
Protective Effects of Wine Polyphenols on Oxidative Stress and Hepatotoxicity Induced by Acrylamide in Rats

 , ,
, ,  , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Wine Samples and Reagents
2.2. Experimental Animals
2.3. Experimental Design
2.4. Collection of Biological Material
2.5. Histopathological Evaluation of Liver Tissue
2.6. Evaluation of Liver Function
2.7. Preparation of Hepatic Tissue Homogenates
2.8. Protein Quantification in Liver Tissue Homogenates
2.9. Evaluation of Oxidative Stress by Means of Biomarkers Determined in Plasma
2.9.1. Determination of Lipid Peroxidation by Quantification of Malondialdehyde in Rat Plasma
2.9.2. Determination of Reduced and Total Glutathione Plasma Levels
2.10. Evaluation of Oxidative Stress in Liver Tissue
2.10.1. Determination of Lipid Peroxidation by Quantifying TBARS in Liver Tissue
2.10.2. Determination of Reduced Glutathione Levels in Liver Tissue
2.10.3. Determination of Antioxidant Enzymes in Liver Tissue
2.11. Statistical Analysis
3. Results
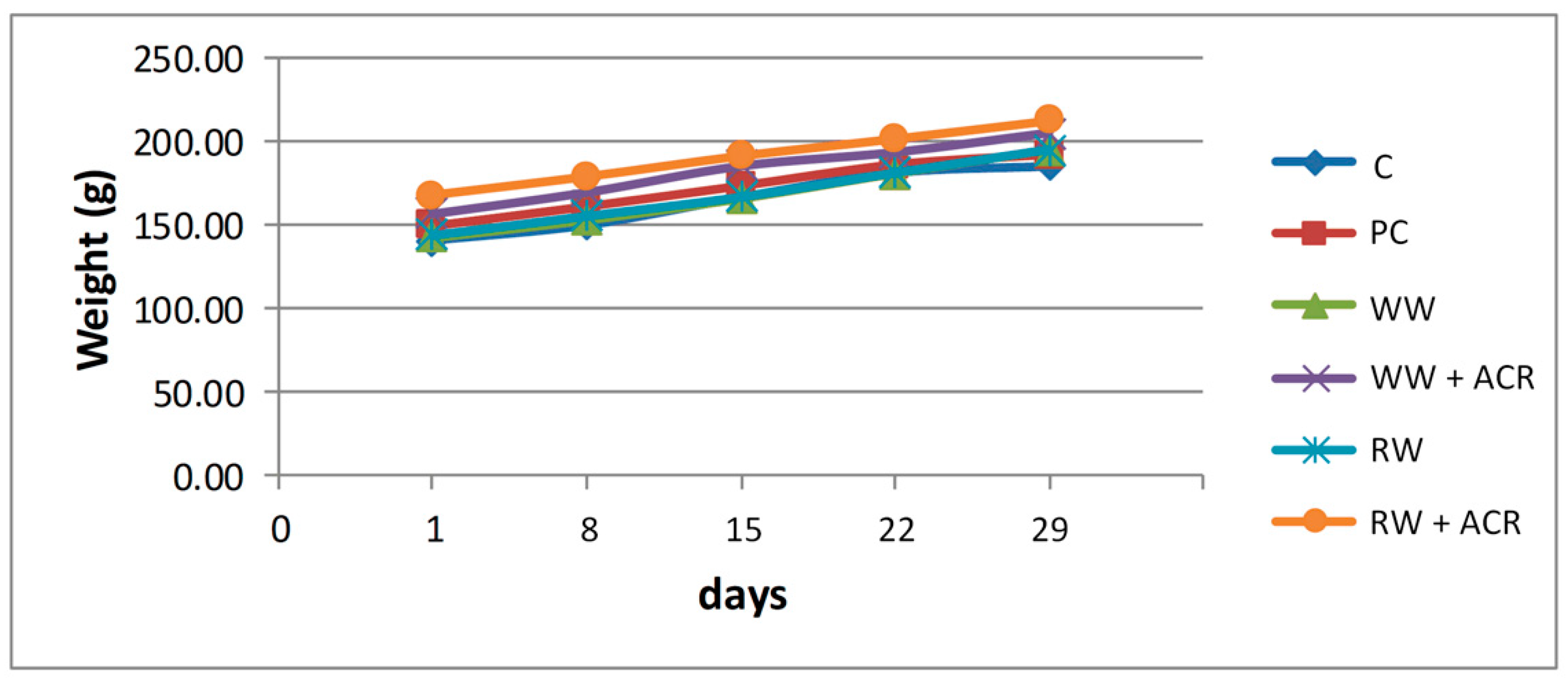
3.1. General Toxicity: Evolution of Animal Body Weight, Absolute and Relative Weight of the Liver
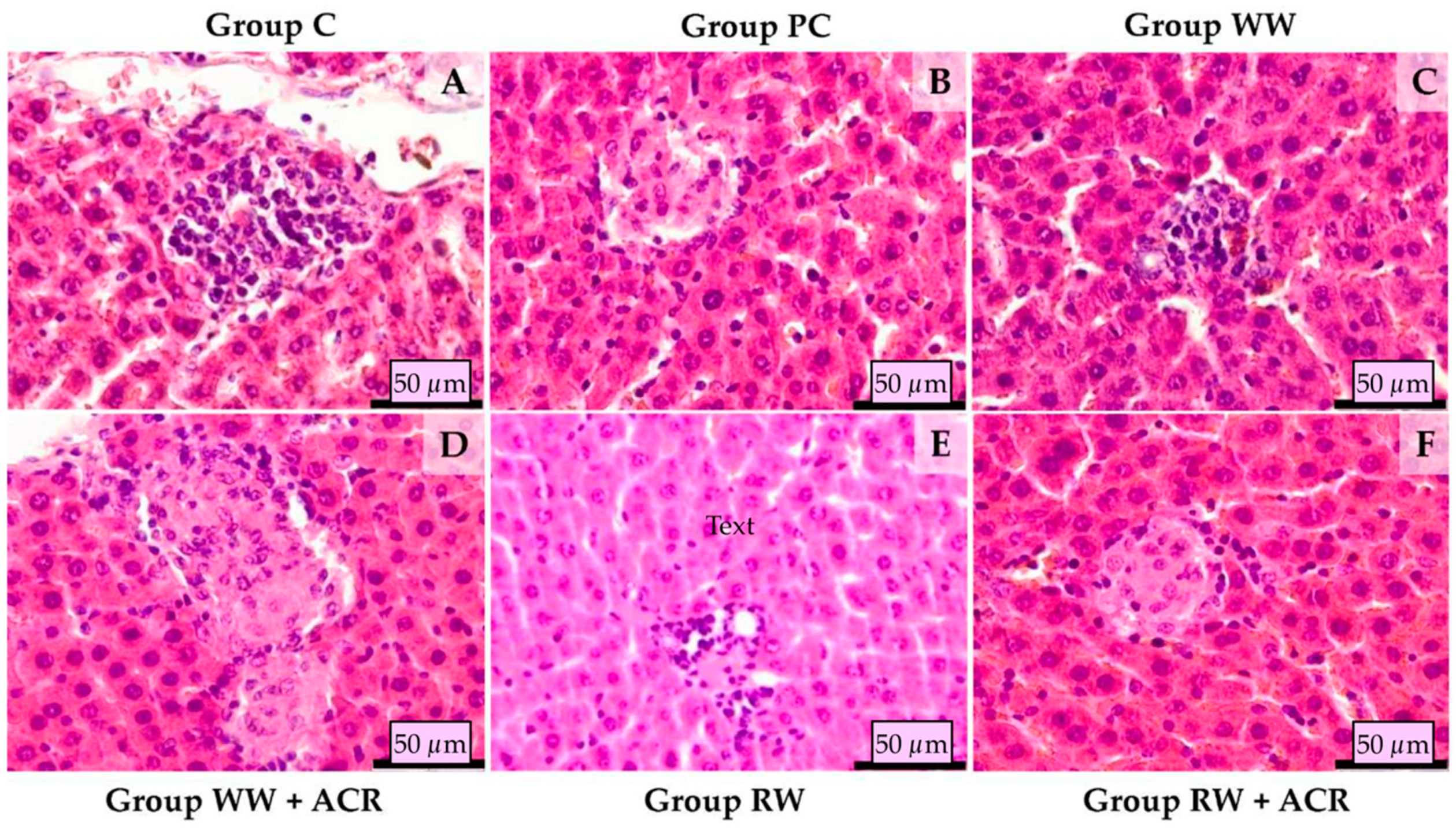
3.2. Histopathological Examination of Rat Liver
3.3. Biochemical Parameters of Liver Damage
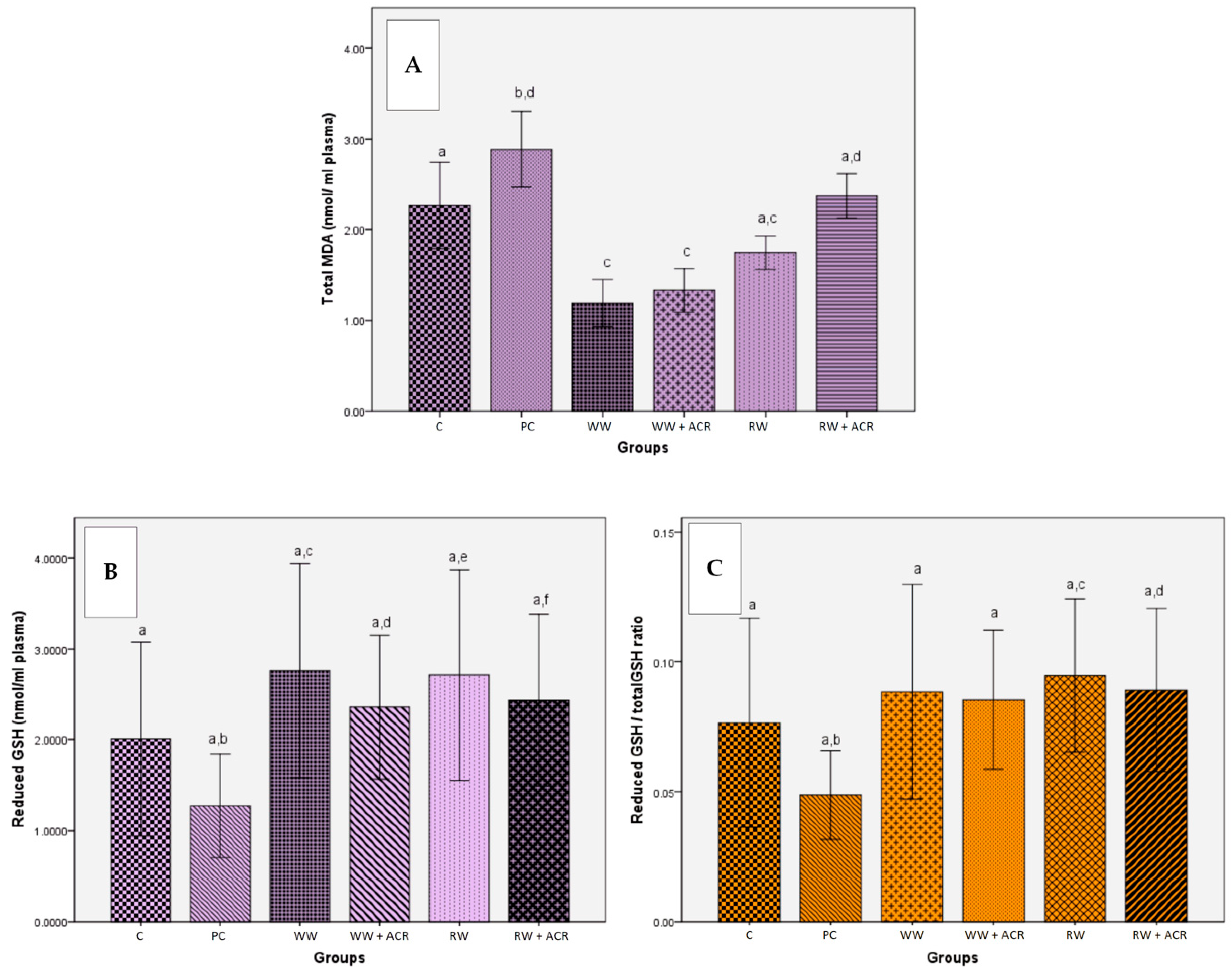
3.4. Effects of Wine Polyphenols on Acrylamide-Induced Oxidative Stress Assessed by Plasma Concentration of Biomarkers
3.4.1. Determination of Lipid Peroxidation by Quantification of Malondialdehyde in Rat Plasma
3.4.2. Determination of Reduced and Total Glutathione Plasma Levels
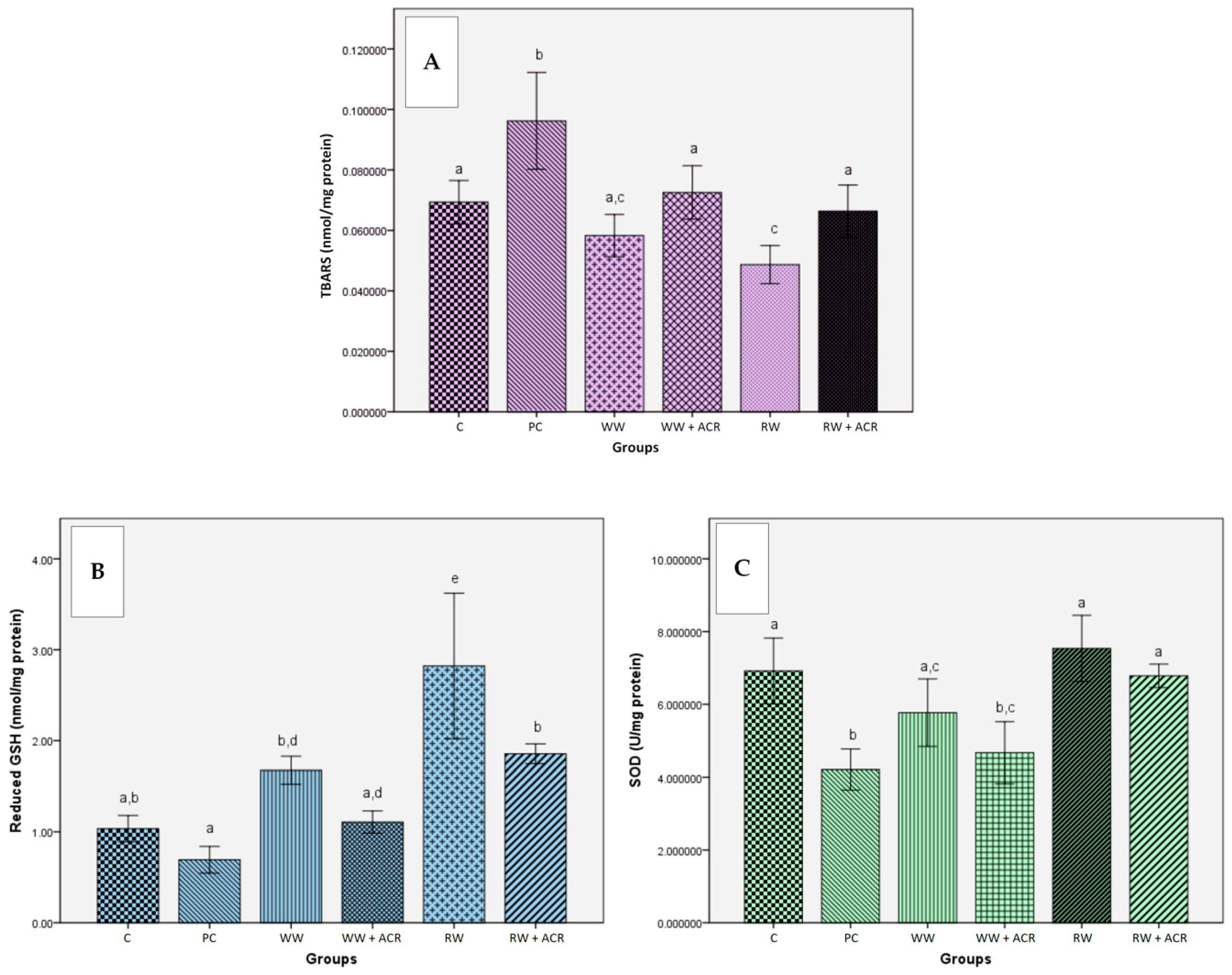
3.5. Effects of Wine Polyphenols on Acrylamide-Induced Oxidative Stress in Liver Tissue
3.5.1. Effects of Wine Polyphenols on Lipid Peroxidation
3.5.2. Effects of Wine Polyphenols on Reduced Glutathione Hepatic Levels
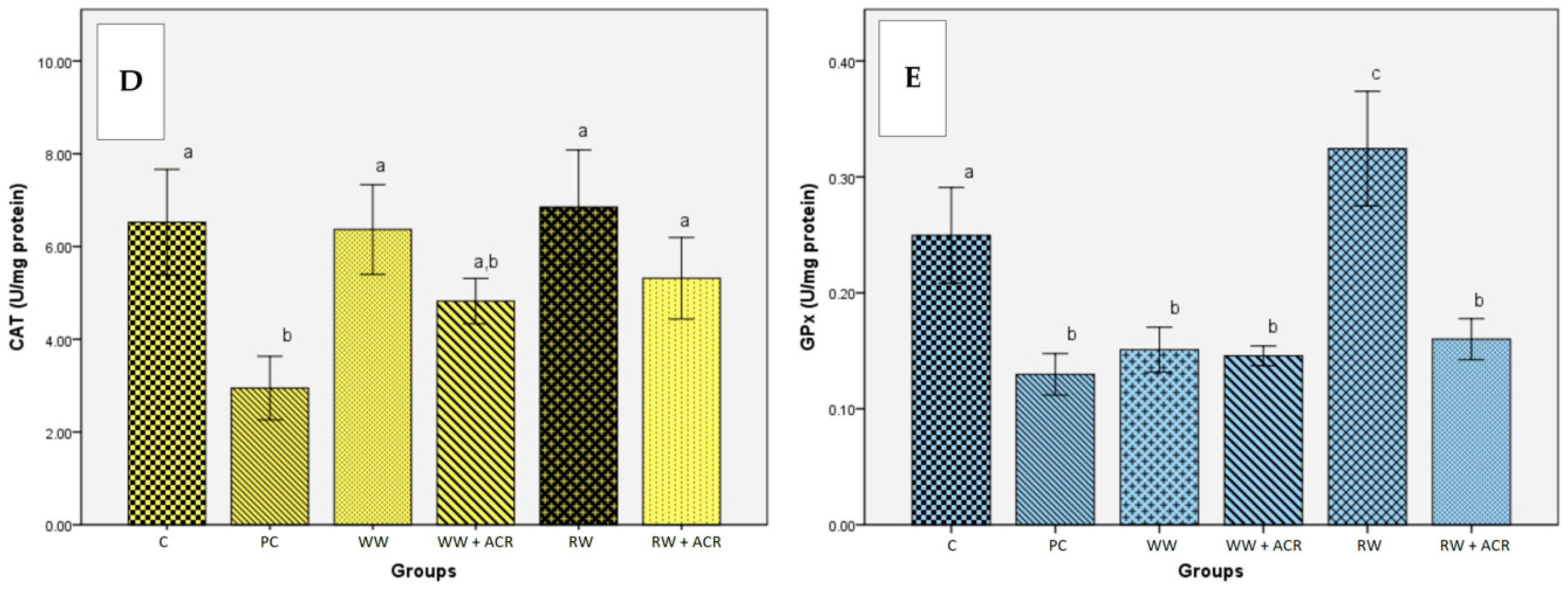
3.5.3. Effects of Wine Polyphenols on Liver Antioxidant Enzymes Activities
