
Oxidative Implications of Substituting a Conserved Cysteine Residue in Sugar Beet Phytoglobin BvPgb 1.2

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Recombinant Protein Production
2.2. Cultivation of Protein-Expressing Cells
2.3. Protein Purification
2.4. Protein Crystallization
2.5. Size-Exclusion Chromatography
2.6. Autoxidation Kinetics
2.7. Thermal Stability
2.8. Heme-Loss Assay
2.9. Peroxidase Activity Measurements
3. Results
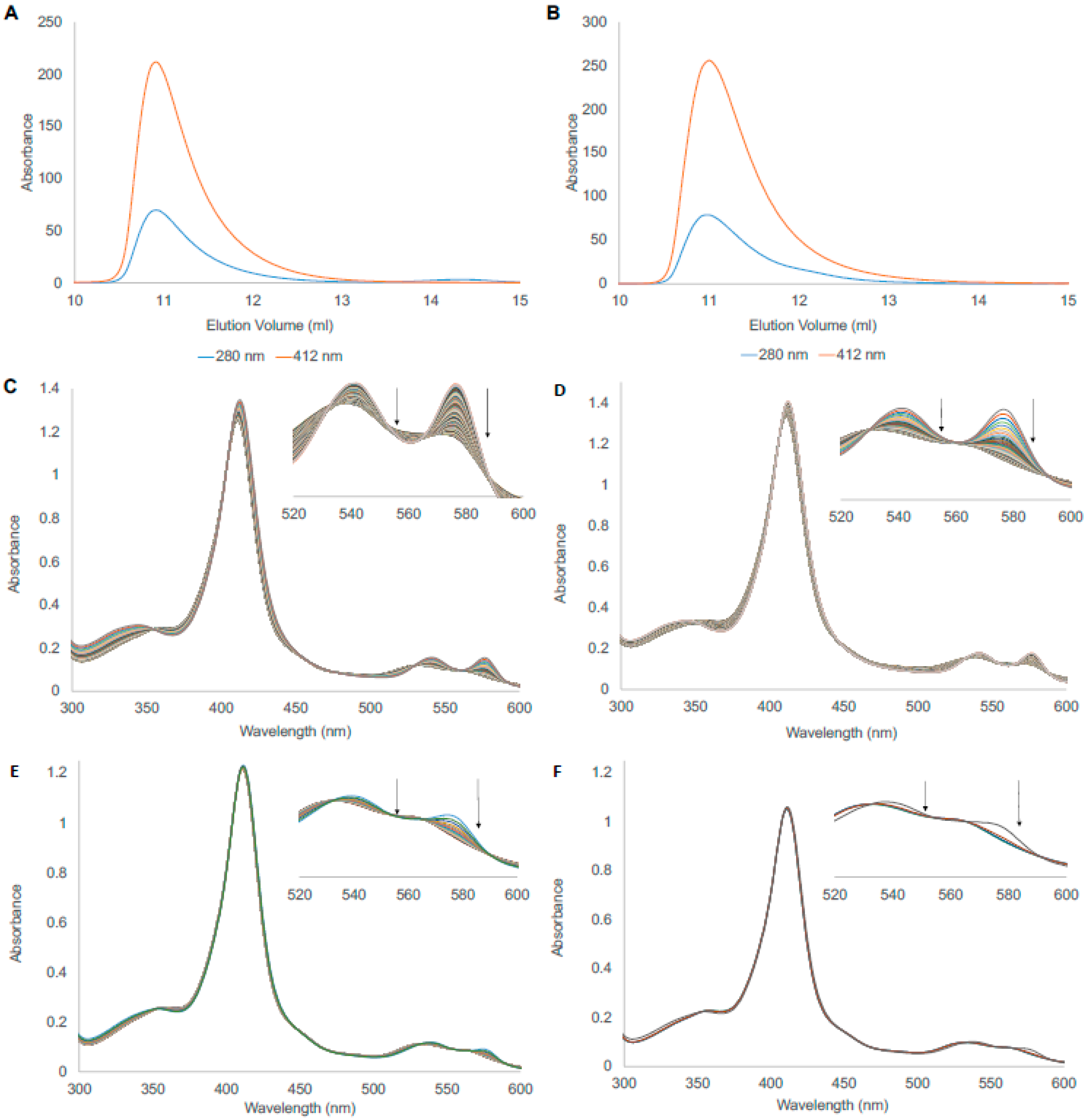
3.1. Protein Expression and Purification
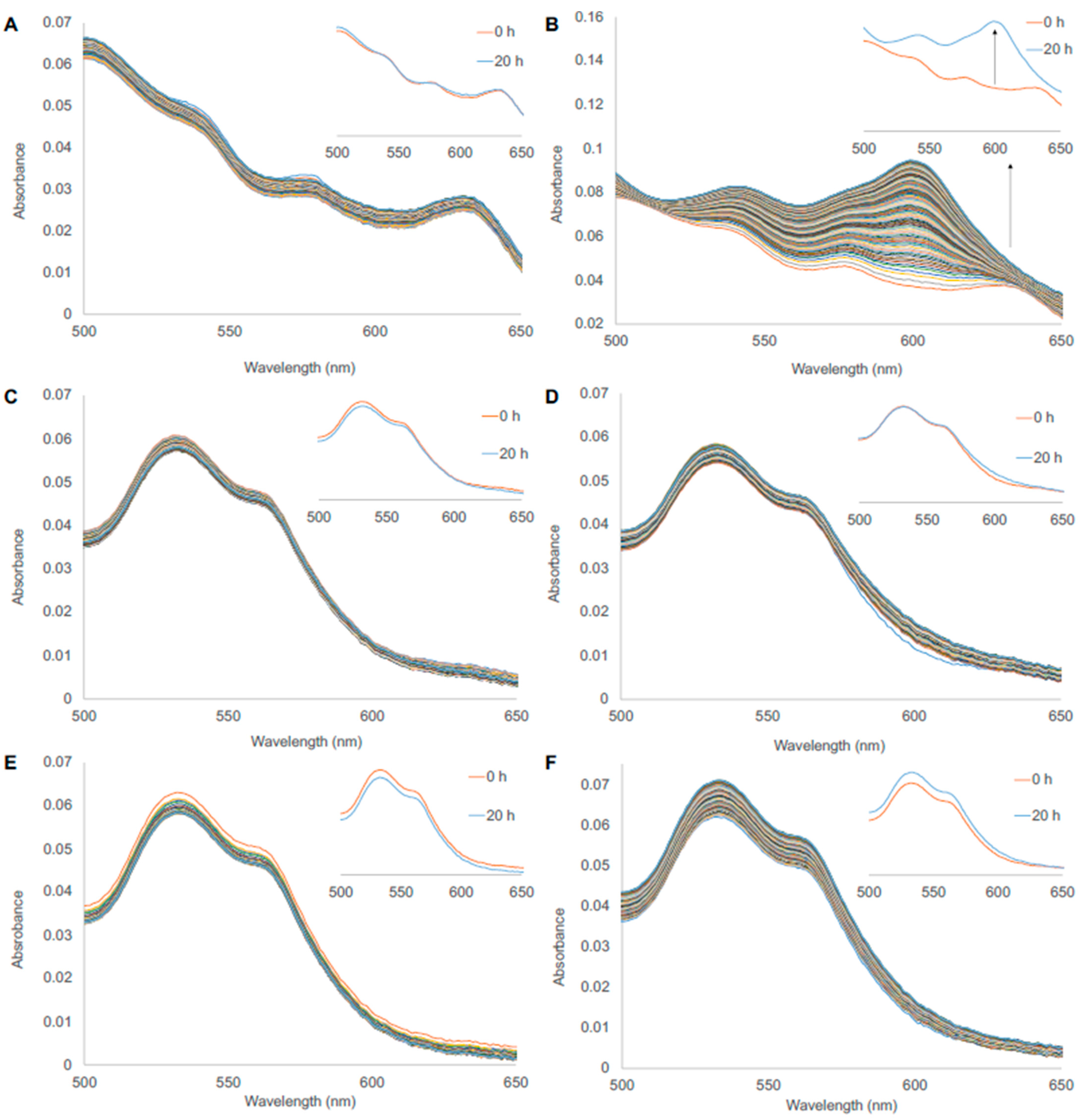
3.2. Autoxidation
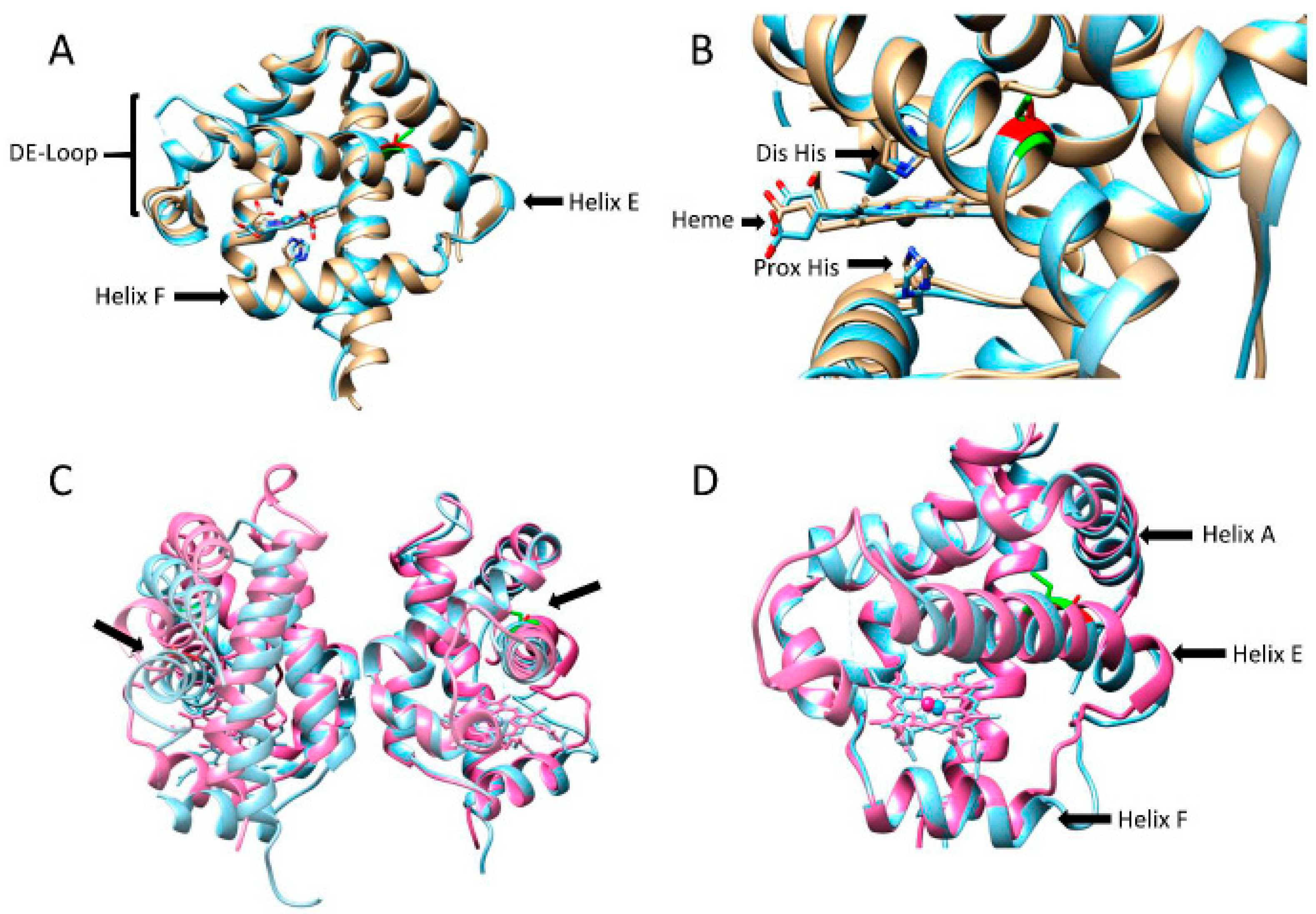
3.3. Structural Comparison
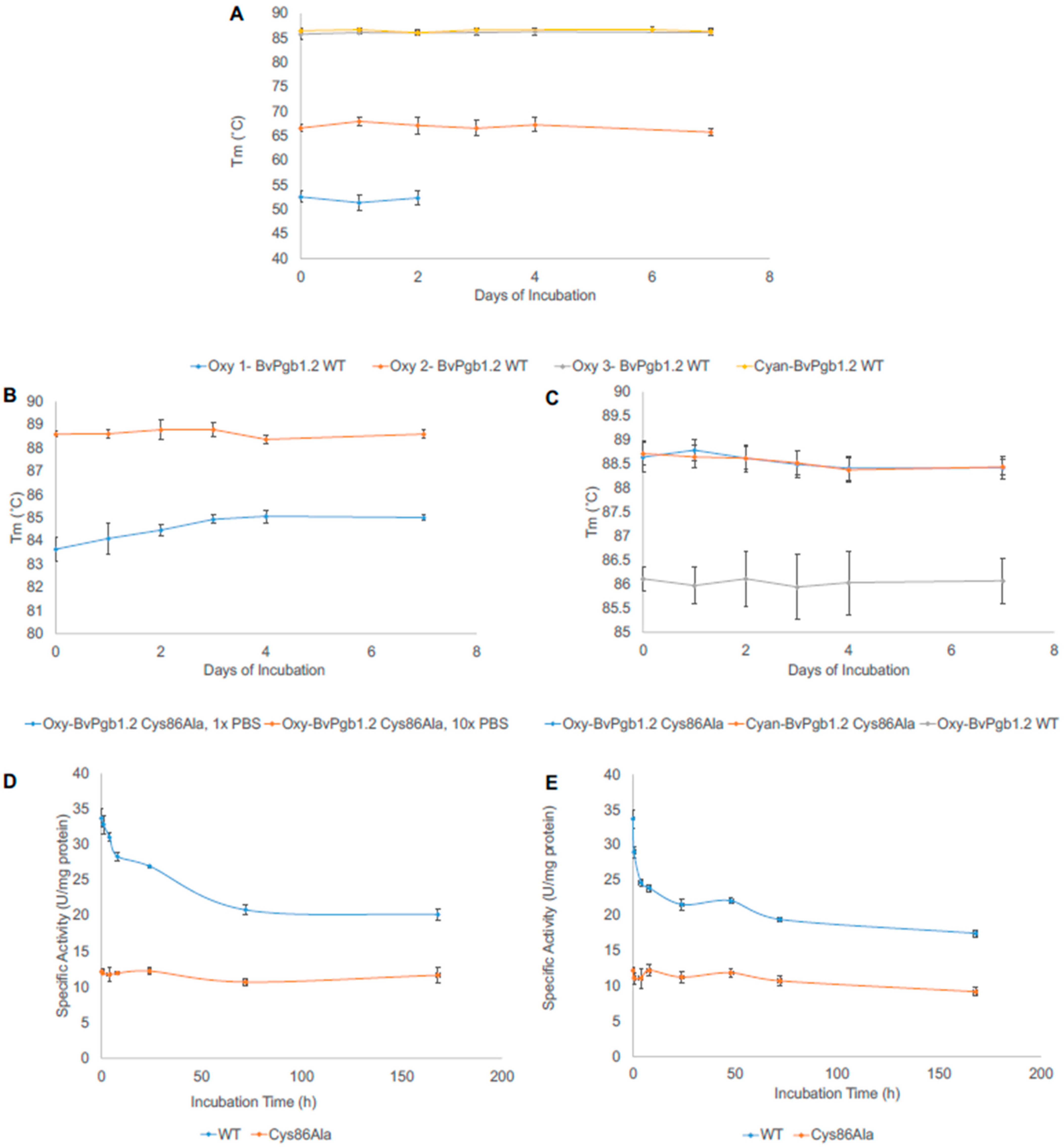
3.4. Thermal Stability of BvHb1.2 WT and Cys86Ala
3.5. Peroxidase Activity
3.6. Heme Loss from WT and Cys86Ala
4. Discussion
5. Conclusions
6. Patents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tejero, J.; Gladwin, M.T. The globin superfamily: Functions in nitric oxide formation and decay. Biol. Chem. 2014, 395, 631–639. [Google Scholar] [PubMed] [Green Version]
- Hardison, R. Hemoglobins from bacteria to man: Evolution of different patterns of gene expression. J. Exp. Biol. 1998, 201, 1099–1117. [Google Scholar] [CrossRef] [PubMed]
- Vázquez-Limón, C.; Hoogewijs, D.; Vinogradov, S.N.; Arredondo-Peter, R. The evolution of land plant hemoglobins. Plant Sci. 2012, 191–192, 71–81. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, M.H.; Ghatge, M.S.; Safo, M.K. Hemoglobin: Structure, Function and Allostery. Sub-Cell. Biochem. 2020, 94, 345–382. [Google Scholar]
