
Effects of Supplementing Sea Buckthorn Leaves (Hippophae rhamnoides L.) and Chromium (III) in Broiler Diet on the Nutritional Quality and Lipid Oxidative Stability of Meat
 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Broiler Performance
2.3. Sample Collection
2.4. Analysis of Plant and Meat Nutrients
2.5. Analysis of Antioxidant Capacity and Lipid Oxidation Status
2.6. Statistical Analysis
3. Results
3.1. Chemical Composition of Sea Buckthorn Leaves (SBL)
3.2. Broiler Performance and Carcass and Cuts Yield
3.3. Proximate Composition of Chicken Meat
3.4. Fatty Acid Profile of Chicken Meat

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fatty Acids | Treatment | SEM | p-value | Treatment | SEM | p-Value | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| T0 | T1 | T2 | T0 | T1 | T2 | |||||
| g/100 g FAME | mg/100 g meat | |||||||||
| C14:0 | 0.60 | 0.42 | 0.48 | 0.018 | 0.161 | 8.84 | 6.49 | 6.94 | 1.120 | 0.285 |
| C15:0 | 0.13 | 0.10 | 0.12 | 0.010 | 0.951 | 1.98 a | 1.60 b | 1.81 ab | 0.261 | 0.048 |
| C16:0 | 18.08 | 18.06 | 18.07 | 0.067 | 0.168 | 266.72 | 269.58 | 279.35 | 0.300 | 0.194 |
| C17:0 | 0.22 b | 0.22 b | 0.20 a | 0.014 | 0.001 | 3.06 | 3.19 | 2.93 | 0.078 | 0.113 |
| C18:0 | 7.57 | 7.76 | 8.20 | 0.117 | 0.139 | 111.14 a | 115.76 ab | 123.01 b | 0.241 | 0.022 |
| C24:0 | 0.25 a | 0.21 b | 0.22 b | 0.012 | 0.0001 | 3.77 a | 3.28 b | 3.35 b | 0.268 | 0.033 |
| Σ Saturated fatty acids | 26.85 | 26.77 | 27.29 | 0.488 | 0.090 | 394.41 | 400.30 | 420.29 | 0.294 | 0.088 |
| C14:1 | 0.09 a | 0.08 ab | 0.07 b | 0.124 | 0.001 | 1.17 | 0.97 | 0.91 | 0.290 | 0.140 |
| C15:1 | 0.55 | 0.63 | 0.71 | 0.181 | 0.121 | 8.19 b | 9.53 ab | 12.17 a | 0.912 | 0.026 |
| C16:1 | 2.52 a | 2.47 ab | 2.39 b | 0.741 | 0.0001 | 37.21 | 36.95 | 35.89 | 0.902 | 0.551 |