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brata, A.M.; Chiciudean, D.I.; Brata, V.D.; Popa, D.; Chiciudean, G.O.; Muresan, I.C. Determinants of Choice and Wine Consumption Behaviour: A Comparative Analysis between Two Counties of Romania. Foods 2022, 11, 1110. [Google Scholar] [CrossRef] [PubMed]
- Castaldo, L.; Narváez, A.; Izzo, L.; Graziani, G.; Gaspari, A.; Di Minno, G.; Ritieni, A. Red Wine Consumption and Cardiovascular Health. Molecules 2019, 24, 3626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buja, L.M. The History, Science, and Art of Wine and the Case for Health Benefits: Perspecitives of an Oenophilic Cardiovascular Pathologist. Cardiovasc. Pathol. 2022, 60, 107446. [Google Scholar] [CrossRef] [PubMed]
- Kołota, A.; Głabska, D.; Oczkowski, M.; Gromadzka-Ostrowska, J. Analysis of Association between Intake of Red Wine Polyphenols and Oxidative Stress Parameters in the Liver of Growing Male Rats. Appl. Sci. 2020, 10, 6389. [Google Scholar] [CrossRef]
- Banc, R.; Socaciu, C.; Miere, D.; Filip, L.; Cozma, A.; Stanciu, O.; Loghin, F. Benefits of Wine Polyphenols on Human Health: A Review. Bull. Univ. Agric. Sci. Vet. Med. Cluj-Napoca. Food Sci. Technol. 2014, 71, 79–87. [Google Scholar] [CrossRef] [Green Version]
- Pavlidou, E.; Mantzorou, M.; Fasoulas, A.; Tryfonos, C.; Petridis, D.; Giaginis, C. Wine: An Aspiring Agent in Promoting Longevity and Preventing Chronic Diseases. Diseases 2018, 6, 73. [Google Scholar] [CrossRef] [Green Version]
- Torres, A.; Cachofeiro, V.; Millán, J.; Lahera, V.; Nieto, M.L.; Martín, R.; Bello, E.; Alvarez-Sala, L.A. El Consumo de Vino Tinto Pero No Otras Bebidas Alcohólicas Aumenta La Capacidad Antioxidante y Mejora El Perfil Pro-Inflamatorio Después de Una Dieta Rica En Grasas Oral En Voluntarios Sanos. Rev. Clin. Esp. 2015, 215, 486–494. [Google Scholar] [CrossRef]
- Kołota, A.; Głąbska, D.; Oczkowski, M.; Gromadzka-Ostrowska, J. Oxidative Stress Parameters in the Liver of Growing Male Rats Receiving Various Alcoholic Beverages. Nutrients 2020, 12, 158. [Google Scholar] [CrossRef] [Green Version]