- Olson, J.; Mathews, A.J.; Rohlfs, R.J.; Springer, B.A.; Egeberg, K.D.; Sligar, S.G.; Tame, J.; Renaud, J.-P.; Nagai, K. The role of the distal histidine in myoglobin and haemoglobin. Nature 1988, 336, 265–266. [Google Scholar]
- Smagghe, B.J.; Hoy, J.A.; Percifield, R.; Kundu, S.; Hargrove, M.S.; Sarath, G.; Hilbert, J.-L.; Watts, R.A.; Dennis, E.S.; Peacock, W.J.; et al. Review: Correlations between oxygen affinity and sequence classifications of plant hemoglobins. Biopolymers 2009, 91, 1083–1096. [Google Scholar] [PubMed]
- Kakar, S.; Hoffman, F.G.; Storz, J.F.; Fabian, M.; Hargrove, M.S. Structure and reactivity of hexacoordinate hemoglobins. Biophys. Chem. 2010, 152, 1–14. [Google Scholar]
- Becana, M.; Yruela, I.; Sarath, G.; Catalán, P.; Hargrove, M.S. Plant hemoglobins: A journey from unicellular green algae to vascular plants. New Phytol. 2020, 227, 1618–1635. [Google Scholar]
- Hill, R.; Hargrove, M.; Arredondo-Peter, R. Phytoglobin: A novel nomenclature for plant globins accepted by the globin community at the 2014 XVIII conference on Oxygen-Binding and Sensing Proteins. F1000Research 2016, 5, 212. [Google Scholar]
- Birukou, I.; Schweers, R.L.; Olson, J.S. Distal Histidine Stabilizes Bound O2 and Acts as a Gate for Ligand Entry in Both Subunits of Adult Human Hemoglobin. J. Biol. Chem. 2010, 285, 8840–8854. [Google Scholar]
- Arredondo-Peter, R.; Hargrove, M.S.; Sarath, G.; Moran, J.F.; Lohrman, J.; Olson, J.S.; Klucas, R.V. Rice Hemoglobins (Gene Cloning, Analysis, and O2-Binding Kinetics of a Recombinant Protein Synthesized in Escherichia coli). Plant Physiol. 1997, 115, 1259–1266. [Google Scholar]