| C17:1 | 0.12 b | 0.15 b | 0.21 a | 0.524 | 0.0001 | 1.93 b | 2.40 b | 3.16 a | 0.197 | 0.003 |
| C18:1 | 29.45 a | 29.62 a | 27.90 b | 0.185 | 0.0001 | 433.22 | 441.99 | 418.79 | 9.501 | 0.336 |
| C22:1n9 | 0.08 a | 0.06 b | 0.09 a | 0.125 | 0.003 | 1.37 a | 0.97 b | 1.36 a | 0.082 | 0.009 |
| C24:1n9 | 1.02 b | 1.07 b | 1.38 a | 0.580 | 0.0001 | 15.19 b | 16.08 b | 20.67 a | 0.137 | <0.0001 |
| Σ Monounsaturated fatty acids | 33.81 a | 34.01 a | 32.84 b | 0.125 | 0.0001 | 497.42 a | 508.89 a | 492.96 b | 0.368 | 0.007 |
| C18:2n6 | 32.41 | 32.34 | 32.23 | 0.294 | 0.125 | 476.65 | 482.51 | 468.73 | 0.341 | 0.707 |
| C18:3n6 | 0.27 | 0.29 | 0.27 | 0.325 | 0.230 | 4.21 | 4.51 | 4.28 | 0.322 | 0.394 |
| C20:2n6 | 0.25 a | 0.21 ab | 0.17 b | 0.101 | 0.0001 | 3.87 a | 3.28 a | 2.50 b | 0.175 | 0.001 |
| C20:3n6 | 0.80 | 0.85 | 0.91 | 0.045 | 0.321 | 11.67 b | 10.44 c | 13.70 a | 0.132 | <0.0001 |
| C20:4n6 | 2.74 b | 2.88 b | 3.64 a | 0.370 | 0.0001 | 38.75 b | 43.20 b | 54.59 a | 1.304 | <0.0001 |
| C22:2n6 | 0.15 | 0.15 | 0.16 | 0.451 | 0.355 | 2.37 | 2.25 | 2.71 | 0.298 | 0.182 |
| C22:3n6 | 0.16 | 0.17 | 0.16 | 0.604 | 0.196 | 1.96 | 2.50 | 1.96 | 0.294 | 0.160 |
| C22:4n6 | 0.33 b | 0.29 b | 0.43 a | 0.725 | 0.0001 | 4.97 b | 4.48 b | 6.48 a | 0.117 | <0.0001 |
| Σ n-6 | 37.25 b | 37.07 b | 37.97 a | 0.147 | 0.0001 | 547.34 | 553.17 | 554.95 | 0.381 | 0.917 |
| C18:3n3 | 0.67 a | 0.58 ab | 0.53 b | 0.021 | 0.010 | 10.10 a | 8.75 ab | 8.06 b | 0.247 | 0.028 |
| C18:4n3 | 0.23 b | 0.29 a | 0.36 a | 0.001 | 0.0001 | 2.84 c | 4.53 b | 5.45 a | <0.0001 | <0.0001 |
| C20:3n3 | 0.49 b | 0.47 b | 0.63 a | 0.009 | 0.0001 | 7.32 b | 7.24 b | 9.51 a | 0.147 | 0.0001 |
| C20:5n3 | 0.21 a | 0.19 b | 0.18 b | 0.002 | 0.0001 | 3.26 a | 2.55 b | 2.65 b | 0.207 | 0.005 |
