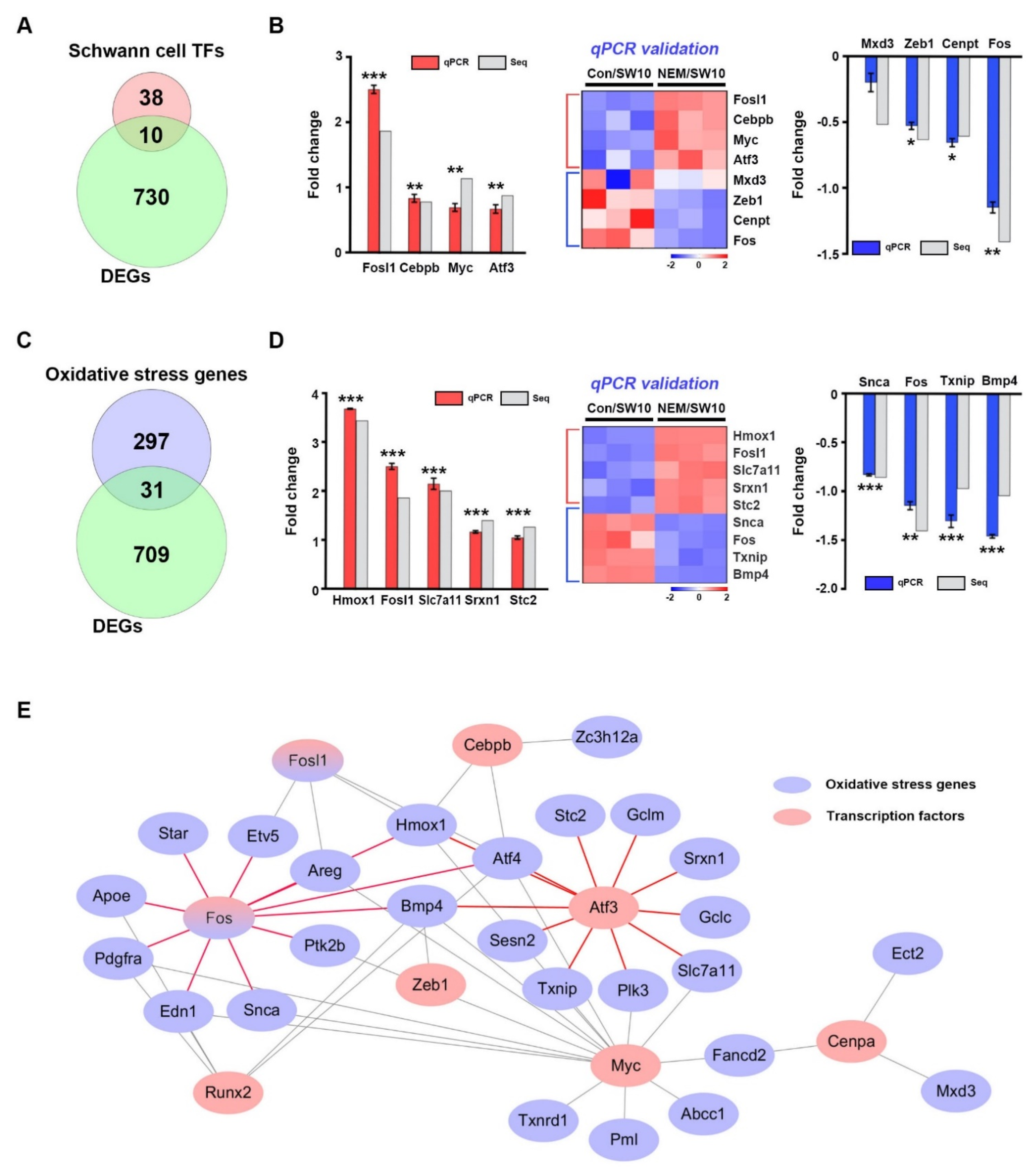
3.2. Transcriptional Regulation of Oxidative Stress in the H2S Signaling Pathway

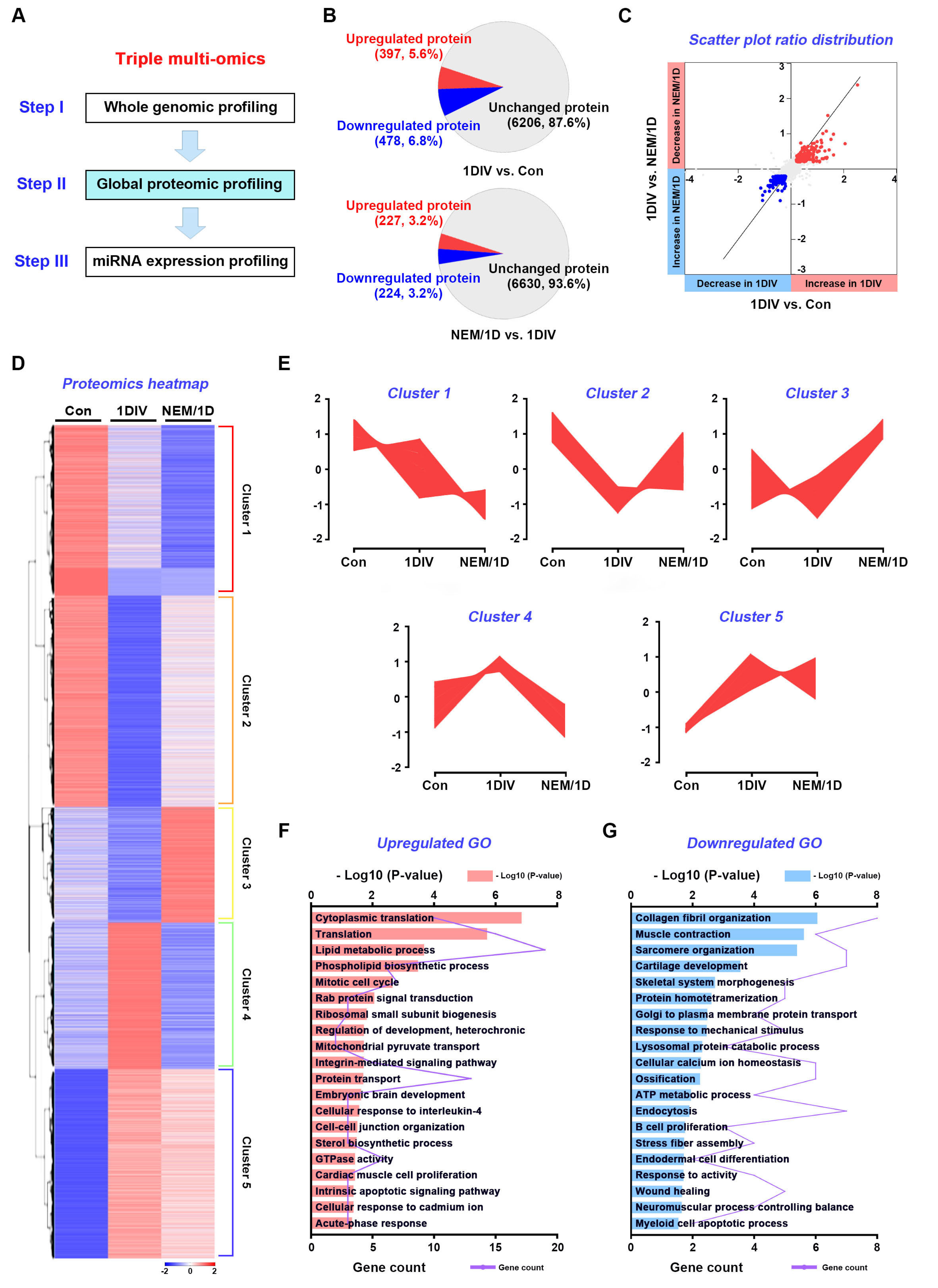
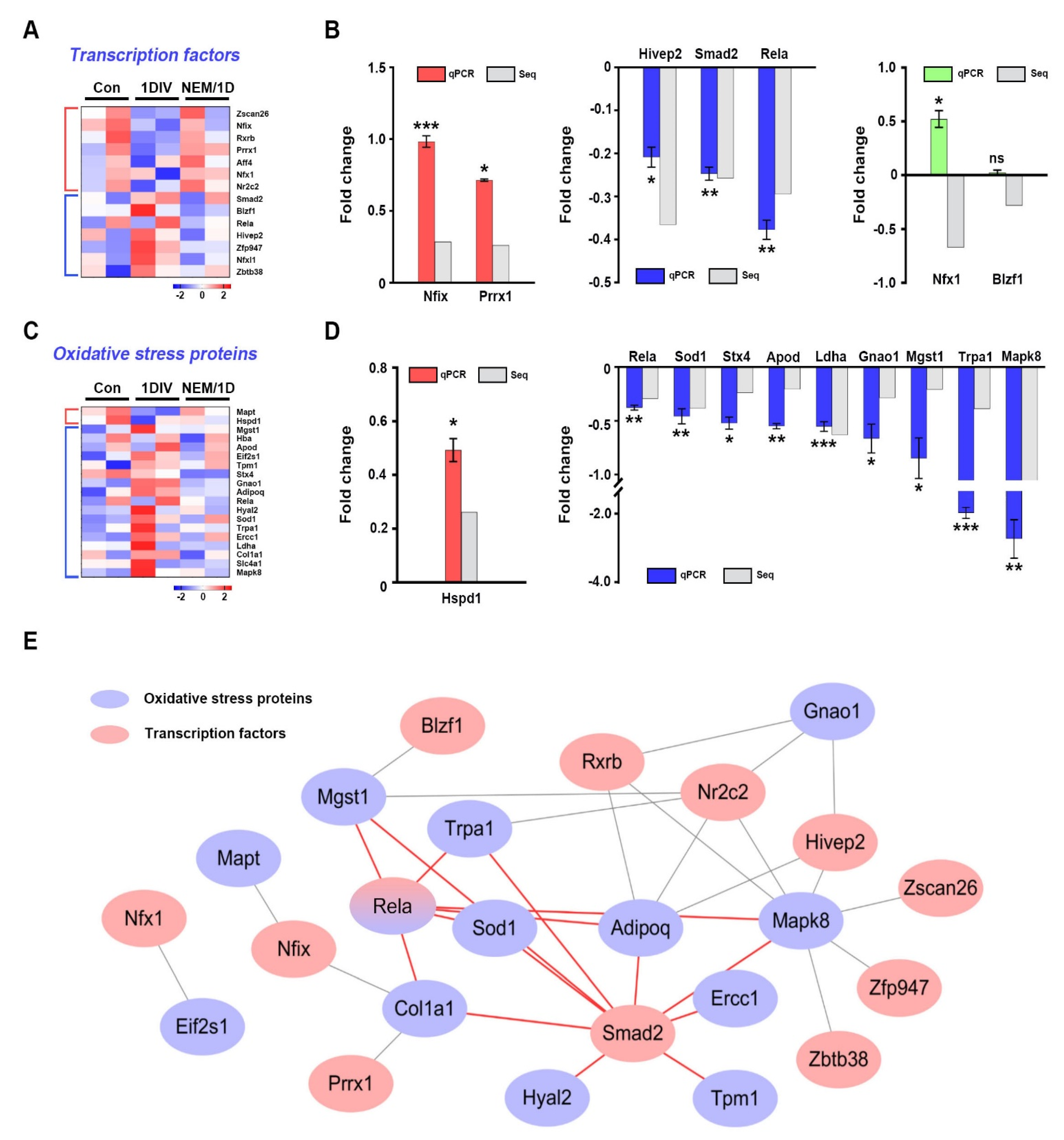
3.3. Quantitative Proteomic Analysis of the H2S Signaling Pathway in Sciatic Nerves Ex Vivo
3.4. Interactions of Transcription Factors and Oxidative Stress Proteins in Sciatic Nerves Ex Vivo
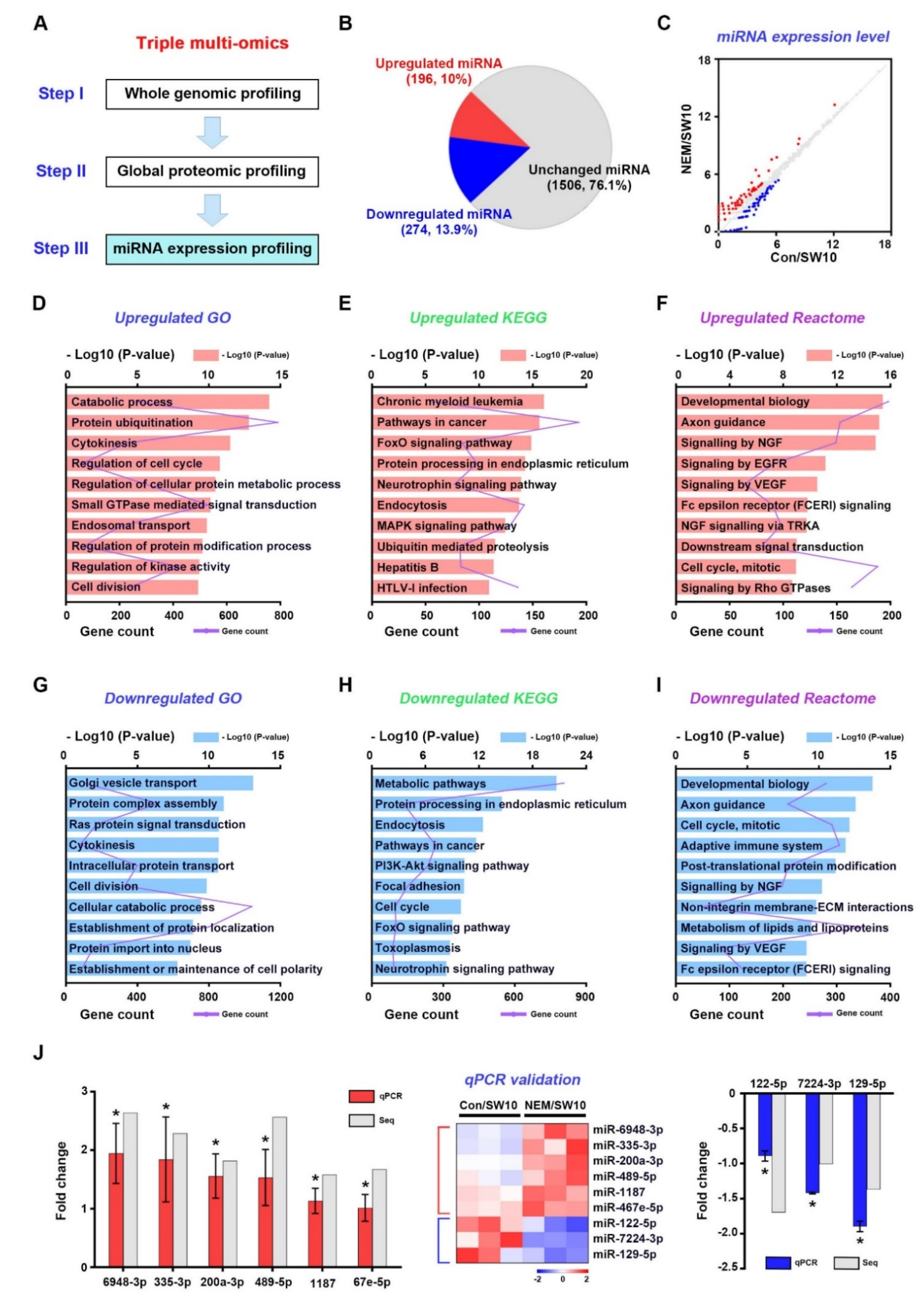
3.5. miRNA Expression Profiling of the H2S Signaling Pathway in Dedifferentiated Schwann Cells
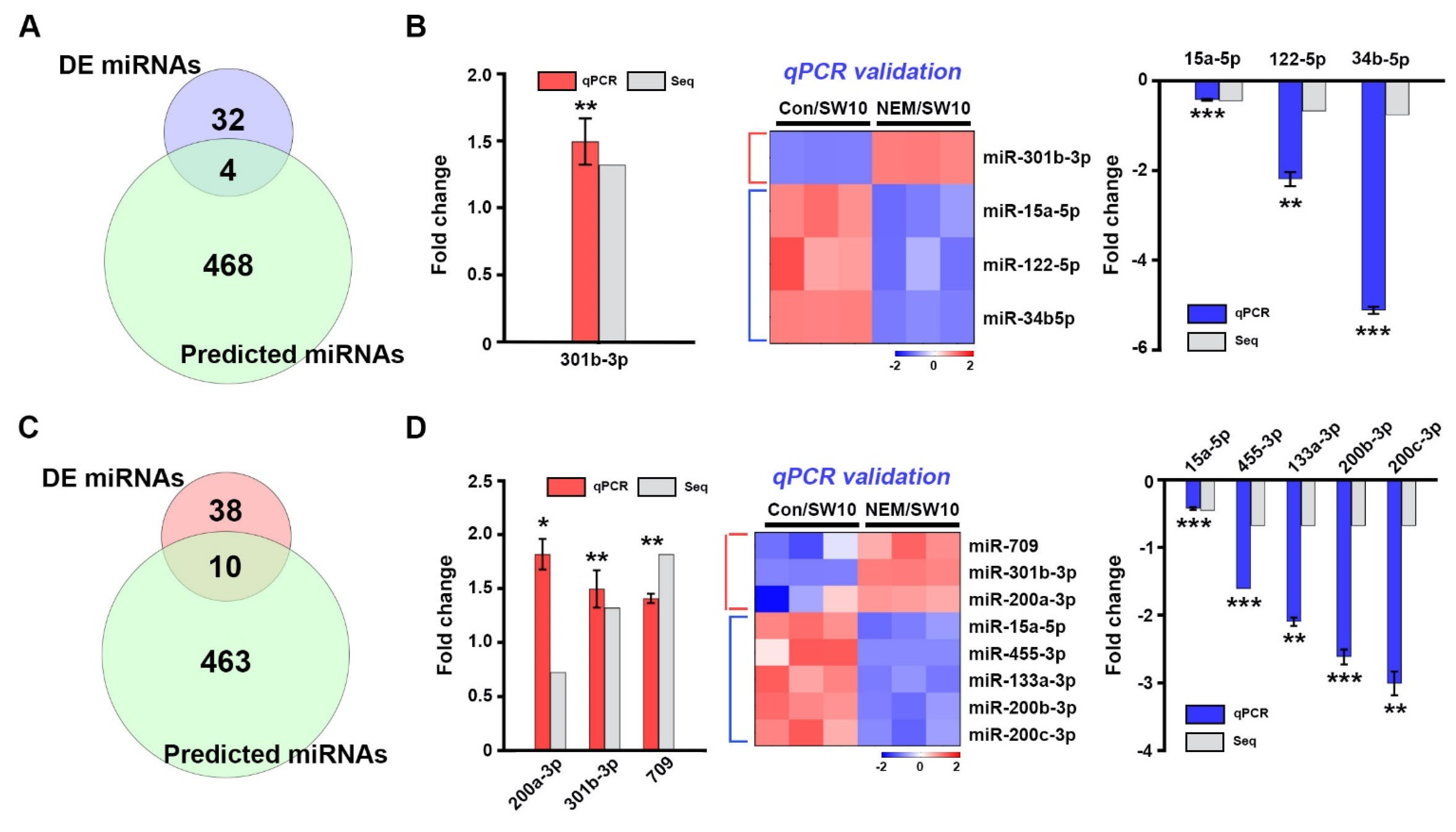
3.6. Post-Transcriptional Regulation of the Response to Oxidative Stress
4. Discussion


5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Jessen, K.R.; Mirsky, R. The success and failure of the Schwann cell response to nerve injury. Front. Cell. Neurosci. 2019, 11, 13–33. [Google Scholar] [CrossRef] [PubMed]
- Sood, A.; Kumar, B.; Singh, S.K.; Prashar, P.; Gautam, A.; Gulati, M.; Pandey, N.K.; Melkani, I.; Awasthi, A.; Saraf, S.A.; et al. Flavonoids as potential therapeutic agents for the management of diabetic neuropathy. Curr. Pharm. Des. 2020, 26, 5468–5487. [Google Scholar] [CrossRef] [PubMed]
- Wang, R. Gasotransmitters: Growing pains and joys. Trends Biochem. Sci. 2014, 39, 227–232. [Google Scholar] [CrossRef] [PubMed]
- Kimura, H. Production and physiological effects of hydrogen sulfide. Antioxid Redox Signal. 2014, 20, 783–793. [Google Scholar] [CrossRef] [PubMed]
- Kabil, O.; Motl, N.; Banerjee, R. H2S and its role in redox signaling. Biochim. Biophys. Acta 2014, 1844, 1355–1366. [Google Scholar] [CrossRef]
