
Modulating Nitric Oxide Dioxygenase and Nitrite Reductase of Cytoglobin through Point Mutations

Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Recombinant Human Cytoglobin Expression and Purification
2.3. Site-Directed Mutagenesis of Human Cytoglobin
2.4. Nitric Oxide Dioxygenase Activity
2.5. Nitric Oxide Binding
2.6. Nitrite Reductase Activity
3. Results
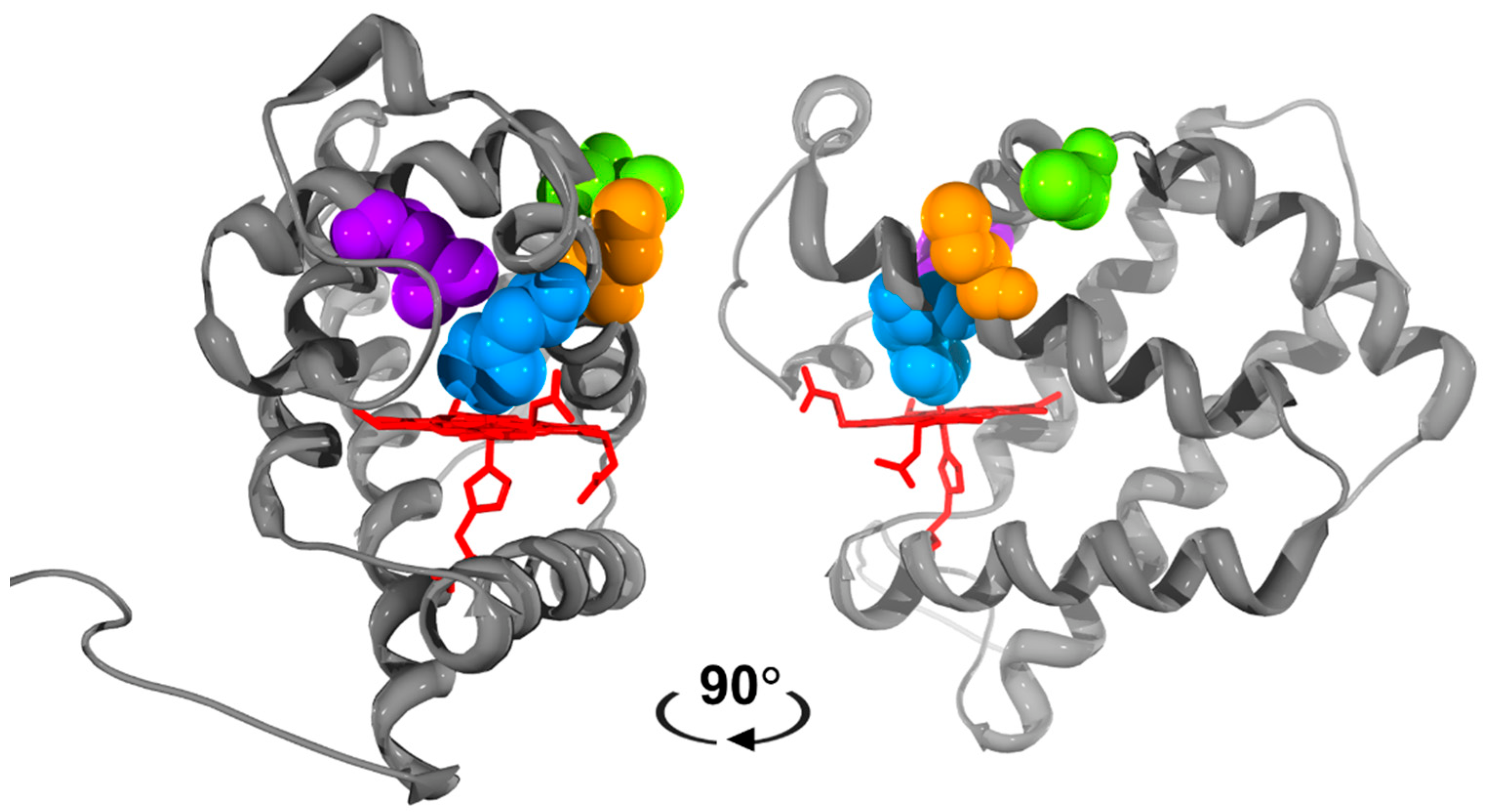
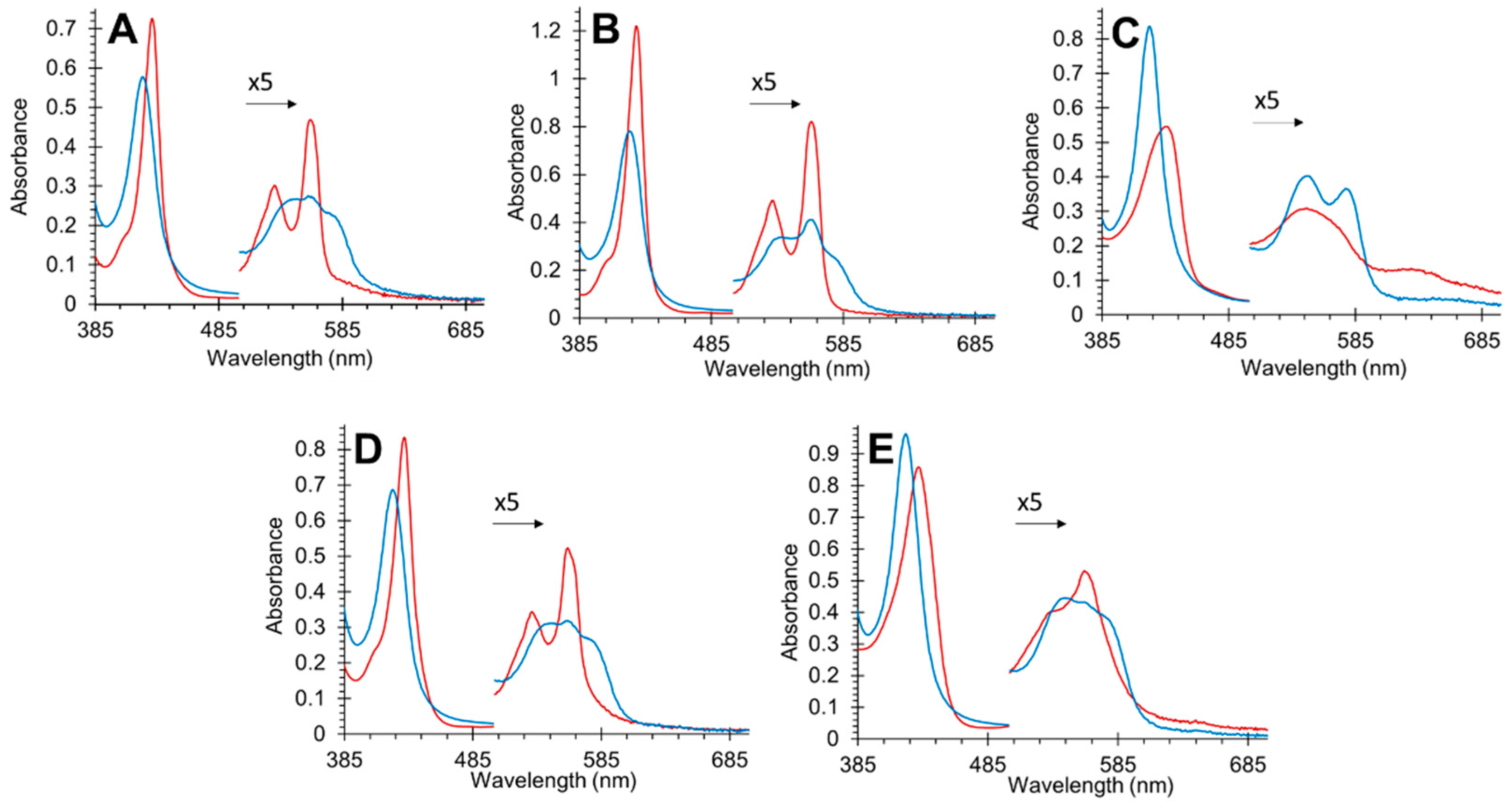
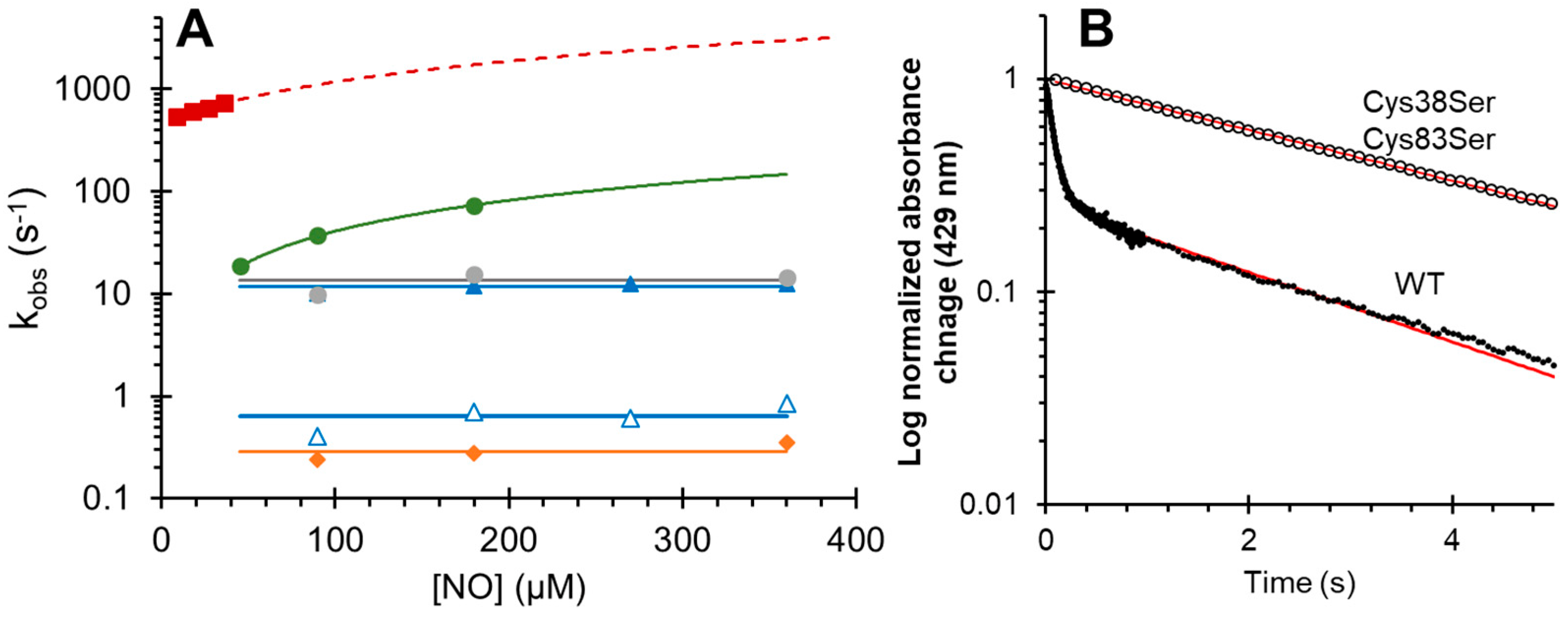
3.1. Effect of Mutations on Nitric Oxide Binding to Cytoglobin
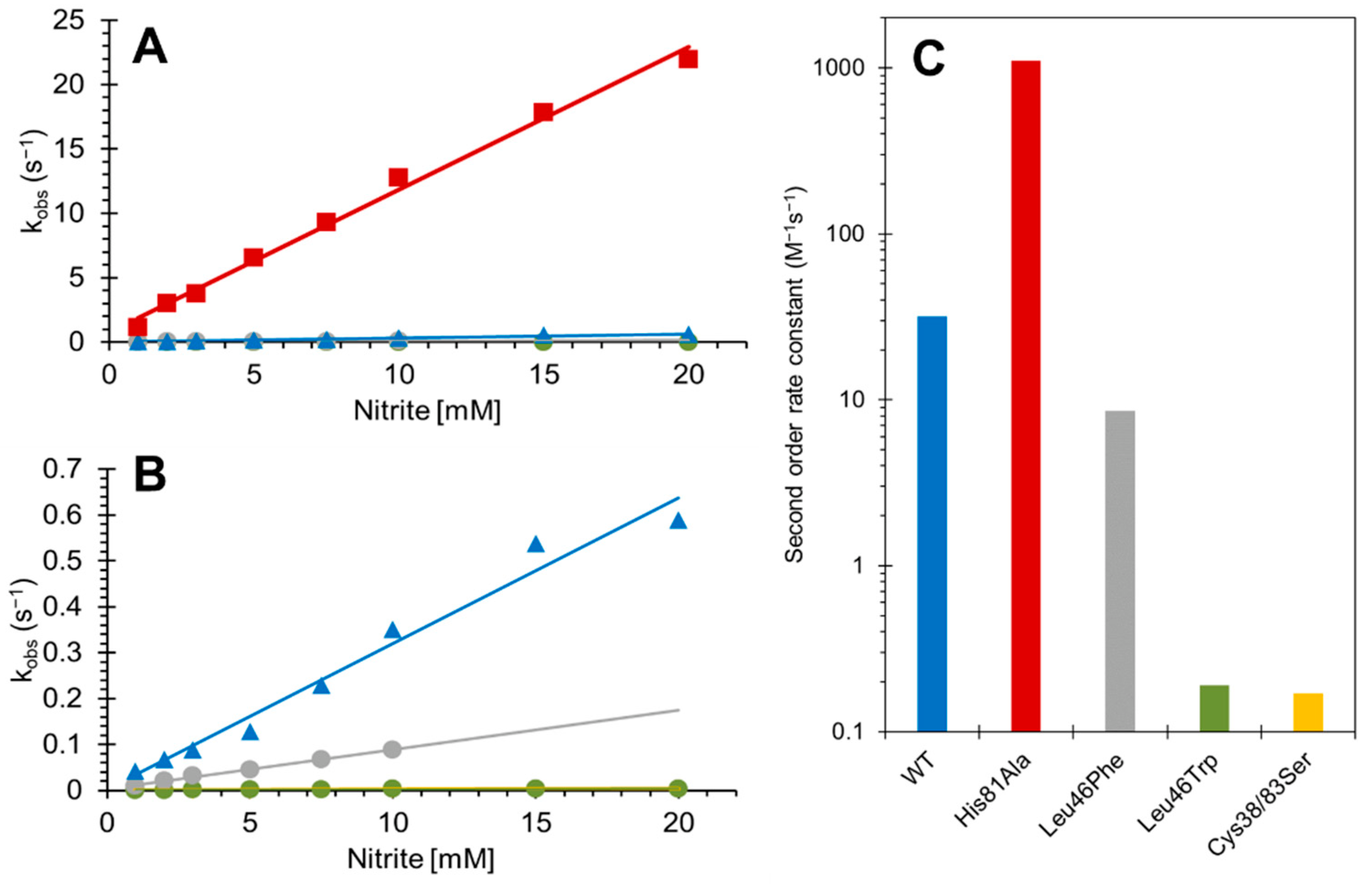
3.2. Effect of Mutations on the Nitrite Reductase Activity of Cytoglobin
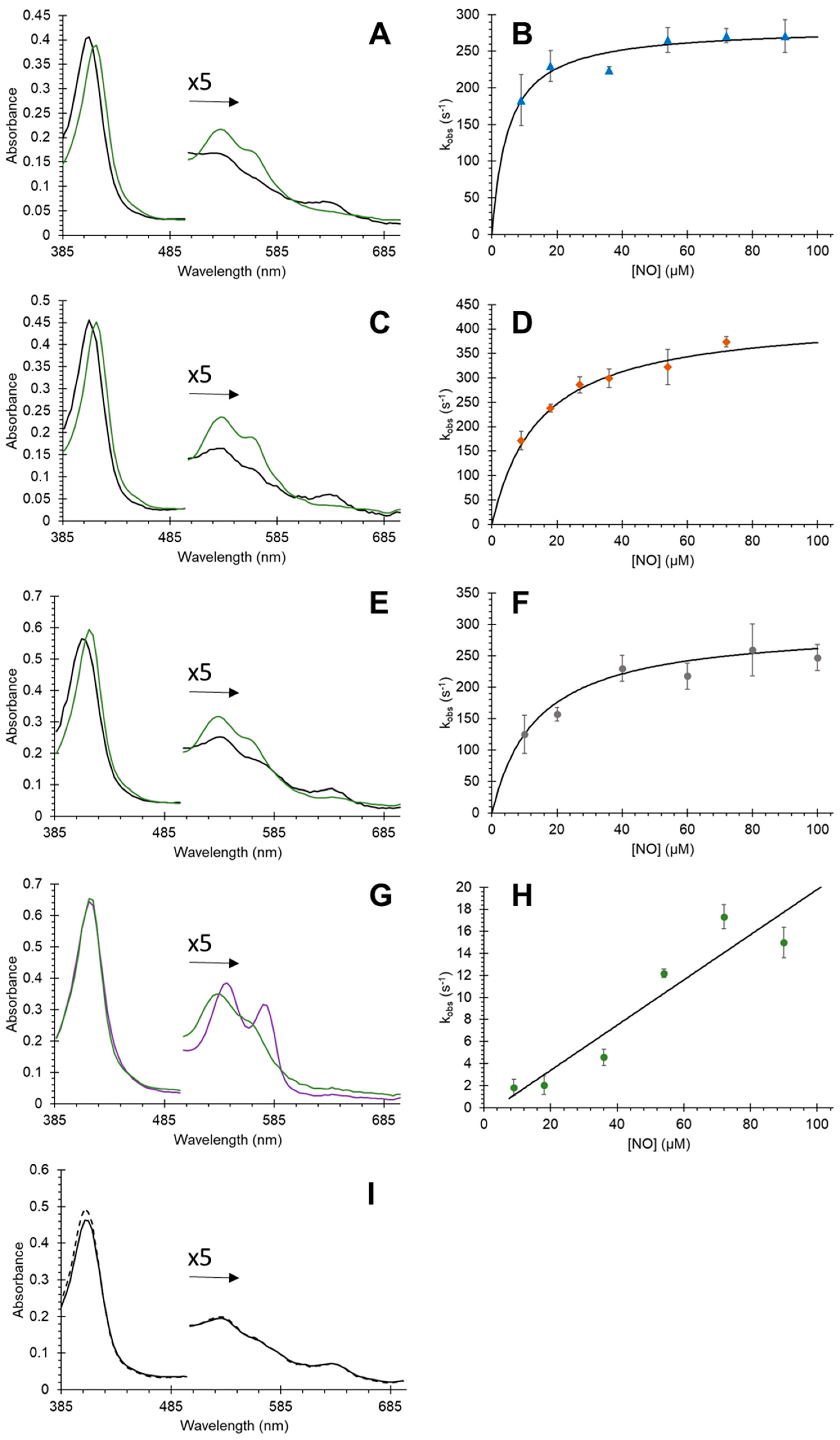
3.3. Effect of Mutations on Nitric Oxide Dioxygenase Activity of Cytoglobin
4. Discussion
5. Conclusions and Future Directions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Burmester, T.; Ebner, B.; Weich, B.; Hankeln, T. Cytoglobin: A novel globin type ubiquitously expressed in vertebrate tissues. Mol. Biol. Evol. 2002, 19, 416–421. [Google Scholar] [CrossRef]
- Hankeln, T.; Ebner, B.; Fuchs, C.; Gerlach, F.; Haberkamp, M.; Laufs, T.L.; Roesner, A.; Schmidt, M.; Weich, B.; Wystub, S.; et al. Neuroglobin and cytoglobin in search of their role in the vertebrate globin family. J Inorg. Biochem. 2005, 99, 110–119. [Google Scholar] [CrossRef]
- Liu, X.; Follmer, D.; Zweier, J.R.; Huang, X.; Hemann, C.; Liu, K.; Druhan, L.J.; Zweier, J.L. Characterization of the function of cytoglobin as an oxygen-dependent regulator of nitric oxide concentration. Biochemistry 2012, 51, 5072–5082. [Google Scholar] [CrossRef]