- Gupta, K.J.; Hebelstrup, K.H.; Mur, L.A.; Igamberdiev, A.U. Plant hemoglobins: Important players at the crossroads between oxygen and nitric oxide. FEBS Lett. 2011, 585, 3843–3849. [Google Scholar]
- Garrocho-Villegas, V.; Gopalasubramaniam, S.K.; Arredondo-Peter, R. Plant hemoglobins: What we know six decades after their discovery. Gene 2007, 398, 78–85. [Google Scholar]
- Hoy, J.A.; Hargrove, M.S. The structure and function of plant hemoglobins. Plant Physiol. Biochem. 2008, 46, 371–379. [Google Scholar]
- Perazzolli, M.; Dominici, P.; Romero-Puertas, M.C.; Zago, E.; Zeier, J.; Sonoda, M.; Lamb, C.; Delledonne, M. Arabidopsis Nonsymbiotic Hemoglobin AHb1 Modulates Nitric Oxide Bioactivity. Plant Cell 2004, 16, 2785–2794. [Google Scholar]
- Mot, A.C.; Puscas, C.; Miclea, P.; Naumova-Letia, G.; Dorneanu, S.; Podar, D.; Dissmeyer, N.; Silaghi-Dumitrescu, R. Redox control and autoxidation of class 1, 2 and 3 phytoglobins from Arabidopsis thaliana. Sci. Rep. 2018, 8, 13714. [Google Scholar]
- Leiva-Eriksson, N.; Pin, P.A.; Kraft, T.; Dohm, J.C.; Minoche, A.E.; Himmelbauer, H.; Bülow, L. Differential Expression Patterns of Non-Symbiotic Hemoglobins in Sugar Beet (Beta vulgaris ssp. vulgaris). Plant Cell Physiol. 2014, 55, 834–844. [Google Scholar]
- Eriksson, N.L.; Reeder, B.J.; Wilson, M.T.; Bülow, L. Sugar beet hemoglobins: Reactions with nitric oxide and nitrite reveal differential roles for nitrogen metabolism. Biochem. J. 2019, 476, 2111–2125. [Google Scholar]
