
Metabolic Profiling and Investigation of the Modulatory Effect of Fagonia cretica L. Aerial Parts on Hepatic CYP3A4 and UGT2B7 Enzymes in Streptozotocin—Induced Diabetic Model
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Preparation of Plant Extracts
2.3. Chemicals and Drugs
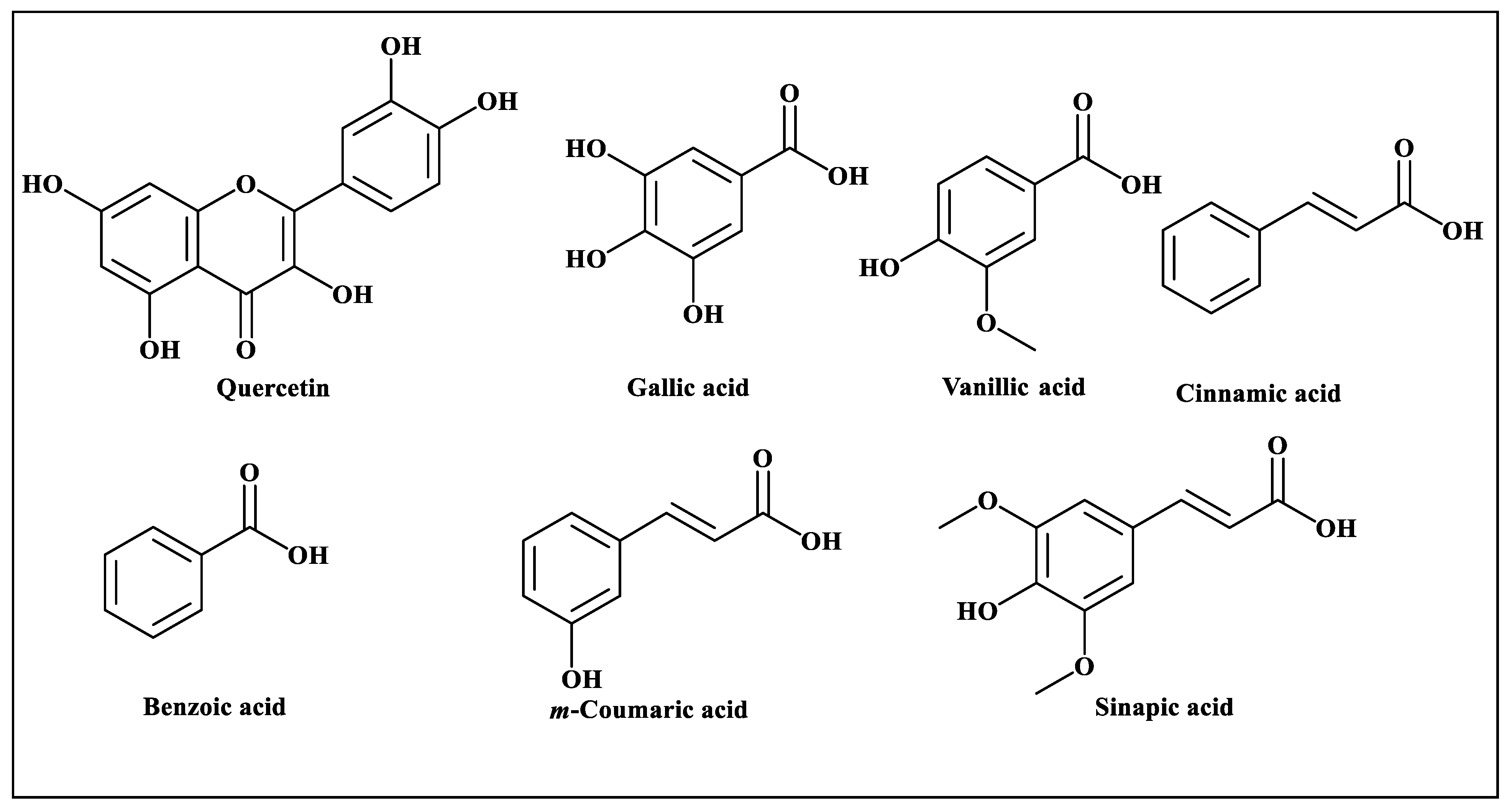
2.4. Phytochemical Analysis of Fagonia cretica Using High-Performance Liquid Chromatography (HPLC)
2.5. In Vivo Investigation of the Modulatory Effect of Fagonia cretica Extracts
2.5.1. Experimental Animals
2.5.2. Induction and Maintenance of Diabetes
2.5.3. Experimental Design
2.5.4. Homogenization of Liver
2.5.5. Isolation of mRNA and Real-Time PCR Quantitative Assay
2.6. Sample Prepration for Detection and Quantitaion of CYP3A4 and UGT Substrates
2.6.1. Sample Preparation for Detection of CYP3A4 and UGT Substrates by ELISA
2.6.2. Liquid–Liquid Extraction for CYP3A4 Substrate Quantitation by GCMS
2.6.3. Solid–Liquid Extraction for UGT2B7 Substrate Quantitation by GCMS
2.7. Preparation of Working Solutions
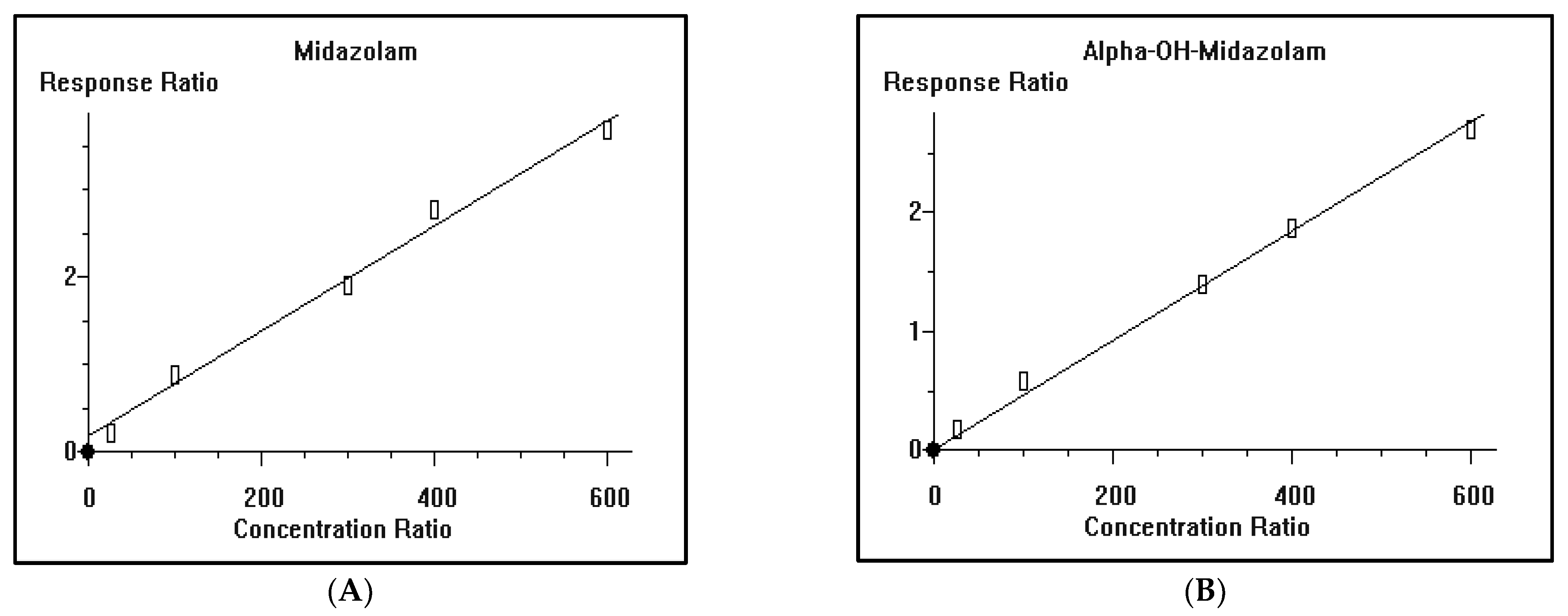
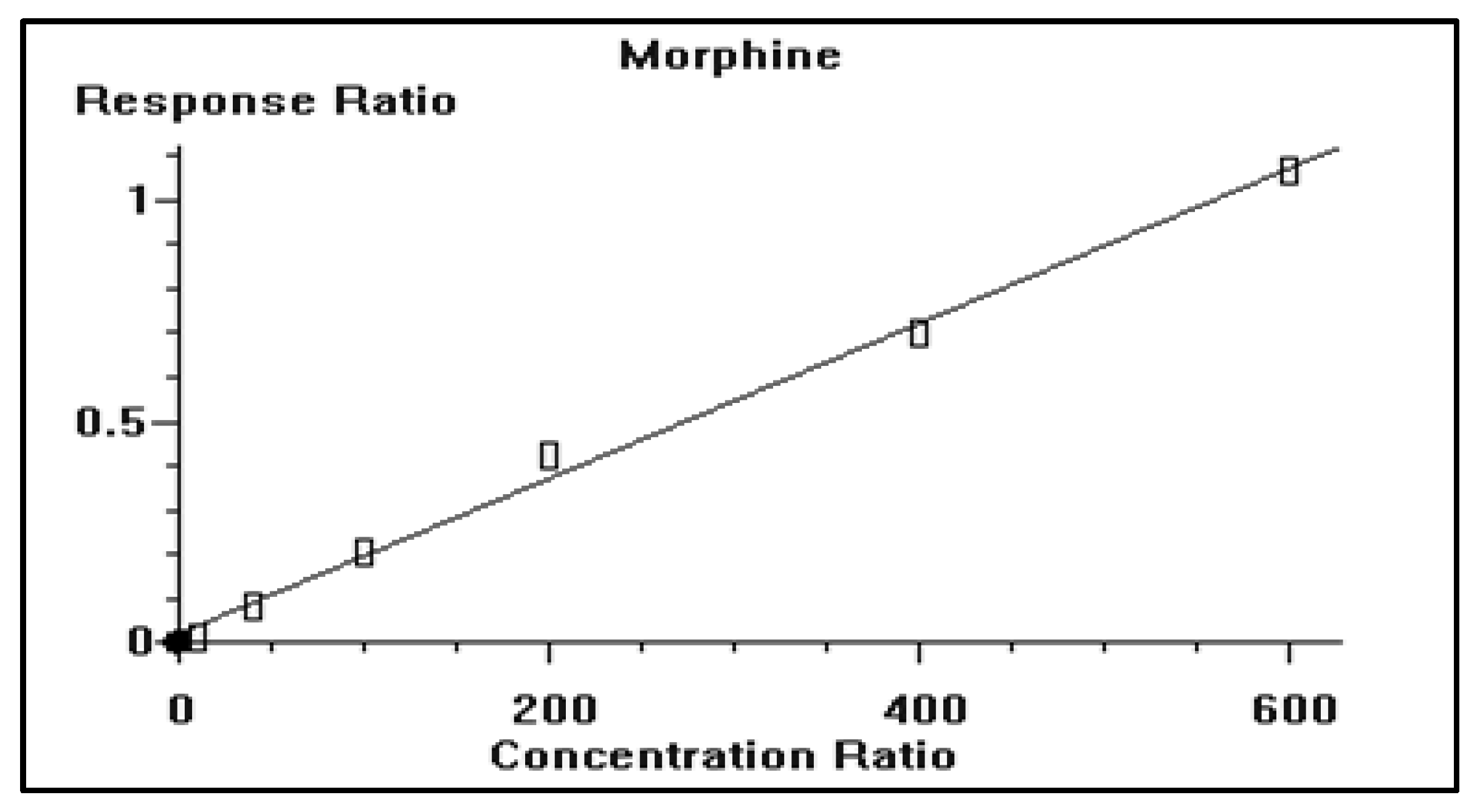
2.8. Method Validation
2.8.1. Specificity
2.8.2. Construction of Calibration Curve and Linearity
2.8.3. Sensitivity
2.8.4. Accuracy and Precision
2.9. Analysis Method for the Detection of CYP3A4 and UGT2B7 Substrates by ELISA
2.10. Analysis Method for the Quantitation of Alpha-Hydroxyl Midazolam and Morphine by GCMS
2.11. Molecular Docking Study
2.12. Statistical Analysis
3. Results
3.1. Phytochemical Analysis of Fagonia cretica Using High-Performance Liquid Chromatography (HPLC)
3.2. GC/MS Method Validation
3.3. Effect of F. cretica Extracts on the Activity and mRNA Expression of CYP3A4 and UGT2B7