- Petrella, C.; Carito, V.; Carere, C.; Ferraguti, G.; Ciafrè, S.; Natella, F.; Bello, C.; Greco, A.; Ralli, M.; Mancinelli, R.; et al. Oxidative Stress Inhibition by Resveratrol in Alcohol-Dependent Mice. Nutrition 2020, 79–80, 110783. [Google Scholar] [CrossRef]
- Hiliţanu, L.N.; Mititelu-Tarţău, L.; Bogdan, M.; Buca, B.R.; Păuna, A.-M.R.; Pavel, L.L.; Pelin, A.-M.; Meca, A.D.; Popa, E.G. The Use of Chitosan-Coated Nanovesicles in Repairing Alcohol-Induced Damage of Liver Cells in Mice. Medicina 2022, 58, 762. [Google Scholar] [CrossRef]
- Kasdallah-Grissa, A.; Mornagui, B.; Aouani, E.; Hammami, M.; El May, M.; Gharbi, N.; Kamoun, A.; El-Fazaâ, S. Resveratrol, a Red Wine Polyphenol, Attenuates Ethanol-Induced Oxidative Stress in Rat Liver. Life Sci. 2007, 80, 1033–1039. [Google Scholar] [CrossRef] [PubMed]
- Muñoz-Bernal, Ó.A.; Coria-Oliveros, A.J.; de la Rosa, L.A.; Rodrigo-García, J.; del Rocío Martínez-Ruiz, N.; Sayago-Ayerdi, S.G.; Alvarez-Parrilla, E. Cardioprotective Effect of Red Wine and Grape Pomace. Food Res. Int. 2021, 140, 110069. [Google Scholar] [CrossRef] [PubMed]
- Choleva, M.; Argyrou, C.; Detopoulou, M.; Donta, M.E.; Gerogianni, A.; Moustou, E.; Papaemmanouil, A.; Skitsa, C.; Kolovou, G.; Kalogeropoulos, P.; et al. Effect of Moderate Wine Consumption on Oxidative Stress Markers in Coronary Heart Disease Patients. Nutrients 2022, 14, 1377. [Google Scholar] [CrossRef]
- Sperkowska, B.; Murawska, J.; Przybylska, A.; Gackowski, M.; Kruszewski, S.; Durmowicz, M.; Rutkowska, D. Cardiovascular Effects of Chocolate and Wine—Narrative Review. Nutrients 2021, 13, 4269. [Google Scholar] [CrossRef] [PubMed]
- Gambini, J.; Gimeno-Mallench, L.; Olaso-Gonzalez, G.; Mastaloudis, A.; Traber, M.G.; Monleón, D.; Borrás, C.; Viña, J. Moderate Red Wine Consumption Increases the Expression of Longevity-Associated Genes in Controlled Human Populations and Extends Lifespan in Drosophila Melanogaster. Antioxidants 2021, 10, 301. [Google Scholar] [CrossRef] [PubMed]