- Wang, R. Physiological implications of hydrogen sulfide: A whiff exploration that blossomed. Physiol. Rev. 2012, 92, 791–896. [Google Scholar] [CrossRef]
- Ishigami, M.; Hiraki, K.; Umemura, K.; Ogasawara, Y.; Ishii, K.; Kimura, H. A source of hydrogen sulfide and a mechanism of its release in the brain. Antioxid. Redox Signal. 2009, 11, 205–214. [Google Scholar] [CrossRef]
- Park, B.S.; Kim, H.-W.; Rhyu, I.J.; Park, C.; Yeo, S.G.; Huh, Y.; Jeong, N.Y.; Jung, J. Hydrogen sulfide is essential for Schwann cell responses to peripheral nerve injury. J. Neurochem. 2015, 132, 230–242. [Google Scholar] [CrossRef]
- Tan-No, K.; Takahashi, H.; Nakagawasai, O.; Niijima, F.; Sakurada, S.; Bakalkin, G.; Terenius, L.; Tadano, T. Nociceptive behavior induced by the endogenous opioid peptides dynorphins in uninjured mice: Evidence with intrathecal N-ethylmaleimide inhibiting dynorphin degradation. Int. Rev. Neurobiol. 2009, 85, 191–205. [Google Scholar] [CrossRef]
- Tyanova, S.; Temu, T.; Cox, J. The MaxQuant computational platform for mass spectrometry-based shotgun proteomics. Nat. Protoc. 2016, 11, 2301–2319. [Google Scholar] [CrossRef]
- Tyanova, S.; Temu, T.; Sinitcyn, P.; Carlson, A.; Hein, M.Y.; Geiger, T.; Mann, M.; Cox, J. The Perseus computational platform for comprehensive analysis of (prote)omics data. Nat. Methods. 2016, 13, 731–740. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.B.; Jang, W.E.; Park, J.H.; Mohammad, H.B.; Lee, J.-Y.; Jeong, W.-H.; Kim, M.-S. Identification of organophosphate modifications by high-resolution mass spectrometry. Bull. Korean Chem. Soc. 2022, 43, 444–449. [Google Scholar] [CrossRef]