- Reeder, B.J.; Ukeri, J. Strong modulation of nitrite reductase activity of cytoglobin by disulfide bond oxidation: Implications for nitric oxide homeostasis. Nitric. Oxide 2018, 72, 16–23. [Google Scholar] [CrossRef]
- McRonald, F.E.; Risk, J.M.; Hodges, N.J. Protection from intracellular oxidative stress by cytoglobin in normal and cancerous oesophageal cells. PLoS ONE 2012, 7, e30587. [Google Scholar] [CrossRef]
- Ostojic, J.; Grozdanic, S.D.; Syed, N.A.; Hargrove, M.S.; Trent, J.T., 3rd; Kuehn, M.H.; Kwon, Y.H.; Kardon, R.H.; Sakaguchi, D.S. Patterns of distribution of oxygen-binding globins, neuroglobin and cytoglobin in human retina. Arch. Ophthalmol. 2008, 126, 1530–1536. [Google Scholar] [CrossRef]
- Ascenzi, P.; Marino, M.; Polticelli, F.; Coletta, M.; Gioia, M.; Marini, S.; Pesce, A.; Nardini, M.; Bolognesi, M.; Reeder, B.J.; et al. Non-covalent and covalent modifications modulate the reactivity of monomeric mammalian globins. Biochim. Biophys. Acta 2013, 1834, 1750–1756. [Google Scholar] [CrossRef]
- Reeder, B.J.; Svistunenko, D.A.; Wilson, M.T. Lipid binding to cytoglobin leads to a change in haem co-ordination: A role for cytoglobin in lipid signalling of oxidative stress. Biochem. J. 2011, 434, 483–492. [Google Scholar] [CrossRef]
- Tejero, J.; Kapralov, A.A.; Baumgartner, M.P.; Sparacino-Watkins, C.E.; Anthonymutu, T.S.; Vlasova, I.I.; Camacho, C.J.; Gladwin, M.T.; Bayir, H.; Kagan, V.E. Peroxidase activation of cytoglobin by anionic phospholipids: Mechanisms and consequences. Biochim. Biophys. Acta 2016, 1861, 391–401. [Google Scholar] [CrossRef]