- Watts, R.A.; Hunt, P.W.; Hvitved, A.N.; Hargrove, M.S.; Peacock, W.J.; Dennis, E.S. A hemoglobin from plants homologous to truncated hemoglobins of microorganisms. Proc. Natl. Acad. Sci. USA 2001, 98, 10119–10124. [Google Scholar]
- Bashford, D.; Chothia, C.; Lesk, A.M. Determinants of a protein fold: Unique features of the globin amino acid sequences. J. Mol. Biol. 1987, 196, 199–216. [Google Scholar]
- Ota, M.; Isogai, Y.; Nishikawa, K. Structural requirement of highly-conserved residues in globins. FEBS Lett. 1997, 415, 129–133. [Google Scholar] [CrossRef] [Green Version]
- Bykova, N.V.; Igamberdiev, A.U.; Ens, W.; Hill, R.D. Identification of an intermolecular disulfide bond in barley hemoglobin. Biochem. Biophys. Res. Commun. 2006, 347, 301–309. [Google Scholar] [CrossRef]
- Waterman, M.R. Role of cysteine residues in hemoglobin structure and function: Transfer of p-mercuribenzoate from alpha subunits to beta subunits during tetramer formation. Biochim. Biophys. Acta 1974, 371, 159–167. [Google Scholar] [CrossRef]
- Blacken, G.R.; Wang, Y.; Lopez, J.A.; Fu, X. Cysteine 93 of Hemoglobin Beta Chain Is the Major Target of Oxidation During Red Blood Cell Storage. Blood 2009, 114, 4040. [Google Scholar] [CrossRef]
- Hargrove, M.S.; Brucker, E.A.; Stec, B.; Sarath, G.; Arredondo-Peter, R.; Klucas, R.V.; Olson, J.S.; Phillips, G.N. Crystal structure of a nonsymbiotic plant hemoglobin. Structure 2000, 8, 1005–1014. [Google Scholar] [CrossRef] [Green Version]
