
Effects of Quercetin on the Intestinal Microflora of Freshwater Dark Sleeper Odontobutis potamophila
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Culture
2.2. Experimental Design and Sample Collection
2.3. Illumina Miseq Sequencing
2.4. Quantitative Real-Time PCR Analysis
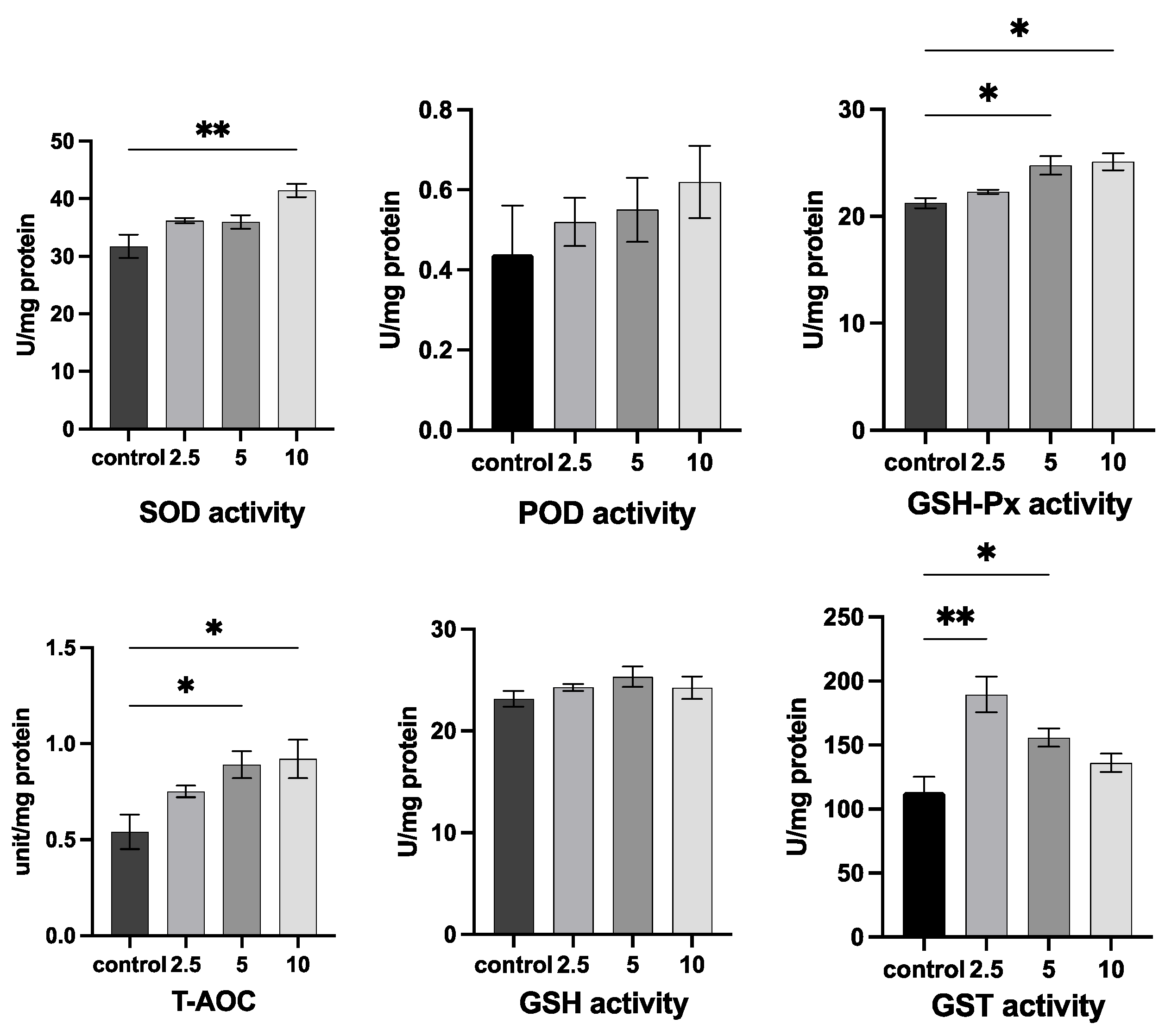
2.5. Antioxidant Enzyme Activity Assay
2.6. Data Analysis
3. Results
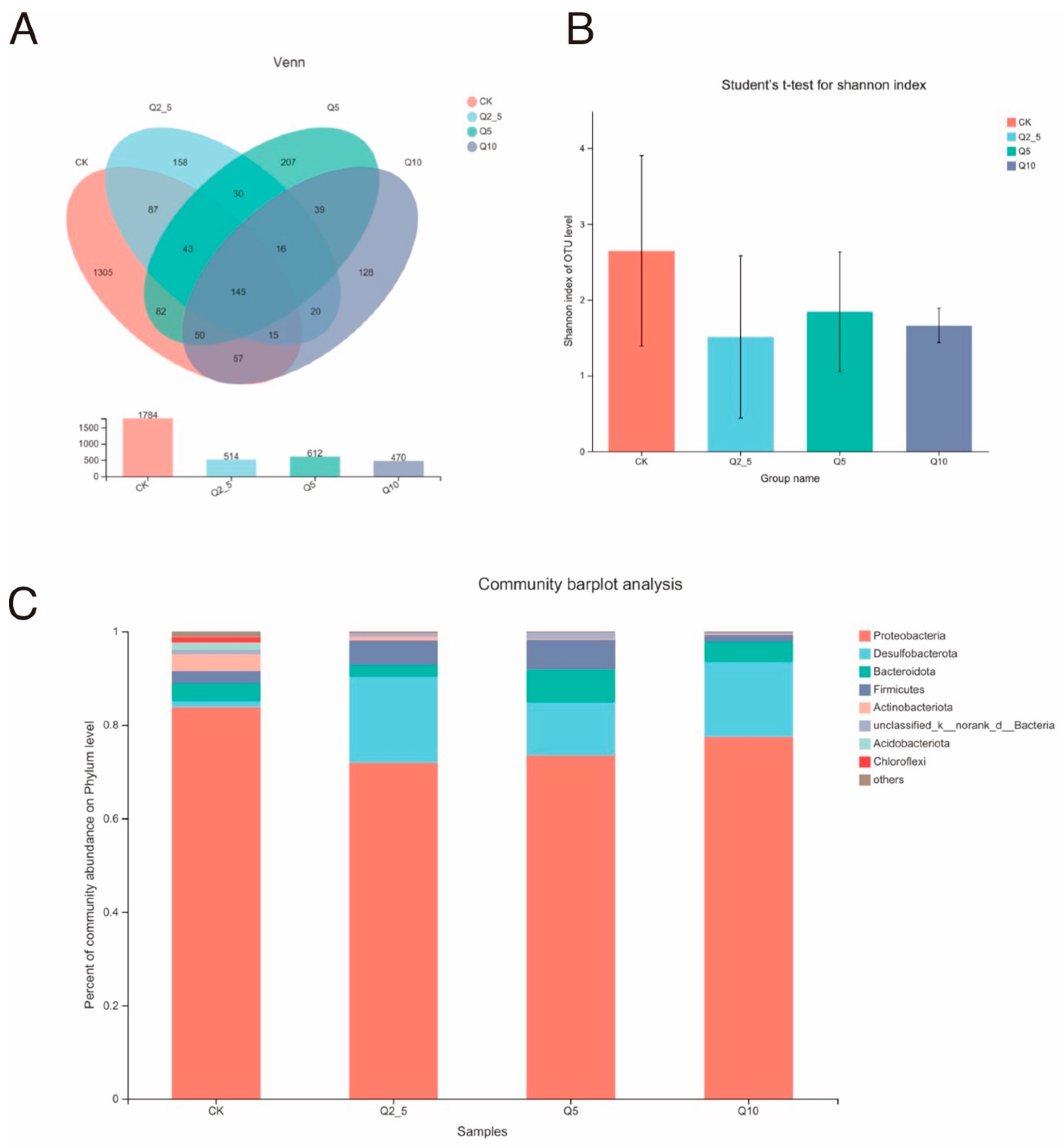
3.1. S rRNA Sequencing Data and Species Evaluation
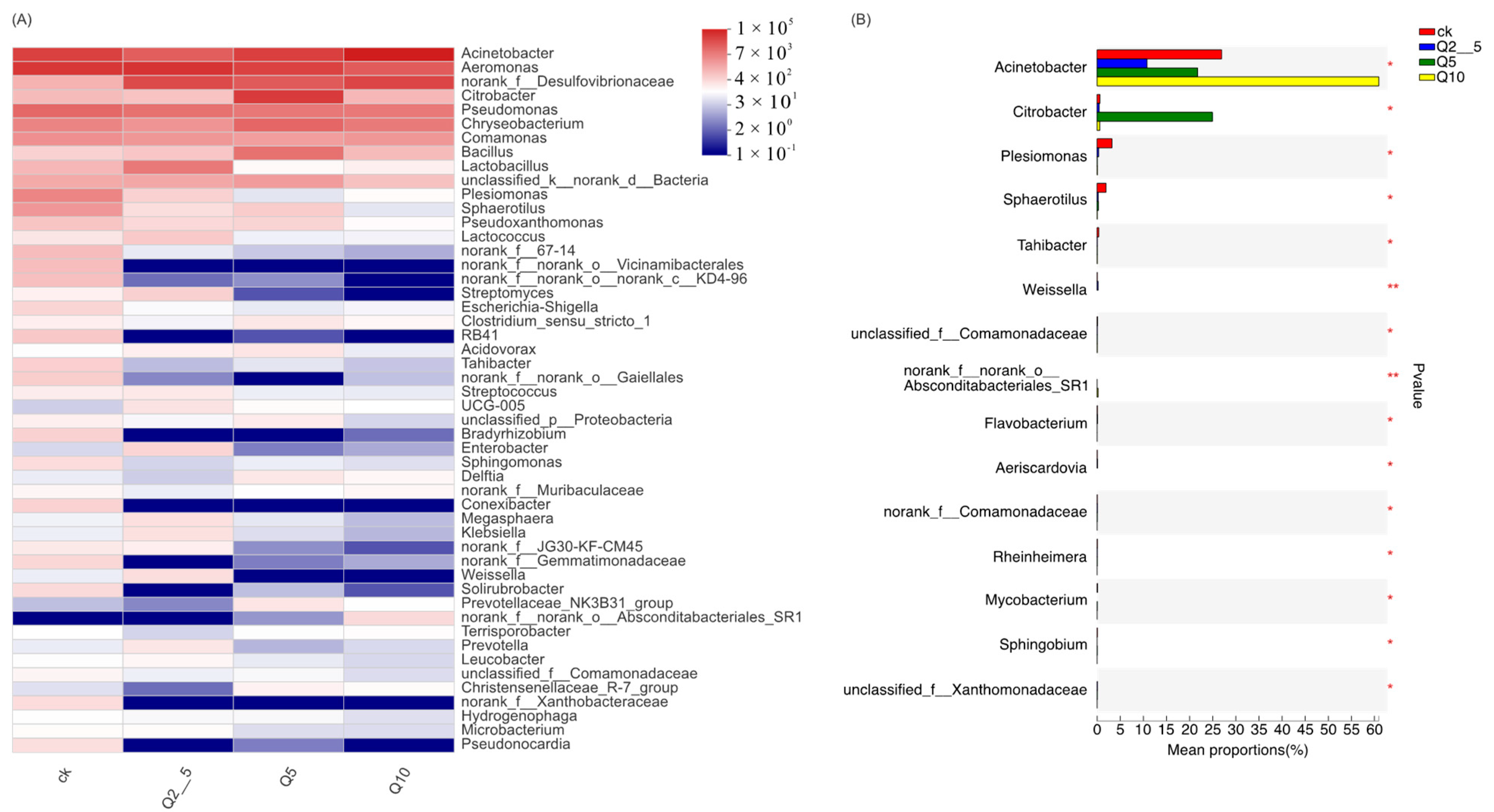
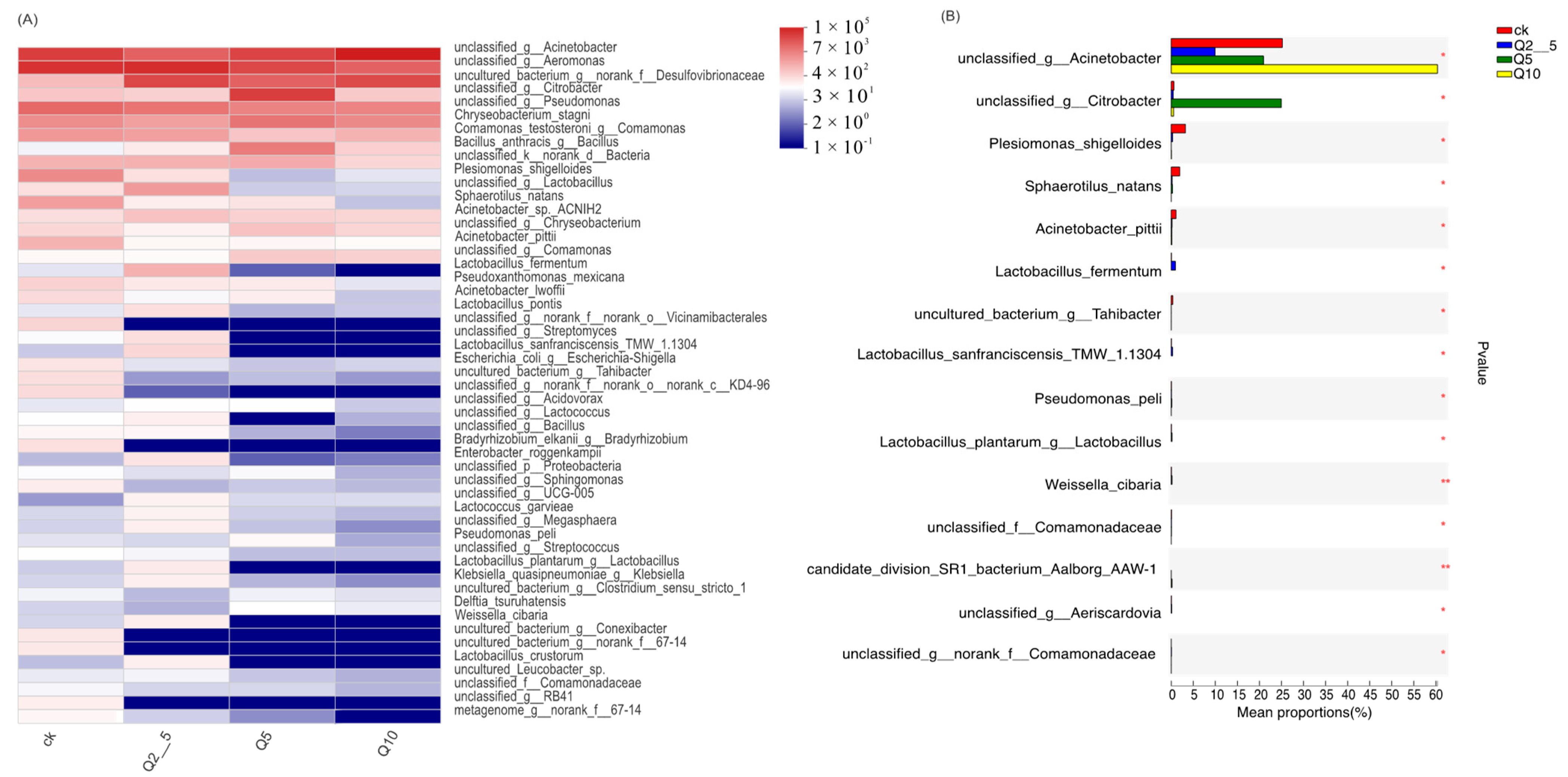
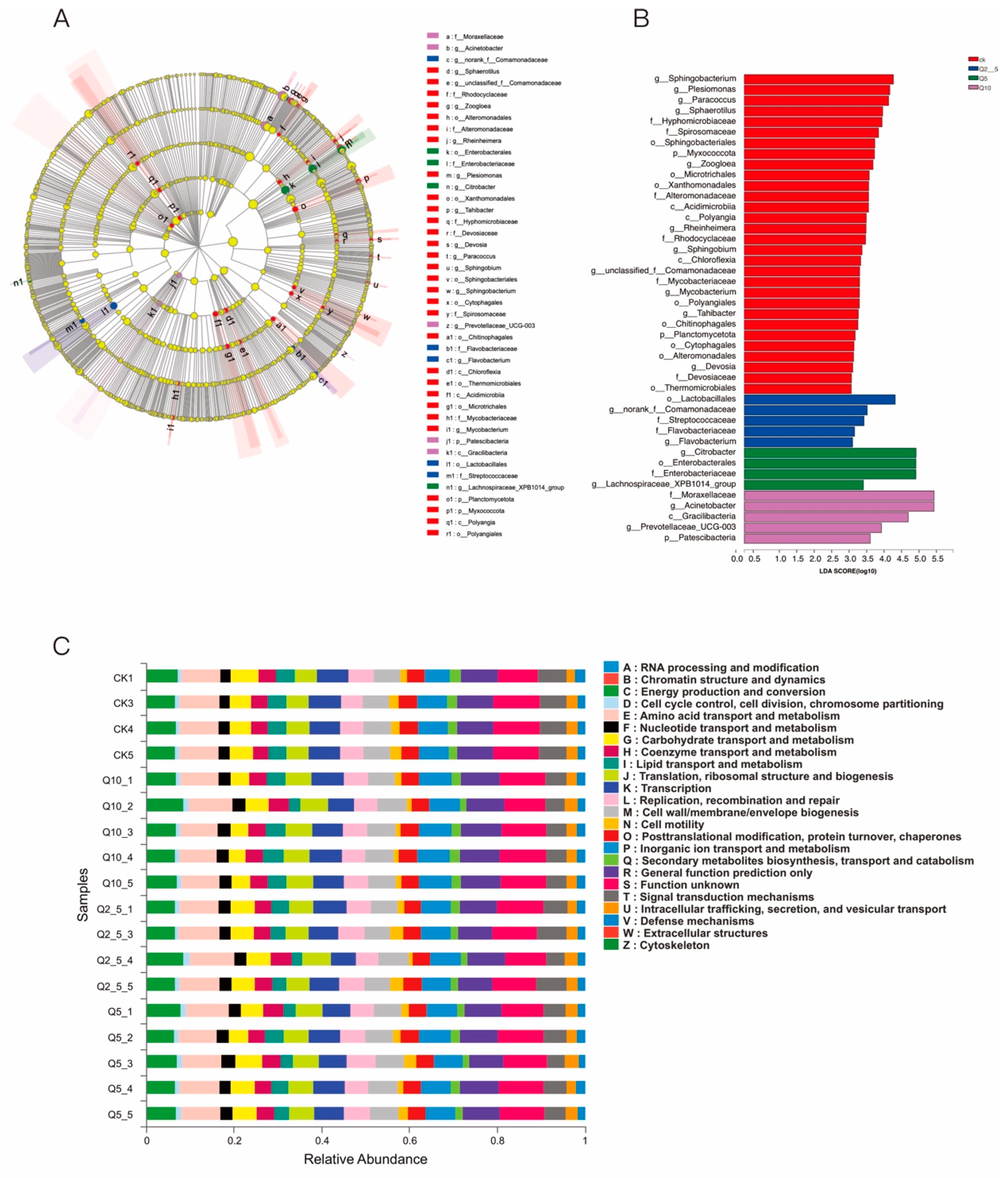
3.2. Species Analysis at the Genus Level
3.3. Analysis of the Biochemical Components
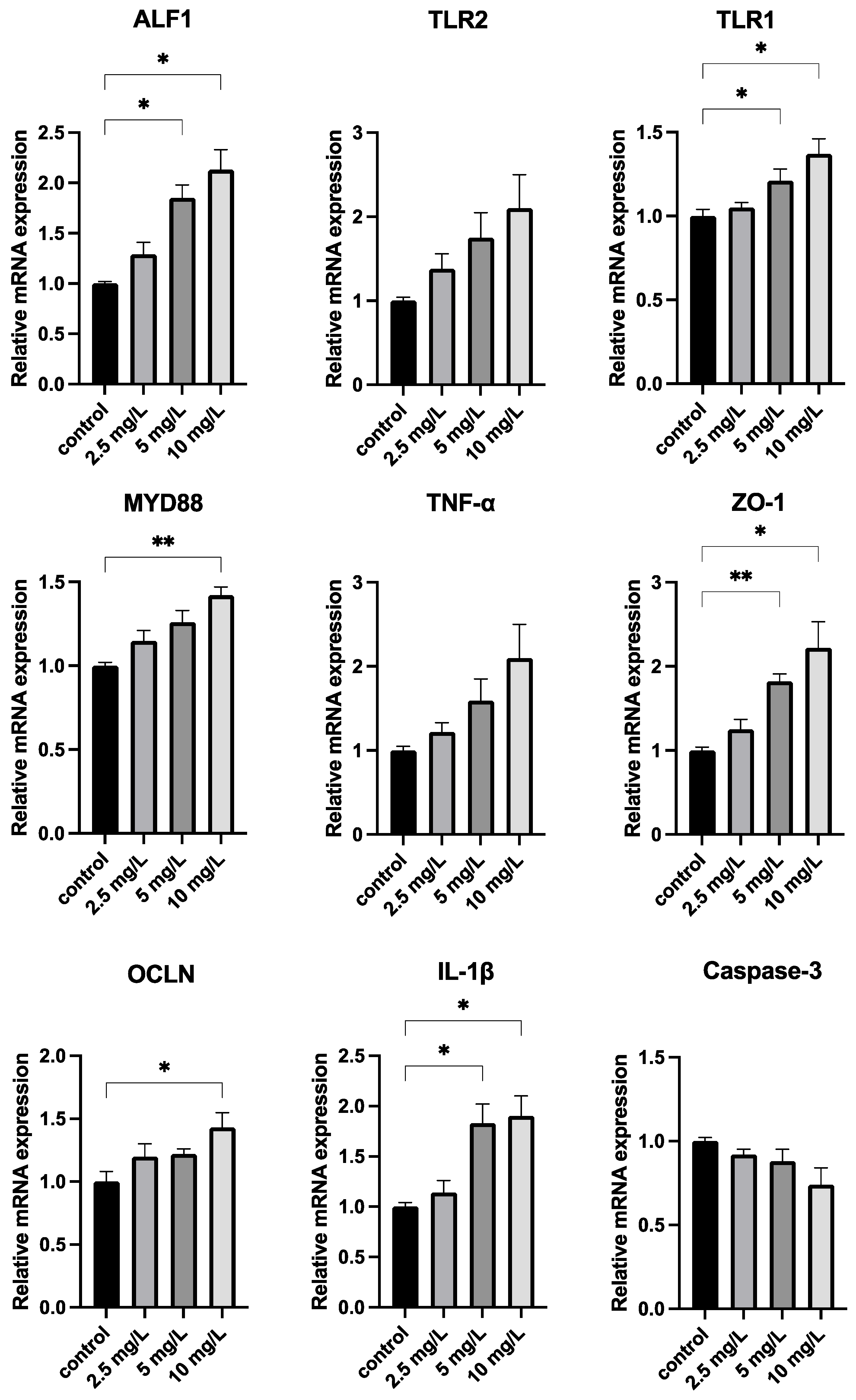
3.4. Quercetin Strengthens the Immune System
3.5. Linear Discriminant Analysis (Lda) Integrated with Effect Size (LEfSe)
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hoseinifar, S.H.; Hosseini, M.; Paknejad, H.; Safari, R.; Jafar, A.; Yousefi, M.; Van Doan, H.; Mozanzadeh, M.T. Enhanced mucosal immune responses, immune related genes and growth performance in common carp (Cyprinus carpio) juveniles fed dietary Pediococcus acidilactici MA18/5M and raffinose. Dev. Comp. Immunol. 2019, 94, 59–65. [Google Scholar] [CrossRef] [PubMed]
- Hoseini, S.M.; Yousefi, M. Beneficial effects of thyme (Thymus vulgaris) extract on oxytetracycline-induced stress response, immunosuppression, oxidative stress and enzymatic changes in rainbow trout (Oncorhynchus mykiss). Aquac. Nutr. 2019, 25, 298–309. [Google Scholar] [CrossRef]
- Omoregie, E.; Oyebanji, S.M. Oxytetracycline-Induced Blood Disorder in Juvenile Nile Tilapia Oreochromis niloticus (Trewavas). J. World Aquac. Soc. 2002, 33, 377–382. [Google Scholar] [CrossRef]
- Solomon, S.L.; Oliver, K.B. Antibiotic resistance threats in the United States: Stepping back from the brink. Am. Fam. Physician 2014, 89, 938–941. [Google Scholar]
- Casewell, M.; Friis, C.; Marco, E.; McMullin, P.; Phillips, I. The European ban on growth-promoting antibiotics and emerging consequences for human and animal health. J. Antimicrob. Chemother. 2003, 52, 159–161. [Google Scholar] [CrossRef] [Green Version]
- Piccinelli, A.L.; De Simone, F.; Passi, S.; Rastrelli, L. Phenolic constituents and antioxidant activity of Wendita calysina leaves (burrito), a folk Paraguayan tea. J. Agric. Food Chem. 2004, 52, 5863–5868. [Google Scholar] [CrossRef]
- Ren, W.; Qiao, Z.; Wang, H.; Zhu, L.; Zhang, L. Flavonoids: Promising anticancer agents. Med. Res. Rev. 2003, 23, 519–534. [Google Scholar] [CrossRef]
- Slimestad, R.; Fossen, T.; Vågen, I.M. Onions: A source of unique dietary flavonoids. J. Agric. Food Chem. 2007, 55, 10067–10080. [Google Scholar] [CrossRef]