- Radeka, S.; Rossi, S.; Bestulic, E.; Budic-Leto, I.; Ganic Kovacevic, K.; Horvat, I.; Lukic, I.; Orbanic, F.; Zaninovic Jurjevic, T.; Dvornik, S. Bioactive Compounds and Antioxidant Activity of Red and White Wines Produced from Autochthonous Croatian Varieties: Effect of Moderate Consumption on Human Health. Foods 2022, 11, 1804. [Google Scholar] [CrossRef]
- Yu, Y.; Tian, Z.Q.; Liang, L.; Yang, X.; Sheng, D.D.; Zeng, J.X.; Li, X.Y.; Shi, R.Y.; Han, Z.P.; Wei, L.X. Babao Dan Attenuates Acute Ethanol-Induced Liver Injury via Nrf2 Activation and Autophagy. Cell Biosci. 2019, 9, 80. [Google Scholar] [CrossRef] [Green Version]
- Fiore, M.; Messina, M.P.; Petrella, C.; D’Angelo, A.; Greco, A.; Ralli, M.; Ferraguti, G.; Tarani, L.; Vitali, M.; Ceccanti, M. Antioxidant Properties of Plant Polyphenols in the Counteraction of Alcohol-Abuse Induced Damage: Impact on the Mediterranean Diet. J. Funct. Foods 2020, 71, 104012. [Google Scholar] [CrossRef]
- Tedesco, I.; Spagnuolo, C.; Russo, G.L.; Russo, M.; Cervellera, C.; Moccia, S. The Pro-Oxidant Activity of Red Wine Polyphenols Induces an Adaptive Antioxidant Response in Human Erythrocytes. Antioxidants 2021, 10, 800. [Google Scholar] [CrossRef]
- Delgado-Andrade, C.; Mesías, M.; Morales, F.J. Introduction to the Special Issue: New Frontiers in Acrylamide Study in Foods-Formation, Analysis and Exposure Assessment. Foods 2020, 9, 1506. [Google Scholar] [CrossRef]
- Sayed, S.; Alotaibi, S.S.; El-Shehawi, A.M.; Hassan, M.M.; Shukry, M.; Alkafafy, M.; Soliman, M.M. The Anti-Inflammatory, Anti-Apoptotic and Antioxidant Effects of a Pomegranate-Peel Extract against Acrylamide-Induced Hepatotoxicity in Rats. Life 2022, 12, 224. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Kim, H.J. Dietary Exposure to Acrylamide and Associated Health Risks for the Korean Population. Int. J. Environ. Res. Public Health 2020, 17, 7619. [Google Scholar] [CrossRef]
- Timmermann, C.A.G.; Mølck, S.S.; Kadawathagedara, M.; Bjerregaard, A.A.; Törnqvist, M.; Brantsæter, A.L.; Pedersen, M. A Review of Dietary Intake of Acrylamide in Humans. Toxics 2021, 9, 155. [Google Scholar] [CrossRef] [PubMed]