- Li, M.; Banton, M.C.; Min, Q.; Parkinson, D.B.; Dun, X. Meta-analysis reveals transcription factor upregulation in cells of injured mouse sciatic nerve. Front. Cell. Neurosci. 2021, 15, 688243. [Google Scholar] [CrossRef] [PubMed]
- Ko, K.R.; Lee, J.; Nho, B.; Kim, S. c-Fos is necessary for HGF-mediated gene regulation and cell migration in Schwann cells. Biochem. Biophys. Res. Commun. 2018, 503, 2855–2860. [Google Scholar] [CrossRef]
- Hunt, D.; Raivich, G.; Anderson, P.N. Activating transcription factor 3 and the nervous system. Front. Mol. Neurosci. 2012, 5, 7. [Google Scholar] [CrossRef]
- Binns, D.; Dimmer, E.; Huntley, R.; Barrell, D.; O’Donovan, C.; Apweiler, R. QuickGO: A web-based tool for Gene Ontology searching. Bioinformatics 2009, 25, 3045–3046. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; Kim, H.; Kim, D.; Kim, D.; Huh, Y.; Park, C.; Chung, H.-J.; Jung, J.; Jeong, N.Y. Heme Oxygenase 1 in Schwann cells regulates peripheral nerve degeneration against oxidative stress. ASN. Neuro 2019, 11, 1759091419838949. [Google Scholar] [CrossRef] [PubMed]
- Lv, W.; Deng, B.; Duan, W.; Li, Y.; Liu, Y.; Li, Z.; Xia, W.; Li, C. Schwann cell plasticity is regulated by a weakened intrinsic antioxidant defense system in acute peripheral nerve injury. Neuroscience 2018, 382, 1–13. [Google Scholar] [CrossRef]
- Wilkins, P.L.; Suchovsky, D.; Berti-Mattera, L.N. Immortalized Schwann cells express endothelin receptors coupled to adenylyl cyclase and phospholipase C. Neurochem. Res. 1997, 22, 409–418. [Google Scholar] [CrossRef]