- Wallace, S.N.; Carrier, D.J.; Clausen, E.C. Batch solvent extraction of flavanolignans from milk thistle (Silybum marianum L. Gaertner). Phytochem. Anal. Int. J. Plant Chem. Biochem. Tech. 2005, 16, 7–16. [Google Scholar] [CrossRef]
- Wamhoff, H.; Gribble, G.W. Wine and heterocycles. Adv. Heterocycl. Chem. 2012, 106, 185–225. [Google Scholar]
- Zhang, H.-Q.; Wang, Y.-J.; Yang, G.-T.; Gao, Q.-L.; Tang, M.-X. Taxifolin inhibits receptor activator of NF-κB ligand-induced osteoclastogenesis of human bone marrow-derived macrophages in vitro and prevents lipopolysaccharide-induced bone loss in vivo. Pharmacology 2019, 103, 101–109. [Google Scholar] [CrossRef]
- Kozhikkadan Davis, C.; Nasla, K.; Anjana, A.; Rajanikant, G. Taxifolin as dual inhibitor of Mtb DNA gyrase and isoleucyl-tRNA synthetase: In silico molecular docking, dynamics simulation and in vitro assays. Silico Pharmacol. 2018, 6, 8. [Google Scholar] [CrossRef]
- Tapas, A.R.; Sakarkar, D.; Kakde, R. Flavonoids as nutraceuticals: A review. Trop. J. Pharm. Res. 2008, 7, 1089–1099. [Google Scholar] [CrossRef] [Green Version]
- Shu, Z.; Yang, Y.; Yang, L.; Jiang, H.; Yu, X.; Wang, Y. Cardioprotective effects of dihydroquercetin against ischemia reperfusion injury by inhibiting oxidative stress and endoplasmic reticulum stress-induced apoptosis via the PI3K/Akt pathway. Food Funct. 2019, 10, 203–215. [Google Scholar] [CrossRef]
- Wasimul, H.; Shakti, P.; Barij, N. Evaluation of taxifolin and phloretin as antiangiogenic flavonoids: An in vivo, in vitro experimental analysis. J. Pharm. Pharm. Sci. 2015, 7, 72–79. [Google Scholar]
- Chen, X.; Gu, N.; Xue, C.; Li, B.R. Plant flavonoid taxifolin inhibits the growth, migration and invasion of human osteosarcoma cells. Mol. Med. Rep. 2018, 17, 3239–3245. [Google Scholar] [CrossRef] [Green Version]
- Pietsch, K.; Saul, N.; Menzel, R.; Stürzenbaum, S.R.; Steinberg, C.E. Quercetin mediated lifespan extension in Caenorhabditis elegans is modulated by age-1, daf-2, sek-1 and unc-43. Biogerontology 2009, 10, 565–578. [Google Scholar] [CrossRef]
- Kobori, M.; Masumoto, S.; Akimoto, Y.; Oike, H. Chronic dietary intake of quercetin alleviates hepatic fat accumulation associated with consumption of a Western-style diet in C57/BL6J mice. Mol. Nutr. Food Res. 2011, 55, 530–540. [Google Scholar] [CrossRef]
- Shoskes, D.A.; Zeitlin, S.I.; Shahed, A.; Rajfer, J. Quercetin in men with category III chronic prostatitis: A preliminary prospective, double-blind, placebo-controlled trial. Urology 1999, 54, 960–963. [Google Scholar] [CrossRef]