- Igamberdiev, A.U.; Bykova, N.V.; Hill, R.D. Nitric oxide scavenging by barley hemoglobin is facilitated by a monodehydroascorbate reductase-mediated ascorbate reduction of methemoglobin. Planta 2005, 223, 1033–1040. [Google Scholar] [CrossRef]
- Vonrhein, C.; Flensburg, C.; Keller, P.; Sharff, A.; Smart, O.; Paciorek, W.; Womack, T.; Bricogne, G. Data processing and analysis with the autoPROC toolbox. Acta Crystallogr. Sect. D Biol. Crystallogr. 2011, 67, 293–302. [Google Scholar] [CrossRef] [Green Version]
- Adams, P.D.; Afonine, P.V.; Bunkóczi, G.; Chen, V.B.; Davis, I.W.; Echols, N.; Headd, J.J.; Hung, L.-W.; Kapral, G.J.; Grosse-Kunstleve, R.W.; et al. PHENIX: A comprehensive Python-based system for macromolecular structure solution. Acta Crystallogr. Sect. D Biol. Crystallogr. 2019, 66, 213–221. [Google Scholar] [CrossRef] [Green Version]
- Emsley, P.; Lohkamp, B.; Scott, W.G.; Cowtan, K. Features and development of Coot. Acta Crystallogr. Sect. D Biol. Crystallogr. 2010, 66, 486–501. [Google Scholar] [CrossRef] [Green Version]
- Hargrove, M.S.; Singleton, E.W.; Quillin, M.L.; Ortiz, A.L.; Phillips, G.N.; Olson, J.S.; Mathews, A.J. His64(E7)-->Tyr apomyoglobin as a reagent for measuring rates of hemin dissociation. J. Biol. Chem. 1994, 269, 4207–4214. [Google Scholar] [CrossRef]
- Silkstone, G.G.; Silkstone, R.S.; Wilson, M.T.; Simons, M.; Bülow, L.; Kallberg, K.; Ratanasopa, K.; Ronda, L.; Mozzarelli, A.; Reeder, B.J.; et al. Engineering tyrosine electron transfer pathways decreases oxidative toxicity in hemoglobin: Implications for blood substitute