- Zha, L.; Sobue, T.; Kitamura, T.; Kitamura, Y.; Ishihara, J.; Kotemori, A.; Liu, R.; Ikeda, S.; Sawada, N.; Iwasaki, M.; et al. Dietary Acrylamide Intake and the Risk of Liver Cancer: The Japan Public Health Center-Based Prospective Study. Nutrients 2020, 12, 2503. [Google Scholar] [CrossRef] [PubMed]
- Markovic Filipovic, J.; Karan, J.; Ivelja, I.; Matavulj, M.; Stošic, M. Acrylamide and Potential Risk of Diabetes Mellitus: Effects on Acrylamide and Potential Risk of Diabetes Mellitus: Effects on Human Glucose Metabolism and Toxicity Human Population, Glucose Metabolism and Beta-Cell Toxicity. Int. J. Mol. Sci. 2022, 23, 6112. [Google Scholar] [CrossRef] [PubMed]
- Michalak, J.; Czarnowska-Kujawska, M.; Klepacka, J.; Gujska, E. Effect of Microwave Heating on the Acrylamide Formation in Foods. Molecules 2020, 25, 4140. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, M.; Vryonidis, E.; Joensen, A.; Törnqvist, M. Hemoglobin Adducts of Acrylamide in Human Blood—What Has Been Done and What Is Next? Food Chem. Toxicol. 2022, 161, 112799. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.; Zhong, H.; Geng, C.; Xue, H.; Wang, C.; Sun, W.; Dang, R.; Han, W.; Jiang, P. Comprehensive Analysis of Metabolic Changes in Rats Exposed to Acrylamide. Environ. Pollut. 2021, 287, 117591. [Google Scholar] [CrossRef]
- European Food Safety Authority. Scientific Opinion on Acrylamide in Food. EFSA Panel on Contaminants in the Food Chain (CONTAM). EFSA J. 2015, 13, 4104. [Google Scholar] [CrossRef] [Green Version]
- Markovic Filipovic, J.; Miler, M.; Kojic, D.; Karan, J.; Ivelja, I.; Cukuranovic Kokoris, J.; Matavulj, M. Effect of Acrylamide Treatment on Cyp2e1 Expression and Redox Status in Rat Hepatocytes. Int. J. Mol. Sci. 2022, 23, 6062. [Google Scholar] [CrossRef]
- World Health Organization; International Agency for Research on Cancer. IARC Working Group on the Evaluation of Carcinogenic Risks to Humans: Some Industrial Chemicals. Lyon, 15–22 February 1994. IARC Monogr. Eval. Carcinog. Risks Hum. 1994, 60, 1–560. [Google Scholar]
- Commission Regulation (EU) 2017/2158; Official Journal of the European Union. Publications Office of the European Union: Luxembourg, 2017.
