
Evaluation of the Anxiolytic and Anti-Epileptogenic Potential of Lactuca Serriola Seed Using Pentylenetetrazol-Induced Kindling in Mice and Metabolic Profiling of Its Bioactive Extract

 and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Preparation of the Different Plant Extracts
2.3. In Vivo Biological Evaluation of Different L. serriola Dried Seeds Extracts
2.3.1. Experimental Animals
2.3.2. Acute Toxicity Study
2.3.3. In Vivo Anxiolytic Activity Evaluation of Different L. serriola Dried Seeds Extracts
Hole-Board Test
Elevated Plus Maze Test
Light/Dark Box Test
2.3.4. In Vivo Sedative Activity Evaluation of Different L. serriola Dried Seeds Extracts
2.3.5. In Vivo Anti-Convulsive Activity Evaluation of Different L. serriola Dried Seeds Extracts
2.3.6. In Vivo Anti-Epileptic Activity Evaluation of Different L. serriola Dried Seeds Extracts
Pentylenetetrazol (PTZ) Induced Kindling in Mice
Evaluation of the Biochemical Parameters in Brain Tissue
Assessment of Catalase (CAT) Activity in Brain
Assessment of Malondialdehyde (MDA) Level in Brain
Assessment of Superoxide Dismutase (SOD) Level in Brain
Assessment of Total Glutathione (tGSH) Activity in Brain
2.3.7. Histopathological Examination
2.4. Phytochemical Analysis of Different L. serriola Dried Seeds Extracts
2.4.1. Estimation of Total Phenol Content
2.4.2. Estimation of Total Flavonoid Content
2.4.3. Metabolic Profiling of L. serriola Dried Seeds Chloroform Extract by GC/MS
2.5. Molecular Docking Study
2.6. Statistical Analysis
3. Results
3.1. In Vivo Biological Evaluation of Different L. serriola Dried Seeds Extracts
3.1.1. Acute Toxicity Study
3.1.2. In Vivo Anxiolytic Activity Evaluation of Different L. serriola Dried Seeds Extracts
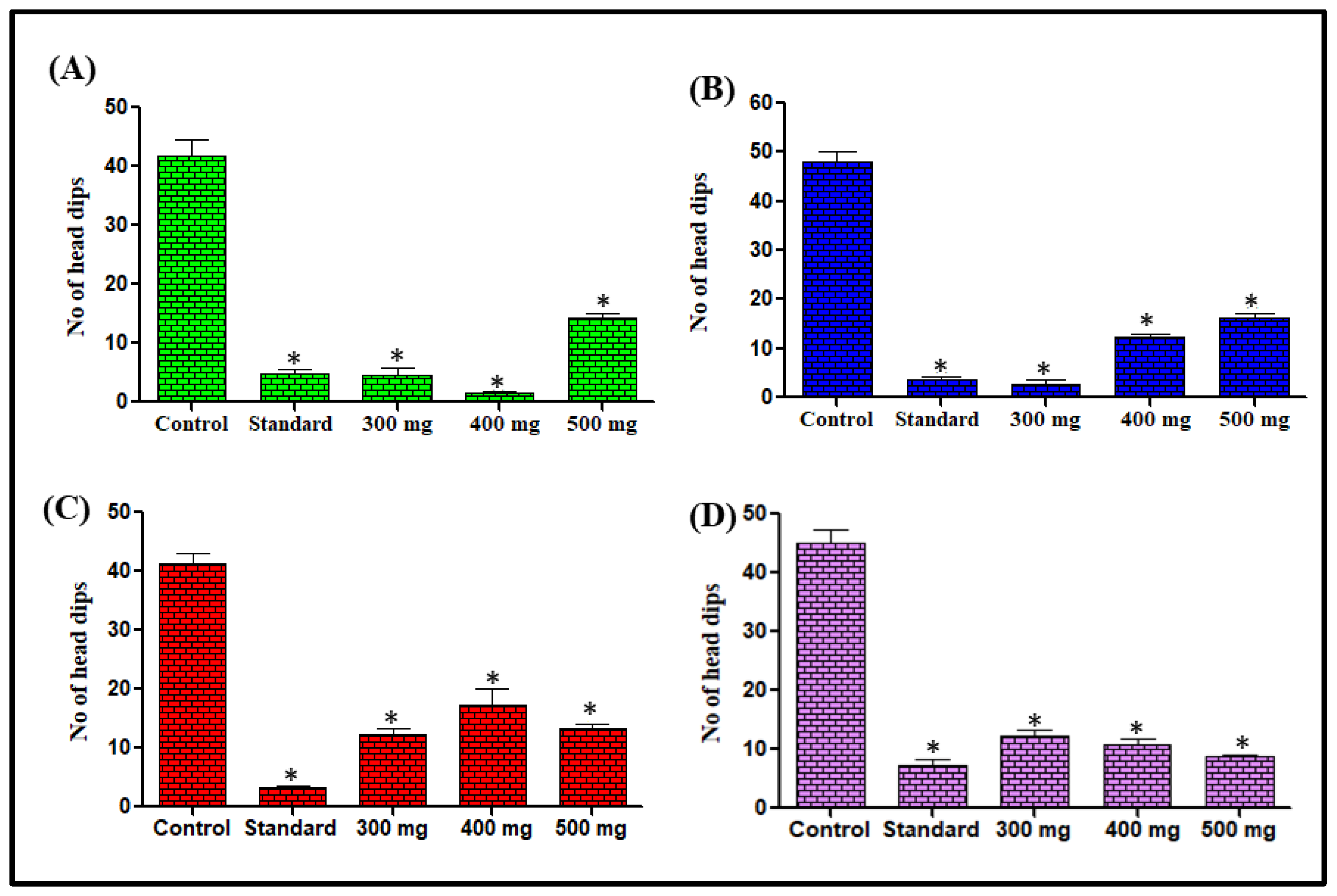
Hole-Board Test and Effect on Number of Head Dips
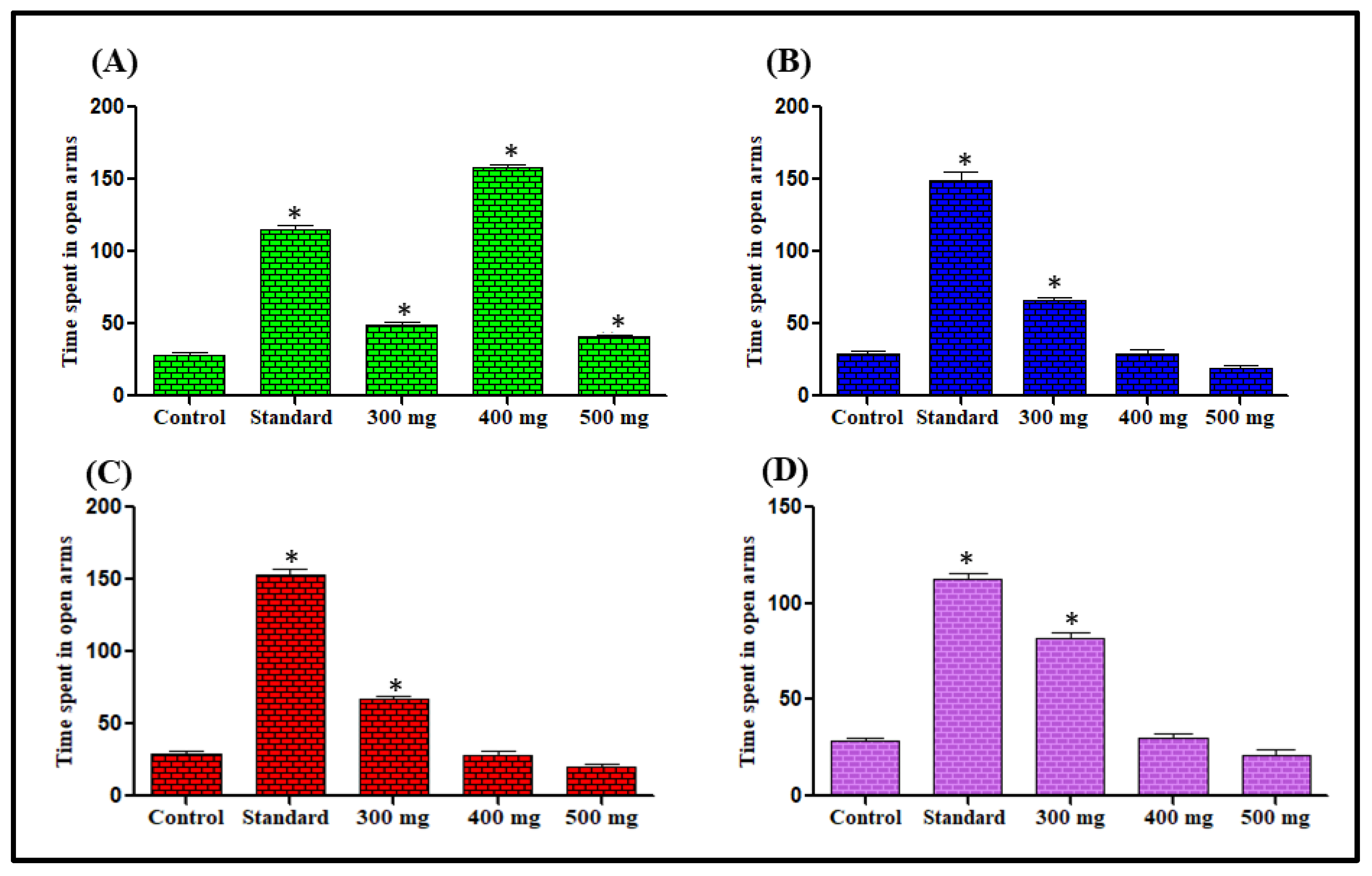
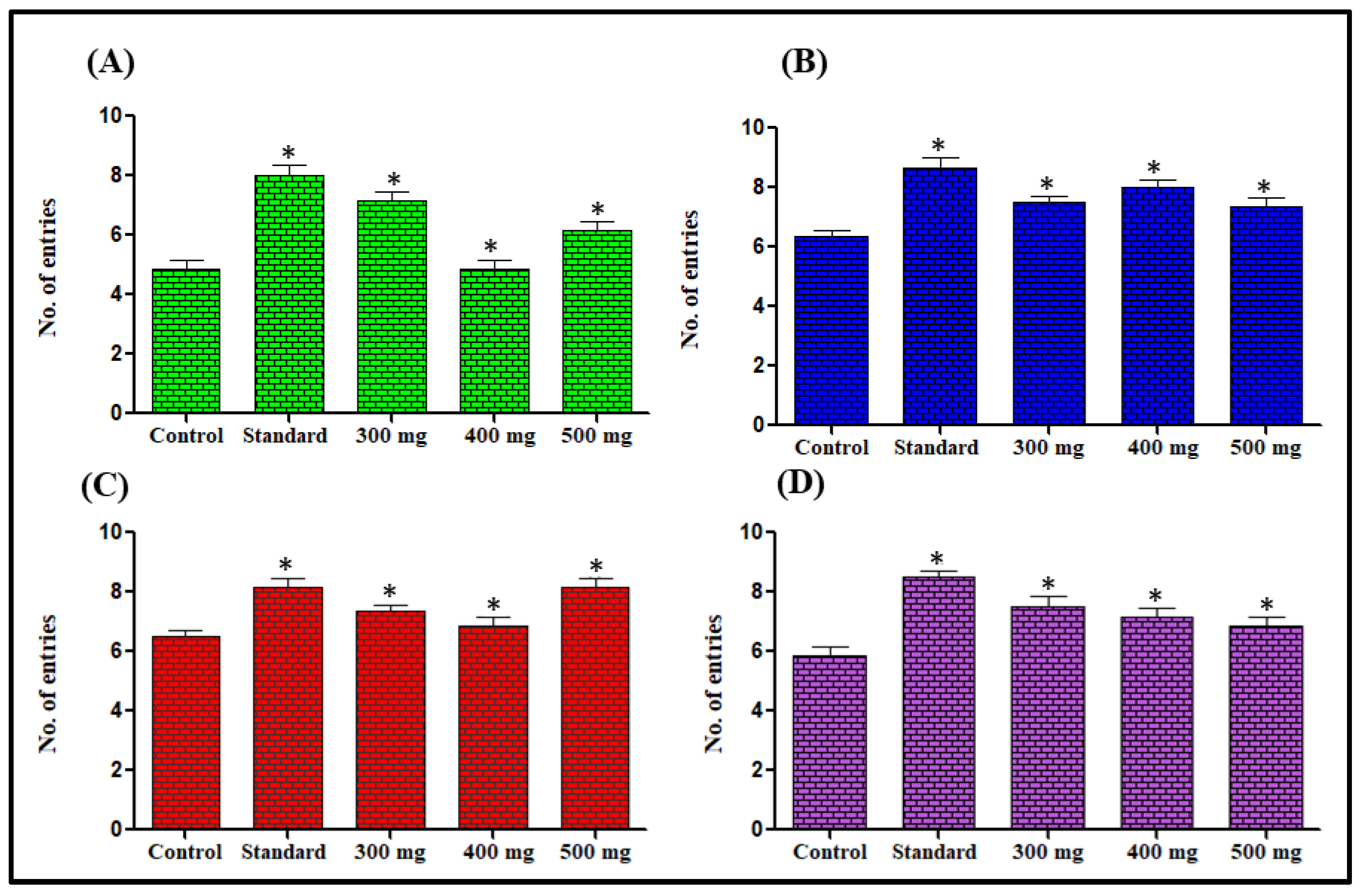
Effect on Time Spent in the Elevated Plus Maze
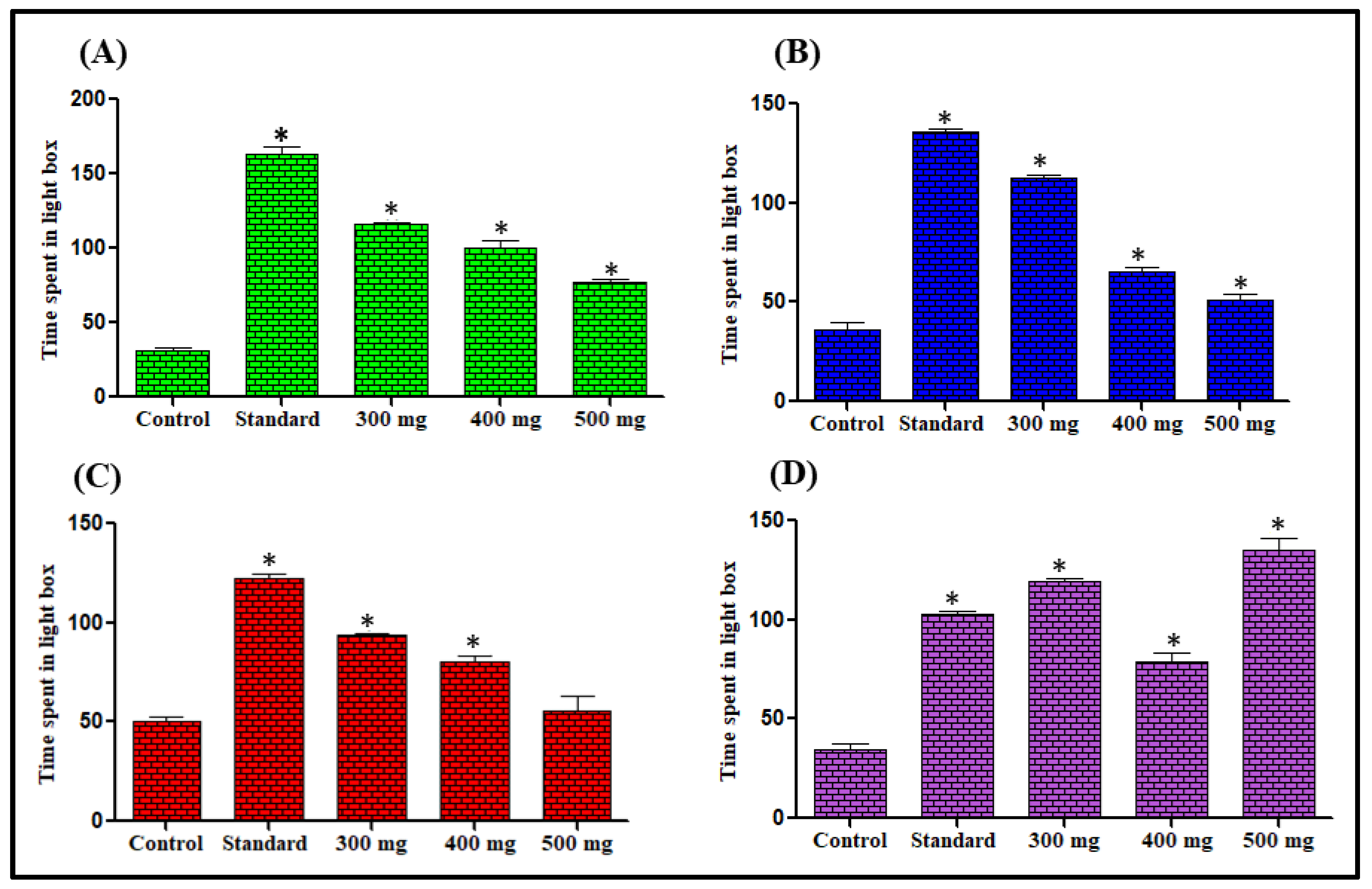
Effect on Time Spent in Light Dark Box
3.1.3. In Vivo Sedative Activity Evaluation of Different L. serriola Dried Seeds Extracts
3.1.4. In Vivo Anti-Convulsive Activity Evaluation of Different L. serriola Dried Seeds Extracts
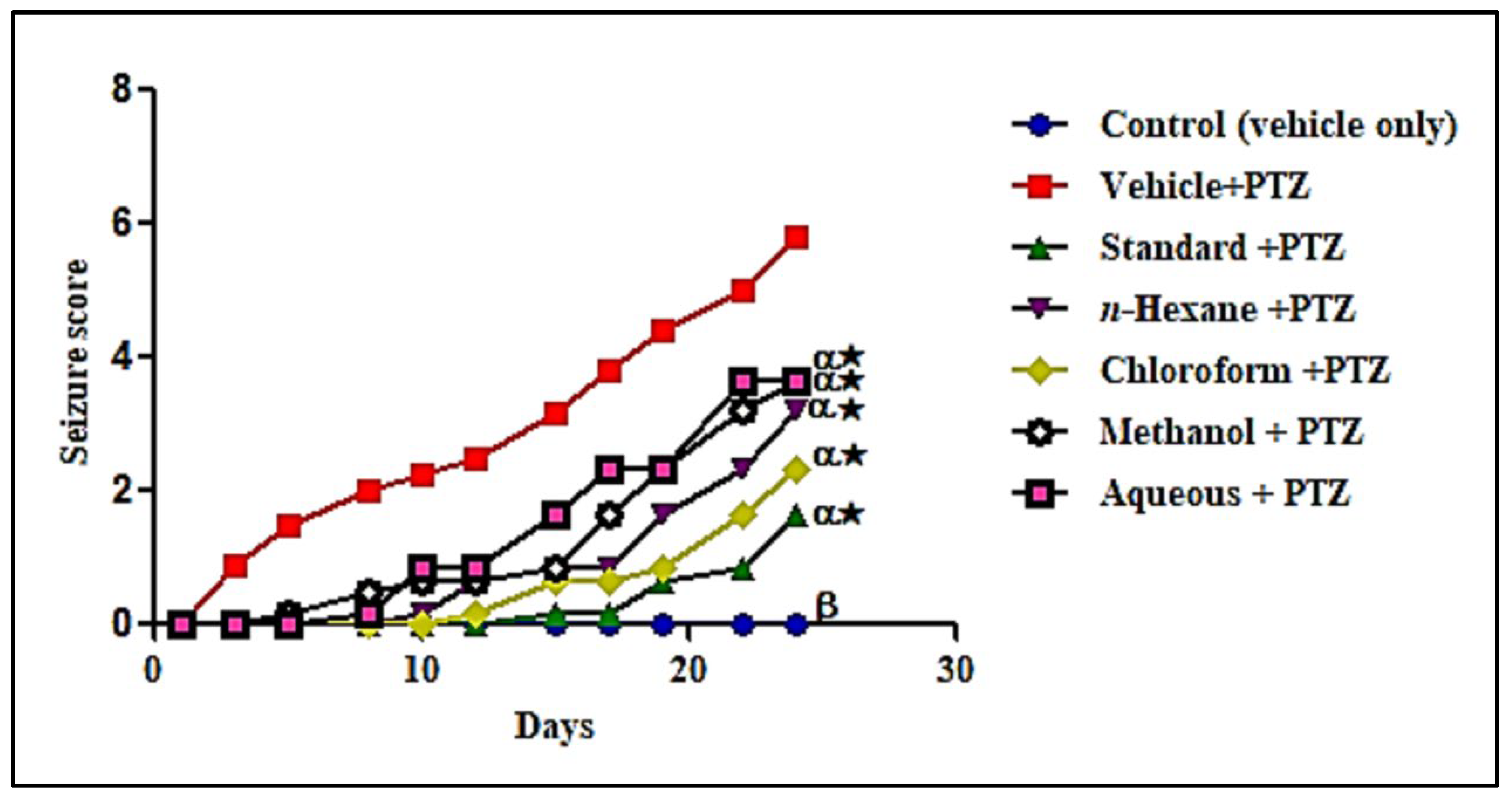
3.1.5. In Vivo Anti-Epileptic Activity Evaluation of Different L. serriola Dried Seeds Extracts
Effect of Different L. serriola Dried Seeds Extracts on Various Oxidative Stress Markers in Brain Tissues
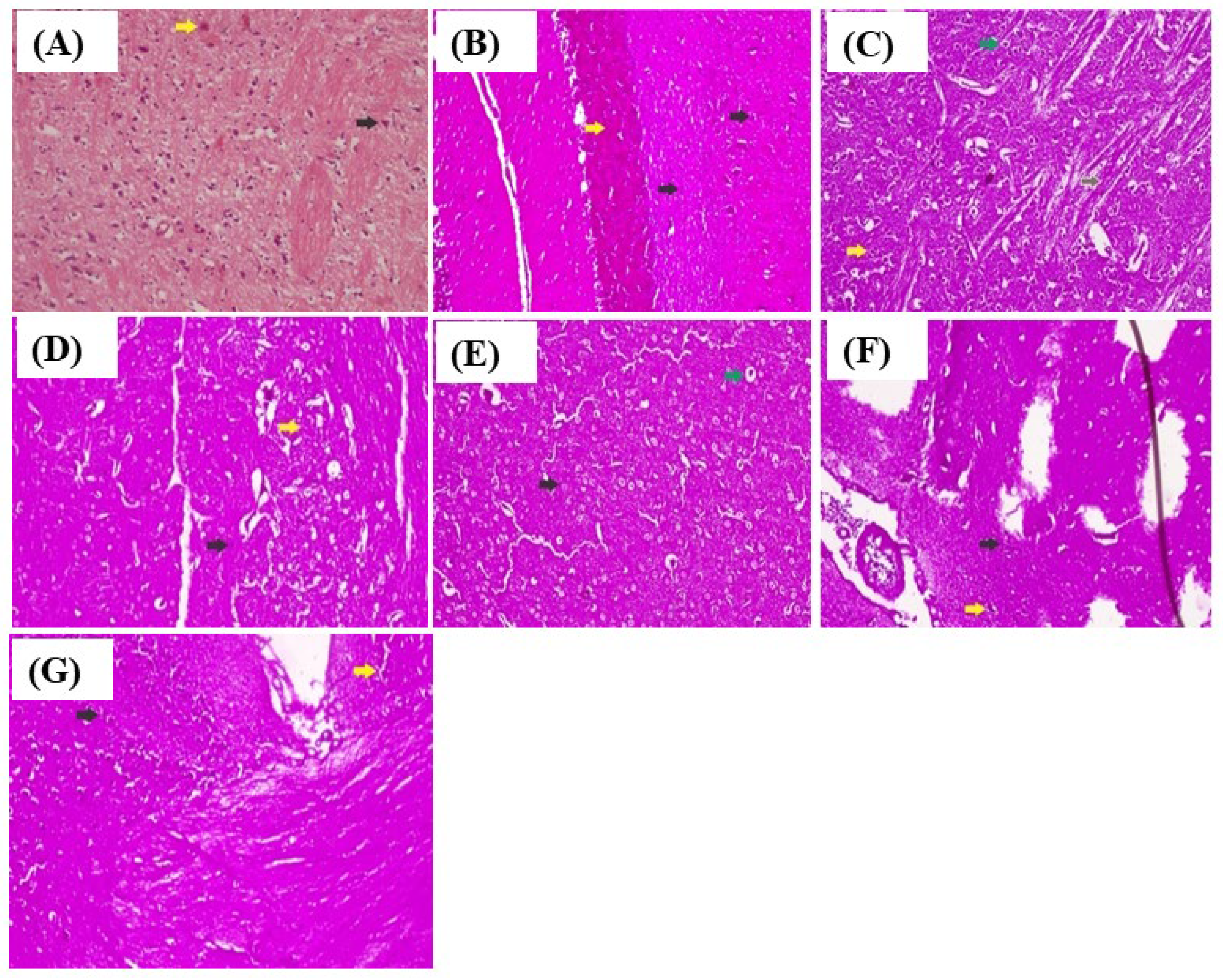
3.1.6. Histopathological Examination
3.2. Phytochemical Analysis of Different L. serriola Dried Seeds Extracts
3.2.1. Estimation of Total Flavonoid and Phenol Contents
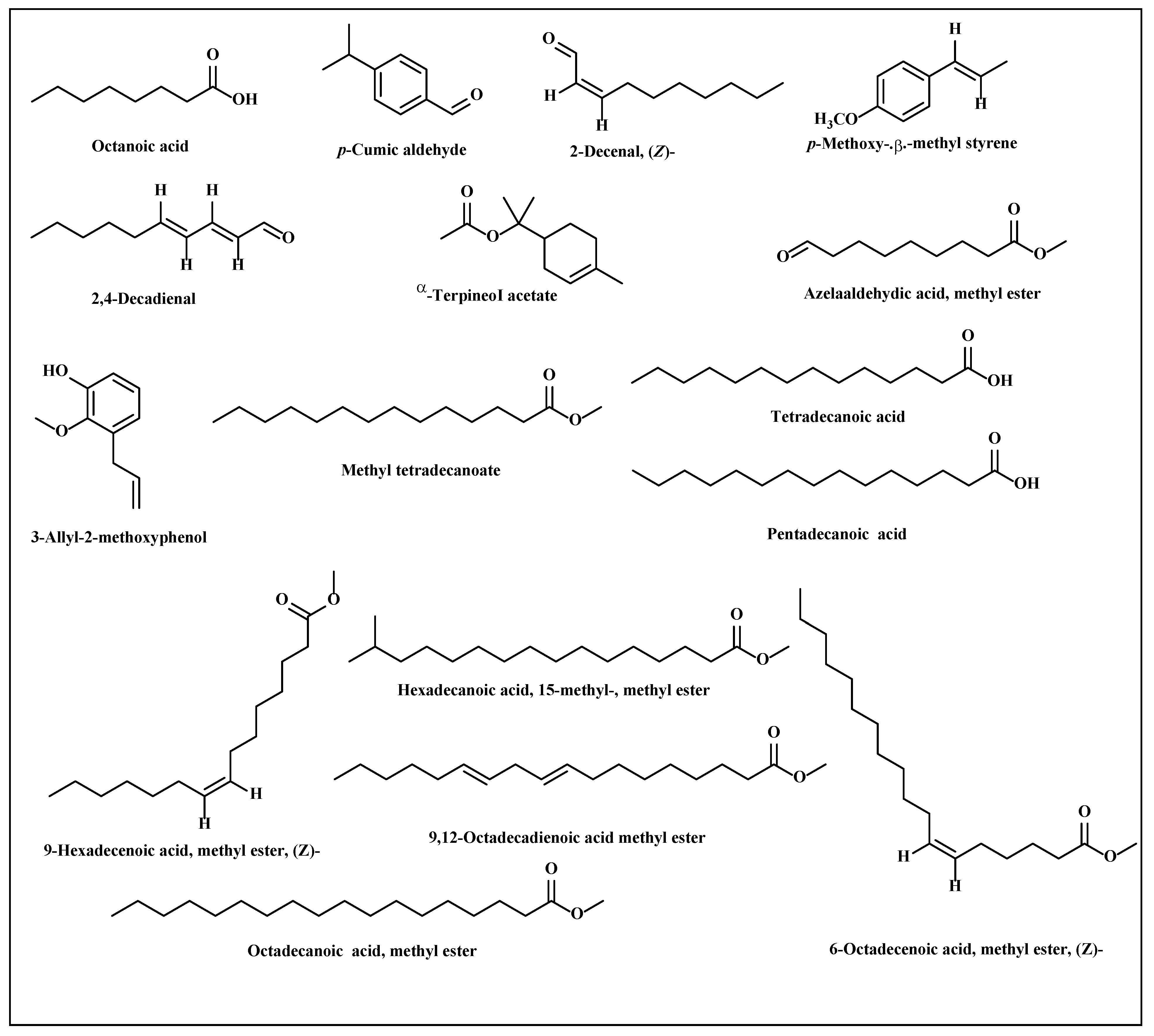
3.2.2. Metabolic Profiling of L. serriola Dried Seeds Chloroform Extract by GC/MS
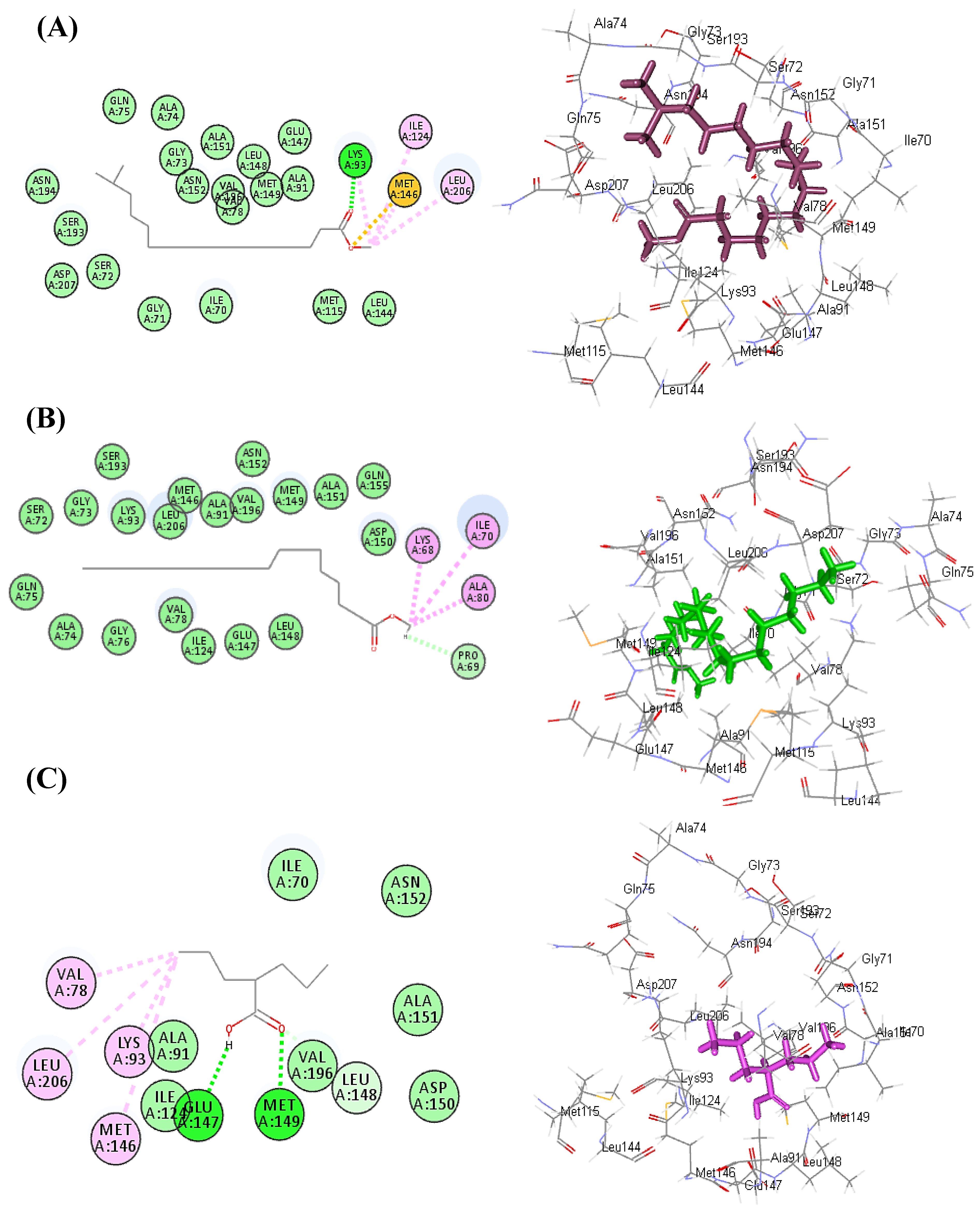
3.3. Molecular Docking Study
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Afolabi, H.A.; bin Zakariya, Z.; Shokri, A.B.A.; Hasim, M.N.B.M.; Vinayak, R.; Afolabi-Owolabi, O.T.; Elesho, R.F. The relationship between obesity and other medical comorbidities. Obes. Med. 2020, 17, 100164. [Google Scholar] [CrossRef]
- Singh, N.; Saha, L.; Kumari, P.; Singh, J.; Bhatia, A.; Banerjee, D.; Chakrabarti, A. Effect of dimethyl fumarate on neuroinflammation and apoptosis in pentylenetetrazol kindling model in rats. Brain Res. Bull. 2019, 144, 233–245. [Google Scholar] [CrossRef] [PubMed]