| C22:5n3 | 0.10 b | 0.11 b | 0.15 a | 0.001 | 0.0001 | 1.66 b | 1.78 b | 2.27 a | 0.079 | 0.001 |
| C22:6n3 | 0.05 b | 0.06 b | 0.10 a | 0.002 | 0.0001 | 1.08 b | 1.08 b | 1.52 a | 0.096 | 0.012 |
| Σ n-3 | 1.75 b | 1.70 b | 1.95 a | 0.029 | 0.0001 | 26.30 b | 25.93 b | 29.46 a | 0.227 | 0.009 |
| Σ Polyunsaturated fatty acids | 39.05 | 38.77 | 39.92 | 0.153 | 0.158 | 573.64 | 579.11 | 584.40 | 0.379 | 0.865 |
| Other fatty acids | 0.23 | 0.23 | 0.22 | 0.074 | 0.111 | 3.33 | 3.42 | 3.26 | 0.351 | 0.950 |
| Fatty Acids | Treatment | SEM | p-Value | Treatment | SEM | p-Value | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| T0 | T1 | T2 | T0 | T1 | T2 | |||||
| g/100 g FAME | mg/100 g meat | |||||||||
| C14:0 | 0.49 | 0.47 | 0.49 | 0.032 | 0.188 | 18.36 b | 17.15 b | 22.16 a | 0.217 | 0.008 |
| C15:0 | 0.18 | 0.18 | 0.19 | 0.011 | 0.063 | 7.15 | 5.64 | 6.69 | 0.272 | 0.249 |
| C16:0 | 19.13 ab | 18.87 b | 19.99 a | 0.720 | 0.040 | 703.12 ab | 699.40 b | 888.05 a | 0.128 | <0.0001 |
| C17:0 | 0.22 b | 0.24 ab | 0.26 a | 0.002 | 0.0001 | 8.22 b | 8.73 ab | 11.36 a | 0.120 | <0.0001 |
| C18:0 | 7.15 b | 7.96 a | 7.31 b | 0.036 | 0.0001 | 262.78 c | 295.24 b | 325.01 a | 0.145 | <0.0001 |
| Σ Saturated fatty acids | 27.44 | 27.69 | 28.52 | 0.933 | 0.061 | 1008.93 b | 1026.15 b | 1267.62 a | 0.109 | <0.0001 |
| C14:1 | 0.09 | 0.07 | 0.07 | 0.005 | 0.324 | 3.56 | 3.06 | 3.28 | 0.290 | 0.648 |
| C15:1 | 0.92 | 0.84 | 0.89 | 0.015 | 0.231 | 33.34 | 30.78 | 39.37 | 0.233 | 0.025 |
| C16:1 | 3.32 a | 2.94 b | 3.15 ab | 0.028 | 0.0001 | 122.46 b | 109.45 c | 140.12 a | 0.147 | <0.0001 |
| C17:1 | 0.17 b | 0.14 b | 0.24 a | 0.025 | 0.002 | 5.79 b | 4.79 b | 10.55 a | 0.170 | 0.0001 |