design. Biochem. J. 2016, 473, 3371–3383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kettisen, K.; Strader, M.B.; Wood, F.; Alayash, A.I.; Bülow, L. Site-directed mutagenesis of cysteine residues alters oxidative stability of fetal hemoglobin. Redox Biol. 2018, 19, 218–225. [Google Scholar] [CrossRef] [PubMed]
- Kvist, M.; Ryabova, E.S.; Nordlander, E.; Bülow, L. An investigation of the peroxidase activity of Vitreoscilla hemoglobin. JBIC J. Biol. Inorg. Chem. 2007, 12, 324–334. [Google Scholar] [CrossRef] [PubMed]
- Igamberdiev, A.U.; Bykova, N.V.; Hill, R.D. Structural and functional properties of class 1 plant hemoglobins. IUBMB Life 2011, 63, 146–152. [Google Scholar] [CrossRef]
- Everse, J.; Johnson, M.C.; Marini, M.A. Peroxidative activities of hemoglobin and hemoglobin derivatives. Methods Enzymol. 1994, 231, 547–561. [Google Scholar]
- Postnikova, G.B.; Komarov, Y.E.; Yumakova, E.M. Fluorescence study of the conformational properties of myoglobin structure. 2. pH- and ligand-induced conformational changes in ferric- and ferrousmyoglobins. JBIC J. Biol. Inorg. Chem. 1991, 198, 233–239. [Google Scholar] [CrossRef]
- Cooper, C.E.; Schaer, D.J.; Buehler, P.W.; Wilson, M.T.; Reeder, B.J.; Silkstone, G.; Svistunenko, D.A.; Bulow, L.; Alayash, A.I. Haptoglobin binding stabilizes hemoglobin ferryl iron and the globin radical on tyrosine β145. Antioxid. Redox Signal. 2013, 18, 2264–2273. [Google Scholar] [CrossRef]
- Moser, C.C.; Keske, J.M.; Warncke, K.; Farid, R.S.; Dutton, P.L. Nature of biological electron transfer. Nature 1992, 355, 796–802. [Google Scholar] [CrossRef]