- Zhu, C.; Liu, G.; Gu, X.; Yin, J.; Xia, A.; Han, M.; Zhang, T.; Jiang, Q. Effect of quercetin on muscle growth and antioxidant status of the dark sleeper Odontobutis potamophila. Front. Genet. 2022, 13, 938526. [Google Scholar] [CrossRef]
- Tan, H.Y.; Chen, S.-W.; Hu, S.-Y. Improvements in the growth performance, immunity, disease resistance, and gut microbiota by the probiotic Rummeliibacillus stabekisii in Nile tilapia (Oreochromis niloticus). Fish Shellfish. Immunol. 2019, 92, 265–275. [Google Scholar] [CrossRef]
- Xiong, J.-B.; Nie, L.; Chen, J. Current understanding on the roles of gut microbiota in fish disease and immunity. Zool. Res. 2019, 40, 70. [Google Scholar] [PubMed] [Green Version]
- He, M.; Wu, T.; Pan, S.; Xu, X. Antimicrobial mechanism of flavonoids against Escherichia coli ATCC 25922 by model membrane study. Appl. Surf. Sci. 2014, 305, 515–521. [Google Scholar] [CrossRef]
- Porras, D.; Nistal, E.; Martínez-Flórez, S.; Pisonero-Vaquero, S.; Olcoz, J.L.; Jover, R.; González-Gallego, J.; García-Mediavilla, M.V.; Sánchez-Campos, S. Protective effect of quercetin on high-fat diet-induced non-alcoholic fatty liver disease in mice is mediated by modulating intestinal microbiota imbalance and related gut-liver axis activation. Free Radic. Biol. Med. 2017, 102, 188–202. [Google Scholar] [CrossRef]
- Steiner, T.; Syed, B. Phytogenic feed additives in animal nutrition. In Medicinal and Aromatic Plants of the World; Springer: Berlin/Heidelberg, Germany, 2015; pp. 403–423. [Google Scholar]
- Hou, X.; Zhu, F.; Yin, S.; Zhang, L.; Hu, Y.; Wang, Y.; Jia, Y.; Zhang, G.; Li, L. Genetic diversity of Odontobutis potamophila from different geographic populations inferred from mtDNA control region. Mitochondrial DNA 2014, 25, 400–406. [Google Scholar] [CrossRef]
- Jia, Y.; Zheng, J.; Liu, S.; Li, F.; Chi, M.; Cheng, S.; Gu, Z. A chromosome-level genome assembly of the dark sleeper Odontobutis potamophila. Genome Biol. Evol. 2021, 13, evaa271. [Google Scholar] [CrossRef]
- Wu, S.; Tian, J.; Wang, G.; Li, W.; Zou, H. Characterization of bacterial community in the stomach of yellow catfish (Pelteobagrus fulvidraco). World J. Microbiol. Biotechnol. 2012, 28, 2165–2174. [Google Scholar] [CrossRef] [PubMed]
- Kawabata, K.; Sugiyama, Y.; Sakano, T.; Ohigashi, H. Flavonols enhanced production of anti-inflammatory substance (s) by Bifidobacterium adolescentis: Prebiotic actions of galangin, quercetin, and fisetin. Biofactors 2013, 39, 422–429. [Google Scholar] [CrossRef]
