
Dietary L-Arginine or N-Carbamylglutamate Alleviates Colonic Barrier Injury, Oxidative Stress, and Inflammation by Modulation of Intestinal Microbiota in Intrauterine Growth-Retarded Suckling Lambs
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Treatments
2.2. Chemical Analyses of MR
2.3. Tissue Sample Collection
2.4. Determination of Epithelial Tight Junction Permeability of the Colon
2.5. Cytokine Analysis of the Colon Tissue
2.6. Colonic VFAs and LPS Concentration
2.7. mRNA Abundance
2.8. Western Blotting
2.9. Bacterial DNA Extraction of Colonic Mucosa
2.10. 16S rRNA Analysis of Colonic Mucosa-Associated Microbiota
2.11. Sequence Processing and Analysis
2.12. Statistical Analysis
3. Results
3.1. Mitochondrial Reactive Oxygen Species (ROS) Generation and Oxidative Status in the Colon
3.2. The Barrier Function of the Colon
3.3. The Cytokine Concentrations in the Colonic Mucosa
3.4. VFAs and LPS Concentrations in the Colonic Digesta
3.5. Gene Expression
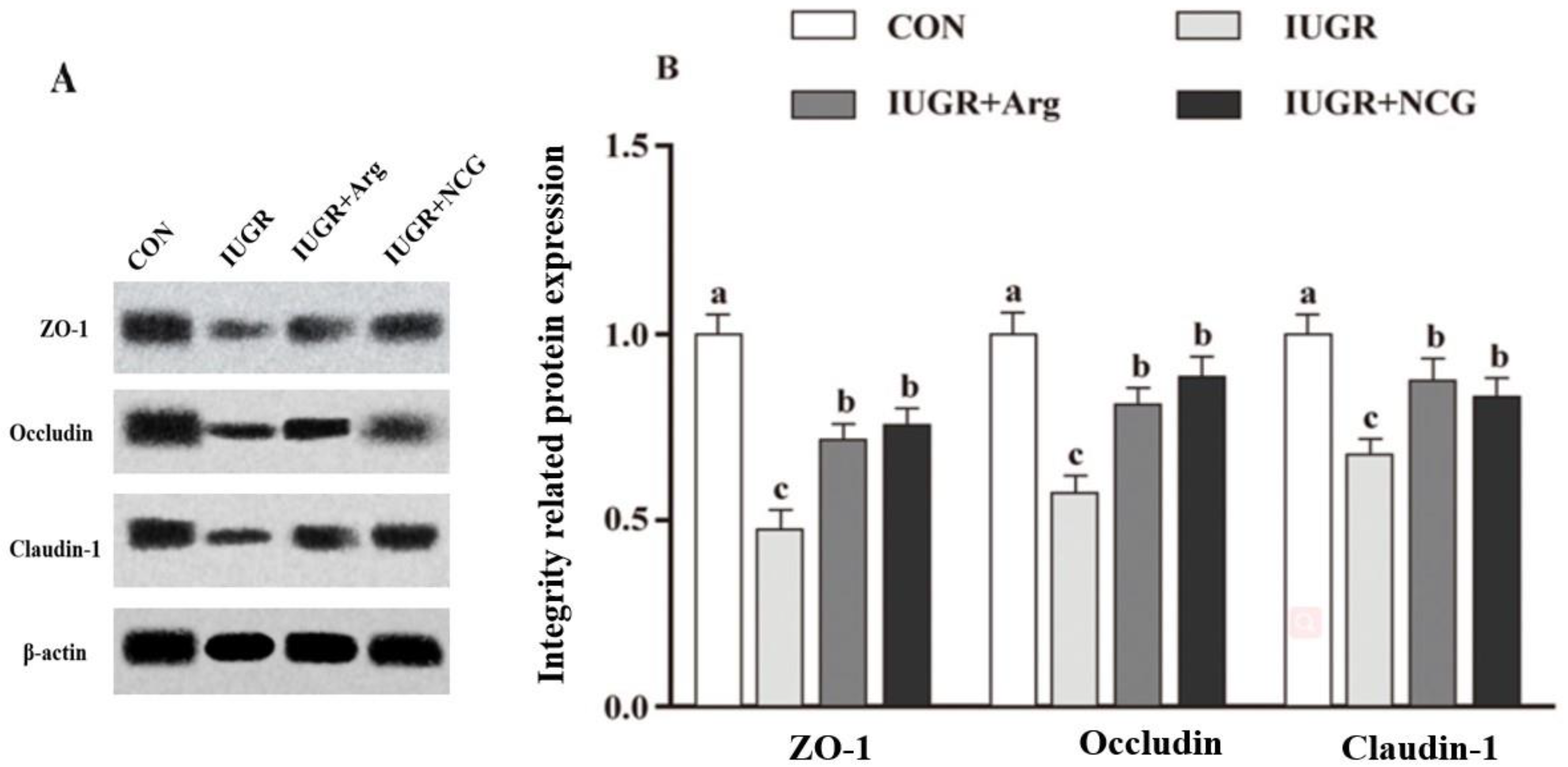
3.6. The Protein Expression
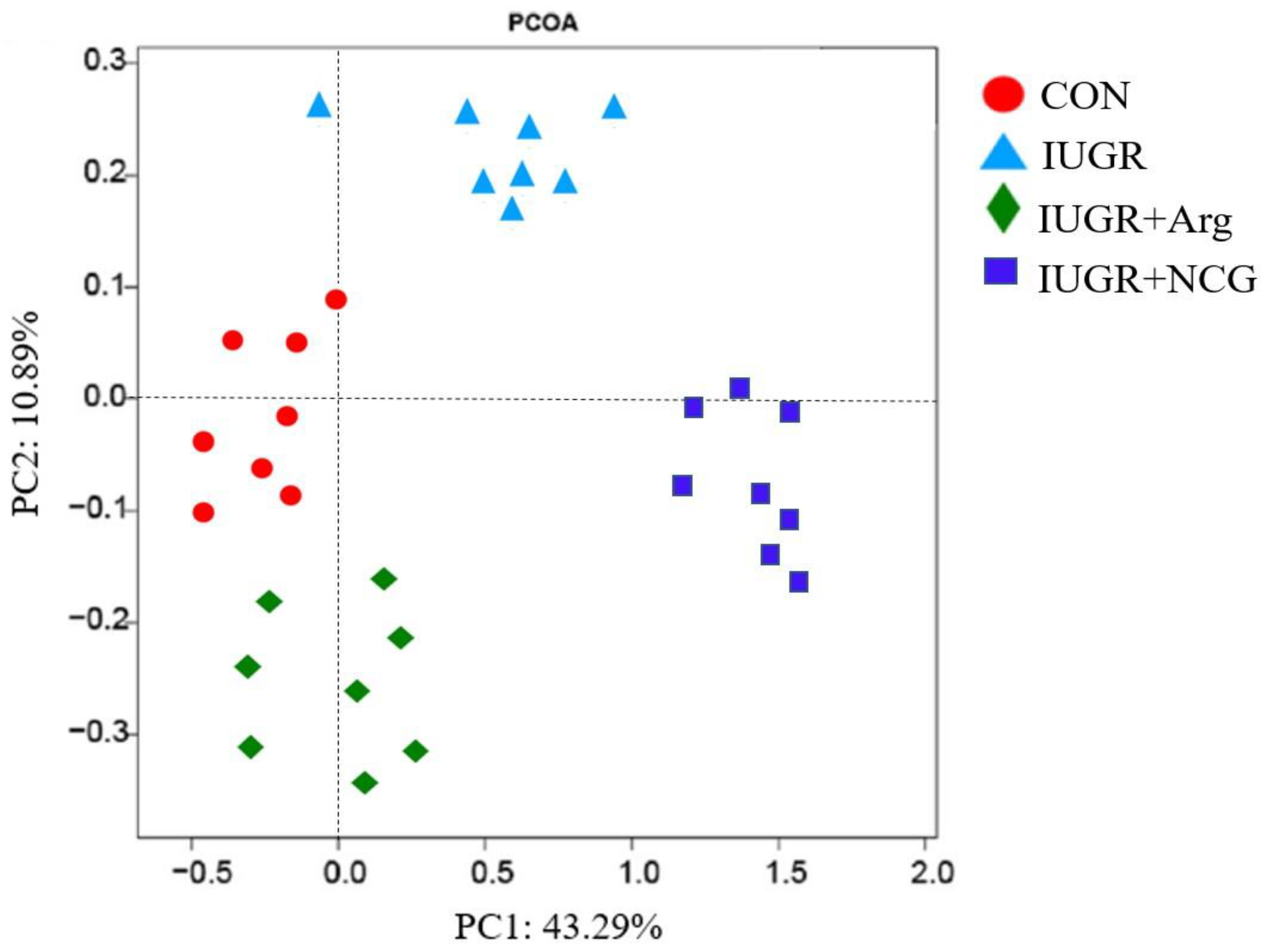
3.7. Diversity of the Colonic Mucosa-Associated Microbiota
3.8. Colonic Mucosal Microbiota Composition
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Désir-Vigné, A.; Haure-Mirande, V.; de Coppet, P.; Darmaun, D.; Le Dréan, G.; Segain, J.-P. Perinatal supplementation of 4-phenylbutyrate and glutamine attenuates endoplasmic reticulum stress and improves colonic epithelial barrier function in rats born with intrauterine growth restriction. J. Nutr. Biochem. 2018, 55, 104–112. [Google Scholar] [CrossRef]
- Bengtson, M.-B.; Ronning, T.; Vatn, M.H.; Harris, J.R. Irritable bowel syndrome in twins: Genes and environment. Gut 2006, 55, 1754–1759. [Google Scholar] [CrossRef] [Green Version]
- Khalili, H.; Ananthakrishnan, A.N.; Higuchi, L.M.; Richter, J.M.; Fuchs, C.S.; Chan, A.T. Early life factors and risk of inflammatory bowel disease in adulthood. Inflamm. Bowel Dis. 2013, 19, 542–547. [Google Scholar] [CrossRef] [Green Version]
- Sandhu, M.S.; Luben, R.; Day, N.E.; Khaw, K.-T. Self-reported birth weight and subsequent risk of colorectal cancer. Cancer Epidemiol. Biomark. Prev. 2002, 11, 935–938. [Google Scholar]
- Wang, W.; Degroote, J.; Van Ginneken, C.; Van Poucke, M.; Vergauwen, H.; Dam, T.M.T.; Vanrompay, D.; Peelman, L.J.; De Smet, S.; Michiels, J. Intrauterine growth restriction in neonatal piglets affects small intestinal mucosal permeability and mRNA expression of redox-sensitive genes. FASEB J. 2016, 30, 863–873. [Google Scholar] [CrossRef] [Green Version]
- O’Hara, A.M.; Shanahan, F. The gut flora as a forgotten organ. EMBO Rep. 2006, 7, 688–693. [Google Scholar] [CrossRef] [Green Version]
- Poulsen, A.-S.R.; De Jonge, N.; Sugiharto, S.; Nielsen, J.L.; Lauridsen, C.; Canibe, N. The microbial community of the gut differs between piglets fed sow milk, milk replacer or bovine colostrum. Br. J. Nutr. 2017, 117, 964–978. [Google Scholar] [CrossRef] [Green Version]