- Ishii, T.; Itoh, K.; Takahashi, S.; Sato, H.; Yanagawa, T.; Katoh, Y.; Bannai, S.; Yamamoto, M. Transcription factor Nrf2 coordinately regulates a group of oxidative stress-inducible genes in macrophages. J. Biol. Chem. 2000, 275, 16023–160239. [Google Scholar] [CrossRef]
- Tsang, C.K.; Liu, Y.; Thomas, J.; Zhang, Y.; Zheng, X.F.S. Superoxide dismutase 1 acts as a nuclear transcription factor to regulate oxidative stress resistance. Nat. Commun. 2014, 5, 3446. [Google Scholar] [CrossRef] [PubMed]
- Zhong, H.; Voll, R.E.; Ghosh, S. Phosphorylation of NF-kappa B p65 by PKA stimulates transcriptional activity by promoting a novel bivalent interaction with the coactivator CBP/p300. Mol. Cell 1998, 1, 661–671. [Google Scholar] [CrossRef]
- Nickols, J.C.; Valentine, W.; Kanwal, S.; Carter, B.D. Activation of the transcription factor NF-kappaB in Schwann cells is required for peripheral myelin formation. Nat. Neurosci. 2003, 6, 161–167. [Google Scholar] [CrossRef] [PubMed]
- Muscella, A.; Vetrugno, C.; Cossa, L.G.; Marsigliante, S. TGF-β1 activates RSC96 Schwann cells migration and invasion through MMP-2 and MMP-9 activities. J. Neurochem. 2020, 153, 525–538. [Google Scholar] [CrossRef]
- Lobsiger, C.S.; Boillee, S.; McAlonis-Downes, M.; Khan, A.M.; Feltri, M.L.; Yamanaka, K.; Cleveland, D.W. Schwann cells expressing dismutase active mutant SOD1 unexpectedly slow disease progression in ALS mice. Proc. Natl. Acad. Sci. USA. 2009, 106, 4465–4470. [Google Scholar] [CrossRef]