- Silkstone, R.S.; Silkstone, G.; Baath, J.A.; Rajagopal, B.; Nicholls, P.; Reeder, B.J.; Ronda, L.; Bulow, L.; Cooper, C.E. The βLys66Tyr Variant of Human Hemoglobin as a Component of a Blood Substitute. Adv. Exp. Med. Biol. 2016, 876, 455–460. [Google Scholar]
- Zhang, L.; Levy, A.; Rifkind, J. Autoxidation of hemoglobin enhanced by dissociation into dimers. J. Biol. Chem. 1991, 266, 24698–24701. [Google Scholar] [CrossRef]
- Ku, T.; Lu, P.; Chan, C.; Wang, T.; Lai, S.; Lyu, P.; Hsiao, N. Predicting melting temperature directly from protein sequences. Comput. Biol. Chem. 2009, 33, 445–450. [Google Scholar] [CrossRef]
- Staniforth, R.A.; Giannini, S.; Bigotti, M.G.; Cutruzzolà, F.; Travaglini-Allocatelli, C.; Brunori, M. A new folding intermediate of apomyoglobin from Aplysia limacina: Stepwise formation of a molten globule. J. Mol. Biol. 2000, 297, 1231–1244. [Google Scholar] [CrossRef]
- Wan, L.; Twitchett, M.B.; Eltis, L.D.; Mauk, A.G.; Smith, M. In vitro evolution of horse heart myoglobin to increase peroxidase activity. Proc. Natl. Acad. Sci. USA 1998, 95, 12825–12831. [Google Scholar] [CrossRef] [Green Version]
- Hamdane, D.; Kiger, L.; Dewilde, S.; Uzan, J.; Burmester, T.; Hankeln, T.; Moens, L.; Marden, M.C. Hyperthermal stability of neuroglobin and cytoglobin. FEBS J. 2005, 272, 2076–2084. [Google Scholar] [CrossRef]
- Plana, L.J.; Nadra, A.D.; Estrin, D.A.; Luque, F.J.; Capece, L. Thermal Stability of Globins: Implications of Flexibility and Heme Coordination Studied by Molecular Dynamics Simulations. J. Chem. Inf. Model. 2018, 59, 441–452. [Google Scholar] [CrossRef]