- Matamoros, S.; Gras-Leguen, C.; Le Vacon, F.; Potel, G.; de La Cochetiere, M.-F. Development of intestinal microbiota in infants and its impact on health. Trends Microbiol. 2013, 21, 167–173. [Google Scholar] [CrossRef]
- Li, N.; Huang, S.; Jiang, L.; Wang, W.; Li, T.; Zuo, B.; Li, Z.; Wang, J. Differences in the Gut Microbiota Establishment and Metabolome Characteristics Between Low- and Normal-Birth-Weight Piglets During Early-Life. Front. Microbiol. 2018, 9, 1798. [Google Scholar] [CrossRef] [Green Version]
- Blachier, F.; Beaumont, M.; Andriamihaja, M.; Davila, A.-M.; Lan, A.; Grauso, M.; Armand, L.; Benamouzig, R.; Tomé, D. Changes in the Luminal Environment of the Colonic Epithelial Cells and Physiopathological Consequences. Am. J. Pathol. 2017, 187, 476–486. [Google Scholar] [CrossRef] [Green Version]
- Luo, Y.; Chen, H.; Yu, B.; He, J.; Zheng, P.; Mao, X.; Yu, J.; Luo, J.; Huang, Z.; Chen, D. Dietary pea fibre alters the microbial community and fermentation with increase in fibre degradation-associated bacterial groups in the colon of pigs. J. Anim. Physiol. Anim. Nutr. 2018, 102, e254–e261. [Google Scholar] [CrossRef]
- Zhang, H.; Peng, A.; Yu, Y.; Guo, S.; Wang, M.; Coleman, D.N.; Loor, J.; Wang, H. N-Carbamylglutamate and l-Arginine Promote Intestinal Absorption of Amino Acids by Regulating the mTOR Signaling Pathway and Amino Acid and Peptide Transporters in Suckling Lambs with Intrauterine Growth Restriction. J. Nutr. 2019, 149, 923–932. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.-D.; Tan, J.-Z.; Qi, J.; Zhang, H.-F. Regulatory effects of dietary L-Arg supplementation on the innate immunity and antioxidant ability in broiler chickens. J. Integr. Agric. 2016, 15, 2578–2587. [Google Scholar] [CrossRef]
- Zheng, X.; He, J.; Wang, L.; Zhou, S.; Peng, X.; Huang, S.; Zheng, L.; Cheng, L.; Hao, Y.; Li, J.; et al. Ecological Effect of Arginine on Oral Microbiota. Sci. Rep. 2017, 7, 7206. [Google Scholar] [CrossRef] [Green Version]
- Si, H.; Han, Y.; Liu, H.; Lou, Y.; Li, Z. Effects of rumen-protected arginine supplementation on the plasma amino acids and gut microbiota of sika deer (Cervus nippon). Anim. Feed Sci. Technol. 2021, 273, 114828. [Google Scholar] [CrossRef]
- Tuchman, M.; Caldovic, L.; Daikhin, Y.; Horyn, O.; Nissim, I.; Nissim, I.; Korson, M.; Burton, B.; Yudkoff, M. N-carbamylglutamate Markedly Enhances Ureagenesis in N-acetylglutamate Deficiency and Propionic Acidemia as Measured by Isotopic Incorporation and Blood Biomarkers. Pediatr. Res. 2008, 64, 213–217. [Google Scholar] [CrossRef] [Green Version]
- Wu, X.; Yin, Y.; Liu, Y.; Liu, X.; Liu, Z.; Li, T.; Huang, R.; Ruan, Z.; Deng, Z. Effect of dietary arginine and N-carbamoylglutamate supplementation on reproduction and gene expression of eNOS, VEGFA and PlGF1 in placenta in late pregnancy of sows. Anim. Reprod. Sci. 2012, 132, 187–192. [Google Scholar] [CrossRef]