3.3.1. Effect of F. cretica Extracts on the Activity of CYP3A and UGT2B7 by ELISA
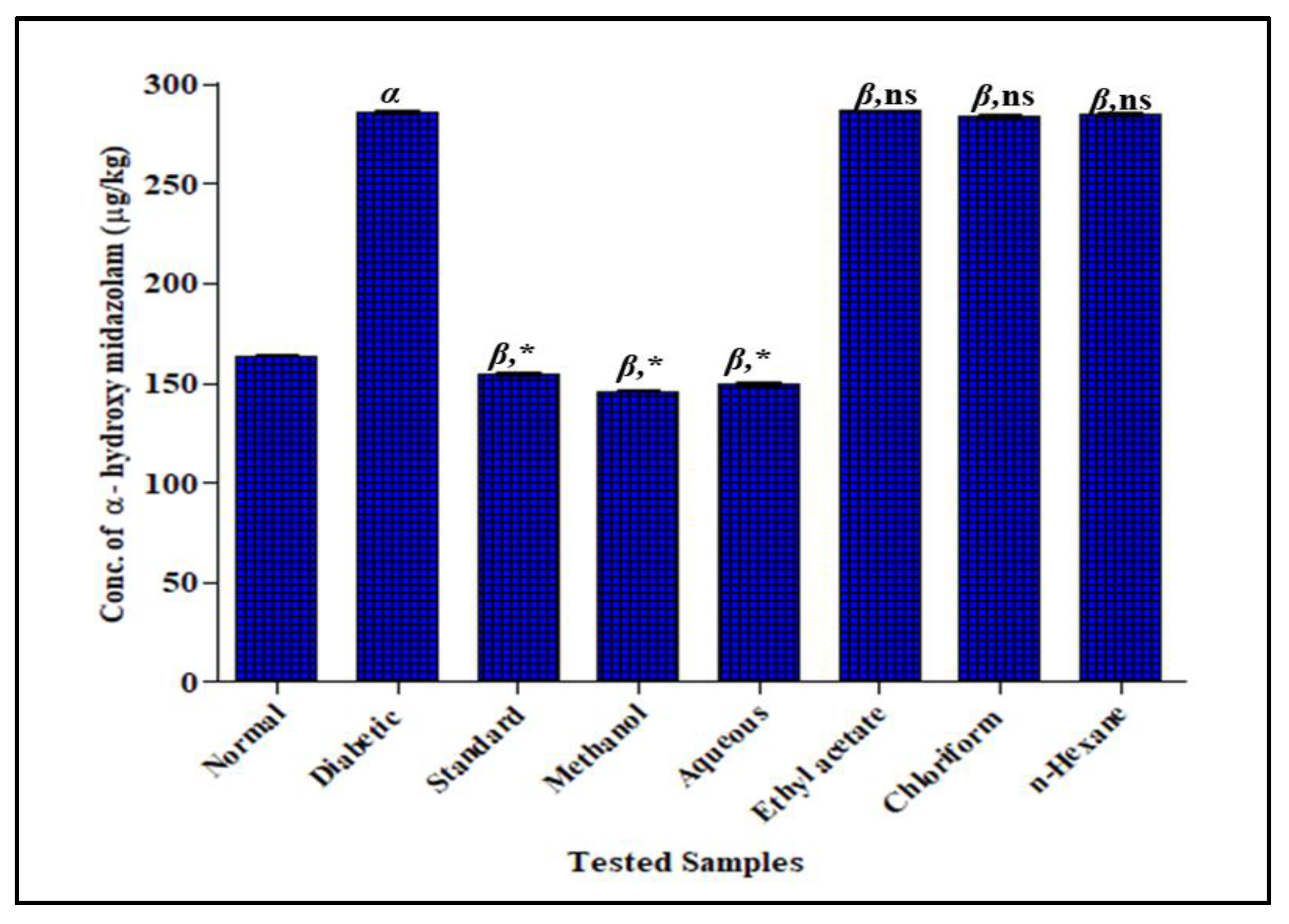
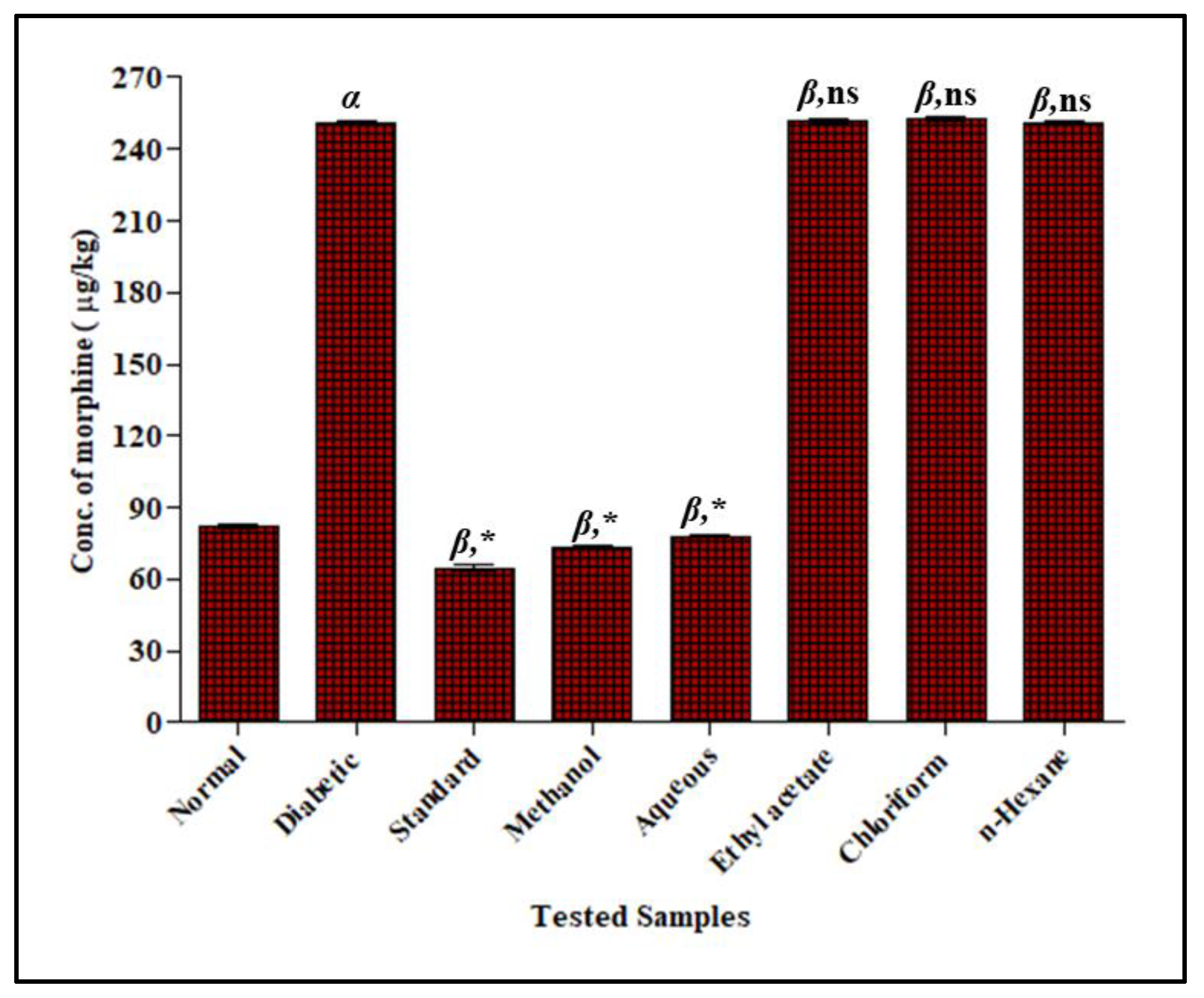
3.3.2. Effect of F. cretica Extracts on the Activity of CYP3A4 by GCMS
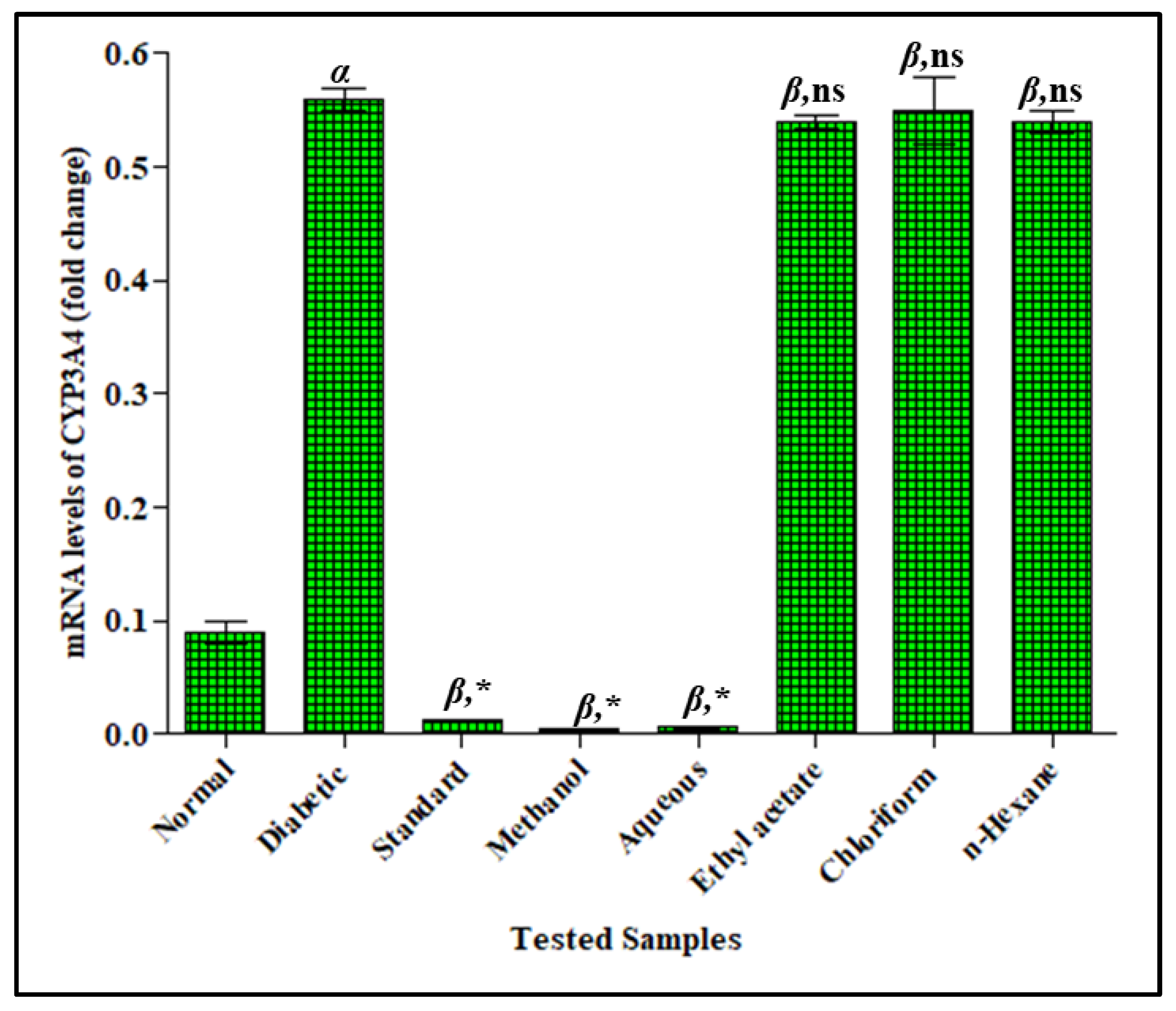
3.3.3. Effect of F. cretica Extracts on Fold-Change mRNA Expression of CYP3A4 by PCR
3.3.4. Effect of F. cretica Extracts on the Activity of UGT2B7 by GCMS
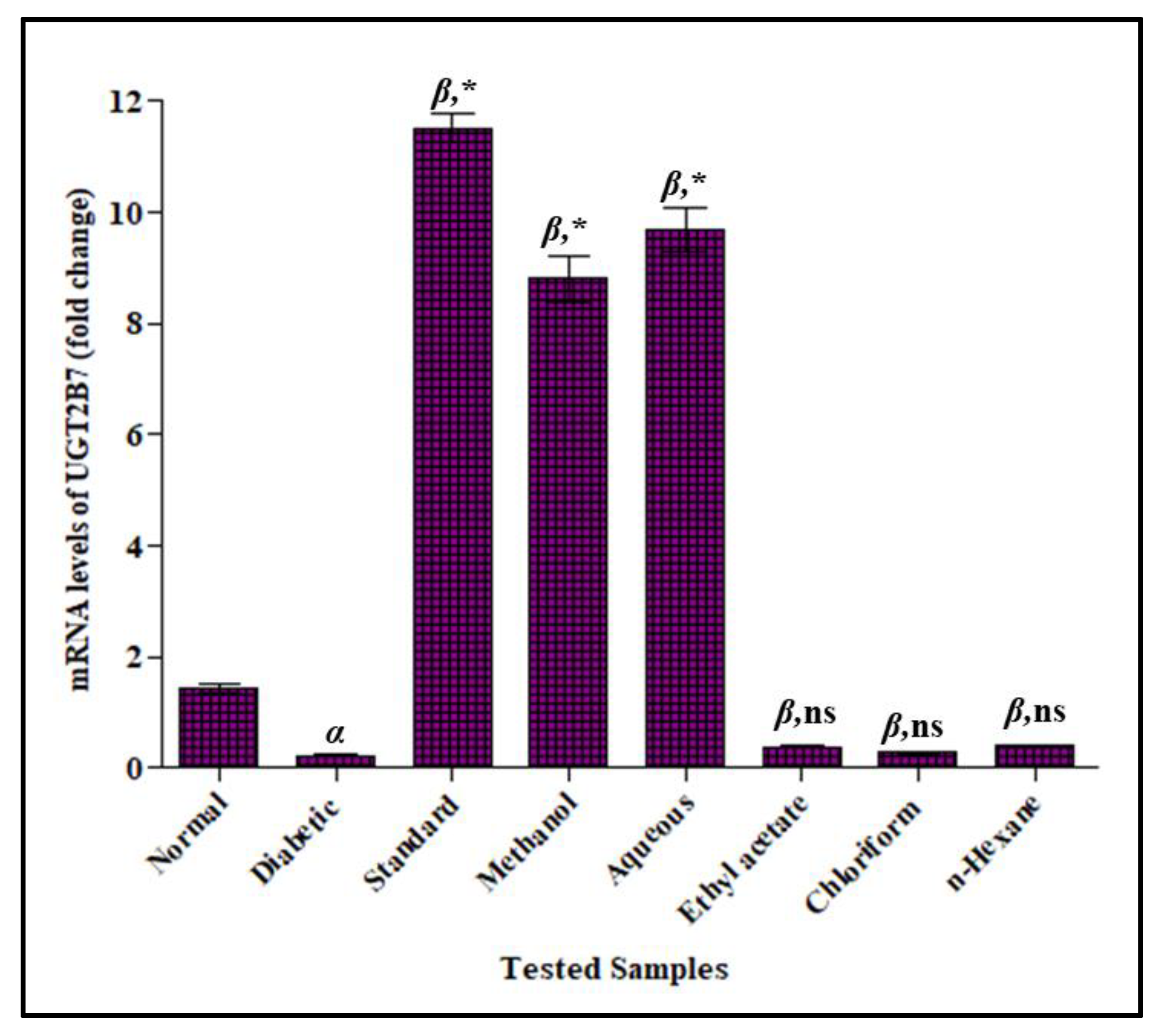
3.3.5. Effect of F. cretica Extracts on Fold-Change mRNA Expression of UGT2B7 by PCR
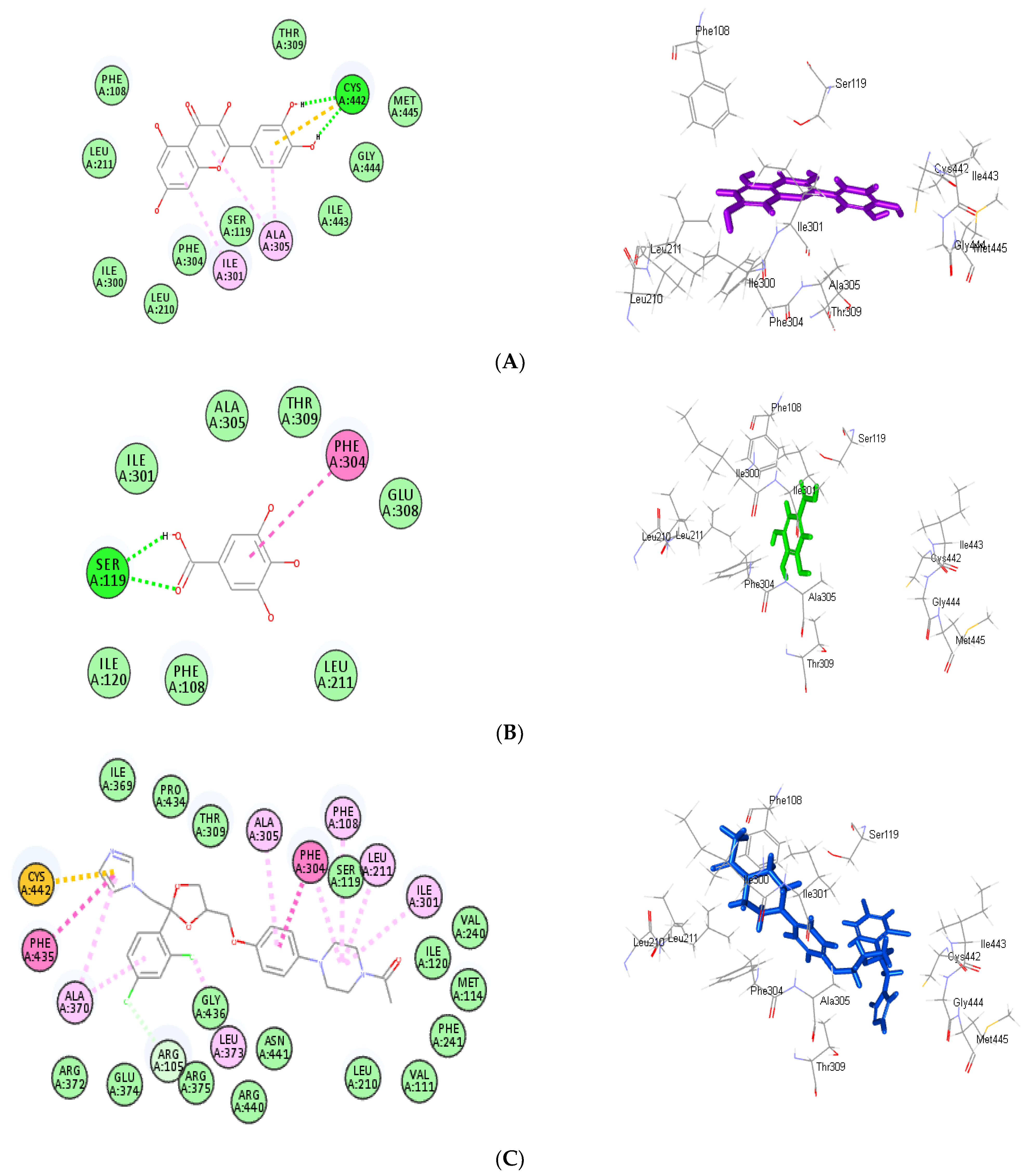
3.4. Molecular Docking Study
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Anwar, R.; Hussin, A.H.; Ismail, S.; Mansor, S.M. In vitro effect of mitragynine (a major alkaloid of Mitragyna speciosa korth) on aminopyrine metabolism in rat hepatocytes. Int. J. Pharm. Sci. Res. 2012, 3, 2238. [Google Scholar]
- Liska, D.A.J. The detoxification enzyme systems. Altern. Med. Rev. 1998, 3, 187–198. [Google Scholar] [PubMed]
- McCarver, D.G.; Hines, R.N.; Therapeutics, E. The ontogeny of human drug-metabolizing enzymes: Phase II conjugation enzymes and regulatory mechanisms. J. Pharmacol. Exp. Ther. 2002, 300, 361–366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, H.; Sun, S.; Cheng, X.; Yan, T.; Zheng, X.; Li, F.; Qi, Q.; Wang, G.; Hao, H. Dysregulations of intestinal and colonic UDP-glucuronosyltransferases in rats with type 2 diabetes. Drug Metab. Pharmacokinet. 2013, 28, 427–434. [Google Scholar] [CrossRef] [PubMed]
- Janibekov, A.A.; Youssef, F.S.; Ashour, M.L.; Mamadalieva, N.Z. New flavonoid glycosides from two Astragalus species (Fabaceae) and validation of their antihyperglycaemic activity using molecular modelling and in vitro studies. Ind. Crops Prod. 2018, 118, 142–148. [Google Scholar] [CrossRef]