- Owoalade, S.A.; Lou, D.M.M.; Yadav, K.; Poudel, S. A Review on anti-epileptic activity of seaweed Ecklonia cava. J. Drug Del. Ther. 2019, 9, 425–432. [Google Scholar] [CrossRef]
- Youssef, F.S.; Menze, E.T.; Ashour, M.L. A potent lignan from Prunes alleviates inflammation and oxidative stress in lithium/pilocarpine-induced epileptic seizures in rats. Antioxidants 2020, 9, 575. [Google Scholar] [CrossRef]
- Saleh, J. MRI Marker of Decreased Parahippocampal White Matter Volume in Temporal Lobe Epilepsy. Master’s Thesis, Rush University, Chicago, IL, USA, 2019. [Google Scholar]
- Zhang, N.; Yao, L. Anxiolytic effect of essential oils and their constituents: A review. J. Agri. Food Chem 2019, 67, 13790–13808. [Google Scholar] [CrossRef]
- Thome-Souza, S.; Kuczynski, E.; Assumpção, F., Jr.; Rzezak, P.; Fuentes, D.; Fiore, L.; Valente, K.D. Which factors may play a pivotal role on determining the type of psychiatric disorder in children and adolescents with epilepsy? Epilepsy Behav. 2004, 5, 988–994. [Google Scholar] [CrossRef]
- Panayotis, N.; Freund, P.A.; Marvaldi, L.; Shalit, T.; Brandis, A.; Mehlman, T.; Tsoory, M.M.; Fainzilber, M. β-sitosterol reduces anxiety and synergizes with established anxiolytic drugs in mice. Cell Rep. Med 2021, 2, 100281. [Google Scholar] [CrossRef]
- Muazu, J.; Kaita, M. A review of traditional plants used in the treatment of epilepsy amongst the Hausa/Fulani tribes of northern Nigeria. Afr. J. Tradit. Complement. Altern. Med. 2008, 5, 387–390. [Google Scholar] [CrossRef] [Green Version]
- Auditeau, E.; Chassagne, F.; Bourdy, G.; Bounlu, M.; Jost, J.; Luna, J.; Ratsimbazafy, V.; Preux, P.-M.; Boumediene, F. Herbal medicine for epilepsy seizures in Asia, Africa and Latin America: A systematic review. J. Ethnopharmacol. 2019, 234, 119–153. [Google Scholar] [CrossRef]
- Nassiri-Asl, M.; Shariati-Rad, S.; Zamansoltani, F. Anticonvulsive effects of intracerebroventricular administration of rutin in rats. Progress Neuro-Psychopharmacol. Biol. Psychiatry 2008, 32, 989–993. [Google Scholar] [CrossRef]
- Zhu, H.L.; Wan, J.B.; Wang, Y.T.; Li, B.C.; Xiang, C.; He, J.; Li, P. Medicinal compounds with antiepileptic/anticonvulsant activities. Epilepsia 2014, 55, 3–16. [Google Scholar] [CrossRef]
- Goyal, M.; Sasmal, D. CNS depressant and anticonvulsant activities of the alcoholic extract of leaves of Ziziyphus nummularia. J. Ethnopharmacol. 2014, 151, 536–542. [Google Scholar] [CrossRef]
- Abdul-Jalil, T.Z. Lactuca serriola: Short Review of its Phytochemical and Pharmacological Profiles. Int. J. Drug Del. Technol. 2020, 10, 505–508. [Google Scholar] [CrossRef]
- Yadava, R.; Jharbade, J. New antibacterial triterpenoid saponin from Lactuca scariola. Fitoterapia 2008, 79, 245–249. [Google Scholar] [CrossRef]
- Janbaz, K.H.; Latif, M.F.; Saqib, F.; Imran, I.; Zia-Ul-Haq, M.; De Feo, V. Pharmacological effects of Lactuca serriola L. in experimental model of gastrointestinal, respiratory, and vascular ailments. Evid. Based Complement. Altern. Med. 2013, 2013, 304394. [Google Scholar] [CrossRef] [Green Version]
- Leila, J.; Ghassem, H.; Hamide, S. Introduction the medicinal species of Asteraceae family in Ilkhji and Sharafaldin regions of Esat Azarbaijan in Iran. J. Am. Sci 2011, 7, 455–458. [Google Scholar]
- Nabavi, S.F.; Nabavi, S.M.; Latifi, A.M.; Mirzaei, M.; Habtemariam, S.; Moghaddam, A.H. Mitigating role of quercetin against sodium fluoride-induced oxidative stress in the rat brain. Pharm. Biol. 2012, 50, 1380–1383. [Google Scholar] [CrossRef]