- Wu, X.; Zhang, Y.; Liu, Z.; Li, T.J.; Yin, Y.L. Effects of oral supplementation with glutamate or combination of glutamate and N-carbamylglutamate on intestinal mucosa morphology and epithelium cell proliferation in weanling piglets1. J. Anim. Sci. 2012, 90, 337–339. [Google Scholar] [CrossRef]
- Zhang, H.; Zheng, Y.; Zha, X.; Liu, X.; Ma, Y.; Loor, J.J.; Elsabagh, M.; Wang, M.; Wang, H.; Jiang, H. Dietary N-carbamylglutamate and L-arginine supplementation improves redox status and suppresses apoptosis in the colon of intrauterine growth-retarded suckling lambs. Anim. Nutr. 2022, 11, 359–368. [Google Scholar] [CrossRef]
- Zhang, H.; Yan, A.; Liu, X.; Ma, Y.; Zhao, F.; Wang, M.; Loor, J.J.; Wang, H. Melatonin ameliorates ochratoxin A induced liver inflammation, oxidative stress and mitophagy in mice involving in intestinal microbiota and restoring the intestinal barrier function. J. Hazard. Mater. 2021, 407, 124489. [Google Scholar] [CrossRef]
- Zhang, H.; Fan, Y.; Elsabagh, M.; Guo, S.; Wang, M.; Jiang, H. Dietary Supplementation of L-Arginine and N-Carbamylglutamate Attenuated the Hepatic Inflammatory Response and Apoptosis in Suckling Lambs with Intrauterine Growth Retardation. Mediat. Inflamm. 2020, 2020, 2453537. [Google Scholar] [CrossRef] [PubMed]
- NRC. Nutrient Requirements of Small Ruminants: Sheep, Goats, Cervids and New World Camelids; The National Academies Press: Washington, DC, USA, 2007. [Google Scholar]
- Zhang, H.; Zhao, F.; Peng, A.; Dong, L.; Wang, M.; Yu, L.; Loor, J.J.; Wang, H. Correction to Effects of Dietary l-Arginine and N-Carbamylglutamate Supplementation on Intestinal Integrity, Immune Function, and Oxidative Status in Intrauterine-Growth-Retarded Suckling Lambs. J. Agric. Food Chem. 2018, 66, 6254. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Ruan, Z.; Gao, Y.; Yin, Y.; Zhou, X.; Wang, L.; Geng, M.; Hou, Y.; Wu, G. Dietary supplementation with l-arginine or N-carbamylglutamate enhances intestinal growth and heat shock protein-70 expression in weanling pigs fed a corn- and soybean meal-based diet. Amino Acids 2010, 39, 831–839. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Jin, Y.; Wang, M.; Loor, J.J.; Wang, H. N-Carbamylglutamate and l-arginine supplementation improve hepatic antioxidant status in intrauterine growth-retarded suckling lambs. RSC Adv. 2020, 10, 11173–11181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- AOAC. Official Methods of Analysis; Association of Official Analytical Chemists: Washington, DC, USA, 1990. [Google Scholar]
- Bidlingmeyer, B.A.; Cohen, S.A.; Tarvin, T.L. Rapid analysis of amino acids using pre-column derivatization. J. Chromatogr. B Biomed. Sci. Appl. 1984, 336, 93–104. [Google Scholar] [CrossRef]
- Qin, W.L. Determination of rumen volatile fatty acids by means of gas chromatography. J. Nanjing Agric. Coll. 1982, 4, 110–116. [Google Scholar]
- Ye, H.; Liu, J.; Feng, P.; Zhu, W.; Mao, S. Grain-rich diets altered the colonic fermentation and mucosa-associated bacterial communities and induced mucosal injuries in goats. Sci. Rep. 2016, 6, 20329. [Google Scholar] [CrossRef] [Green Version]