- Lin, M.; Jiang, M.; Ding, F.; Cao, Z. Syntaxin-4 and SNAP23 act as exocytic SNAREs to release NGF from cultured Schwann cells. Neurosci. Lett. 2017, 653, 97–104. [Google Scholar] [CrossRef]
- Scapin, C.; Ferri, C.; Pettinato, E.; Bianchi, F.; Carro, U.D.; Feltri, M.L.; Kaufman, R.J.; Wrabetz, L.; D’Antonio, M. Phosphorylation of eIF2α promotes Schwann cell differentiation and myelination in CMT1B mice with activated UPR. J. Neurosci. 2020, 40, 8174–8187. [Google Scholar] [CrossRef]
- Yi, S.; Liu, Q.; Wang, X.; Qian, T.; Wang, H.; Zha, G.; Yu, J.; Wang, P.; Gu, X.; Chu, D.; et al. Tau modulates Schwann cell proliferation, migration and differentiation following peripheral nerve injury. J. Cell. Sci. 2019, 132, jcs222059. [Google Scholar] [CrossRef]
- Abe, K.; Kimura, H. The possible role of hydrogen sulfide as an endogenous neuromodulator. J. Neurosci. 1996, 16, 1066–1071. [Google Scholar] [CrossRef]
- Griffiths-Jones, S.; Grocock, R.J.; Dongen, S.; Bateman, A.; Enright, A.J. MiRBase: MicroRNA sequences, targets and gene nomenclature. Nucleic Acids Res. 2006, 34, D140–D144. [Google Scholar] [CrossRef]
- Geng, B.; Yang, J.; Qi, Y.; Zhao, J.; Pang, Y.; Du, J.; Tang, C. H2S generated by heart in rat and its effects on cardiac function. Biochem. Biophys. Res. Commun. 2004, 313, 362–368. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Ahmad, S.; Cai, M.; Rennie, J.; Fujisawa, T.; Crispi, F.; Baily, J.; Miller, M.R.; Cudmore, M.; Hadoke, P.W.F.; et al. Dysregulation of hydrogen sulfide producing enzyme cystathionine γ-lyase contributes to maternal hypertension and placental abnormalities in preeclampsia. Circulation. 2013, 127, 2514–2522. [Google Scholar] [CrossRef] [PubMed]