- Li, H.; Hemann, C.; Abdelghany, T.M.; El-Mahdy, M.A.; Zweier, J.L. Characterization of the mechanism and magnitude of cytoglobin-mediated nitrite reduction and nitric oxide generation under anaerobic conditions. J. Biol. Chem. 2012, 287, 36623–36633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Avivi, A.; Gerlach, F.; Joel, A.; Reuss, S.; Burmester, T.; Nevo, E.; Hankeln, T. Neuroglobin, cytoglobin, and myoglobin contribute to hypoxia adaptation of the subterranean mole rat Spalax. Proc. Natl. Acad. Sci. USA 2010, 107, 21570–21575. [Google Scholar] [CrossRef] [PubMed]
- Emara, M.; Turner, A.R.; Allalunis-Turner, J. Hypoxic regulation of cytoglobin and neuroglobin expression in human normal and tumor tissues. Cancer Cell Int. 2010, 10, 33. [Google Scholar] [CrossRef]
- Shaw, R.J.; Omar, M.M.; Rokadiya, S.; Kogera, F.A.; Lowe, D.; Hall, G.L.; Woolgar, J.A.; Homer, J.; Liloglou, T.; Field, J.K.; et al. Cytoglobin is upregulated by tumour hypoxia and silenced by promoter hypermethylation in head and neck cancer. Br. J. Cancer 2009, 101, 139–144. [Google Scholar] [CrossRef] [PubMed]
- Yoshizato, K.; Thuyle, T.T.; Shiota, G.; Kawada, N. Discovery of cytoglobin and its roles in physiology and pathology of hepatic stellate cells. Proc. Jpn. Acad. Ser. B Phys. Biol. Sci. 2016, 92, 77–97. [Google Scholar] [CrossRef] [PubMed]
- Fang, J.; Ma, I.; Allalunis-Turner, J. Knockdown of cytoglobin expression sensitizes human glioma cells to radiation and oxidative stress. Radiat. Res. 2011, 176, 198–207. [Google Scholar] [CrossRef]
- Halligan, K.E.; Jourd’heuil, F.L.; Jourd’heuil, D. Cytoglobin is expressed in the vasculature and regulates cell respiration and proliferation via nitric oxide dioxygenation. J. Biol. Chem. 2009, 284, 8539–8547. [Google Scholar] [CrossRef]