- Hamard, A.; Mazurais, D.; Boudry, G.; Luron, I.; Sève, B.; Le Floc’h, N. A moderate threonine deficiency affects gene expression profile, paracellular permeability and glucose absorption capacity in the ileum of piglets. J. Nutr. Biochem. 2010, 21, 914–921. [Google Scholar] [CrossRef] [Green Version]
- Hu, C.H.; Xiao, K.; Luan, Z.S.; Song, J. Early weaning increases intestinal permeability, alters expression of cytokine and tight junction proteins, and activates mitogen-activated protein kinases in pigs1. J. Anim. Sci. 2013, 91, 1094–1101. [Google Scholar] [CrossRef] [Green Version]
- Xue, Y.; Hu, F.; Guo, C.; Mei, S.; Xie, F.; Zeng, H.; Mao, S. Undernutrition shifted colonic fermentation and digest-associated bacterial communities in pregnant ewes. Appl. Microbiol. Biotechnol. 2020, 104, 5973–5984. [Google Scholar] [CrossRef]
- Xu, P.; Wang, J.; Hong, F.; Wang, S.; Jin, X.; Xue, T.; Jia, L.; Zhai, Y. Melatonin prevents obesity through modulation of gut microbiota in mice. J. Pineal Res. 2017, 62, e12399. [Google Scholar] [CrossRef]
- Tao, S.; Bai, Y.; Li, T.; Li, N.; Wang, J. Original low birth weight deteriorates the hindgut epithelial barrier function in pigs at the growing stage. FASEB J. 2019, 33, 9897–9912. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) method. Methods 2001, 25, 402–408. [Google Scholar] [PubMed]
- Hnasko, T.S.; Hnasko, R.M. The western blot. Methods Mol. Biol. 2015, 1318, 87–96. [Google Scholar] [PubMed]
- Wang, Y.; Xu, L.; Liu, J.; Zhu, W.; Mao, S. A High Grain Diet Dynamically Shifted the Composition of Mucosa-Associated Microbiota and Induced Mucosal Injuries in the Colon of Sheep. Front. Microbiol. 2017, 8, 2080. [Google Scholar] [CrossRef] [Green Version]
- Caporaso, J.G.; Bittinger, K.; Bushman, F.D.; DeSantis, T.Z.; Andersen, G.L.; Knight, R. PyNAST: A flexible tool for aligning sequences to a template alignment. Bioinformatics 2010, 26, 266–267. [Google Scholar] [CrossRef] [Green Version]
- Campbell, B.J.; Polson, S.; Hanson, T.; Mack, M.C.; Schuur, E.A.G. The effect of nutrient deposition on bacterial communities in Arctic tundra soil. Environ. Microbiol. 2010, 12, 1842–1854. [Google Scholar] [CrossRef]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naive Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microb. 2007, 73, 5261–5267. [Google Scholar] [CrossRef] [Green Version]
- DeSantis, T.Z.; Hugenholtz, P.; Larsen, N.; Rojas, M.; Brodie, E.L.; Keller, K.; Huber, T.; Dalevi, D.; Hu, P.; Andersen, G.L. Greengenes, a Chimera-Checked 16S rRNA Gene Database and Workbench Compatible with ARB. Appl. Environ. Microbiol. 2006, 72, 5069–5072. [Google Scholar] [CrossRef] [Green Version]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Gonzalez Peña, A.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schloss, P.D.; Westcott, S.L.; Ryabin, T.; Hall, J.R.; Hartmann, M.; Hollister, E.B.; Lesniewski, R.A.; Oakley, B.B.; Parks, D.H.; Robinson, C.J.; et al. Introducing mothur: Open-Source, Platform-Independent, Community-Supported Software for Describing and Comparing Microbial Communities. Appl. Environ. Microbiol. 2009, 75, 7537–7541. [Google Scholar] [CrossRef] [PubMed]