- Rösen, P.; Nawroth, P.; King, G.; Möller, W.; Tritschler, H.J.; Packer, L. The role of oxidative stress in the onset and progression of diabetes and its complications: Asummary of a Congress Series sponsored byUNESCO-MCBN, the American Diabetes Association and the German Diabetes Society. Diabetes Metab. Res. Rev. 2001, 17, 189–212. [Google Scholar] [CrossRef]
- Sotaniemi, E.A.; Pelkonen, O.; Arranto, A.J.; Tapanainen, P.; Rautio, A.; Pasanen, M. Diabetes and elimination of antipyrine in man: An analysis of 298 patients classified by type of diabetes, age, sex, duration of disease and liver involvement. Pharmacol. Toxicol 2002, 90, 155–160. [Google Scholar] [CrossRef]
- Anwar, R.; Hussin, H.; Ismail, S.; Mansor, M. In vitro effect of mitragynine on activity of drug metabolizing enzymes, n-demethylase and glutathione s-transferase in streptozotocin-induced diabetic rats. Pharmacologyonline 2012, 1, 68–75. [Google Scholar]
- Ashour, M.L.; Youssef, F.S.; Gad, H.A.; Wink, M. Inhibition of cytochrome P450 (CYP3A4) activity by extracts from 57 plants used in traditional chinese medicine (TCM). Pharmacog. Mag. 2017, 13, 300–308. [Google Scholar]
- Wang, J.-S.; Backman, J.T.; Kivistö, K.T.; Neuvonen, P. Effects of metronidazole on midazolam metabolism in vitro and in vivo. Eur. J. Clin. Pharmacol. 2000, 56, 555–559. [Google Scholar] [CrossRef]
- Schenkman, J.B.; Thummel, K.E.; Favreau, L.V. Physiological and pathophysiological alterations in rat hepatic cytochrome P450. Drug Metab. Rev. 1989, 20, 557–584. [Google Scholar] [CrossRef] [PubMed]
- Volberding, P.; Greene, W.; Lange, J.M.; Gallant, J.E.; Sewankambo, N. Sande’s HIV/AIDS Medicine: Medical Management of AIDS 2013; Elsevier Health Sciences: Amsterdam, The Netherlands, 2012. [Google Scholar]