- Liu, X.; Tong, J.; Zweier, J.R.; Follmer, D.; Hemann, C.; Ismail, R.S.; Zweier, J.L. Differences in oxygen-dependent nitric oxide metabolism by cytoglobin and myoglobin account for their differing functional roles. FEBS J. 2013, 280, 3621–3631. [Google Scholar] [CrossRef]
- Lilly, B.; Dammeyer, K.; Marosis, S.; McCallinhart, P.E.; Trask, A.J.; Lowe, M.; Sawant, D. Endothelial cell-induced cytoglobin expression in vascular smooth muscle cells contributes to modulation of nitric oxide. Vasc. Pharmacol. 2018, 110, 7–15. [Google Scholar] [CrossRef]
- Liu, X.; El-Mahdy, M.A.; Boslett, J.; Varadharaj, S.; Hemann, C.; Abdelghany, T.M.; Ismail, R.S.; Little, S.C.; Zhou, D.; Thuy, L.T.; et al. Cytoglobin regulates blood pressure and vascular tone through nitric oxide metabolism in the vascular wall. Nat. Commun. 2017, 8, 14807. [Google Scholar] [CrossRef]
- Reuss, S.; Wystub, S.; Disque-Kaiser, U.; Hankeln, T.; Burmester, T. Distribution of Cytoglobin in the Mouse Brain. Front. Neuroanat. 2016, 10, 47. [Google Scholar] [CrossRef] [Green Version]
- Van Thuy, T.T.; Thuy, L.T.; Yoshizato, K.; Kawada, N. Possible Involvement of Nitric Oxide in Enhanced Liver Injury and Fibrogenesis during Cholestasis in Cytoglobin-deficient Mice. Sci. Rep. 2017, 7, 41888. [Google Scholar] [CrossRef] [PubMed]
- Gardner, A.M.; Cook, M.R.; Gardner, P.R. Nitric-oxide dioxygenase function of human cytoglobin with cellular reductants and in rat hepatocytes. J. Biol. Chem. 2010, 285, 23850–23857. [Google Scholar] [CrossRef] [PubMed]
- Petersen, M.G.; Dewilde, S.; Fago, A. Reactions of ferrous neuroglobin and cytoglobin with nitrite under anaerobic conditions. J. Inorg. Biochem. 2008, 102, 1777–1782. [Google Scholar] [CrossRef] [PubMed]