- Teng, H.; Wu, B.; Zhao, K.; Yang, G.; Wu, L.; Wang, R. Oxygen-sensitive mitochondrial accumulation of cystathionine β-synthase mediated by Lon protease. Proc. Natl. Acad. Sci. USA. 2013, 110, 12679–12684. [Google Scholar] [CrossRef] [PubMed]
- Shen, M.; Ji, Y.; Zhang, S.; Shi, H.; Chen, G.; Gu, X.; Ding, F. A proteome map of primary cultured rat Schwann cells. Proteome Sci. 2012, 10, 20. [Google Scholar] [CrossRef]
- Weiss, T.; Taschner-Mandl, S.; Bileck, A.; Slany, A.; Kromp, F.; Rifatbegovic, F.; Frech, C.; Windhager, R.; Kitzinger, H.; Tzou, C.-H.; et al. Proteomics and transcriptomics of peripheral nerve tissue and cells unravel new aspects of the human Schwann cell repair phenotype. Glia 2016, 64, 2133–2153. [Google Scholar] [CrossRef]
- Arthur-Farraj, P.J.; Morgan, C.C.; Adamowicz, M.; Gomez-Sanchez, J.A.; Fazal, S.V.; Beucher, A.; Razzaghi, B.; Mirsky, R.; Jessen, K.R.; Aitman, T.J. Changes in the coding and non-coding transcriptome and DNA methylome that define the Schwann cell repair phenotype after nerve injury. Cell Rep. 2017, 20, 2719–2734. [Google Scholar] [CrossRef]
- Schira, J.; Heinen, A.; Poschmann, G.; Ziegler, B.; Hartung, H.-P.; Stuhler, K.; Kury, P. Secretome analysis of nerve repair mediating Schwann cells reveals Smad-dependent trophism. FASEB J. 2019, 33, 4703–4715. [Google Scholar] [CrossRef]
- Barata, B.; Loehr, T.M.; Sigel, H.; Burgess, B.K.; Magnus, K.A.; Solomon, E.I.; Dilworth, J.R.; Manne, S.; Stiefel, I.; Fluck, E.; et al. Glossary of terms used in bioinorganic chemistry. Pure Appl. Chem. 1997, 69, 1251–1304. [Google Scholar] [CrossRef]