- Kliewer, S.A.; Goodwin, B.; Willson, T.M. The nuclear pregnane X receptor: A key regulator of xenobiotic metabolism. Endocr. Rev. 2002, 23, 687–702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lehmann, J.M.; McKee, D.D.; Watson, M.A.; Willson, T.M.; Moore, J.T.; Kliewer, S.A. The human orphan nuclear receptor PXR is activated by compounds that regulate CYP3A4 gene expression and cause drug interactions. J. Clin. Investig. 1998, 102, 1016–1023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, N.; Hu, M.; Duan, R.; Liu, C.; Guo, H.; Zhang, M.; Yu, Y.; Wang, X.; Liu, L.; Liu, X. Increased levels of fatty acids contributed to induction of hepatic CYP3A4 activity induced by diabetes—In vitro evidence from HepG2 cell and Fa2N-4 cell lines. J. Pharmacol. Sci. 2014, 124, 433–444. [Google Scholar] [CrossRef] [Green Version]
- Ramsay, R.R.; Tipton, K.F. Assessment of enzyme inhibition: A review with examples from the development of monoamine oxidase and cholinesterase inhibitory drugs. Molecules 2017, 22, 1192. [Google Scholar] [CrossRef] [Green Version]
- Meech, R.; Miners, J.O.; Lewis, B.C.; Mackenzie, P.I. The glycosidation of xenobiotics and endogenous compounds: Versatility and redundancy in the UDP glycosyltransferase superfamily. Pharmacol. Therap. 2012, 134, 200–218. [Google Scholar] [CrossRef]
- Buckley, D.B.; Klaassen, C.D. Induction of mouse UDP-glucuronosyltransferase mRNA expression in liver and intestine by activators of aryl-hydrocarbon receptor, constitutive androstane receptor, pregnane X receptor, peroxisome proliferator-activated receptor α, and nuclear factor erythroid 2-related factor 2. Drug Metab. Dispos. 2009, 37, 847–856. [Google Scholar]