- Duda-Chodak, A.; Tarko, T.; Satora, P.; Sroka, P. Interaction of dietary compounds, especially polyphenols, with the intestinal microbiota: A review. Eur. J. Nutr. 2015, 54, 325–341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, S.; Liu, Y.; Zhou, Z.; He, S.; Cao, Y.; Shi, P.; Yao, B.; Ringø, E. Analysis of bacterial diversity in the intestine of grass carp (Ctenopharyngodon idellus) based on 16S rDNA gene sequences. Aquac. Res. 2010, 42, 47–56. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.H.; Brunt, J.; Austin, B. Microbial diversity of intestinal contents and mucus in rainbow trout (Oncorhynchus mykiss). J. Appl. Microbiol. 2007, 102, 1654–1664. [Google Scholar] [CrossRef]
- Shin, N.-R.; Whon, T.W.; Bae, J.-W. Proteobacteria: Microbial signature of dysbiosis in gut microbiota. Trends Biotechnol. 2015, 33, 496–503. [Google Scholar] [CrossRef]
- Zhu, L.; Baker, S.S.; Gill, C.; Liu, W.; Alkhouri, R.; Baker, R.D.; Gill, S.R. Characterization of gut microbiomes in nonalcoholic steatohepatitis (NASH) patients: A connection between endogenous alcohol and NASH. Hepatology 2013, 57, 601–609. [Google Scholar] [CrossRef]
- Verschuere, L.; Rombaut, G.; Sorgeloos, P.; Verstraete, W. Probiotic bacteria as biological control agents in aquaculture. Microbiol. Mol. Biol. Rev. 2000, 64, 655–671. [Google Scholar] [CrossRef] [Green Version]
- Al-Muzafar, H.M.; Amin, K.A. Probiotic mixture improves fatty liver disease by virtue of its action on lipid profiles, leptin, and inflammatory biomarkers. BMC Complement. Altern. Med. 2017, 17, 43. [Google Scholar] [CrossRef] [Green Version]
- Do, H.J.; Chung, J.H.; Hwang, J.W.; Kim, O.Y.; Lee, J.-Y.; Shin, M.-J. 1-Deoxynojirimycin isolated from Bacillus subtilis improves hepatic lipid metabolism and mitochondrial function in high-fat–fed mice. Food Chem. Toxicol. 2015, 75, 1–7. [Google Scholar] [CrossRef]
- Kobyliak, N.; Conte, C.; Cammarota, G.; Haley, A.P.; Styriak, I.; Gaspar, L.; Fusek, J.; Rodrigo, L.; Kruzliak, P. Probiotics in prevention and treatment of obesity: A critical view. Nutr. Metab. 2016, 13, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blain Kennedy, S.; Tucker, J.W., Jr.; Neidig, C.L.; Vermeer, G.K.; Cooper, V.R.; Jarrell, J.L.; Sennett, D.G. Bacterial management strategies for stock enhancement of warmwater marine fish: A case study with common snook (Centropomus undecimalis). Bull. Mar. Sci. 1998, 62, 573–588. [Google Scholar]
- Hartviksen, M.; Vecino, J.; Ringø, E.; Bakke, A.M.; Wadsworth, S.; Krogdahl, Å.; Ruohonen, K.; Kettunen, A. Alternative dietary protein sources for A tlantic salmon (Salmo salar L.) effect on intestinal microbiota, intestinal and liver histology and growth. Aquac. Nutr. 2014, 20, 381–398. [Google Scholar] [CrossRef] [Green Version]