| C18:1 | 29.40 a | 29.41 a | 28.31 b | 0.215 | 0.0001 | 1088.74 b | 1089.95 b | 1259.48 a | 0.157 | <0.0001 |
| C22:1n9 | 0.08 | 0.07 | 0.07 | 0.002 | 0.125 | 2.63 | 2.41 | 2.97 | 0.280 | 0.392 |
| C24:1n9 | 0.58 b | 0.87 a | 0.74 ab | 0.022 | 0.025 | 21.58 b | 32.61 a | 33.19 a | 0.221 | 0.011 |
| Σ Monounsaturated fatty acids | 34.56 a | 34.34 a | 33.48 b | 0.124 | 0.0001 | 1278.11 b | 1273.05 b | 1488.96 a | 0.139 | <0.0001 |
| C18:2n6 | 31.26 | 31.16 | 30.92 | 0.122 | 0.185 | 1150.38 b | 1154.51 b | 1375.25 a | 0.128 | <0.0001 |
| C18:3n6 | 0.20 b | 0.22 ab | 0.25 a | 0.004 | 0.0001 | 7.83 b | 8.46 b | 11.32 a | 0.141 | <0.0001 |
| C20:2n6 | 0.29 | 0.27 | 0.28 | 0.006 | 0.077 | 10.36 | 9.80 | 7.88 | 0.275 | 0.292 |
| C20:3n6 | 0.49 | 0.50 | 0.45 | 0.003 | 0.233 | 18.07 b | 19.96 ab | 22.32 a | 0.232 | 0.023 |
| C20:4n6 | 2.82 | 2.84 | 2.80 | 0.583 | 0.987 | 103.68 b | 115.12 ab | 122.88 a | 0.233 | 0.025 |
| C22:2n6 | 0.18 | 0.19 | 0.21 | 0.019 | 0.088 | 6.91 | 6.49 | 9.19 | 0.246 | 0.056 |
| C22:3n6 | 0.16 c | 0.32 a | 0.23 b | 0.143 | 0.0001 | 5.55 c | 19.96 a | 10.10 b | 0.079 | <0.0001 |
| C22:4n6 | 0.22 | 0.24 | 0.24 | 0.150 | 0.058 | 6.90 b | 7.33 b | 10.53 a | 0.212 | 0.006 |
| Σ n-6 | 35.62 | 35.74 | 35.38 | 0.350 | 0.125 | 1309.68 b | 1324.67 b | 1569.47 a | 0.136 | <0.0001 |
| C18:3n3 | 0.57 a | 0.52 b | 0.54 ab | 0.002 | 0.0001 | 19.52 b | 18.68 b | 23.10 a | 0.415 | <0.0001 |
| C18:4n3 | 0.35 | 0.34 | 0.29 | 0.021 | 0.241 | 13.86 | 11.93 | 13.57 | 0.264 | 0.158 |
| C20:3n3 | 0.34 | 0.35 | 0.40 | 0.084 | 0.711 | 12.59 | 13.58 | 17.85 | 0.252 | 0.082 |
| C20:5n3 | 0.43 | 0.49 | 0.47 | 0.018 | 0.115 | 16.11 | 18.81 | 13.39 | 1.765 | 0.129 |
| C22:5n3 | 0.11 | 0.09 | 0.07 | 0.005 | 0.065 | 4.23 | 4.30 | 3.70 | 0.284 | 0.484 |