- Dostalek, M.; Hazarika, S.; Akhlaghi, F. Diabetes mellitus reduces activity of human UDP-glucuronosyltransferase 2B7 in liver and kidney leading to decreased formation of mycophenolic acid acyl-glucuronide metabolite. Drug Metab. Dispos. 2011, 39, 448–455. [Google Scholar] [CrossRef] [Green Version]
- Thabet, A.A.; Youssef, F.S.; Korinek, M.; Chang, F.-R.; Wu, Y.-C.; Chen, B.-H.; El-Shazly, M.; Singab, A.N.B.; Hwang, T.-L. Study of the anti-allergic and anti-inflammatory activity of Brachychiton rupestris and Brachychiton discolor leaves (Malvaceae) using in vitro models. BMC Complement. Altern. Med. 2018, 18, 299. [Google Scholar] [CrossRef] [Green Version]
- Al-Madhagy, S.A.; Mostafa, N.M.; Youssef, F.S.; Awad, G.E.; Eldahshan, O.A.; Singab, A.N.B. Metabolic profiling of a polyphenolic-rich fraction of Coccinia grandis leaves using LC-ESI-MS/MS and in vivo validation of its antimicrobial and wound healing activities. Food Funct. 2019, 10, 6267–6275. [Google Scholar] [CrossRef]
- Qureshi, H.; Asif, S.; Ahmed, H.; Al-Kahtani, H.A.; Hayat, K. Chemical composition and medicinal significance of Fagonia cretica: A review. Nat. Prod. Res. 2016, 30, 625–639. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, A.A.; Elsammani, T.O.; Elfeil, M.E.; Ahmed, S.K. The pharmacological effects of Fagonia cretica Linn ethanolic extract on isolated rabbit intestine. Int. J. Pharmacol. Toxicol. 2013, 1, 91–98. [Google Scholar] [CrossRef]
- Anjum, M.I.; Ahmed, E.; Jabbar, A.; Malik, A.; Ashraf, M.; Moazzam, M.; Rasool, M.A. Antimicrobial constituents from Fagonia cretica. J. Chem. Soc. Pakistan. 2007, 29, 634–639. [Google Scholar]