- Velmurugan, S.; Rajagopal, S. Beneficial uses of probiotics in mass scale production of marine ornamental fish. Afr. J. Microbiol. Res. 2009, 3, 185–190. [Google Scholar]
- Benhamed, F.; Denechaud, P.-D.; Lemoine, M.; Robichon, C.; Moldes, M.; Bertrand-Michel, J.; Ratziu, V.; Serfaty, L.; Housset, C.; Capeau, J. The lipogenic transcription factor ChREBP dissociates hepatic steatosis from insulin resistance in mice and humans. J. Clin. Investig. 2012, 122, 2176–2194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fabbrini, E.; Sullivan, S.; Klein, S. Obesity and nonalcoholic fatty liver disease: Biochemical, metabolic, and clinical implications. Hepatology 2010, 51, 679–689. [Google Scholar] [CrossRef] [Green Version]
- Jung, C.H.; Cho, I.; Ahn, J.; Jeon, T.I.; Ha, T.Y. Quercetin reduces high-fat diet-induced fat accumulation in the liver by regulating lipid metabolism genes. Phytother. Res. 2013, 27, 139–143. [Google Scholar] [CrossRef]
- Spiegelman, B.M. Transcriptional control of mitochondrial energy metabolism through the PGC1 coactivators. In Novartis Foundation Symposium, Chichester; John Wiley: New York, NY, USA, 2007; p. 60. [Google Scholar]
- Tsuduki, T.; Kikuchi, I.; Kimura, T.; Nakagawa, K.; Miyazawa, T. Intake of mulberry 1-deoxynojirimycin prevents diet-induced obesity through increases in adiponectin in mice. Food Chem. 2013, 139, 16–23. [Google Scholar] [CrossRef]
- Li, Y.-G.; Ji, D.-F.; Zhong, S.; Lv, Z.-Q.; Lin, T.-B. Cooperative anti-diabetic effects of deoxynojirimycin-polysaccharide by inhibiting glucose absorption and modulating glucose metabolism in streptozotocin-induced diabetic mice. PLoS ONE 2013, 8, e65892. [Google Scholar] [CrossRef]
- Duda-Chodak, A. The inhibitory effect of polyphenols on human gut microbiota. J. Physiol. Pharmacol. 2012, 63, 497–503. [Google Scholar]
- Zhang, Y.; Wen, B.; David, M.A.; Gao, J.-Z.; Chen, Z.-Z. Comparative analysis of intestinal microbiota of discus fish (Symphysodon haraldi) with different growth rates. Aquaculture 2021, 540, 736740. [Google Scholar] [CrossRef]
- Alexander, J.; Knopp, G.; Dötsch, A.; Wieland, A.; Schwartz, T. Ozone treatment of conditioned wastewater selects antibiotic resistance genes, opportunistic bacteria, and induce strong population shifts. Sci. Total Environ. 2016, 559, 103–112. [Google Scholar] [CrossRef]
- Tsugawa, H.; Ogawa, A.; Takehara, S.; Kimura, M.; Okawa, Y. Primary structure and function of a cytotoxic outer-membrane protein (ComP) of Plesiomonas shigelloides. FEMS Microbiol. Lett. 2008, 281, 10–16. [Google Scholar] [CrossRef]
- Bodhidatta, L.; McDaniel, P.; Sornsakrin, S.; Srijan, A.; Serichantalergs, O.; Mason, C.J. Case-control study of diarrheal disease etiology in a remote rural area in Western Thailand. Am. J. Trop. Med. Hyg. 2010, 83, 1106. [Google Scholar] [CrossRef] [Green Version]