- Awan, A.F.; Akhtar, M.S.; Anjum, I.; Mushtaq, M.N.; Fatima, A.; Mannan, A.; Ali, I. Anti-oxidant and hepatoprotective effects of Lactuca serriola and its phytochemical screening by HPLC and FTIR analysis. Pak. J. Pharma. Sci. 2020, 33, 2823–2830. [Google Scholar] [CrossRef]
- Elsharkawy, E.; Alshathly, M. Anticancer activity of Lactuca steriolla growing under dry desert condition of Northern Region in Saudi Arabia. J. Nat. Sci. 2013, 3, 5–18. [Google Scholar]
- El-Esawi, M.A.; Elkelish, A.; Elansary, H.O.; Ali, H.M.; Elshikh, M.; Witczak, J.; Ahmad, M. Genetic transformation and hairy root induction enhance the antioxidant potential of Lactuca serriola L. Oxid. Med. Cell. Longev. 2017, 2017, 5604746. [Google Scholar] [CrossRef] [Green Version]
- Mohammad, A. Traditional use of kahu (Lactuca scariola L.)-a review. Glob. J. Res. Med. Plants Indigen. Med. 2013, 2, 465–474. [Google Scholar]
- Mushtaq, A.; Anwar, R.; Gohar, U.F.; Ahmad, M.; Marc, R.A.; Mureşan, C.C.; Irimie, M.; Bobescu, E. Biomolecular evaluation of Lavandula stoechas L. for nootropic activity. Plants 2021, 10, 1259. [Google Scholar] [CrossRef] [PubMed]
- Echandia, E.R.; Broitman, S.; Foscolo, M. Effect of the chronic ingestion of chlorimipramine and desipramine on the hole board response to acute stresses in male rats. Pharmacol. Biochem. Behav. 1987, 26, 207–210. [Google Scholar] [CrossRef]
- Franchini, S.; Sorbi, C.; Linciano, P.; Carnevale, G.; Tait, A.; Ronsisvalle, S.; Buccioni, M.; Del Bello, F.; Cilia, A.; Pirona, L. 1, 3-Dioxane as a scaffold for potent and selective 5-HT1AR agonist with in-vivo anxiolytic, anti-depressant and anti-nociceptive activity. Eur. J. Med. Chem. 2019, 176, 310–325. [Google Scholar] [CrossRef] [PubMed]
- Fageyinbo, M.S.; Odumeso, A.O.; Rotimi, K.; Fasipe, O.J.; Olayemi, S.O. Anticonvulsant, anxiolytic and sedative activities of hydro-ethanolic leaf extract of Mitragyna stipulosa. J. Herbs Spices Med. Plants 2019, 25, 43–54. [Google Scholar] [CrossRef]
- Sowemimo, A.A.; Adio, O.; Fageyinbo, S. Anticonvulsant activity of the methanolic extract of Justicia extensa T. Anders. J. Eethnopharmacol. 2011, 138, 697–699. [Google Scholar] [CrossRef]
- Racine, R.J. Modification of seizure activity by electrical stimulation: I. After-discharge threshold. Electroencephalogr. Clin. Neurophysiol. 1972, 32, 269–279. [Google Scholar] [CrossRef]
- Ilhan, A.; Iraz, M.; Kamisli, S.; Yigitoglu, R. Pentylenetetrazol-induced kindling seizure attenuated by Ginkgo biloba extract (EGb 761) in mice. Progress Neuro-Psychopharmacol. Biol. Psychiatry 2006, 30, 1504–1510. [Google Scholar] [CrossRef]
- Mushtaq, A.; Anwar, R.; Ahmad, M. Lavandula stoechas (L) a very potent antioxidant attenuates dementia in scopolamine induced memory deficit mice. Front. Pharmacol. 2018, 9, 1375. [Google Scholar] [CrossRef] [Green Version]
- Sinha, A.K. Colorimetric assay of catalase. Anal. Biochem. 1972, 47, 389–394. [Google Scholar] [CrossRef]
- Anwar, R.; Sultan, R.; Batool, F. Ameliorating effect of Berberis lycium root bark extracts against cisplatin-induced nephropathy in rat. Bangladesh J. Pharmacol. 2018, 13, 248–254. [Google Scholar] [CrossRef] [Green Version]
- Ohkawa, H.; Ohishi, N.; Yagi, K. Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal. Biochem. 1979, 95, 351–358. [Google Scholar] [CrossRef]