| C22:6n3 | 0.07 ab | 0.05 b | 0.10 a | 0.007 | 0.026 | 2.51 b | 2.01 b | 4.47 a | 0.213 | 0.006 |
| Σ n-3 | 1.87 | 1.84 | 1.87 | 0.025 | 0.624 | 56.73 b | 68.45 b | 78.90 a | 0.142 | <0.0001 |
| Σ Polyunsaturated fatty acids | 37.50 | 37.58 | 37.24 | 0.551 | 0.175 | 1380.98 | 1393.12 | 1645.55 | 0.259 | 0.122 |
| Other fatty acids | 0.50 a | 0.40 a | 0.76 b | 0.071 | 0.234 | 13.17 b | 17.79 b | 44.76 a | 0.125 | <0.0001 |
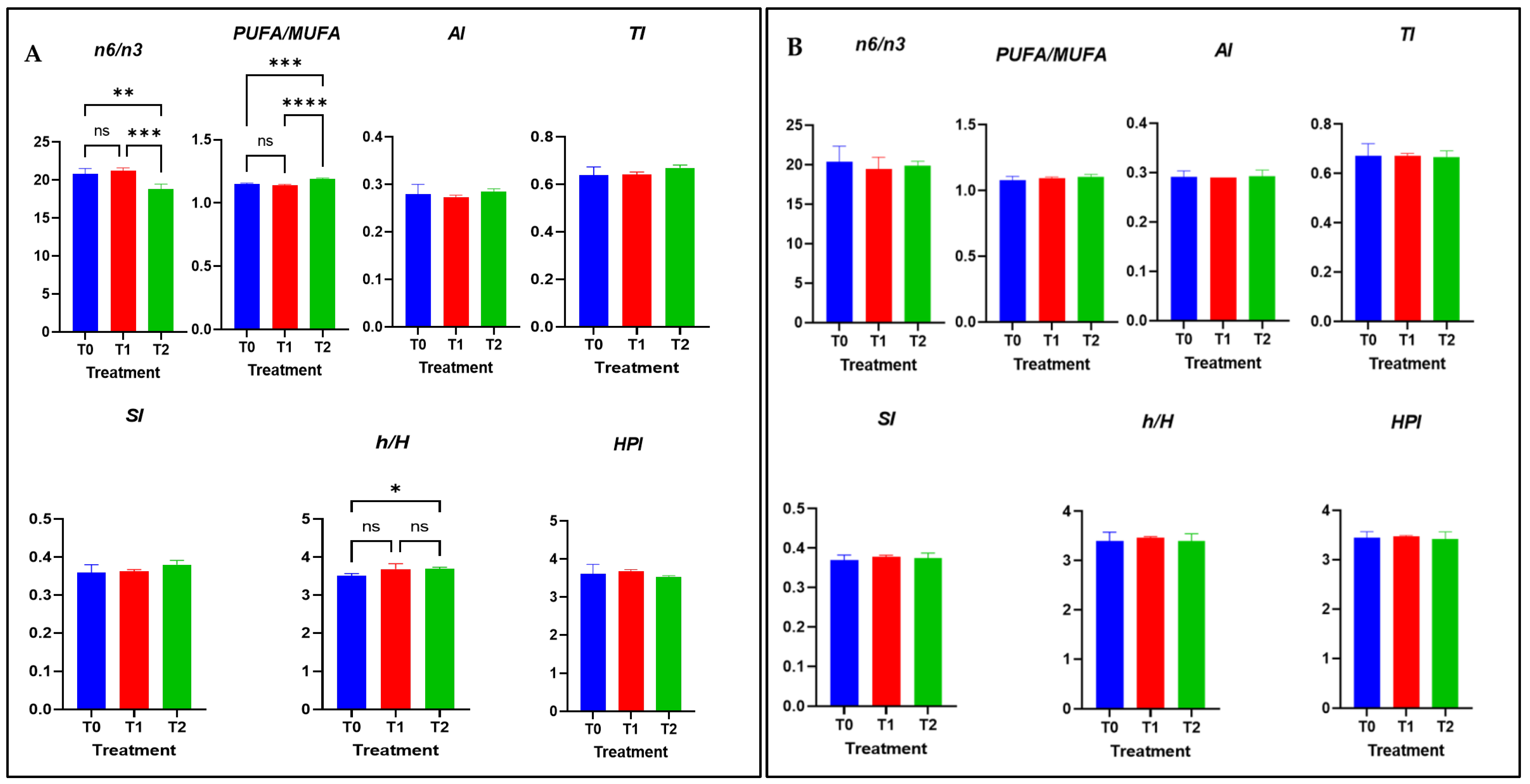
3.5. Nutritional Quality Indices of Meat Lipids
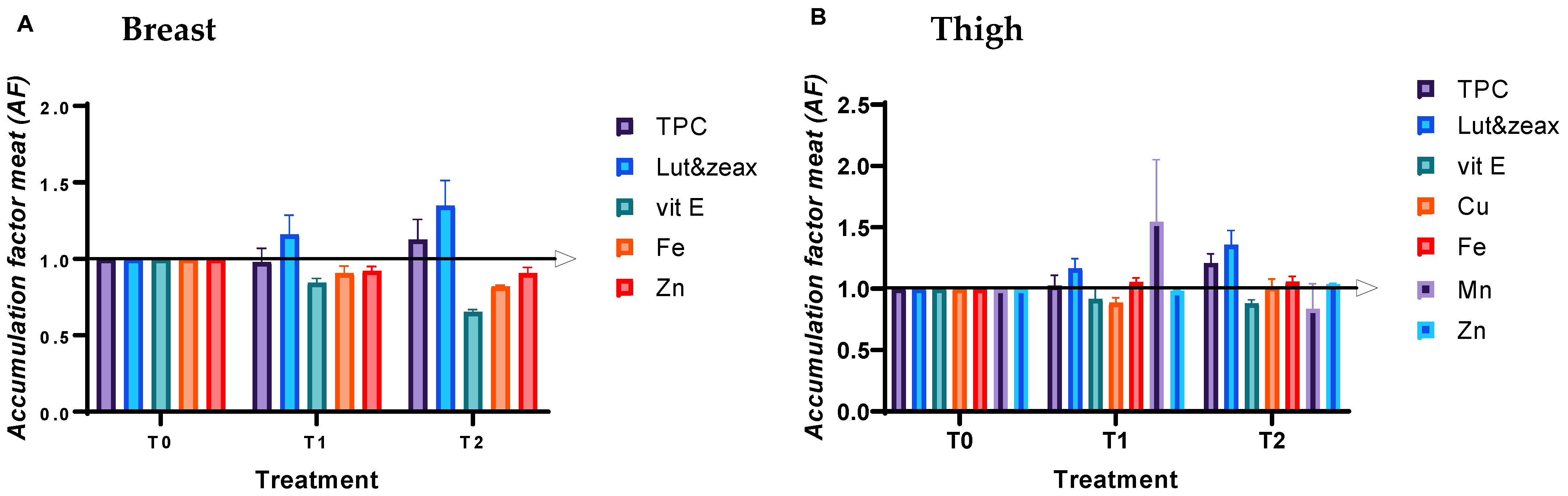
3.6. Bioactive Nutrient Content of Breast and Thigh Meat
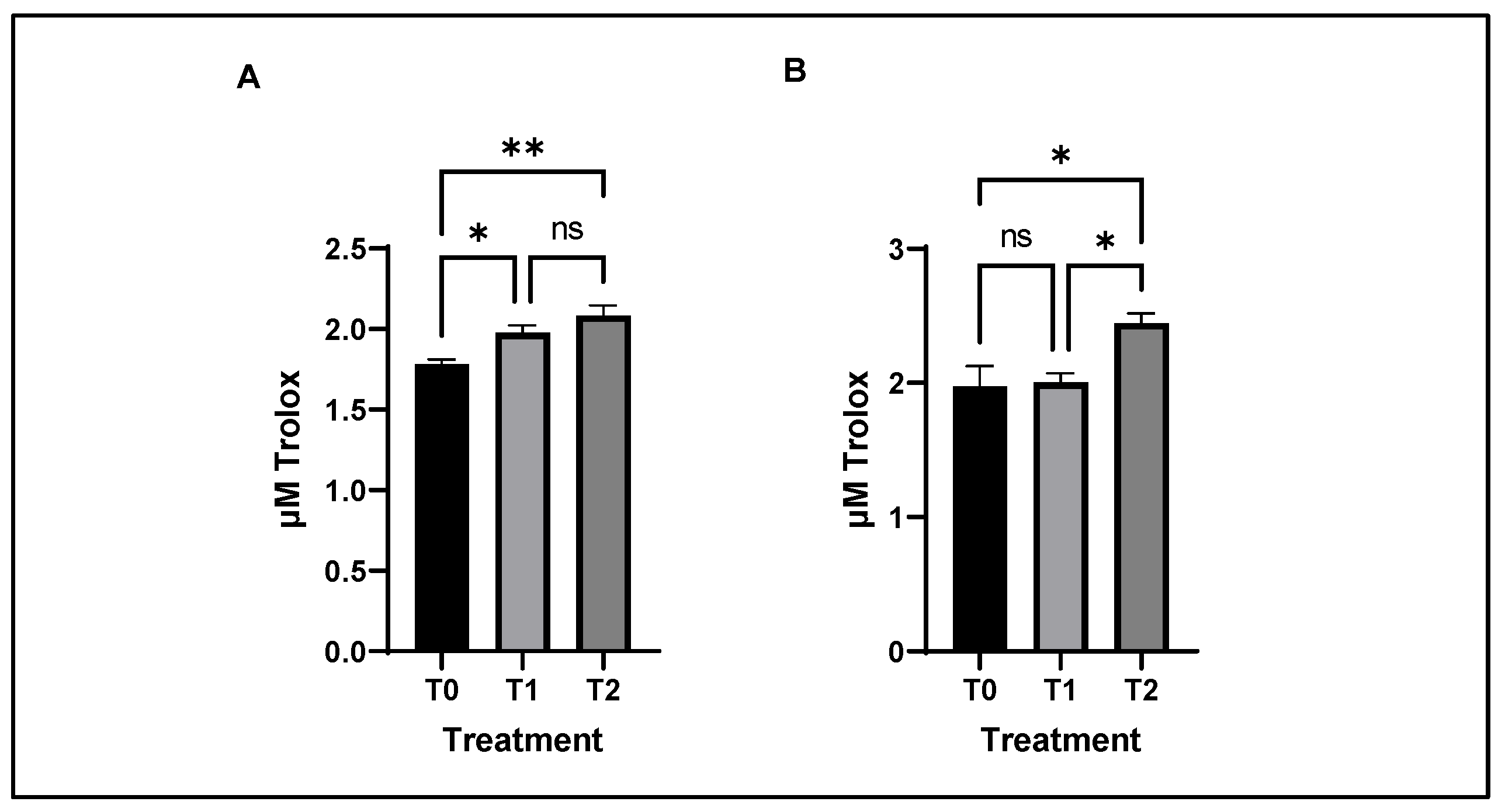
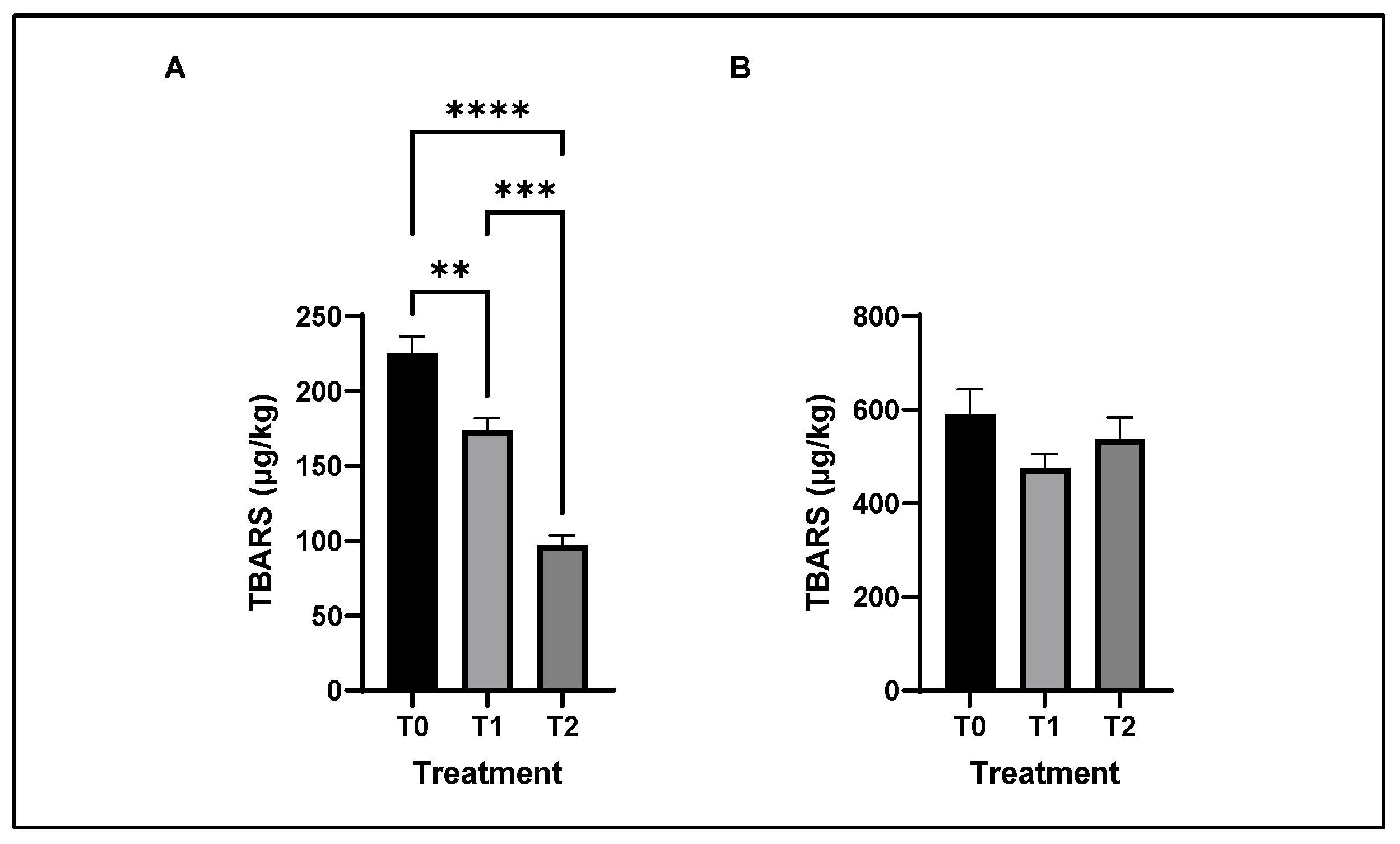
3.7. Antioxidant and Lipid Oxidative Stability of Chicken Meat
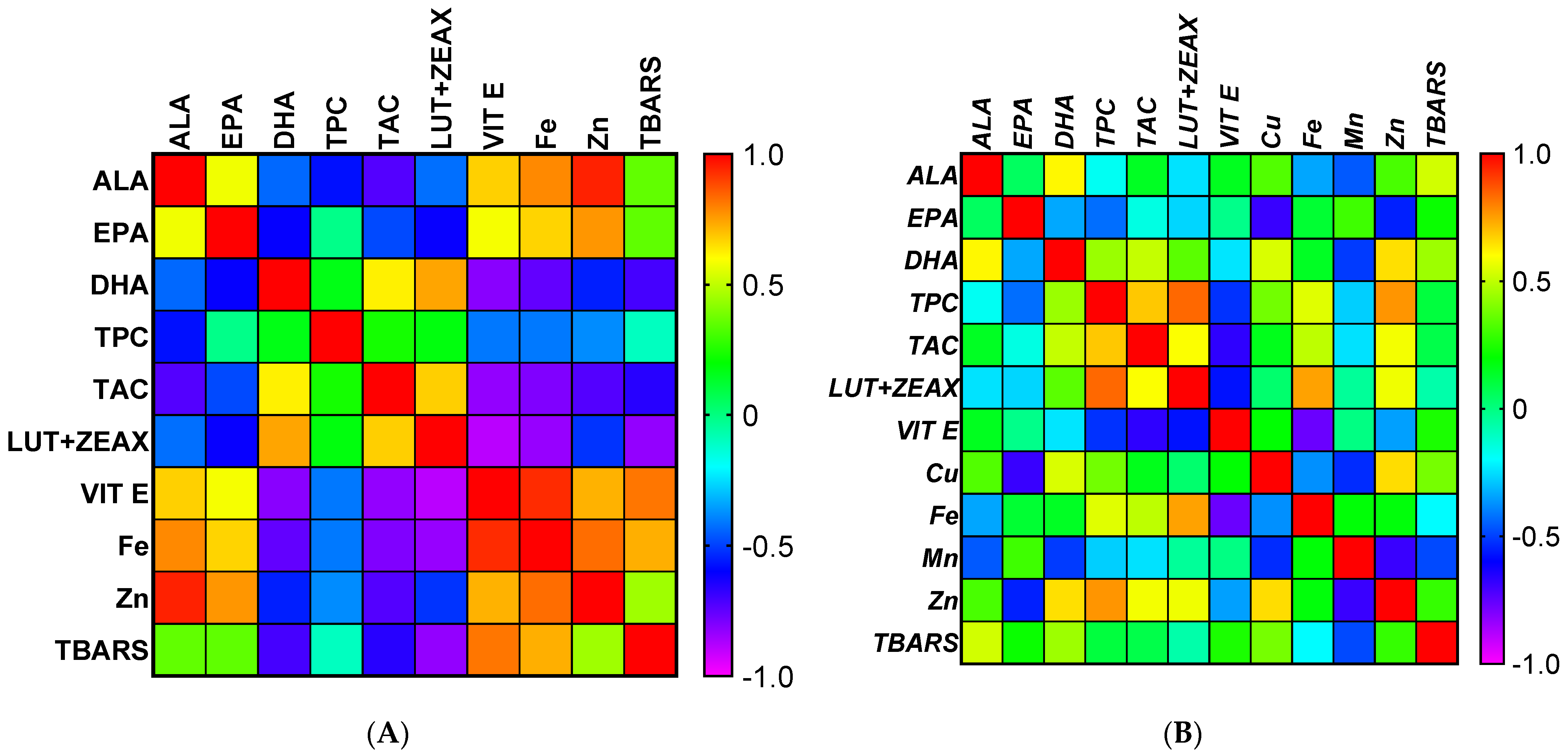
3.8. The Relationship between Meat Characteristics Given by Pearson Correlation Matrix
4. Discussion
4.1. Chemical Composition of Sea Buckthorn Leaves (SBL)
4.2. Broiler Performance and Carcass and Cuts Yiled
4.3. Proximate Composition of Chicken Meat
4.4. Fatty Acid Profile of Chicken Meat
4.5. Nutritional Quality Indices of Meat Lipids
4.6. Bioactive Nutrient Content of Breast and Thigh Meat
4.7. Lipid Oxidation Status of Chicken Meat
4.8. The Relationship between Meat Characteristics Given by Pearson Correlation Matrix
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Petracci, M.; Bianchi, M.; Mudalal, S.; Cavani, C. Functional ingredients for poultry meat products. Trends Food Sci. Technol. 2013, 33, 27–39. [Google Scholar] [CrossRef]
- Ravindran, V. FAO Poultry Development Review. Poultry Feed Availability and Nutrition in Developing Countries: Main Ingredients Used in Poultry Feed Formulations. 2013. Available online: http://www.fao.org/docrep/019/i3531e/i3531e.pdf (accessed on 2 November 2022).