- van Kessel, M.A.; Dutilh, B.E.; Neveling, K.; Kwint, M.P.; Veltman, J.A.; Flik, G.; Jetten, M.S.; Klaren, P.H.; Op den Camp, H.J. Pyrosequencing of 16S rRNA gene amplicons to study the microbiota in the gastrointestinal tract of carp (Cyprinus carpio L.). AMB Express 2011, 1, 41. [Google Scholar] [CrossRef] [Green Version]
- Wu, S.; Wang, G.; Angert, E.R.; Wang, W.; Li, W.; Zou, H. Composition, diversity, and origin of the bacterial community in grass carp intestine. PLoS ONE 2012, 7, e30440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swathy, J.S.; Mishra, P.; Thomas, J.; Mukherjee, A.; Chandrasekaran, N. Nanometric neem oil emulsification through microfluidization, and its therapeutic potential against Aeromonas culicicola infection in Cyprinus carpio. Flavour Fragr. J. 2018, 33, 340–350. [Google Scholar] [CrossRef]
- Asakura, H.; Makino, S.-I.; Watanabe, K.; Tuchida, Y.; Kawabe, M.; Sakurai, D. Kuma bamboo grass (Sasa veitchii) extracts exhibit protective effects against atypical Aeromonas salmonicida infection in goldfish (Carassius auratus). Biocontrol Sci. 2019, 24, 145–154. [Google Scholar] [CrossRef] [Green Version]
- Chassaing, B.; Koren, O.; Carvalho, F.A.; Ley, R.E.; Gewirtz, A.T. AIEC pathobiont instigates chronic colitis in susceptible hosts by altering microbiota composition. Gut 2014, 63, 1069–1080. [Google Scholar] [CrossRef] [PubMed]
- Mittal, M.; Siddiqui, M.R.; Tran, K.; Reddy, S.P.; Malik, A.B. Reactive oxygen species in inflammation and tissue injury. Antioxid. Redox Signal. 2014, 20, 1126–1167. [Google Scholar] [CrossRef]
- Nair, S.; Rocha-Ferreira, E.; Fleiss, B.; Nijboer, C.H.; Gressens, P.; Mallard, C.; Hagberg, H. Neuroprotection offered by mesenchymal stem cells in perinatal brain injury: Role of mitochondria, inflammation, and reactive oxygen species. J. Neurochem. 2021, 158, 59–73. [Google Scholar] [CrossRef]
- Al-Baqami, N.; Hamza, R. Synergistic antioxidant capacities of vanillin and chitosan nanoparticles against reactive oxygen species, hepatotoxicity, and genotoxicity induced by aging in male Wistar rats. Hum. Exp. Toxicol. 2021, 40, 183–202. [Google Scholar] [CrossRef] [PubMed]
- Bu, T.; Mi, Y.; Zeng, W.; Zhang, C. Protective effect of quercetin on cadmium-induced oxidative toxicity on germ cells in male mice. Anat. Rec. Adv. Integr. Anat. Evol. Biol. 2011, 294, 520–526. [Google Scholar] [CrossRef]
- Farombi, E.; Adedara, I.; Akinrinde, S.; Ojo, O.; Eboh, A. Protective effects of kolaviron and quercetin on cadmium-induced testicular damage and endocrine pathology in rats. Andrologia 2012, 44, 273–284. [Google Scholar] [CrossRef]