- Demirci-Çekiç, S.; Özkan, G.; Avan, A.N.; Uzunboy, S.; Çapanoğlu, E.; Apak, R. Biomarkers of Oxidative Stress and Antioxidant Defense. J. Pharm. Biomed. Anal. 2022, 209, 114477. [Google Scholar] [CrossRef] [PubMed]
- Hamdy, S.M.; Shabaan, A.M.; Abdel Latif, A.K.M.; Abdel-Aziz, A.M.; Amin, A.M. Protective Effect of Hesperidin and Tiger Nut against Acrylamide Toxicity in Female Rats. Exp. Toxicol. Pathol. 2017, 69, 580–588. [Google Scholar] [CrossRef]
- Lobo, V.; Patil, A.; Phatak, A.; Chandra, N. Free Radicals, Antioxidants and Functional Foods: Impact on Human Health. Pharmacogn. Rev. 2010, 4, 118–126. [Google Scholar] [CrossRef] [Green Version]
- Qu, D.; Liu, C.; Jiang, M.; Feng, L.; Chen, Y.; Han, J. After In Vitro Digestion, Jackfruit Flake Affords Protection against Acrylamide-Induced Oxidative Damage. Molecules 2019, 24, 3322. [Google Scholar] [CrossRef] [Green Version]
- Popa, D.S.; Bolfa, P.; Kiss, B.; Vlase, L.; Pǎltinean, R.; Pop, A.; Cǎtoi, C.; Crişan, G.; Loghin, F. Influence of Genista Tinctoria L or Methylparaben on Subchronic Toxicity of Bisphenol A in Rats. Biomed. Environ. Sci. 2014, 27, 85–96. [Google Scholar] [CrossRef]
- Vedeanu, N.; Voica, C.; Magdas, D.A.; Kiss, B.; Stefan, M.G.; Simedrea, R.; Georgiu, C.; Berce, C.; Vostinaru, O.; Boros, R.; et al. Subacute Co-Exposure to Low Doses of Ruthenium (III) Changes the Distribution, Excretion and Biological Effects of Silver Ions in Rats. Environ. Chem. 2020, 17, 163–172. [Google Scholar] [CrossRef]
- Banc, R.; Loghin, F.; Miere, D.; Ranga, F.; Socaciu, C. Phenolic Composition and Antioxidant Activity of Red, Rosé and White Wines Originating from Romanian Grape Cultivars. Not. Bot. Horti Agrobot. Cluj-Napoca 2020, 48, 716–734. [Google Scholar] [CrossRef]
- Bradford, M. A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Kiss, B.; Popa, D.S.; Loghin, F.; Iepure, R.; Bojiţă, M. Dozarea Malondialdehidei Libere Şi Totale Prin HPLC. Farmacia 2005, 53, 17–25. [Google Scholar]
- Porfire, A.S.; Leucuţa, S.E.; Kiss, B.; Loghin, F.; Pârvu, A.E. Investigation into the Role of Cu/Zn-SOD Delivery System on Its Antioxidant and Antiinflammatory Activity in Rat Model of Peritonitis. Pharmacol. Rep. 2014, 66, 670–676. [Google Scholar] [CrossRef]
- Kiss, B.; Popa, D.S.; Crişan, G.; Bojiţǎ, M.; Loghin, F. The Evaluation of Antioxidant Potential of Veronica Officinalis and Rosmarinus Officinalis Extracts by Monitoring Malondialdehide and Glutathione Levels in Rats. Farmacia 2009, 57, 432–441. [Google Scholar]
- Conti, M.; Morand, P.C.; Levillain, P.; Lemonnier, A. Improved Fluorometric Determination of Malonaldehyde. Clin. Chem. 1991, 37, 1273–1275. [Google Scholar] [CrossRef]
- Hu, M.L. Measurement of Protein Thiol Groups and Glutathione in Plasma. Methods Enzymol. 1994, 233, 380–385. [Google Scholar] [CrossRef] [PubMed]
- Flohé, L.; Otting, F. Superoxide Dismutase Assays. Methods Enzym. 1984, 105, 93–104. [Google Scholar]
- Pippenger, C.E.; Browne, R.W.; Armstrong, D. Regulatory Antioxidant Enzymes. Methods Mol. Biol. 1998, 108, 299–313. [Google Scholar] [CrossRef]
- Flohé, L.; Günzler, W.A. Assays of Gluthathione Peroxidase. Methods Enzymol. 1984, 105, 114–120. [Google Scholar]
- Alturfan, A.A.; Tozan-Beceren, A.; Sehirli, A.O.; Demiralp, E.; Sener, G.; Omurtag, G. Resveratrol Ameliorates Oxidative DNA Damage and Protects against Acrylamide-Induced Oxidative Stress in Rats. Mol. Biol. Rep. 2012, 39, 4589–4596. [Google Scholar] [CrossRef]
- Rifai, L.; Saleh, F.A. A Review on Acrylamide in Food: Occurrence, Toxicity, and Mitigation Strategies. Int. J. Toxicol. 2020, 39, 93–102. [Google Scholar] [CrossRef]
- Yousef, M.I.; El-Demerdash, F.M. Acrylamide-Induced Oxidative Stress and Biochemical Perturbations in Rats. Toxicology 2006, 219, 133–141. [Google Scholar] [CrossRef]
- Li, J.T.; Liao, Z.X.; Ping, J.; Xu, D.; Wang, H. Molecular Mechanism of Hepatic Stellate Cell Activation and Antifibrotic Therapeutic Strategies. J. Gastroenterol. 2008, 43, 419–428. [Google Scholar] [CrossRef] [PubMed]
- Sun, R.; Chen, W.; Cao, X.; Guo, J.; Wang, J. Protective Effect of Curcumin on Acrylamide-Induced Hepatic and Renal Impairment in Rats: Involvement of CYP2E1. Nat. Prod. Commun. 2020, 15, 1934578X20910548. [Google Scholar] [CrossRef] [Green Version]
- El-Beltagi, H.S.; Ahmed, M.M. Assessment the Protective Role of Quercetin on Acrylamide-Induced Oxidative Stress in Rats. J. Food Biochem. 2016, 40, 715–723. [Google Scholar] [CrossRef]
- Begum Sheikh, R.; Kedam, T. Effect of Acrylamide on Chick Embryonic Liver Glutathione S-Transferases. Med. J. Nutr. Metab. 2010, 3, 31–38. [Google Scholar] [CrossRef]