- Wu, G.; Bazer, F.W.; Davis, T.A.; Kim, S.W.; Li, P.; Rhoads, J.M.; Satterfield, M.C.; Smith, S.B.; Spencer, T.E.; Yin, Y. Arginine metabolism and nutrition in growth, health and disease. Amino Acids 2009, 37, 153–168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crenn, P.; Messing, B.; Cynober, L. Citrulline as a biomarker of intestinal failure due to enterocyte mass reduction. Clin. Nutr. 2008, 27, 328–339. [Google Scholar] [CrossRef]
- Turner, J.R. Intestinal mucosal barrier function in health and disease. Nat. Rev. Immunol. 2009, 9, 799–809. [Google Scholar] [CrossRef] [PubMed]
- Cao, S.; Shen, Z.; Wang, C.; Zhang, Q.; Hong, Q.; He, Y.; Hu, C. Resveratrol improves intestinal barrier function, alleviates mitochondrial dysfunction and induces mitophagy in diquat challenged piglets1. Food Funct. 2019, 10, 344–354. [Google Scholar] [CrossRef]
- Runkle, E.A.; Mu, D. Tight junction proteins: From barrier to tumorigenesis. Cancer Lett. 2013, 337, 41–48. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Jin, Y.; Peng, A.; Guo, S.; Loor, J.; Wang, H. L-Arginine protects ovine intestinal epithelial cells from lipopolysaccharide-induced intestinal barrier injury. Food Agric. Immunol. 2019, 30, 1067–1084. [Google Scholar] [CrossRef]
- Denning, T.L.; Takaishi, H.; Crowe, S.E.; Boldogh, I.; Jevnikar, A.; Ernst, P.B. Oxidative stress induces the expression of Fas and Fas ligand and apoptosis in murine intestinal epithelial cells. Free. Radic. Biol. Med. 2002, 33, 1641–1650. [Google Scholar] [CrossRef]
- Tang, X.; Xiong, K. Intrauterine Growth Retardation Affects Intestinal Health of Suckling Piglets via Altering Intestinal Antioxidant Capacity, Glucose Uptake, Tight Junction, and Immune Responses. Oxidative Med. Cell. Longev. 2022, 2022, 2644205. [Google Scholar] [CrossRef]
- Navarro-Yepes, J.; Burns, M.; Anandhan, A.; Khalimonchuk, O.; Del Razo, L.M.; Quintanilla-Vega, B.; Pappa, A.; Panayiotidis, M.I.; Franco, R. Oxidative stress, redox signaling, and autophagy: Cell death versus survival. Antioxid. Redox Signal. 2014, 21, 66–85. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, C.; Song, P.; Fan, P.; Hou, C.; Thacker, P.; Ma, X. Dietary Sodium Butyrate Decreases Postweaning Diarrhea by Modulating Intestinal Permeability and Changing the Bacterial Communities in Weaned Piglets. J. Nutr. 2015, 145, 2774–2780. [Google Scholar] [CrossRef]
- Sabroe, I.; Parker, L.; Dower, S.; Whyte, M. The role of TLR activation in inflammation. J. Pathol. 2008, 214, 126–135. [Google Scholar] [CrossRef] [PubMed]
- Kawai, T.; Akira, S. The roles of TLRs, RLRs and NLRs in pathogen recognition. Int. Immunol. 2019, 21, 317–337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, S.T.; Wang, Y.; Xue, Y.; Feng, D.C.; Xu, Y.; Xu, L.Y. Tetrandrine suppresses LPS-induced astrocyte activation via modulating IKKs-IkappaBalpha-NF-kappaB signaling pathway. Mol. Cell. Biochem. 2008, 315, 41–49. [Google Scholar] [CrossRef] [PubMed]