- Ilangovan, G.; Khaleel, S.A.; Kundu, T.; Hemann, C.; El-Mahdy, M.A.; Zweier, J.L. Defining the reducing system of the NO dioxygenase cytoglobin in vascular smooth muscle cells and its critical role in regulating cellular NO decay. J. Biol. Chem. 2021, 296, 100196. [Google Scholar] [CrossRef]
- Mahgoup, E.M.; Khaleel, S.A.; El-Mahdy, M.A.; Abd-Allah, A.R.; Zweier, J.L. Role of cytoglobin in cigarette smoke constituent-induced loss of nitric oxide bioavailability in vascular smooth muscle cells. Nitric Oxide 2022, 119, 9–18. [Google Scholar] [CrossRef] [PubMed]
- Astudillo, L.; Bernad, S.; Derrien, V.; Sebban, P.; Miksovska, J. Reduction of the internal disulfide bond between Cys 38 and 83 switches the ligand migration pathway in cytoglobin. J. Inorg. Biochem. 2013, 129, 23–29. [Google Scholar] [CrossRef]
- Lechauve, C.; Chauvierre, C.; Dewilde, S.; Moens, L.; Green, B.N.; Marden, M.C.; Celier, C.; Kiger, L. Cytoglobin conformations and disulfide bond formation. FEBS J. 2010, 277, 2696–2704. [Google Scholar] [CrossRef] [PubMed]
- Tsujino, H.; Yamashita, T.; Nose, A.; Kukino, K.; Sawai, H.; Shiro, Y.; Uno, T. Disulfide bonds regulate binding of exogenous ligand to human cytoglobin. J. Inorg. Biochem. 2014, 135, 20–27. [Google Scholar] [CrossRef]
- Beckerson, P.; Reeder, B.J.; Wilson, M.T. Coupling of disulfide bond and distal histidine dissociation in human ferrous cytoglobin regulates ligand binding. FEBS Lett. 2015, 589, 507–512. [Google Scholar] [CrossRef]
- Beckerson, P.; Svistunenko, D.; Reeder, B. Effect of the distal histidine on the peroxidatic activity of monomeric cytoglobin. F1000Res 2015, 4, 87. [Google Scholar] [CrossRef] [Green Version]