- Ijaz, M.; Fatima, M.; Anwar, R.; Uroos, M. Green synthesis of gold nanoparticles from Manilkara zapota L. extract and the evaluation of its intrinsic in vivo antiarthritic potential. RSC Adv. 2021, 11, 27092–27106. [Google Scholar] [CrossRef] [PubMed]
- Magnani, L.; Gaydou, E.M.; Hubaud, J.C. Spectrophotometric measurement of antioxidant properties of flavones and flavonols against superoxide anion. Anal. Chim. Acta 2000, 411, 209–216. [Google Scholar] [CrossRef]
- Sedlak, J.; Lindsay, R.H. Estimation of total, protein-bound, and nonprotein sulfhydryl groups in tissue with Ellman’s reagent. Anal. Biochem. 1968, 25, 192–205. [Google Scholar] [CrossRef]
- Youssef, F.S.; Ashour, M.L.; El-Beshbishy, H.A.; Ahmed Hamza, A.; Singab, A.N.B.; Wink, M. Pinoresinol-4-O-β-D-glucopyranoside: A lignan from prunes (Prunus domestica) attenuates oxidative stress, hyperglycaemia and hepatic toxicity in vitro and in vivo. J. Pharm. Pharmacol. 2020, 72, 1830–1839. [Google Scholar] [CrossRef]
- Saeed, N.; Khan, M.R.; Shabbir, M. Antioxidant activity, total phenolic and total flavonoid contents of whole plant extracts Torilis leptophylla L. BMC Complement. Altern. Med. 2012, 12, 221. [Google Scholar] [CrossRef] [Green Version]
- Malla, M.Y.; Sharma, M.; Saxena, R.; Mir, M.I.; Mir, A.H.; Bhat, S.H. Phytochemical screening and spectroscopic determination of total phenolic and flavonoid contents of Eclipta Alba Linn. J. Nat. Prod. Plant Resour. 2013, 3, 86–91. [Google Scholar]
- Youssef, F.S.; Hamoud, R.; Ashour, M.L.; Singab, A.N.; Wink, M. Volatile oils from the aerial parts of Eremophila maculata and their antimicrobial activity. Chem. Biodivers. 2014, 11, 831–841. [Google Scholar] [CrossRef]
- Mamadalieva, N.Z.; Youssef, F.S.; Ashour, M.L.; Sasmakov, S.A.; Tiezzi, A.; Azimova, S.S. Chemical composition, antimicrobial and antioxidant activities of the essential oils of three Uzbek Lamiaceae species. Nat. Prod. Res. 2019, 33, 2394–2397. [Google Scholar] [CrossRef]
- Altyar, A.E.; Ashour, M.L.; Youssef, F.S. Premna odorata: Seasonal metabolic variation in the essential oil composition of its leaf and verification of its anti-ageing potential via in vitro assays and molecular modelling. Biomolecules 2020, 10, 879. [Google Scholar] [CrossRef]
- Storici, P.; De Biase, D.; Bossa, F.; Bruno, S.; Mozzarelli, A.; Peneff, C.; Silverman, R.B.; Schirmer, T. Structures of γ-aminobutyric acid (GABA) aminotransferase, a pyridoxal 5′-phosphate, and [2Fe-2S] cluster-containing enzyme, complexed with γ-ethynyl-GABA and with the antiepilepsy drug vigabatrin. J. Biol. Chem. 2004, 279, 363–373. [Google Scholar] [CrossRef] [Green Version]
- Thabet, A.A.; Youssef, F.S.; El-Shazly, M.; El-Beshbishy, H.A.; Singab, A.N.B. Validation of the antihyperglycaemic and hepatoprotective activity of the flavonoid rich fraction of Brachychiton rupestris using in vivo experimental models and molecular modelling. Food Chem. Toxicol. 2018, 114, 302–310. [Google Scholar] [CrossRef]
- Talaat, A.N.; Ebada, S.S.; Labib, R.M.; Esmat, A.; Youssef, F.S.; Singab, A.N.B. Verification of the anti-inflammatory activity of the polyphenolic-rich fraction of Araucaria bidwillii Hook. using phytohaemagglutinin-stimulated human peripheral blood mononuclear cells and virtual screening. J. Ethnopharmacol. 2018, 226, 44–47. [Google Scholar] [CrossRef]
- Labib, R.; Youssef, F.; Ashour, M.; Abdel-Daim, M.; Ross, S. Chemical composition of Pinus roxburghii bark volatile oil and validation of its anti-inflammatory activity using molecular modelling and bleomycin-induced inflammation in albino mice. Molecules 2017, 22, 1384. [Google Scholar] [CrossRef] [Green Version]