- Doroshyenko, O.; Fuhr, U.; Kunz, D.; Frank, D.; Kinzig, M.; Jetter, A.; Reith, Y.; Lazar, A.; Taubert, D.; Kirchheiner, J.; et al. In Vivo Role of Cytochrome P450 2E1 and Glutathione-S-Transferase Activity for Acrylamide Toxicokinetics in Humans. Cancer Epidemiol. Biomark. Prev. 2009, 18, 433–443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uzma, N.; Kumar, B.S.; Anees, S. Red Wine Ameliorates CCl4–Induced Acute Liver Injury in Rats. Aust. J. Biomed. Sci. 2011, 1, 1–7. [Google Scholar]
- Lakshmi, B.V.S.; Sudhakar, M.; Aparna, M. Protective Potential of Black Grapes against Lead Induced Oxidative Stress in Rats. Environ. Toxicol. Pharmacol. 2013, 35, 361–368. [Google Scholar] [CrossRef]
- Pizzino, G.; Irrera, N.; Cucinotta, M.; Pallio, G.; Mannino, F.; Arcoraci, V.; Squadrito, F.; Altavilla, D.; Bitto, A. Oxidative Stress: Harms and Benefits for Human Health. Oxid. Med. Cell. Longev. 2017, 2017, 8416763. [Google Scholar] [CrossRef]
- Petitpas, F.; Sichel, F.; Hébert, B.; Lagadu, S.; Beljean, M.; Pottier, D.; Laurentie, M.; Prevost, V. Effects of Alcohol Consumption on Biomarkers of Oxidative Damage to DNA and Lipids in Ethanol-Fed Pigs. Exp. Toxicol. Pathol. 2013, 65, 263–269. [Google Scholar] [CrossRef]
- Al-Qabba, M.M.; El-Mowafy, M.A.; Althwab, S.A.; Alfheeaid, H.A.; Aljutaily, T.; Barakat, H. Phenolic Profile, Antioxidant Activity, and Ameliorating Efficacy of Chenopodium Quinoa Sprouts against CCl4-Induced Oxidative Stress in Rats. Nutrients 2020, 12, 2904. [Google Scholar] [CrossRef]
- Montilla, P.; Espejo, I.; Muñoz, M.C.; Bujalance, I.; Muñoz-Castañeda, J.R.; Túnez, I. Effect of Red Wine on Oxidative Stress and Hypercholesterolemia Induced by Feeding a High-Cholesterol Diet in Rat. J. Physiol. Biochem. 2004, 60, 259–264. [Google Scholar] [CrossRef] [PubMed]
- Macedo, L.F.L.; Rogero, M.M.; Guimarães, J.P.; Granato, D.; Lobato, L.P.; Castro, I.A. Effect of Red Wines with Different in Vitro Antioxidant Activity on Oxidative Stress of High-Fat Diet Rats. Food Chem. 2013, 137, 122–129. [Google Scholar] [CrossRef] [PubMed]
- Ou, J.; Zheng, J.; Huang, J.; Ho, C.T.; Ou, S. Interaction of Acrylamide, Acrolein, and 5-Hydroxymethylfurfural with Amino Acids and DNA. J. Agric. Food Chem. 2020, 68, 5039–5048. [Google Scholar] [CrossRef] [PubMed]
- Gris, E.F.; Mattivi, F.; Ferreira, E.A.; Vrhovsek, U.; Filho, D.W.; Pedrosa, R.C.; Bordignon-Luiz, M.T. Phenolic Profile and Effect of Regular Consumption of Brazilian Red Wines on in Vivo Antioxidant Activity. J. Food Compos. Anal. 2013, 31, 31–40. [Google Scholar] [CrossRef] [Green Version]
- Khan, M.R.; Afzaal, M.; Saeed, N.; Shabbir, M. Protective Potential of Methanol Extract of Digera Muricata on Acrylamide Induced Hepatotoxicity in Rats. African J. Biotechnol. 2011, 10, 8456–8464. [Google Scholar] [CrossRef]
- Rocha, K.K.R.; Souza, G.A.; Ebaid, G.X.; Seiva, F.R.F.; Cataneo, A.C.; Novelli, E.L.B. Resveratrol Toxicity: Effects on Risk Factors for Atherosclerosis and Hepatic Oxidative Stress in Standard and High-Fat Diets. Food Chem. Toxicol. 2009, 47, 1362–1367. [Google Scholar] [CrossRef]
- Rodriguez, C.; Mayo, J.C.; Sainz, R.M.; Antolín, I.; Herrera, F.; Martín, V.; Reiter, R.J. Regulation of Antioxidant Enzymes: A Significant Role for Melatonin. J. Pineal Res. 2004, 36, 1–9. [Google Scholar] [CrossRef]
- Rusu, M.E.; Simedrea, R.; Gheldiu, A.M.; Mocan, A.; Vlase, L.; Popa, D.S.; Ferreira, I.C.F.R. Benefits of Tree Nut Consumption on Aging and Age-Related Diseases: Mechanisms of Actions. Trends Food Sci. Technol. 2019, 88, 104–120. [Google Scholar] [CrossRef]
- Feillet-Coudray, C.; Sutra, T.; Fouret, G.; Ramos, J.; Wrutniak-Cabello, C.; Cabello, G.; Cristol, J.P.; Coudray, C. Oxidative Stress in Rats Fed a High-Fat High-Sucrose Diet and Preventive Effect of Polyphenols: Involvement of Mitochondrial and NAD(P)H Oxidase Systems. Free Radic. Biol. Med. 2009, 46, 624–632. [Google Scholar] [CrossRef]
- Obrenovich, M.E.; Nair, N.G.; Beyaz, A.; Aliev, G.; Reddy, V.P. The Role of Polyphenolic Antioxidants in Health, Disease, and Aging. Rejuvenation Res. 2010, 13, 631–643. [Google Scholar] [CrossRef] [Green Version]
- Hou, L.; Liu, S.; Zhao, C.; Fan, L.; Hu, H.; Yin, S. The Combination of T-2 Toxin and Acrylamide Synergistically Induces Hepatotoxicity and Nephrotoxicity via the Activation of Oxidative Stress and the Mitochondrial Pathway. Toxicon 2021, 189, 65–72. [Google Scholar] [CrossRef] [PubMed]
- Ghasemzadeh Rahbardar, M.; Cheraghi Farmad, H.; Hosseinzadeh, H.; Mehri, S. Protective Effects of Selenium on Acrylamide-Induced Neurotoxicity and Hepatotoxicity in Rats. Iran. J. Basic Med. Sci. 2021, 24, 1041–1049. [Google Scholar] [CrossRef] [PubMed]