- Kasubuchi, M.; Hasegawa, S.; Hiramatsu, T.; Ichimura, A.; Kimura, I. Dietary Gut Microbial Metabolites, Short-chain Fatty Acids, and Host Metabolic Regulation. Nutrients 2015, 7, 2839–2849. [Google Scholar] [CrossRef] [Green Version]
- Sun, X.; Cui, Y.; Su, Y.; Gao, Z.; Diao, X.; Li, J.; Zhu, X.; Li, D.; Li, Z.; Wang, C.; et al. Dietary Fiber Ameliorates Lipopolysaccharide-Induced Intestinal Barrier Function Damage in Piglets by Modulation of Intestinal Microbiome. mSystems 2021, 6, e01374–20. [Google Scholar] [CrossRef]
- Metzler-Zebeli, B.U.; Lawlor, P.G.; Magowan, E.; Zebeli, Q. Interactions between metabolically active bacteria and host gene expression at the cecal mucosa in pigs of diverging feed efficiency. J. Anim. Sci. 2018, 96, 2249–2264. [Google Scholar] [CrossRef]
- Wang, H.; Wang, G.; Banerjee, N.; Liang, Y.; Du, X.; Boor, P.J.; Hoffman, K.L.; Khan, M.F. Aberrant Gut Microbiome Contributes to Intestinal Oxidative Stress, Barrier Dysfunction, Inflammation and Systemic Autoimmune Responses in MRL/lpr Mice. Front. Immunol. 2021, 12, 651191. [Google Scholar] [CrossRef]
- Ruan, D.; Fouad, A.; Fan, Q.; Huo, X.; Kuang, Z.; Wang, H.; Guo, C.; Deng, Y.; Zhang, C.; Zhang, J.; et al. Dietary L-arginine supplementation enhances growth performance, intestinal antioxidative capacity, immunity and modulates gut microbiota in yellow-feathered chickens. Poult. Sci. 2020, 99, 6935–6945. [Google Scholar] [CrossRef]
- Huang, Y.; Shi, X.; Li, Z.; Shen, Y.; Shi, X.; Wang, L.; Li, G.; Yuan, Y.; Wang, J.; Zhang, Y.; et al. Possible association of Firmicutes in the gut microbiota of patients with major depressive disorder. Neuropsychiatr. Dis. Treat. 2018, 14, 3329–3337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madigan, M.T.; Martinko, J.M.; Parker, J. Brock Biology of Microorganisms; Prentice Hall: Upper Saddle River, NJ, USA, 2003. [Google Scholar]
- Clemente, J.C.; Manasson, J.; Scher, J.U. The role of the gut microbiome in systemic inflammatory disease. BMJ 2018, 360, j5145. [Google Scholar] [CrossRef] [PubMed]
- Henao-Mejia, J.; Elinav, E.; Jin, C.; Hao, L.; Mehal, W.Z.; Strowig, T.; Thaiss, C.A.; Kau, A.L.; Eisenbarth, S.C.; Jurczak, M.J.; et al. Inflammasome-mediated dysbiosis regulates progression of NAFLD and obesity. Nature 2012, 482, 179–185. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; He, Z.; Ma, N.; Chen, Z.-Y. Beneficial Effects of Dietary Polyphenols on High-Fat Diet-Induced Obesity Linking with Modulation of Gut Microbiota. J. Agric. Food Chem. 2020, 68, 33–47. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Su, H.; Li, Q.; Wu, H.; Liu, M.; Huang, J.; Zeng, M.; Zheng, Y.; Sun, X. Oral administration of Clostridium butyricum CGMCC0313-1 inhibits β-lactoglobulin-induced intestinal anaphylaxis in a mouse model of food allergy. Gut Pathog. 2017, 9, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Louis, P.; Hold, G.L.; Flint, H.J. The gut microbiota, bacterial metabolites and colorectal cancer. Nat. Rev. Microbiol. 2014, 12, 661–672. [Google Scholar] [CrossRef]