- Silverman, R.B. Design and mechanism of GABA aminotransferase inactivators. Treatments for epilepsies and addictions. Chem. Rev. 2018, 118, 4037–4070. [Google Scholar] [CrossRef]
- Stojanović, N.M.; Mladenović, M.Z.; Maslovarić, A.; Stojiljković, N.I.; Randjelović, P.J.; Radulović, N.S. Lemon balm (Melissa officinalis L.) essential oil and citronellal modulate anxiety-related symptoms–In vitro and in vivo studies. J. Ethnopharmacol. 2022, 284, 114788. [Google Scholar] [CrossRef]
- Löscher, W. Animal models of seizures and epilepsy: Past, present, and future role for the discovery of antiseizure drugs. Neurochem. Res. 2017, 42, 1873–1888. [Google Scholar] [CrossRef]
- Dhir, A. Pentylenetetrazol (PTZ) kindling model of epilepsy. Curr. Prot. Neurosci. 2012, 58, 9.37.1–9.37.12. [Google Scholar] [CrossRef]
- Tourov, A.; Ferri, R.; Del Gracco, S.; Elia, M.; Musumeci, S.A.; Stefanini, M.C. Spike morphology in PTZ-induced generalized and cobalt-induced partial experimental epilepsy. Funct. Neurol. 1996, 11, 237–245. [Google Scholar]
- Phillipson, J.D.; Anderson, L.A. Ethnopharmacology and western medicine. J. Ethnopharmacol. 1989, 25, 61–72. [Google Scholar] [CrossRef]
- Takeda, H.; Tsuji, M.; Matsumiya, T. Changes in head-dipping behavior in the hole-board test reflect the anxiogenic and/or anxiolytic state in mice. Eur. J. Pharmacol. 1998, 350, 21–29. [Google Scholar] [CrossRef]
- Sofidiya, M.O.; Alokun, A.M.; Fageyinbo, M.S.; Akindele, A.J. Central nervous system depressant activity of ethanol extract of Motandra guineensis (Thonn) AD. aerial parts in mice. Phytomed. Plus 2022, 2, 100186. [Google Scholar] [CrossRef]
- Foyet, H.S.; Tsala, D.E.; Bouba, A.A.; Hritcu, L. Anxiolytic and antidepressant-like effects of the aqueous extract of Alafia multiflora stem barks in rodents. Adv. Pharmacol. Sci. 2012, 2012, 912041. [Google Scholar] [CrossRef] [Green Version]
- Young, R.; Johnson, D.N. A fully automated light/dark apparatus useful for comparing anxiolytic agents. Pharmacol. Biochem. Behav. 1991, 40, 739–743. [Google Scholar] [CrossRef]
- Macdonald, R.L.; Kelly, K.M. Antiepileptic drug mechanisms of action. Epilepsia 1995, 36, S2–S12. [Google Scholar] [CrossRef] [Green Version]
- Emam, G.M.; Heydari, H.G. Sedative-hypnotic activity of extracts and essential oil of coriander seeds. Iran. J. Med. Sci. 2006, 31, 22–27. [Google Scholar]
- N’gouemo, P.; Nguemby-Bina, C.; Baldy-Moulinier, M. Some neuropharmacological effects of an ethanolic extract of Maprounea africana in rodents. J. Eethnopharmacol. 1994, 43, 161–166. [Google Scholar] [CrossRef]
- Goodman, L.S.; Gilman, A. Goodman and Gilman’s the Pharmacological Basis of Therapeutics, 13th ed.; McGraw-Hill Education: New York, NY, USA, 2011. [Google Scholar]
- Beyenburg, S.; Mitchell, A.J.; Schmidt, D.; Elger, C.E.; Reuber, M. Anxiety in patients with epilepsy: Systematic review and suggestions for clinical management. Epilepsy Behav. 2005, 7, 161–171. [Google Scholar] [CrossRef]
- Murray, J.B. Effects of valium and librium on human psychomotor and cognitive functions. Genet. Psychol. Monogr. 1984, 109, 167–197. [Google Scholar]
- Rang, H.P.; Dale, M.M.; Ritter, J.M.; Moore, P. Pharmacology; Churchill Livingstone: London, UK, 2003. [Google Scholar]