- Zhang, C.; Jiang, D.; Wang, J.; Qi, Q. The effects of TPT and dietary quercetin on growth, hepatic oxidative damage and apoptosis in zebrafish. Ecotoxicol. Environ. Saf. 2021, 224, 112697. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wang, L.; Wang, L.; Yang, J.; Gai, Y.; Qiu, L.; Song, L. The second anti-lipopolysaccharide factor (EsALF-2) with antimicrobial activity from Eriocheir sinensis. Dev. Comp. Immunol. 2010, 34, 945–952. [Google Scholar] [CrossRef] [PubMed]
- Molteni, M.; Gemma, S.; Rossetti, C. The role of toll-like receptor 4 in infectious and noninfectious inflammation. Mediat. Inflamm. 2016, 2016, 6978936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, D.; Zhang, G.; Hayden, M.S.; Greenblatt, M.B.; Bussey, C.; Flavell, R.A.; Ghosh, S. A toll-like receptor that prevents infection by uropathogenic bacteria. Science 2004, 303, 1522–1526. [Google Scholar] [CrossRef]
- Iwasaki, A.; Medzhitov, R. Toll-like receptor control of the adaptive immune responses. Nat. Immunol. 2004, 5, 987–995. [Google Scholar] [CrossRef]
- Muhammad, T.; Ikram, M.; Ullah, R.; Rehman, S.U.; Kim, M.O. Hesperetin, a citrus flavonoid, attenuates LPS-induced neuroinflammation, apoptosis and memory impairments by modulating TLR4/NF-κB signaling. Nutrients 2019, 11, 648. [Google Scholar] [CrossRef]
- Gonzalez-Mariscal, L.; Betanzos, A.; Nava, P.; Jaramillo, B. Tight junction proteins. Prog. Biophys. Mol. Biol. 2003, 81, 1–44. [Google Scholar] [CrossRef]
- Ugalde-Silva, P.; Gonzalez-Lugo, O.; Navarro-Garcia, F. Tight junction disruption induced by type 3 secretion system effectors injected by enteropathogenic and enterohemorrhagic Escherichia coli. Front. Cell. Infect. Microbiol. 2016, 6, 87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, W.; Liu, M.; Li, Y.; Yu, H.; Wang, D.; Chen, Q.; Chen, Y.; Zhang, Y.; Wang, T. Flavonoids from Citrus aurantium ameliorate TNBS-induced ulcerative colitis through protecting colonic mucus layer integrity. Eur. J. Pharmacol. 2019, 857, 172456. [Google Scholar] [CrossRef] [PubMed]
- Noda, S.; Tanabe, S.; Suzuki, T. Differential effects of flavonoids on barrier integrity in human intestinal Caco-2 cells. J. Agric. Food Chem. 2012, 60, 4628–4633. [Google Scholar] [CrossRef] [PubMed]
- Kosińska, A.; Andlauer, W. Modulation of tight junction integrity by food components. Food Res. Int. 2013, 54, 951–960. [Google Scholar] [CrossRef]
- Secombes, C.; Wang, T.; Hong, S.; Peddie, S.; Crampe, M.; Laing, K.; Cunningham, C.; Zou, J. Cytokines and innate immunity of fish. Dev. Comp. Immunol. 2001, 25, 713–723. [Google Scholar] [CrossRef]