- O’Sullivan, S.A.; Velasco-Estevez, M.; Dev, K.K. Demyelination induced by oxidative stress is regulated by sphingosine 1-phosphate receptors. Glia 2017, 65, 1119–1136. [Google Scholar] [CrossRef]
- Ravera, S.; Bartolucci, M.; Cuccarolo, P.; Litame, E.; Illarcio, M.; Calzia, D.; Degan, P.; Morelli, A.; Panfoli, I. Oxidative stress in myelin sheath: The other face of the extramitochondrial oxidative phosphorylation ability. Free Radic. Res. 2015, 49, 1156–1164. [Google Scholar] [CrossRef]
- Iida, H.; Schmeichel, A.M.; Wang, Y.; Schmelzer, J.D.; Low, P.A. Schwann cell is a target in ischemia-reperfusion injury to peripheral nerve. Muscle Nerve 2004, 30, 761–766. [Google Scholar] [CrossRef] [PubMed]
- Lisak, R.P.; Bealmear, B.; Benjamins, J.A.; Skoff, A.M. Interferon-gamma, tumor necrosis factor-alpha, and transforming growth factor-beta inhibit cyclic AMP-induced Schwann cell differentiation. Glia 2001, 36, 354–363. [Google Scholar] [CrossRef] [PubMed]
- Jung, J.; Cai, W.; Jang, S.Y.; Shin, Y.K.; Suh, D.J.; Kim, J.K.; Park, H.T. Transient lysosomal activation is essential for p75 nerve growth factor receptor expression in myelinated Schwann cells during Wallerian degeneration. Anat. Cell Biol. 2011, 44, 41–49. [Google Scholar] [CrossRef]
- Min, Q.; Parkinson, D.B.; Dun, X.-P. Migrating Schwann cells direct axon regeneration within the peripheral nerve bridge. Glia 2021, 69, 235–254. [Google Scholar] [CrossRef]