- Nicoll, R.A. Introduction to the pharmacology of the central nervous system. Basic Clin. Pharmacol. 2001. [Google Scholar]
- Cardenas-Rodriguez, N.; Huerta-Gertrudis, B.; Rivera-Espinosa, L.; Montesinos-Correa, H.; Bandala, C.; Carmona-Aparicio, L.; Coballase-Urrutia, E. Role of oxidative stress in refractory epilepsy: Evidence in patients and experimental models. Int. J. Mol. Sci. 2013, 14, 1455–1476. [Google Scholar] [CrossRef] [Green Version]
- Jain, S.; Bharal, N.; Khurana, S.; Mediratta, P.K.; Sharma, K.K. Anticonvulsant and antioxidant actions of trimetazidine in pentylenetetrazole-induced kindling model in mice. Naunyn-Schmiedebergs Arch. Pharmacol. 2011, 383, 385–392. [Google Scholar] [CrossRef]
- Bikjdaouene, L.; Escames, G.; Leon, J.; Ferrer, J.M.; Khaldy, H.; Vives, F.; Acuña-Castroviejo, D. Changes in brain amino acids and nitric oxide after melatonin administration in rats with pentylenetetrazole-induced seizures. J. Pineal Res. 2003, 35, 54–60. [Google Scholar] [CrossRef] [Green Version]
- Fernández, S.P.; Wasowski, C.; Loscalzo, L.M.; Granger, R.E.; Johnston, G.A.; Paladini, A.C.; Marder, M. Central nervous system depressant action of flavonoid glycosides. Eur. J. Pharmacol. 2006, 539, 168–176. [Google Scholar] [CrossRef]
- Gangwal, A. Neuropharmacological effects of triterpenoids. Phytopharmacology 2013, 4, 354–372. [Google Scholar]
- Zeng, L.; Zhang, H.; Xu, C.; Bian, Y.; Xu, X.; Xie, Q.; Zhang, R. Neuroprotective effects of flavonoids extracted from licorice on kainate-induced seizure in mice through their antioxidant properties. J. Zhejiang Univ. Sci. B 2013, 14, 1004–1012. [Google Scholar] [CrossRef] [Green Version]
- Lasoń, W.; Leśkiewicz, M. Effect of plant polyphenols on seizures–animal studies. J. Epileptol. 2013, 21, 79–87. [Google Scholar] [CrossRef]
- Diniz, T.C.; Silva, J.C.; Lima-Saraiva, S.R.G.d.; Ribeiro, F.P.R.d.A.; Pacheco, A.G.M.; de Freitas, R.M.; Quintans-Júnior, L.J.; Quintans, J.d.S.S.; Mendes, R.L.; Almeida, J.R.G.d.S. The role of flavonoids on oxidative stress in epilepsy. Oxid. Med. Cell. Longev. 2015, 2015, 171756. [Google Scholar] [CrossRef]
- Choudhary, N.; Bijjem, K.R.V.; Kalia, A.N. Antiepileptic potential of flavonoids fraction from the leaves of Anisomeles malabarica. J. Ethnopharmacol. 2011, 135, 238–242. [Google Scholar] [CrossRef]
- Costa Marques, T.H.; Santos De Melo, C.H.; Fonseca De Carvalho, R.B.; Costa, L.M.; De Souza, A.A.; David, J.M.; De Lima David, J.P.; De Freitas, R.M. Phytochemical profile and qualification of biological activity of an isolated fraction of Bellis perennis. Biol. Res. 2013, 46, 231–238. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Extract | Control | Standard | 300 mg/kg | 400 mg/kg | 500 mg/kg |
|---|---|---|---|---|---|
| n-Hexane | 10.88 ± 0.04 | 10.06 ± 0.03 * | 10.08 ± 0.04 * | 9.98 ± 0.03 * | 9.51 ± 0.06 * |
| Chloroform | 11.35 ± 0.04 | 10.98 ± 0.04 * | 9.76 ± 0.04 * | 9.06 ± 0.03 * | 8.33 ± 0.04 * |
| Methanol | 10.03 ± 0.03 | 9.81 ± 0.03 * | 8.95 ± 0.04 * | 7.6 ± 0.05 * | 5.41 ± 0.04 * |
| Aqueous | 10.11 ± 0.03 | 9.96 ± 0.03 * | 8.1 ± 0.02 * | 7.55 ± 0.05 * | 7 ± 0.03 * |
| Extract | Control | Standard | 300 mg/kg | 400 mg/kg | 500 mg/kg |
|---|---|---|---|---|---|
| n-Hexane | 189.8 ± 0.60 | 202.1 ± 0.83 * | 241.6 ± 0.55 * | 263.3 ± 0.88 * | 291.1 ± 0.70 * |
| Chloroform | 204.83 ± 0.47 | 214.33 ± 0.61 * | 247 ± 0.57 * | 273 ± 0.81 * | 302.3 ± 0.71 * |
| Methanol | 211.3 ± 0.61 | 190.66 ± 0.49 * | 262.6 ± 0.71 * | 304.1 ± 0.70 * | 326.5 ± 1.08 * |