- Mahmood, A.; Qureshi, R.A.; Mahmood, A.; Sangi, Y.; Shaheen, H.; Ahmad, I.; Nawaz, Z.J.J.M.P.R. Ethnobotanical survey of common medicinal plants used by people of district Mirpur, AJK, Pakistan. J. Med. Plant Res. 2011, 5, 4493–4498. [Google Scholar]
- Yaseen, H.S.; Asif, M.; Saadullah, M.; Asghar, S.; Shams, M.U.; Bazmi, R.R.; Saleem, M.; Yousaf, H.M.; Yaseen, M.J.I. Methanolic extract of Ephedra ciliata promotes wound healing and arrests inflammatory cascade in vivo through downregulation of TNF-α. Inflammopharmacology 2020, 28, 1691–1704. [Google Scholar] [CrossRef]
- Chang, W.-C.; Huang, D.-W.; Chen, J.-A.; Chang, Y.-F.; Wu, J.S.-B.; Shen, S.-C.J.R.a. Protective effect of Ruellia tuberosa L. extracts against abnormal expression of hepatic detoxification enzymes in diabetic rats. RSC Adv. 2018, 8, 21596–21605. [Google Scholar] [CrossRef]
- Tsunoda, S.M.; Velez, R.L.; von Moltke, L.L.; Greenblatt, D. Differentiation of intestinal and hepatic cytochrome P450 3A activity with use of midazolam as an in vivo probe: Effect of ketoconazole. Clin. Pharmacol. Therap. 1999, 66, 461–471. [Google Scholar] [CrossRef]
- Fromm, M.F.; Eckhardt, K.; Li, S.; Schänzle, G.; Hofmann, U.; Mikus, G.; Eichelbaum, M.J.P. Loss of analgesic effect of morphine due to coadministration of rifampin. Pain 1997, 72, 261–267. [Google Scholar] [CrossRef]
- Graham, J.J.T. Homogenization of mammalian tissues. Sci. World J. 2002, 2, 1626–1629. [Google Scholar] [CrossRef] [Green Version]
- Fang, C.J. Expression of Hepatic Cytochrome P450 Drug Metabolizing Enzymes in Diabetes and Diabetic Nephropathy. Ph.D. Thesis, The University of Western Ontario, London, ON, Canada, 2018. [Google Scholar]
- Invitrogen Life Technologies. Available online: https://www.biorxiv.org/content/10.1101/2022.03.23.485541v1.full.pdf (accessed on 23 November 2022).