- Ishii, A.; Furusho, M.; Bansal, R. Mek/ERK1/2-MAPK and PI3K/Akt/mTOR signaling plays both independent and cooperative roles in Schwann cell differentiation, myelination and dysmyelination. Glia 2021, 69, 2429–2446. [Google Scholar] [CrossRef]
- Grigoryan, T.; Stein, S.; Qi, J.; Wende, H.; Garratt, A.N.; Nave, K.-A.; Birchmeier, C.; Birchmeier, W. Wnt/Rspondin/β-catenin signals control axonal sorting and lineage progression in Schwann cell development. Proc. Natl. Acad. Sci. USA 2013, 110, 18174–18179. [Google Scholar] [CrossRef] [PubMed]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chun, Y.L.; Eom, W.-J.; Lee, J.H.; Nguyen, T.N.C.; Park, K.-H.; Chung, H.-J.; Seo, H.; Huh, Y.; Kim, S.H.; Yeo, S.G.; et al. Investigation of the Hydrogen Sulfide Signaling Pathway in Schwann Cells during Peripheral Nerve Degeneration: Multi-Omics Approaches. Antioxidants 2022, 11, 1606. https://doi.org/10.3390/antiox11081606
Chun YL, Eom W-J, Lee JH, Nguyen TNC, Park K-H, Chung H-J, Seo H, Huh Y, Kim SH, Yeo SG, et al. Investigation of the Hydrogen Sulfide Signaling Pathway in Schwann Cells during Peripheral Nerve Degeneration: Multi-Omics Approaches. Antioxidants. 2022; 11(8):1606. https://doi.org/10.3390/antiox11081606
Chicago/Turabian StyleChun, Yoo Lim, Won-Joon Eom, Jun Hyung Lee, Thy N. C. Nguyen, Ki-Hoon Park, Hyung-Joo Chung, Han Seo, Youngbuhm Huh, Sang Hoon Kim, Seung Geun Yeo, and et al. 2022. "Investigation of the Hydrogen Sulfide Signaling Pathway in Schwann Cells during Peripheral Nerve Degeneration: Multi-Omics Approaches" Antioxidants 11, no. 8: 1606. https://doi.org/10.3390/antiox11081606