- World Cancer Research Fund/American Institute for Cancer Research. Food, Nutrition, Physical Activity, and the Prevention of Cancer: A Global Perspective; AICR: Washington, DC, USA, 2007. [Google Scholar]
- Esposito, K.; Chiodini, P.; Maiorino, M.I.; Bellastella, G.; Panagiotakos, D.; Giugliano, D. Which diet for prevention of type 2 diabetes? A meta-analysis of prospective studies. Endocrine 2014, 47, 107–116. [Google Scholar] [CrossRef] [PubMed]
- EFSA Panel on Dietetic Products, Nutrition, and Allergies (NDA). Scientific Opinion on Dietary Reference Values for fats, including saturated fatty acids, polyunsaturated fatty acids, monounsaturated fatty acids, trans fatty acids, and cholesterol. EFSA J. 2010, 8, 1461. [Google Scholar]
- Albergamo, A.; Vadalà, R.; Nava, V.; Bartolomeo, G.; Rando, R.; Colombo, N.; Gualtieri, R.; Petracci, M.; Di Bella, G.; Costa, R. Effect of dietary enrichment with flaxseed, vitamin e and selenium, and of market class on the broiler breast meat—Part 1: Nutritional and functional traits. Nutrients 2022, 14, 1666. [Google Scholar] [CrossRef]
- Vlaicu, P.A.; Untea, A.E.; Turcu, R.P.; Saracila, M.; Panaite, T.D.; Cornescu, G.M. Nutritional composition and bioactive compounds of basil, thyme and sage plant additives and their functionality on broiler thigh meat quality. Foods 2022, 11, 1105. [Google Scholar] [CrossRef]
- Ma, X.; Yang, W.; Kallio, H.; Yang, B. Health promoting properties and sensory characteristics of phytochemicals in berries and leaves of sea buckthorn (Hippophaë rhamnoides). Crit. Rev. Food Sci. Nutr. 2021, 62, 3798–3816. [Google Scholar] [CrossRef]
- Górnaś, P.; Šnē, E.; Siger, A.; Segliņa, D. Sea buckthorn (Hippophae rhamnoides L.) leaves as valuable source of lipophilic antioxidants: The effect of harvest time, sex, drying and extraction methods. Ind. Crops Prod. 2014, 60, 1–7. [Google Scholar] [CrossRef]
- Raudone, L.; Puzerytė, V.; Vilkickyte, G.; Niekyte, A.; Lanauskas, J.; Viskelis, J.; Viskelis, P. Sea buckthorn leaf powders: The impact of cultivar and drying mode on antioxidant, phytochemical, and chromatic profile of valuable resource. Molecules 2021, 26, 4765. [Google Scholar] [CrossRef]
- Radenkovs, V.; Püssa, T.; Juhnevica-Radenkova, K.; Anton, D.; Seglina, D. Phytochemical characterization and antimicrobial evaluation of young leaf/shoot and press cake extracts from Hippophae rhamnoides L. Food Biosci. 2018, 24, 56–66. [Google Scholar] [CrossRef]
- Pap, N.; Reshamwala, D.; Korpinen, R.; Kilpeläinen, P.; Fidelis, M.; Furtado, M.M.; Granato, D. Toxicological and bioactivity evaluation of blackcurrant press cake, sea buckthorn leaves and bark from Scots pine and Norway spruce extracts under a green integrated approach. Food Chem. Toxicol. 2021, 153, 112284. [Google Scholar] [CrossRef]
- Hathwar, S.C.; Rai, A.K.; Modi, V.K.; Narayan, B. Characteristics and consumer acceptance of healthier meat and meat product formulations—A review. J. Food Sci. Technol. 2012, 49, 653–664. [Google Scholar] [CrossRef] [PubMed]
- Aschemann-Witzel, J.; Stangherlin, I.D.C. Upcycled by-product use in agri-food systems from a consumer perspective: A review of what we know, and what is missing. Technol. Forecast. Soc. 2021, 168, 120749. [Google Scholar] [CrossRef]
- FAO. The State of Food and Agriculture 2019. Moving Forward on Food Loss and Waste Reduction. Rome. 2019. Available online: https://www.fao.org/3/ca6030en/ca6030en.pdf (accessed on 28 September 2022).
- Ma, J.S.; Chang, W.H.; Liu, G.H.; Zhang, S.; Zheng, A.J.; Li, Y.; Cai, H.Y. Effects of flavones of sea buckthorn fruits on growth performance, carcass quality, fat deposition and lipometabolism for broilers. Poult. Sci. 2015, 94, 2641–2649. [Google Scholar] [CrossRef] [PubMed]
- Singh, D.N.; Shukla, P.K.; Bhattacharyya, A.; Singh, Y.; Sirohi, R. Effect of breeder and post-hatch dietary supplementation of sea buckthorn leaf meal on growth performance of coloured broiler during summer season. Indian J. Poult. Sci. 2019, 54, 257–262. [Google Scholar] [CrossRef]
- Dhanze, H.; Khurana, S.K.; Mane, B.G. Effect of seabuckthorn leaf extract on microbiological quality of raw chicken during extended periods of storage. J. Food Qual. 2013, 36, 59–65. [Google Scholar] [CrossRef]
- Roussel, A.M.; Andriollo-Sanchez, M.; Ferry, M.; Bryden, N.A.; Anderson, R.A. Food chromium content, dietary chromium intake and related biological variables in French free-living elderly. Br. J. Nutr. 2007, 98, 326–331. [Google Scholar] [CrossRef] [Green Version]
- Arif, M.; Hussain, I.; Mahmood, M.A.; Abd El-Hack, M.E.; Swelum, A.A.; Alagawany, M. Effect of varying levels of chromium propionate on growth performance and blood biochemistry of broilers. Animals 2019, 9, 935. [Google Scholar] [CrossRef] [Green Version]
- Lu, L.; Zhao, L.L.; Dong, S.Y.; Liao, X.D.; Dong, X.Y.; Zhang, L.Y. Dietary supplementation of organic or inorganic chromium modulates the immune responses of broilers vaccinated with avian influenza virus vaccine. Animal 2018, 13, 983–991. [Google Scholar] [CrossRef]
- White, P.E.; Vincent, J.B. Systematic review of the effects of chromium (III) on chickens. Biol. Trace Elem. Res. 2019, 188, 99–126. [Google Scholar] [CrossRef]
- Saracila, M.; Panaite, T.D.; Mironeasa, S.; Untea, A.E. Dietary supplementation of some antioxidants as attenuators of heat stress on chicken meat characteristics. Agriculture 2021, 11, 638. [Google Scholar] [CrossRef]
- Lee, M.H. Official Methods of Analysis of the Association of Official Analytical Chemists, 16th ed.; AOAC International: Gaithersburg, MD, USA, 1996. [Google Scholar]
- Saracila, M.; Untea, A.E.; Panaite, T.D.; Varzaru, I.; Oancea, A.; Turcu, R.P.; Vlaicu, P.A. Creeping wood sorrel and chromium picolinate effect on the nutritional composition and lipid oxidative stability of broiler meat. Antioxidants 2022, 11, 780. [Google Scholar] [CrossRef]