- Koh, A.; De Vadder, F.; Kovatcheva-Datchary, P.; Bäckhed, F. From Dietary Fiber to Host Physiology: Short-Chain Fatty Acids as Key Bacterial Metabolites. Cell 2016, 165, 1332–1345. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Bosque, A.; Amat, C.; Polo, J.; Campbell, J.M.; Crenshaw, J.; Russell, L.; Moretό, M. Spray-Dried Animal Plasma Prevents the Effects of Staphylococcus aureus Enterotoxin B on Intestinal Barrier Function in Weaned Rats. J. Nutr. 2006, 136, 2838–2843. [Google Scholar] [CrossRef] [Green Version]
- Chung, H.; Pamp, S.J.; Hill, J.A.; Surana, N.K.; Edelman, S.M.; Troy, E.B.; Reading, N.C.; Villablanca, E.J.; Wang, S.; Mora, J.R.; et al. Gut Immune Maturation Depends on Colonization with a Host-Specific Microbiota. Cell 2012, 149, 1578–1593. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Item | CON 2 | IUGR 3 | IUGR + Arg 4 | IUGR + NCG 5 | SEM | p-Value |
|---|---|---|---|---|---|---|
| TER, Ω·cm2 | 17.2 a | 9.48 c | 12.9 b | 13.1 b | 1.28 | 0.008 |
| FD4, μg·cm−2·h−1 | 14.5 c | 23.1 a | 17.4 b | 18.3 b | 1.37 | 0.027 |
| Item | CON 2 | IUGR 3 | IUGR + Arg 4 | IUGR + NCG 5 | SEM | p-Value |
|---|---|---|---|---|---|---|
| TNF-α, ng g−1 protein | 9.11 c | 17.89 a | 13.16 b | 12.98 b | 1.062 | 0.009 |
| IL-1β, ng g−1 protein | 14.12 c | 22.26 a | 17.89 b | 18.01 b | 1.291 | 0.012 |
| IL-6, ng g−1 protein | 10.47 c | 19.89 a | 14.68 b | 14.72 b | 0.876 | 0.018 |
| Item | CON 2 | IUGR 3 | IUGR + Arg 4 | IUGR + NCG 5 | SEM | p-Value |
|---|---|---|---|---|---|---|
| LPS, ng/mL | 9.86 c | 21.45 a | 15.13 b | 14.99 b | 1.513 | 0.009 |
| Acetate, μmol/g | 62.96 a | 38.19 c | 47.43 b | 50.11 b | 2.872 | 0.019 |
| Propionate, μmol/g | 17.16 a | 10.42 c | 13.89 b | 14.02 b | 1.461 | 0.008 |
| Butyrate, μmol/g | 4.68 a | 2.68 c | 3.59 b | 3.64 b | 0.794 | 0.021 |
| Isobutyrate, μmol/g | 6.48 a | 2.59 c | 4.08 b | 4.21 b | 0.287 | 0.005 |
| Valerate, μmol/g | 1.69 | 1.72 | 1.67 | 1.70 | 0.208 | 0.098 |
| Isovalerate, μmol/g | 0.78 | 0.81 | 0.77 | 0.80 | 0.083 | 0.207 |
| Item | CON 2 | IUGR 3 | IUGR + Arg 4 | IUGR + NCG 5 | SEM | p-Value |
|---|---|---|---|---|---|---|
| Barrier function-related genes | ||||||
| ZO-1 | 1.00 a | 0.52 c | 0.69 b | 0.92 a | 0.121 | 0.009 |
| Occludin | 1.00 a | 0.61 c | 0.82 b | 0.79 b | 0.152 | 0.006 |
| Claudin-1 | 1.00 a | 0.48 c | 0.69 b | 0.71 b | 0.093 | 0.012 |
| Immune function-related genes | ||||||
| MyD88 | 1.00 c | 1.63 a | 1.31 b | 1.35 b | 0.078 | 0.009 |
| TLR-4 | 1.00 b | 1.33 a | 1.19 ab | 1.14 ab | 0.098 | 0.023 |
| NF-κB | 1.00 c | 1.65 a | 1.30 b | 1.27 b | 0.162 | 0.019 |
| TNF-α | 1.00 c | 1.70 a | 1.42 b | 1.39 b | 0.094 | 0.008 |
| IL-1β | 1.00 b | 1.58 a | 1.05 b | 0.97 b | 0.069 | 0.013 |
| IL-6 | 1.00 c | 1.69 a | 1.36 b | 1.37 b | 0.128 | 0.006 |
| Item | CON 2 | IUGR 3 | IUGR + Arg 4 | IUGR + NCG 5 | SEM | p-Value |