- Ramakers, C.; Ruijter, J.M.; Deprez, R.H.L.; Moorman, A.F. Assumption-free analysis of quantitative real-time polymerase chain reaction (PCR) data. Neurosci. Lett. 2003, 339, 62–66. [Google Scholar] [CrossRef]
- Matheux, A.; Gassiot, M.; Fromont, G.; Leenhardt, F.; Boulahtouf, A.; Fabbrizio, E.; Pourquier, P. PXR modulates the prostate cancer cell response to afatinib by regulating the expression of the monocarboxylate transporter SLC16A1. Cancers 2021, 13, 3635. [Google Scholar] [CrossRef] [PubMed]
- Hao, H.; Zhang, L.; Jiang, S.; Sun, S.; Gong, P.; Xie, Y.; Zhou, X.; Wang, G. Thioacetamide intoxication triggers transcriptional up-regulation but enzyme inactivation of UDP-glucuronosyltransferases. Drug Metab. Dispos. 2011, 39, 1815–1822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kemp, P.; Sneed, G.; Kupiec, T.; Spiehler, V. Validation of a microtiter plate ELISA for screening of postmortem blood for opiates and benzodiazepines. J. Anal.Toxicol. 2002, 26, 504–512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tiscione, N.B.; Shan, X.; Alford, I.; Yeatman, D.T. Quantitation of benzodiazepines in whole blood by electron impact-gas chromatography-mass spectrometry. J. Anal.Toxicol. 2008, 32, 644–652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaki, Y.; Fujishiro, M.; Lee, X.-P.; Kato, A.; Sakamoto, Y.; Hashimoto, M.; Minohara, S.; Hasegawa, C.; Hasegawa, I.; Dohi, K. Sensitive determination of midazolam and propofol in human plasma by GC–MS/MS. Forensic Toxicol. 2020, 38, 409–419. [Google Scholar] [CrossRef]
- Coulter, C.; Tuyay, J.; Taruc, M.; Moore, C. Semi-quantitative analysis of drugs of abuse, including tetrahydrocannabinol in hair using aqueous extraction and immunoassay. Forensic Sci. Int. 2010, 196, 70–73. [Google Scholar] [CrossRef]
- Kaur, P.; Chamberlin, A.R.; Poulos, T.L.; Sevrioukova, I.F. Structure-based inhibitor design for evaluation of a CYP3A4 pharmacophore model. J. Med.Chem. 2016, 59, 4210–4220. [Google Scholar] [CrossRef] [Green Version]
- Labib, R.M.; Srivedavyasasri, R.; Youssef, F.S.; Ross, S.A. Secondary metabolites isolated from Pinus roxburghii and interpretation of their cannabinoid and opioid binding properties by virtual screening and in vitro studies. Saudi Pharm. J. 2018, 26, 437–444. [Google Scholar] [CrossRef]
- Talaat, A.N.; Ebada, S.S.; Labib, R.M.; Esmat, A.; Youssef, F.S.; Singab, A.N.B. Verification of the anti-inflammatory activity of the polyphenolic-rich fraction of Araucaria bidwillii Hook. Using phytohaemagglutinin-stimulated human peripheral blood mononuclear cells and virtual screening. J. Ethnopharmacol. 2018, 226, 44–47. [Google Scholar] [CrossRef]
- Sweilam, S.H.; Alqarni, M.H.; Youssef, F.S. Antimicrobial alkaloids from marine-derived fungi as drug leads versus COVID-19 infection: A computational approach to explore their Anti-COVID-19 activity and ADMET properties. Evid. Based Complement. Alt. Med. 2022, 2022, 5403757. [Google Scholar] [CrossRef]
- Altyar, A.E.; Ashour, M.L.; Youssef, F.S. Premna odorata: Seasonal metabolic variation in the essential oil composition of its leaf and verification of its anti-ageing potential via in vitro assays and molecular modelling. Biomolecules 2020, 10, 879. [Google Scholar] [CrossRef] [PubMed]
- Patel, D.; Kumar, V. Phytochemical analysis & in-vitro anti obesity activity of different fractions of methanolic extract of Fagonia cretica L. Int. J. Pharm.Sci. Drug Res. 2020, 12, 282–286. [Google Scholar]
- Choi, J.-S.; Piao, Y.-J.; Kang, K.W. Effects of quercetin on the bioavailability of doxorubicin in rats: Role of CYP3A4 and P-gp inhibition by quercetin. Arch. Pharmacal Res. 2011, 34, 607–613. [Google Scholar] [CrossRef]
- Pu, Q.-H.; Shi, L.; Yu, C. Time-dependent inhibition of CYP3A4 by gallic acid in human liver microsomes and recombinant systems. Xenobiotica 2015, 45, 213–217. [Google Scholar] [CrossRef]
- Elhady, S.S.; Youssef, F.S.; Alahdal, A.M.; Almasri, D.M.; Ashour, M.L. Anti-hyperglycaemic evaluation of Buddleia indica leaves using in vitro, in vivo and in silico studies and its correlation with the major phytoconstituents. Plants 2021, 10, 2351. [Google Scholar] [CrossRef]
- Gwilt, P.R.; Nahhas, R.R.; Tracewell, W.G. The effects of diabetes mellitus on pharmacokinetics and pharmacodynamics in humans. Clin. Pharmacokinet. 1991, 20, 477–490. [Google Scholar] [CrossRef] [PubMed]
- Kamran, S.H.; Shoaib, R.M.; Ahmad, M.; Ishtiaq, S.; Anwar, R. Antidiabetic and renoprotective effect of Fagonia cretica L. methanolic extract and Citrus paradise Macfad. Juice in alloxan induced diabetic rabbits. J. Pharm Pharmcog. Res. 2017, 5, 365–380. [Google Scholar]
- Mandona, J.W.; Tuk, B.; van Steveninck, A.L.; Breimer, D.D.; Cohen, A.F.; Danhof, M. Pharmacokinetic-pharmacodynamic modeling of the central nervous system effects of midazolam and its main metabolite α-hydroxymidazolam in healthy volunteers. Clin. Pharm. Ther. 1992, 51, 715–728. [Google Scholar] [CrossRef]
- De Gregori, S.; De Gregori, M.; Ranzani, G.N.; Allegri, M.; Minella, C.; Regazzi, M.J. Morphine metabolism, transport and brain disposition. Metab. Brain Dis. 2012, 27, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Armstrong, S.C.; Cozza, K.L. Pharmacokinetic drug interactions of morphine, codeine, and their derivatives: Theory and clinical reality, Part II. Psychosomatics 2003, 44, 515–520. [Google Scholar] [CrossRef] [Green Version]
- Yuan, R.; Flockhart, D.A.; Balian, J.D. Pharmacokinetic and pharmacodynamic consequences of metabolism-based drug interactions with alprazolam, midazolam, and triazolam. J. Clin. Pharmacol. 1999, 39, 1109–1125. [Google Scholar] [CrossRef] [PubMed]
- Thummel, K.; Wilkinson, G. In vitro and in vivo drug interactions involving human CYP3A. Annu. Rev. Pharmacol. Toxicol. 1998, 38, 389–430. [Google Scholar] [CrossRef] [PubMed]