- Gao, H.; Xue, Y.; Wu, L.; Huo, J.; Pang, Y.; Chen, J.; Gao, Q. Protective Effect of Lycium Ruthenicum Polyphenols on Oxidative Stress against Acrylamide Induced Liver Injury in Rats. Mol. Nutr. Food Res. 2022, 27, 4100. [Google Scholar] [CrossRef]
- Seydi, E.; Rajabi, M.; Salimi, A.; Pourahmad, J. Involvement of Mitochondrial-Mediated Caspase-3 Activation and Lysosomal Labilization in Acrylamide-Induced Liver Toxicity. Toxicol. Environ. Chem. 2015, 97, 563–575. [Google Scholar] [CrossRef]
- Martín, J.; Kuskoski, E.M.; Navas, M.J.; Asuero, A.G. Antioxidant Capacity of Anthocyanin Pigments. In Flavonoids—From Biosynthesis to Human Health; Justino, G.C., Ed.; IntechOpen: London, UK, 2017. [Google Scholar] [CrossRef] [Green Version]
- Vejarano, R.; Luján-Corro, M. Red Wine and Health: Approaches to Improve the Phenolic Content During Winemaking. Front. Nutr. 2022, 9, 890066. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.; Lewis Wang, F.S.; Hu, X.; Chen, F.; Chan, H.M. Acrylamide-Induced Neurotoxicity in Primary Astrocytes and Microglia: Roles of the Nrf2-ARE and NF-ΚB Pathways. Food Chem. Toxicol. 2017, 106, 25–35. [Google Scholar] [CrossRef]
- Davuljigari, C.B.; Ekuban, F.A.; Zong, C.; Fergany, A.A.M.; Morikawa, K.; Ichihara, G. Nrf2 Activation Attenuates Acrylamide-Induced Neuropathy in Mice. Int. J. Mol. Sci. 2021, 22, 5995. [Google Scholar] [CrossRef]
- Fang, Z.; Luo, Y.; Ma, C.; Dong, L.; Chen, F. Blueberry Anthocyanins Extract Attenuates Acrylamide-Induced Oxidative Stress and Neuroinflammation in Rats. Oxid. Med. Cell. Longev. 2022, 2022, 7340881. [Google Scholar] [CrossRef]
- Pan, X.; Wu, X.; Yan, D.; Peng, C.; Rao, C.; Yan, H. Acrylamide-Induced Oxidative Stress and Inflammatory Response Are Alleviated by N-Acetylcysteine in PC12 Cells: Involvement of the Crosstalk between Nrf2 and NF-ΚB Pathways Regulated by MAPKs. Toxicol. Lett. 2018, 288, 55–64. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| White Wine Sample | TPC (mg GAE/L) | DPPH Radical Scavenging Activity (%) | TAA (mM TE/L) |
|---|---|---|---|
| FRJid2011 | 245 * | 51 * | 0.93 * |
| Red Wine Sample | TPC (mg GAE/L) | DPPH Radical Scavenging Activity (%) | TAA (mM TE/L) |
| FNToh2010 | 2359 * | 95 * | 9.84 * |
| Group | Number of Animals | Intragastric Gavage | Diet |
|---|---|---|---|
| Control (C) | 10 | 12.5% (v/v) hydroalcoholic solution | standard |
| Positive control (PC) | 10 | 12.5% (v/v) hydroalcoholic solution + acrylamide 250 µg/kg of weight, 1% (m/v) aqueous solution | standard |
| White wine (WW) | 10 | FRJid2011 white wine | standard |
| White wine + acrylamide (WW + ACR) | 10 | FRJid2011 white wine + acrylamide 250 µg/kg of weight, 1% (m/v) aqueous solution | standard |
| Red wine (RW) | 10 | FNToh2010 red wine | standard |
| Red wine + acrylamide (RW + ACR) | 10 | FNToh2010 red wine+ acrylamide 250 µg/kg of weight, 1% (m/v) aqueous solution | standard |
| Experimental Groups | ||||||
|---|---|---|---|---|---|---|
| C | PC | WW | WW + ACR | RW | RW + ACR | |
| Absolute liver weight (g) | 4.78 ± 0.15 a | 6.04 ± 0.17 b | 4.66 ± 0.18 a | 6.02 ± 0.15 b | 4.79 ± 0.17 a | 5.99 ± 0.03 b |
| Relative liver weight (%) | 2.58 ± 0.08 a | 3.15 ± 0.09 b | 2.39 ± 0.09 a | 2.94 ± 0.08 b | 2.46 ± 0.09 a | 2.82 ± 0.02 a,b |
| Experimental Groups | ||||||
|---|---|---|---|---|---|---|
| C | PC | WW | WW + ACR | RW | RW + ACR | |
| AST (U/mL plasma) | 101.29 ± 4.96 | 162.06 ± 23.15 a | 116.31 ± 15.06 | 130 ± 12.29 | 105.17 ± 6.45 b | 108.3 ± 10.06 b |
| ALT (U/mL plasma) | 60.34 ± 9.71 | 88.93 ± 10.27 a | 52.93 ± 3.83 b | 70.77 ± 14.79 | 44.94 ± 5.56 b | 62.68 ± 11.61 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Banc, R.; Popa, D.-S.; Cozma-Petruţ, A.; Filip, L.; Kiss, B.; Fărcaş, A.; Nagy, A.; Miere, D.; Loghin, F. Protective Effects of Wine Polyphenols on Oxidative Stress and Hepatotoxicity Induced by Acrylamide in Rats. Antioxidants 2022, 11, 1347. https://doi.org/10.3390/antiox11071347
Banc R, Popa D-S, Cozma-Petruţ A, Filip L, Kiss B, Fărcaş A, Nagy A, Miere D, Loghin F. Protective Effects of Wine Polyphenols on Oxidative Stress and Hepatotoxicity Induced by Acrylamide in Rats. Antioxidants. 2022; 11(7):1347. https://doi.org/10.3390/antiox11071347
Chicago/Turabian StyleBanc, Roxana, Daniela-Saveta Popa, Anamaria Cozma-Petruţ, Lorena Filip, Béla Kiss, Anca Fărcaş, Andras Nagy, Doina Miere, and Felicia Loghin. 2022. "Protective Effects of Wine Polyphenols on Oxidative Stress and Hepatotoxicity Induced by Acrylamide in Rats" Antioxidants 11, no. 7: 1347. https://doi.org/10.3390/antiox11071347