- Beckerson, P.; Wilson, M.T.; Svistunenko, D.A.; Reeder, B.J. Cytoglobin ligand binding regulated by changing haem-co-ordination in response to intramolecular disulfide bond formation and lipid interaction. Biochem. J. 2015, 465, 127–137. [Google Scholar] [CrossRef] [PubMed]
- Tejero, J.; Sparacino-Watkins, C.E.; Ragireddy, V.; Frizzell, S.; Gladwin, M.T. Exploring the mechanisms of the reductase activity of neuroglobin by site-directed mutagenesis of the heme distal pocket. Biochemistry 2015, 54, 722–733. [Google Scholar] [CrossRef] [PubMed]
- Varnado, C.L.; Mollan, T.L.; Birukou, I.; Smith, B.J.; Henderson, D.P.; Olson, J.S. Development of recombinant hemoglobin-based oxygen carriers. Antioxid Redox Signal. 2013, 18, 2314–2328. [Google Scholar] [CrossRef] [PubMed]
- Kaliszuk, S.J.; Morgan, N.I.; Ayers, T.N.; Sparacino-Watkins, C.E.; DeMartino, A.W.; Bocian, K.; Ragireddy, V.; Tong, Q.; Tejero, J. Regulation of nitrite reductase and lipid binding properties of cytoglobin by surface and distal histidine mutations. Nitric Oxide 2022, 125–126, 12–22. [Google Scholar] [CrossRef] [PubMed]
- Birukou, I.; Maillett, D.H.; Birukova, A.; Olson, J.S. Modulating distal cavities in the alpha and beta subunits of human HbA reveals the primary ligand migration pathway. Biochemistry 2011, 50, 7361–7374. [Google Scholar] [CrossRef]
- Draghi, F.; Miele, A.E.; Travaglini-Allocatelli, C.; Vallone, B.; Brunori, M.; Gibson, Q.H.; Olson, J.S. Controlling ligand binding in myoglobin by mutagenesis. J. Biol. Chem. 2002, 277, 7509–7519. [Google Scholar] [CrossRef]