- Huang, P.; Chu, S.K.S.; Frizzo, H.N.; Connolly, M.P.; Caster, R.W.; Siegel, J.B. Evaluating Protein Engineering Thermostability Prediction Tools Using an Independently Generated Dataset. ACS Omega 2020, 5, 6487–6493. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| N0 | λ (h−1) | τ (h) | t1/2 (h) | |
|---|---|---|---|---|
| WT 25 °C | 0.096 ± 0.002 | 0.014 ± 0.001 | 72.2 ± 0.180 | 50.1 + 0.125 |
| WT 37 °C | 0.082 ± 0.005 | 0.214 ± 0.003 | 4.68 + 0.069 | 3.24 + 0.005 |
| Cys86Ala 25 °C | 0.043 ± 0.005 | 0.234 ± 0.012 | 4.28 + 0.226 | 2.97 + 0.157 |
| Cys86Ala 37 °C | 0.035 ± 0.003 | 3.08 ± 0.035 | 0.342 + 0.004 | 0.225 + 0.003 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Christensen, S.; Groth, L.; Leiva-Eriksson, N.; Nyblom, M.; Bülow, L. Oxidative Implications of Substituting a Conserved Cysteine Residue in Sugar Beet Phytoglobin BvPgb 1.2. Antioxidants 2022, 11, 1615. https://doi.org/10.3390/antiox11081615
Christensen S, Groth L, Leiva-Eriksson N, Nyblom M, Bülow L. Oxidative Implications of Substituting a Conserved Cysteine Residue in Sugar Beet Phytoglobin BvPgb 1.2. Antioxidants. 2022; 11(8):1615. https://doi.org/10.3390/antiox11081615
Chicago/Turabian StyleChristensen, Simon, Leonard Groth, Nélida Leiva-Eriksson, Maria Nyblom, and Leif Bülow. 2022. "Oxidative Implications of Substituting a Conserved Cysteine Residue in Sugar Beet Phytoglobin BvPgb 1.2" Antioxidants 11, no. 8: 1615. https://doi.org/10.3390/antiox11081615