| Aqueous | 197.5 ± 0.42 | 203.16 ± 0.47 * | 260.16 ± 0.30 * | 281.5 ± 0.67 * | 292 ± 0.57 * |
| Treatment Groups | Seizure Latency | Seizure Duration | Mortality (Yes/No) |
|---|---|---|---|
| PCT | 679 ± 3.22 | 213.3 ± 2.44 | No |
| PCT + Phenobarbitone | - | - | No |
| PCT + n-hexane extract (100 mg) | 1514 ± 2.95 * | 110.8 ± 1.42 * | No |
| PCT + n-hexane extract (200 mg) | 2011.1 ± 1.74 * | 20.5 ± 1.17 * | No |
| PCT + n-hexane extract (300 mg) | - | - | No |
| PCT + chloroform extract (100 mg) | 2253.8 ± 8.46 * | 16.7 ± 2.15 * | No |
| PCT + chloroform extract (200 mg) | - | - | No |
| PCT + chloroform extract (300 mg) | - | - | No |
| PCT + methanol extract (100 mg) | 821.5 ± 4.12 * | 218.3 ± 2.23 | No |
| PCT + methanol extract (200 mg) | 1036.1 ± 6.73 * | 184.8 ± 0.30 * | No |
| PCT + methanol extract (300 mg) | 1194.5 ± 3.94 * | 120.3 ± 3.06 * | No |
| PCT + aqueous extract (100 mg) | 820.6 ± 4.06 * | 214.6 ± 2.15 | No |
| PCT + aqueous extract (200 mg) | 1031.1 ± 5.26 * | 168.1 ± 8.89 * | No |
| PCT + aqueous extract (300 mg) | 1329.3.3 ± 8.22 * | 112.6 ± 3.26 * | No |
| Treatment Groups | Seizure Latency | Seizure Duration | Mortality (Yes/No) |
|---|---|---|---|
| STR | 124.0 ± 0.93 | 48.33 ± 1.2 | Yes |
| STR + Diazepam | 214.5 ± 0.84 * | 20.33 ± 1.87 * | Yes |
| STR + n-hexane extract (100 mg) | 142.6 ± 0.88 * | 31.00 ± 0.57 * | Yes |
| STR + n-hexane extract (200 mg) | 178.3 ± 1.14 * | 26.00 ± 0.44 * | Yes |
| STR + n-hexane extract (300 mg) | 242.6 ± 0.84 * | 20.33 ± 0.66 * | Yes |
| STR + chloroform extract (100 mg) | 194.1 ± 2.73 * | 20.50 ± 0.42 * | Yes |
| STR + chloroform extract (200 mg) | 242.1 ± 0.94 * | 17.83± 0.4 * | Yes |
| STR + chloroform extract (300 mg) | 254.5 ± 1.05 * | 16.10 ± 0.6 * | Yes |
| STR + methanol extract (100 mg) | 147.1 ± 1.47 * | 31.30 ± 0.66 * | Yes |
| STR + methanol extract (200 mg) | 170.5 ± 0.76 * | 28.83 ± 0.30 * | Yes |
| STR + methanol extract (300 mg) | 190.5 ± 0.76 * | 24.66 ± 0.49 * | Yes |
| STR + aqueous extract (100 mg) | 153 ± 1.03 * | 29.5 ± 0.67 * | Yes |
| STR + aqueous extract (200 mg) | 162.6 ± 1.05 * | 26.50 ± 0.67 * | Yes |
| STR + aqueous extract (300 mg) | 180.5 ± 0.76 * | 19.83 ± 0.60 * | Yes |
| Treatment | Seizure Latency | Seizure Intensity | Mortality |
|---|---|---|---|
| PTZ only | 240± 0.57 | 5.75 ± 0.28 | 5/6 |
| Standard (VA) | 680 ± 0.88 * | 3.25 ± 0.28 * | 1/6 * |
| n-Hexane | 580 ± 1.76 * | 3.66 ± 0.33 * | 1/6 * |
| Chloroform | 710 ± 1.20 * | 3.33 ± 0.33 * | 1/6 * |
| Methanol | 380 ± 0.33 * | 3.75 ± 0.28 * | 3/6 * |
| Aqueous | 520 ± 1.20 * | 4.33 ± 0.33 * | 3/6 * |
| Treatment Group | MDA (µM/mg Tissue) | CAT (µM/mg Tissue) | SOD (U/mL) | Total GSH (mM/g) |
|---|---|---|---|---|
| Normal | 8.51 ± 0.28 | 40.8 ± 0.44 | 61.76 ± 0.50 | 1.44 ± 0.00 |
| PTZ | 27.38 ± 0.08 | 8.50 ± 0.57 | 20.80 ± 0.19 | 0.37 ± 0.01 |
| Standard | 18.75 ± 0.23 * | 16.6 ± 0.60 * | 50.88 ± 1.56 * | 1.01 ± 0.01 * |
| n-Hexane | 17.04 ± 0.39 * | 16.6 ± 0.88 * | 48.89 ± 2.51 * | 0.87 ± 0.01 * |
| Chloroform | 17.39 ± 0.06 * | 15.6 ± 0.92 * | 47.01 ± 0.97 * | 0.92 ± 0.00 * |
| Methanol | 25.54 ± 0.05 * | 13.16 ± 0.44 * | 38.31 ± 0.39 * | 0.62 ± 0.00 * |
| Aqueous | 25.52 ± 0.18 * | 11.66 ± 0.44 * | 31.38 ± 0.62 * | 0.54 ± 0.00 * |
| No. | Name | Retention Time | Retention Index | Concentration (%) |
|---|---|---|---|---|
| 1 | Octanoic acid | 7.47 | 1179 | 0.63 |
| 2 | p-Cumic aldehyde | 9.20 | 1239 | 0.35 |