- Makino, M.; Sugimoto, H.; Sawai, H.; Kawada, N.; Yoshizato, K.; Shiro, Y. High-resolution structure of human cytoglobin: Identification of extra N- and C-termini and a new dimerization mode. Acta Crystallogr. D Biol. Crystallogr. 2006, 62, 671–677. [Google Scholar] [CrossRef]
- Simons, M.; Gretton, S.; Silkstone, G.G.A.; Rajagopal, B.S.; Allen-Baume, V.; Syrett, N.; Shaik, T.; Leiva-Eriksson, N.; Ronda, L.; Mozzarelli, A.; et al. Comparison of the oxidative reactivity of recombinant fetal and adult human hemoglobin: Implications for the design of hemoglobin-based oxygen carriers. Biosci. Rep. 2018, 38, BSR20180370. [Google Scholar] [CrossRef]
- Antonini, E.; Brunori, M. Hemoglobin and myoglobin and their reaction with ligands. In Frontiers in Biology; Neuberger, A., Tatum, E.L., Eds.; North-Holland Publishing Company: Amsterdam, The Netherlands; London, UK, 1971; Volume 21, pp. 13–52. [Google Scholar]
- Shiva, S.; Huang, Z.; Grubina, R.; Sun, J.; Ringwood, L.A.; MacArthur, P.H.; Xu, X.; Murphy, E.; Darley-Usmar, V.M.; Gladwin, M.T. Deoxymyoglobin is a nitrite reductase that generates nitric oxide and regulates mitochondrial respiration. Circ. Res. 2007, 100, 654–661. [Google Scholar] [CrossRef]
- Mansy, S.S.; Olson, J.S.; Gonzalez, G.; Gilles-Gonzalez, M.A. Imidazole is a sensitive probe of steric hindrance in the distal pockets of oxygen-binding heme proteins. Biochemistry 1998, 37, 12452–12457. [Google Scholar] [CrossRef]
- Eich, R.F.; Li, T.; Lemon, D.D.; Doherty, D.H.; Curry, S.R.; Aitken, J.F.; Mathews, A.J.; Johnson, K.A.; Smith, R.D.; Phillips, G.N., Jr.; et al. Mechanism of NO-induced oxidation of myoglobin and hemoglobin. Biochemistry 1996, 35, 6976–6983. [Google Scholar] [CrossRef] [PubMed]