- Glare, P.; Walsh, T.J. Clinical pharmacokinetics of morphine. Ther. Drug Monit. 1991, 13, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Lee, M.G. Telithromycin pharmacokinetics in rat model of diabetes mellitus induced by alloxan or streptozotocin. Pharm. Res. 2008, 25, 1915–1924. [Google Scholar] [CrossRef] [PubMed]
- Sinz, M.W. Evaluation of pregnane X receptor (PXR)-mediated CYP3A4 drug-drug interactions in drug development. Drug Metab. Rev. 2013, 45, 3–14. [Google Scholar] [CrossRef]
- Xu, C.; Gao, J.; Zhang, H.-F.; Gao, N.; Guo, Y.-Y.; Fang, Y.; Wen, Q.; Qiao, H.-L. Content and activities of UGT2B7 in human liver in vitro and predicted in vivo: A bottom-up approach. Drug Metab. Dispos. 2018, 46, 1351–1359. [Google Scholar] [CrossRef] [PubMed]
- Jeong, H.; Choi, S.; Song, J.; Chen, H.; Fischer, J.H. Regulation of UDP-glucuronosyltransferase (UGT) 1A1 by progesterone and its impact on labetalol elimination. Xenobiotica 2008, 38, 62–75. [Google Scholar] [CrossRef] [Green Version]
- Xie, W.; Uppal, H.; Saini, S.P.; Mu, Y.; Little, J.M.; Radominska-Pandya, A.; Zemaitis, M.A. Orphan nuclear receptor-mediated xenobiotic regulation in drug metabolism. Drug Dis. Today 2004, 9, 442–449. [Google Scholar] [CrossRef]
- Jones, S.A.; Moore, L.B.; Shenk, J.L.; Wisely, G.B.; Hamilton, G.A.; McKee, D.D.; Tomkinson, N.C.; LeCluyse, E.L.; Lambert, M.H.; Willson, T.M. The pregnane X receptor: A promiscuous xenobiotic receptor that has diverged during evolution. Mol. Endocrinol. 2000, 14, 27–39. [Google Scholar] [CrossRef]
- Zanger, U.M.; Schwab, M. Cytochrome P450 enzymes in drug metabolism: Regulation of gene expression, enzyme activities, and impact of genetic variation. Pharmacol. Ther. 2013, 138, 103–141. [Google Scholar] [CrossRef]
- Salam, R.; Batool Rizvi, S.N.; Hussain, N.; Firdous, S.; Zaheer, M.; Naeem, M. Role of hesperidin and fresh orange juice in altering the bioavailability of beta-blocker, metoprolol tartrate. An in vivo model. Xenobiotica 2022, 52, 295–300. [Google Scholar] [CrossRef] [PubMed]
- Dasgupta, A. Alcohol, Drugs, Genes and the Clinical Laboratory; Elsevie: Amsterdam, The Netherlands, 2017. [Google Scholar]
- Heiskanen, T.; Kalso, E. Non-analgesic effects of opioids: Interactions between opioids and other drugs. Curr. Pharm. Des. 2012, 18, 6079–6089. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds Name | Methanol Extract | Aqueous Extract | ||
|---|---|---|---|---|
| Retention Time (min) | Amount (ppm) | Retention Time (min) | Amount (ppm) | |
| Quercetin | 2.707 | 8.28 | 2.727 | 7.82 |
| Gallic acid | 4.247 | 2.52 | 4.240 | 2.43 |
| Vanillic acid | 13.627 | 6.72 | 13.400 | 5.64 |
| Benzoic acid | 14.667 | 11.21 | 14.533 | 8.66 |
| m-Coumaric acid | 20.367 | 2.14 | 19.680 | 0.97 |
| Cinnamic acid | 25.040 | 4.25 | - | - |
| Sinapic acid | 26.013 | 2.01 | 26.453 | 1.64 |
| Drug | Nominal Value | Day-1 | Day-2 | Day-3 | Grand Average | STD | % CV | % Bias | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Run1 | Run2 | Run3 | Run1 | Run2 | Run3 | Run1 | Run2 | Run3 | ||||||
| Midazolam | 100 | 112.5 | 91.8 | 105.7 | 100.8 | 97.8 | 93.4 | 111.7 | 103.9 | 101.5 | 102.1 | 7.24 | 7.09 | 2.12 |
| 300 | 325.4 | 311.6 | 305.4 | 325.8 | 304.5 | 298.8 | 278.2 | 318.7 | 308.5 | 308.6 | 14.74 | 4.78 | 2.85 | |
| 500 | 517.2 | 467.8 | 506.8 | 496.7 | 546.9 | 508.9 | 510.2 | 481.7 | 555.9 | 510.2 | 28.04 | 5.50 | 2.05 | |
| Alpha Hydroxyl Midazolam | 100 | 104.6 | 99.6 | 105.5 | 109.5 | 97.8 | 95.6 | 102.8 | 106.8 | 110.3 | 103.6 | 5.11 | 4.93 | 3.61 |
| 300 | 291.2 | 301.5 | 316.5 | 308.6 | 278.9 | 292.7 | 310.8 | 322.7 | 333.4 | 306.3 | 17.03 | 5.56 | 2.08 | |
| 500 | 496.8 | 472.7 | 545.8 | 542.9 | 545.8 | 509.4 | 515.7 | 529.3 | 501.9 | 517.8 | 25.33 | 4.89 | 3.56 | |
| Morphine | 40 | 36.3 | 39.2 | 41.8 | 37.2 | 43.7 | 41.0 | 44.1 | 40.4 | 46.2 | 41.1 | 3.25 | 7.91 | 2.75 |
| 200 | 197.5 | 211.6 | 178.6 | 217.7 | 208.3 | 205.5 | 217.1 | 195.6 | 222.6 | 206.1 | 13.71 | 6.65 | 3.03 | |
| 400 | 439.0 | 417.9 | 388.2 | 391.7 | 412.9 | 444.7 | 369.8 | 433.0 | 420.2 | 413.0 | 25.23 | 6.11 | 3.26 | |
| Compound | CYP3A4 (4D7D) | Number of Formed Hydrogen Bonds and C-H Bonds | Number of Formed π-π and π-alkyl Bonds |
|---|---|---|---|
| Benzoic acid | −12.30 | 2; Ser119 | 1; Phe304 |
| Cinnamic acid | −17.32 | 2; Cys442, Ile443 | - |
| Gallic acid | −23.12 | 2; Ser119 | 1; Phe304 |
| m-Coumaric_acid | −18.41 | 1; Cys442 | 1; Ala305 |
| Quercetin | −30.83 | 2; Cys442 | 3; Ala305, Ile301 |
| Sinapic acid | −22.47 | 5; Arg375, Arg105, Leu373, Phe435, Arg372 | 1; Ala370 |
| Vanillic acid | −19.42 | 1; Glu308 | - |
| Ketoconazole | −30.16 | - | 10; Phe435, Ala370, Phe108, Leu211, Ala305, Phe304, Ile301 |
| Co-crystalized ligand | −46.88 | 1; Arg375 | 6; Leu, Ala370, Leu211, Phe304, Ile301, Phe108 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kamran, S.; Anwar, R.; Noor, A.; Ullah, M.I.; Bagalagel, A.A.; Aldurdunji, M.M.; Ishtiaq, S. Metabolic Profiling and Investigation of the Modulatory Effect of Fagonia cretica L. Aerial Parts on Hepatic CYP3A4 and UGT2B7 Enzymes in Streptozotocin—Induced Diabetic Model. Antioxidants 2023, 12, 119. https://doi.org/10.3390/antiox12010119
Kamran S, Anwar R, Noor A, Ullah MI, Bagalagel AA, Aldurdunji MM, Ishtiaq S. Metabolic Profiling and Investigation of the Modulatory Effect of Fagonia cretica L. Aerial Parts on Hepatic CYP3A4 and UGT2B7 Enzymes in Streptozotocin—Induced Diabetic Model. Antioxidants. 2023; 12(1):119. https://doi.org/10.3390/antiox12010119
Chicago/Turabian StyleKamran, Shahzad, Rukhsana Anwar, Afifa Noor, Muhammad Ihsan Ullah, Alaa A. Bagalagel, Mohammed M. Aldurdunji, and Saiqa Ishtiaq. 2023. "Metabolic Profiling and Investigation of the Modulatory Effect of Fagonia cretica L. Aerial Parts on Hepatic CYP3A4 and UGT2B7 Enzymes in Streptozotocin—Induced Diabetic Model" Antioxidants 12, no. 1: 119. https://doi.org/10.3390/antiox12010119