- Wang, J.-L.; Meng, X.-l.; Lu, R.-h.; Wu, C.; Luo, Y.-T.; Yan, X.; Li, X.-J.; Kong, X.-H.; Nie, G.-X. Effects of Rehmannia glutinosa on growth performance, immunological parameters and disease resistance to Aeromonas hydrophila in common carp (Cyprinus carpio L.). Aquaculture 2015, 435, 293–300. [Google Scholar] [CrossRef]
- Watanuki, H.; Ota, K.; Tassakka, A.C.M.A.; Kato, T.; Sakai, M. Immunostimulant effects of dietary Spirulina platensis on carp, Cyprinus carpio. Aquaculture 2006, 258, 157–163. [Google Scholar] [CrossRef]
- Xian, J.-A.; Miao, Y.-T.; Li, B.; Guo, H.; Wang, A.-L. Apoptosis of tiger shrimp (Penaeus monodon) haemocytes induced by Escherichia coli lipopolysaccharide. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2013, 164, 301–306. [Google Scholar] [CrossRef]
- Choudhary, G.S.; Al-Harbi, S.; Almasan, A. Caspase-3 activation is a critical determinant of genotoxic stress-induced apoptosis. In Apoptosis and Cancer; Springer: Berlin/Heidelberg, Germany, 2015; pp. 1–9. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Sequence (5′ to 3′) |
|---|---|
| ALF1 F | CTGCTTCCTGCATGGACCTA |
| ALF1 R | CACTGAACTCTCCCGCTTGT |
| TLR1 F | CAAAACGTCCAAAGTGCGGT |
| TLR1 R | AGTCTCCACGATTGCCAGTG |
| TLR2 F | ACTCTGAGCCGCTGATAACG |
| TLR2 R | CCAGGTTCATCCGAGTCACC |
| MYD88 F | CGCTGAGCTCATGGGATTCT |
| MYD88 R | TCAGGGCTCGTCCAGTATGA |
| OCLN F | TCAGGGCTCGTCCAGTATGA |
| OCLN R | AACCAGGAAGCCACAAACCA |
| ZO-1 F | CGCAGGTAGACGGCTCTAAA |
| ZO-1 R | GAGCTGATTGGTCTCCGTCC |
| TNF F | ACCGTAACAACGTGCCTCAT |
| TNF R | ACTGGCTTTCAGGACTGTCG |
| IL1B F | CCGAGGCACACTTGAAGACT |
| IL1B R | GAGTCCGGCTCACACATCTC |
| Caspase-3 F | AGAGCGTCATATACGGCACG |
| Caspase-3 R | CCTCTATGTCTTCGTCCGGC |
| Beta-actin F | CTCTTCCAGCCATCCTTCCT |
| Beta-actin R | TCAGGTGGGGCAATGATCTT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, C.; Liu, G.; Gu, X.; Zhang, T.; Xia, A.; Zheng, Y.; Yin, J.; Han, M.; Jiang, Q. Effects of Quercetin on the Intestinal Microflora of Freshwater Dark Sleeper Odontobutis potamophila. Antioxidants 2022, 11, 2015. https://doi.org/10.3390/antiox11102015
Zhu C, Liu G, Gu X, Zhang T, Xia A, Zheng Y, Yin J, Han M, Jiang Q. Effects of Quercetin on the Intestinal Microflora of Freshwater Dark Sleeper Odontobutis potamophila. Antioxidants. 2022; 11(10):2015. https://doi.org/10.3390/antiox11102015
Chicago/Turabian StyleZhu, Chenxi, Guoxing Liu, Xiankun Gu, Tongqing Zhang, Aijun Xia, You Zheng, Jiawen Yin, Mingming Han, and Qichen Jiang. 2022. "Effects of Quercetin on the Intestinal Microflora of Freshwater Dark Sleeper Odontobutis potamophila" Antioxidants 11, no. 10: 2015. https://doi.org/10.3390/antiox11102015