- Scott, E.E.; Gibson, Q.H.; Olson, J.S. Mapping the pathways for O2 entry into and exit from myoglobin. J. Biol. Chem. 2001, 276, 5177–5188. [Google Scholar] [CrossRef] [PubMed]
- Brunori, M.; Giuffre, A.; Nienhaus, K.; Nienhaus, G.U.; Scandurra, F.M.; Vallone, B. Neuroglobin, nitric oxide, and oxygen: Functional pathways and conformational changes. Proc. Natl. Acad. Sci. USA 2005, 102, 8483–8488. [Google Scholar] [CrossRef]
- Choudhari, S.K.; Chaudhary, M.; Bagde, S.; Gadbail, A.R.; Joshi, V. Nitric oxide and cancer: A review. World J. Surg. Oncol. 2013, 11, 118. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mutation | NO Binding | NiR (M−1 s−1) | NOD (M−1 s−1) | NOD Spectra Relaxation (s−1) |
|---|---|---|---|---|
| Wild Type (intramolecular disulfide) | 11.7 (s−1) | 31.7 | >1 × 108 | 284 |
| Cys38Ser/Cys83Ser | 0.29 (s−1) | 0.17 | >1 × 108 | 427 |
| His81Ala | 7.0 × 106 (M−1 s−1) | 1108 | >1 × 108 | ND |
| Leu46Phe | 13.3 (s−1) | 8.54 | >1 × 108 | 298 |
| Leu46Trp | 4.0 × 105 (M−1 s−1) | 0.19 | 2.1 × 103 | ND |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ukeri, J.; Wilson, M.T.; Reeder, B.J. Modulating Nitric Oxide Dioxygenase and Nitrite Reductase of Cytoglobin through Point Mutations. Antioxidants 2022, 11, 1816. https://doi.org/10.3390/antiox11091816
Ukeri J, Wilson MT, Reeder BJ. Modulating Nitric Oxide Dioxygenase and Nitrite Reductase of Cytoglobin through Point Mutations. Antioxidants. 2022; 11(9):1816. https://doi.org/10.3390/antiox11091816
Chicago/Turabian StyleUkeri, John, Michael T. Wilson, and Brandon J. Reeder. 2022. "Modulating Nitric Oxide Dioxygenase and Nitrite Reductase of Cytoglobin through Point Mutations" Antioxidants 11, no. 9: 1816. https://doi.org/10.3390/antiox11091816