|---|---|---|---|---|---|---|
| OTUs | 1198 a | 639 c | 895 b | 903 b | 34.2 | 0.007 |
| ACE | 1386 a | 692 c | 912 b | 957 b | 40.8 | 0.005 |
| Chao 1 | 1279 a | 603 c | 903 b | 893 b | 39.3 | 0.010 |
| Shannon index | 5.12 a | 3.25 c | 4.02 b | 4.13 b | 0.277 | 0.009 |
| Simpson index | 0.94 | 0.90 | 0.95 | 0.91 | 0.052 | 0.085 |
| Item | CON 2 | IUGR 3 | IUGR + Arg 4 | IUGR + NCG 5 | SEM | p-Value |
|---|---|---|---|---|---|---|
| Acidobacteria | 1.35 | 1.32 | 1.36 | 1.33 | 0.228 | 0.118 |
| Actinobacteria | 4.27 | 4.19 | 4.63 | 4.81 | 0.392 | 0.094 |
| Bacteroidetes | 9.12 c | 16.47 a | 12.68 b | 13.03 b | 1.121 | 0.009 |
| Chloroflexi | 1.24 | 1.17 | 1.23 | 1.19 | 0.327 | 0.103 |
| Cyanobacteria | 4.13 | 4.32 | 4.09 | 4.26 | 0.183 | 0.079 |
| Firmicutes | 55.34 a | 31.19 c | 43.12 b | 42.32 b | 3.476 | 0.005 |
| Proteobacteria | 4.78 c | 9.03 a | 6.83 b | 7.02 b | 1.032 | 0.013 |
| Phylum | Genus | CON 2 | IUGR 3 | IUGR + Arg 4 | IUGR + NCG 5 | SEM | p-Value |
|---|---|---|---|---|---|---|---|
| Actinobacteria | Actinomyces Bifidobacterium Rothia | 1.56 0.67 0.12 | 1.64 0.72 0.13 | 1.62 0.66 0.11 | 1.59 0.71 0.09 | 0.287 0.182 0.047 | 0.077 0.126 0.083 |
| Bacteroidetes | Bacteroides Prevotella | 0.18 c 1.26 | 0.36 a 1.31 | 0.29 b 1.29 | 0.25 b 1.30 | 0.083 0.152 | 0.007 0.208 |
| Firmicutes | Blautia Butyrivibrio Clostridium Lactobacillus Ruminococcus Staphylococcus Streptococcus Veillonella | 0.88 2.11 0.89 a 2.01 a 1.68 1.04 c 2.21 a 0.31 | 1.03 1.98 0.46 c 1.35 c 1.73 3.12 a 1.28 c 0.29 | 0.97 1.99 0.62 b 1.67 b 1.65 1.94 b 1.68 b 0.34 | 0.91 2.06 0.70 b 1.72 b 1.72 1.86 b 1.71 b 0.32 | 0.121 0.757 0.124 0.272 0.321 0.537 0.366 0.082 | 0.097 0.162 0.008 0.013 0.095 0.008 0.006 0.203 |
| Proteobacteria | Escherichia | 0.78 c | 1.36 a | 0.97 b | 1.02 b | 0.321 | 0.019 |
| Pseudomonas | 1.03 c | 1.67 a | 1.43 b | 1.39 b | 0.286 | 0.007 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, H.; Zheng, Y.; Zha, X.; Ma, Y.; Liu, X.; Elsabagh, M.; Wang, H.; Wang, M. Dietary L-Arginine or N-Carbamylglutamate Alleviates Colonic Barrier Injury, Oxidative Stress, and Inflammation by Modulation of Intestinal Microbiota in Intrauterine Growth-Retarded Suckling Lambs. Antioxidants 2022, 11, 2251. https://doi.org/10.3390/antiox11112251
Zhang H, Zheng Y, Zha X, Ma Y, Liu X, Elsabagh M, Wang H, Wang M. Dietary L-Arginine or N-Carbamylglutamate Alleviates Colonic Barrier Injury, Oxidative Stress, and Inflammation by Modulation of Intestinal Microbiota in Intrauterine Growth-Retarded Suckling Lambs. Antioxidants. 2022; 11(11):2251. https://doi.org/10.3390/antiox11112251
Chicago/Turabian StyleZhang, Hao, Yi Zheng, Xia Zha, Yi Ma, Xiaoyun Liu, Mabrouk Elsabagh, Hongrong Wang, and Mengzhi Wang. 2022. "Dietary L-Arginine or N-Carbamylglutamate Alleviates Colonic Barrier Injury, Oxidative Stress, and Inflammation by Modulation of Intestinal Microbiota in Intrauterine Growth-Retarded Suckling Lambs" Antioxidants 11, no. 11: 2251. https://doi.org/10.3390/antiox11112251