- Jay, M.; Gonnet, J.F.; Wollenweber, E.; Voirin, B. Sur l’analyse qualitative des aglycones flavoniques dans une optique chimiotaxinomique. Phytochemistry 1975, 14, 1605–1612. [Google Scholar] [CrossRef]
- Varzaru, I.; Untea, A.E.; Van, I. Distribution of nutrients with benefic potential for the eyes in several medicinal plants. Rom. Biotechnol. Lett. 2015, 20, 10773–11078. [Google Scholar]
- Untea, A.E.; Criste, R.C.; Vladescu, L. Development and validation of a microwave digestion–FAAS procedure for Cu, Mn and Zn determination in liver. Rev. Chim 2012, 63, 341–346. [Google Scholar]
- Stef, D.S.; Gergen, I. Effect of mineral-enriched diet and medicinal herbs on Fe, Mn, Zn, and Cu uptake in chicken. Chem. Cent. J. 2012, 6, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Untea, A.; Lupu, A.; Saracila, M.; Panaite, T. Comparison of ABTS, DPPH, phosphomolybdenum assays for estimating antioxidant activity and phenolic compounds in five different plant extracts. Bull. UASVM Anim. Sci. Biotechnol. 2018, 75, 111–114. [Google Scholar] [CrossRef] [Green Version]
- Untea, A.E.; Varzaru, I.; Panaite, T.D.; Gavris, T.; Lupu, A.; Ropota, M. The effects of dietary inclusion of bilberry and walnut leaves in laying hens’ diets on the antioxidant properties of eggs. Animals 2020, 10, 191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bittová, M.; Krejzová, E.; Roblová, V.; Kubán, P.; Kubáň, V. Monitoring of HPLC profiles of selected polyphenolic compounds in sea buckthorn (Hippophaë rhamnoides L.) plant parts during annual growth cycle and estimation of their antioxidant potential. Cent. Eur. J. Chem. 2014, 12, 1152–1161. [Google Scholar] [CrossRef]
- Criste, A.; Urcan, A.C.; Bunea, A.; Pripon Furtuna, F.R.; Olah, N.K.; Madden, R.H.; Corcionivoschi, N. Phytochemical composition and biological activity of berries and leaves from four romanian sea buckthorn (Hippophae Rhamnoides L.) varieties. Molecules 2020, 25, 1170. [Google Scholar] [CrossRef] [Green Version]
- Pop, R.M.; Socaciu, C.; Pintea, A.; Buzoianu, A.D.; Sanders, M.G.; Gruppen, H.; Vincken, J.-P. UHPLC/PDA-ESI/MS analysis of the main berry and leaf flavonol glycosides from different Carpathian Hippophaë rhamnoides L. varieties. Phytochem. Anal. 2013, 24, 484–492. [Google Scholar] [CrossRef]
- Burčová, Z.; František, K.; Schmidt, Š.; Jablonský, M.; Ház, A.; Sládková, A.; Šurina, I. Composition of fatty acids and tocopherols in peels, seeds and leaves of Sea buckthorn. Acta Chim. Slovaca 2017, 10, 29–34. [Google Scholar] [CrossRef] [Green Version]
- Sytařová, I.; Orsavová, J.; Snopek, L.; Mlček, J.; Byczyński, Ł. Mišurcová, LImpact of phenolic compounds and vitamins C and E on antioxidant activity of sea buckthorn (Hippophaë rhamnoides L.) berries and leaves of diverse ripening times. Food Chem. 2020, 310, 125784. [Google Scholar] [CrossRef] [PubMed]
- Pop, R.M.; Weesepoel, Y.; Socaciu, C.; Pintea, A.; Vincken, J.-P.; Gruppen, H. Carotenoid composition of berries and leaves from six Romanian sea buckthorn (Hippophae rhamnoides L.) varieties. Food Chem. 2014, 147, 1–9. [Google Scholar] [CrossRef]
- Aaby, K.; Martinsen, B.K.; Borge, G.I.; Røen, D. Bioactive compounds and color of sea buckthorn (Hippophae rhamnoides L.) purees as affected by heat treatment and high-pressure homogenization. Int. J. Food Prop. 2020, 23, 651–664. [Google Scholar] [CrossRef] [Green Version]
- Jaroszewska, A.; Biel, W.; Stankowski, S.; Bośko, P. Evaluation of the influence of symbiotic mycorrhizal fungi on basic chemical compounds and minerals of sea buckthorn leaves. J. Elem. 2016, 21, 1029–1041. [Google Scholar]
- Biel, W.; Jaroszewska, A. The nutritional value of leaves of selected berry species. Sci. Agric. 2017, 74, 405–410. [Google Scholar] [CrossRef]
- Chen, X.; Zhao, W.; Liu, H.; Su, J.; Wang, P.; Zhang, Z.; Li, Y. Effect of sea buckthorn leaves on growth performance and calcium metabolism in Arbor Acres broilers. Dongbei Nongye Daxue Xuebao 2011, 42, 19–24. [Google Scholar]
- Akbari, M.; Torki, M. Effects of dietary chromium picolinate and peppermint essential oil on growth performance and blood biochemical parameters of broiler chicks reared under heat stress conditions. Int. J. Biometeorol. 2014, 58, 1383–1391. [Google Scholar] [CrossRef]
- Haq, Z.; Jain, R.K.; Khan, N.; Dar, M.Y.; Ali, S.; Gupta, M.; Varun, T.K. Recent advances in role of chromium and its antioxidant combinations in poultry nutrition: A review. Vet. World 2016, 9, 1392. [Google Scholar] [CrossRef] [Green Version]
- Qin, X.; Zhang, T.; Cao, Y.; Deng, B.; Zhang, J.; Zhao, J. Effects of dietary sea buckthorn pomace supplementation on skeletal muscle mass and meat quality in lambs. Meat Sci. 2020, 166, 108141. [Google Scholar] [CrossRef]
- Kwon, E.-Y.; Lee, J.; Kim, Y.J.; Do, A.; Choi, J.-Y.; Cho, S.-J.; Jung, U.J.; Lee, M.-K.; Park, Y.B.; Choi, M.-S. Sea buckthorn leaves extract and flavonoid glycosides extract from sea buckthorn leaves ameliorates adiposity, hepatic steatosis, insulin resistance, and inflammation in diet-induced obesity. Nutrients 2017, 9, 569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pathak, G.P.; Sharma, N.; Mane, B.G.; Sharma, D.; Krofa, D.; Khurana, S.K. Effect of Sea buckthorn (Hippophae rhamnoides)-leaves, pulp and oil on growth performance, carcass characteristics and meat quality of broilers chicken. J. Poult. Sci. Technol. 2015, 3, 20–23. [Google Scholar]
- Suksombat, W.; Kanchanatawee, S. Effects of various sources and levels of chromium on performance of broilers. Asian-Australas. J. Anim. Sci. 2005, 18, 1628–1633. [Google Scholar] [CrossRef]