| 3 | 2-Decenal, (Z)- | 9.66 | 1250 | l. 10 |
| 4 | p-Methoxy-.beta.-methyl styrene | 10.28 | 1303 | 0.38 |
| 5 | 2,4-Decadienal | 10.44 | 1309 | 4.93 |
| 6 | α-Terpineol acetate | I 1.85 | 1354 | 0.73 |
| 7 | 3-Allyl-2-methoxyphenol | 12.01 | 1392 | 1.38 |
| 8 | Azelaaldehydic acid, methyl ester | 13.81 | 1436 | 0.30 |
| 9 | Methyl tetradecanoate | 20.57 | 1706 | 0.84 |
| 10 | Tetradecanoic acid | 21.37 | 1770 | 1.34 |
| 11 | Pentadecanoic acid | 23.45 | 1857 | 0.45 |
| 12 | 9-Hexadecenoic acid, methyl ester, (Z)- | 24.32 | 1879 | 0.37 |
| 13 | Hexadecanoic acid, 15-methyl-, methyl ester | 24.78 | 1996 | 23.25 |
| 14 | 9,12-octadecadienoic acid methyl ester | 28.10 | 2094 | 49.03 |
| 15 | 6-Octadecenoic acid, methyl ester, (Z)- | 28.30 | 2104 | 1.18 |
| 16 | Octadecanoic acid, methyl ester | 28.64 | 2112 | 12.82 |
| Total identified compounds | 97.98% |
| Compound | GABA-AT (1OHV) | Number of Formed Hydrogen Bonds and C-H Bonds | Number of Formed Alkyl and π-Alkyl Bonds |
|---|---|---|---|
| 2,4-Decadienal | −20.37 | 3; Gln75, Gly76 | - |
| 2-Decenal, (Z)- | −21.41 | 2; Gly76, Lys93 | - |
| 3-Allyl-2-methoxyphenol | −14.92 | 2; Met149 | 4; Leu206, Val196, Val78, Ala91 |
| 6-Octadecenoic acid, methyl ester, (Z)- | −29.92 | 3; Lys93, Glu111, Asp207 | - |
| 9,12-octadecadienoic acid methyl ester | −15.46 | - | 3; Lys93, Leu206 |
| 9-Hexadecenoic acid, methyl ester, (Z)- | −26.32 | 3; Met448, Asp150, Ala151 | 1; Ile70 |
| α-TerpineoI acetate | −0.69 | 1; Ala151 | 7; Val78, Val196, Leu206, Ala91, Ile70 |
| Azelaaldehydic acid, methyl ester | −36.09 | 2; Asn152, Lys193 | - |
| Hexadecanoic acid, 15-methyl-, methyl ester | −51.40 | 1; Lys93 | 2; Ile124, Leu206 |
| Methyl tetradecanoate | −43.56 | 2; Asn152, Lys93 | 1; Leu206 |
| Octadecanoic acid, methyl ester | −44.74 | 1; Pro69 | 3; Lys58, Ile70, Ala80 |
| Octanoic acid | −33.64 | 2; Asn152, Gln155 | - |
| p-Cumic aldehyde | −24.44 | - | 5; Val78, Val196, Ala91, Ile70 |
| p-Methoxy-.beta.-methyl styrene | −11.04 | 3; Gln155, Ile70, Ala151 | 2; Val196, Ile70 |
| Pentadecanoic acid | −44.01 | 2; Lys93, Asn194 | - |
| Tetradecanoic acid | −42.94 | 2; Lys93, Asn194 | - |
| Valproic acid | −30.19 | 3; Glu147, Met149, Leu148 | 4; Val78, Leu206, Lys93, Met146 |
| Co-crystalized ligand | −7.74 | 2; Glu147, Met149 | 11; Val78, Val196, Ala91, Ile70, Leu206 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ullah, M.I.; Anwar, R.; Kamran, S.; Gul, B.; Elhady, S.S.; Youssef, F.S. Evaluation of the Anxiolytic and Anti-Epileptogenic Potential of Lactuca Serriola Seed Using Pentylenetetrazol-Induced Kindling in Mice and Metabolic Profiling of Its Bioactive Extract. Antioxidants 2022, 11, 2232. https://doi.org/10.3390/antiox11112232
Ullah MI, Anwar R, Kamran S, Gul B, Elhady SS, Youssef FS. Evaluation of the Anxiolytic and Anti-Epileptogenic Potential of Lactuca Serriola Seed Using Pentylenetetrazol-Induced Kindling in Mice and Metabolic Profiling of Its Bioactive Extract. Antioxidants. 2022; 11(11):2232. https://doi.org/10.3390/antiox11112232
Chicago/Turabian StyleUllah, Muhammad Ihsan, Rukhsana Anwar, Shahzad Kamran, Bazgha Gul, Sameh S. Elhady, and Fadia S. Youssef. 2022. "Evaluation of the Anxiolytic and Anti-Epileptogenic Potential of Lactuca Serriola Seed Using Pentylenetetrazol-Induced Kindling in Mice and Metabolic Profiling of Its Bioactive Extract" Antioxidants 11, no. 11: 2232. https://doi.org/10.3390/antiox11112232