- Javed, M.T.; Ellahi, M.; Abbas, N.; Yasmin, R.; Mazhar, M. Effects of dietary chromium chloride, nicotinic acid and copper sulphate on meat of broilers. Br. Poult. Sci. 2010, 51, 354–360. [Google Scholar] [CrossRef] [PubMed]
- Król, B.; Słupczyńska, M.; Kinal, S.; Bodarski, R.; Tronina, W.; Mońka, M. Bioavailability of organic and inorganic sources of chromium in broiler chicken feeds. J. Elem. 2017, 22, 283–294. [Google Scholar]
- Białek, A.; Białek, M.; Lepionka, T.; Kaszperuk, K.; Banaszkiewicz, T.; Tokarz, A. The effect of pomegranate seed oil and grapeseed oil on cis-9, trans-11 CLA (rumenic acid), n-3 and n-6 fatty acids deposition in selected tissues of chickens. J. Anim. Physiol. Anim. Nutr. 2018, 102, 962–976. [Google Scholar] [CrossRef]
- Elkin, R.G.; Ying, Y.; Fan, Y.; Harvatine, K.J. Influence of feeding stearidonic acid (18:4n-3)-enriched soybean oil, as compared to conventional soybean oil, on tissue deposition of very long-chain omega-3 fatty acids in meat-type chickens. Anim. Feed. Sci. Technol. 2016, 217, 1–12. [Google Scholar] [CrossRef]
- Foglia, T.A.; Cartwright, A.L.; Gyurik, R.J.; Philips, J.G. Fatty acid turnover rates in the adipose tissues of the growing chicken (Gallus domesticus). Lipids 1994, 29, 497–502. [Google Scholar] [CrossRef]
- Mancinelli Cartoni, A.; Mattioli, S.; Twining, C.; Dal Bosco, A.; Donoghue, A.M.; Arsi, K.; Angelucci, E.; Chiattelli, D.; Castellini, C. Poultry meat and eggs as an alternative source of n-3 long-chain polyunsaturated fatty acids for human nutrition. Nutrients 2022, 14, 1969. [Google Scholar] [CrossRef]
- Czauderna, M.; Białek, M.; Białek, A.; Karpińska, M. Diet supplemented with lycopene and selenized yeast change contents of fatty acids in the liver and femoral muscles of rabbits. Livest. Sci. 2021, 250, 104598. [Google Scholar] [CrossRef]
- Lauritzen, L.; Brambilla, P.; Mazzocchi, A.; Harsløf, L.B.; Ciappolino, V.; Agostoni, C. DHA Effects in brain development and function. Nutrients 2016, 8, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Pora, B.L.R.; Dong, K.; Hasjim, J. Health benefits of docosahexaenoic acid and its bioavailability: A review. Food Sci. Nutr. 2021, 9, 5229–5243. [Google Scholar] [CrossRef] [PubMed]
- Moran, C.; Keegan, J.; Vienola, K.; Apajalahti, J. Broiler tissue enrichment with docosahexaenoic acid (DHA) through dietary supplementation with Aurantiochytrium limacinum algae. Food Nutr. Sci. 2018, 9, 1160–1173. [Google Scholar]
- Food and Agriculture Organization of the United Nations. FAO Fats and Fatty Acids in Human Nutrition; Report of an Expert Consultation; Food and Agriculture Organization of the United Nations: Rome, Italy, 2010. [Google Scholar]
- Dal Bosco, A.; Cartoni Mancinelli, A.; Vaudo, G.; Cavallo, M.; Castellini, C.; Mattioli, S. Indexing of fatty acids in poultry meat for its characterization in healthy human nutrition: A comprehensive application of the scientific literature and new proposals. Nutrients 2022, 14, 3110. [Google Scholar] [CrossRef] [PubMed]
- Tan, Z.; Halter, B.; Liu, D.; Gilbert, E.R.; Cline, M.A. Dietary flavonoids as modulators of lipid metabolism in poultry. Front. Physiol. 2022, 13, 863860. [Google Scholar] [CrossRef]
- Lee, J.; Jung, E.; Lee, J.; Kim, S.; Huh, S.; Kim, Y.; Kim, Y.; Byun, S.Y.; Kim, Y.S.; Park, D. Isorhamnetin represses adipogenesis in 3T3-L1 cells. Obesity 2009, 17, 226–232. [Google Scholar] [CrossRef]
- Yan, S.X.; Li, X.; Sun, C.D.; Chen, K.S. Hypoglycemic and hypolipidemic effects of quercetin and its glycosides. Zhongguo Zhong Yao Za Zhi/China J. Chin. Mater. Med. 2015, 40, 4560–4567. [Google Scholar]
- Khan, R.U.; Naz, S.; Javadani, M.; Nikousefat, Z.; Selvaggi, M.; Tufarelli, V.; Laudadio, V. The use of turmeric (Curcuma longa) in poultry diets. World Poult. Sci. J. 2012, 68, 97–103. [Google Scholar] [CrossRef]
- Khan, R.U.; Nikosefat, Z.; Tufarelli, V.; Naz, S.; Javdani, M.; Laudadio, V. Garlic (Allium sativa) supplementation in poultry diet: Effect on production and physiology. World Poult. Sci. J. 2012, 68, 417–424. [Google Scholar] [CrossRef]
- Chand, N.; Naz, S.; Irfan, M.; Khan, R.U.; Ur Rehman, Z. Effect of sea buckthorn (Hippophae rhamnoides L.) seed supplementation on egg quality and cholesterol of Rhode Island Red×Fayoumi laying hens. Korean J. Food Sci. Anim. Resour. 2018, 38, 468–475. [Google Scholar]
- Laudadio, V.; Tufarelli, V. Influence of substituting dietary soybean meal for dehulled-micronized lupin (Lupinus albus cv. Multitalia) on early phase laying hens production and egg quality. Livest Sci. 2010, 140, 184–188. [Google Scholar] [CrossRef]
- Attia, Y.A.; Al-Harthi, M.A.; Korish, M.A.; Shiboob, M.M. Fatty acid and cholesterol profiles and hypocholesterolemic, atherogenic, and thrombogenic indices of table eggs in the retail market. Lipids Health Dis. 2015, 14, 136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Czaplicki, S.; Ogrodowska, D.; Zadernowski, R. Effect of sea-buckthorn (Hippophaë rhamnoides L.) pulp oil consumption on fatty acids and vitamin A and E accumulation in adipose tissue and liver of rats. Plant Foods Hum. Nutr. 2017, 72, 198–204. [Google Scholar] [CrossRef] [PubMed]
- Untea, A.E.; Varzaru, I.; Turcu, R.P.; Panaite, T.D.; Saracila, M. The use of dietary chromium associated with vitamins and minerals (synthetic and natural source) to improve some quality aspects of broiler thigh meat reared under heat stress condition. Ital. J. Anim. Sci. 2021, 20, 1491–1499. [Google Scholar] [CrossRef]
- Mäkinen, S.; Hellström, J.; Mäki, M.; Korpinen, R.; Mattila, P.H. Bilberry and sea buckthorn leaves and their subcritical water extracts prevent lipid oxidation in meat products. Foods 2020, 9, 265. [Google Scholar] [CrossRef] [Green Version]
- Papuc, C.; Goran, G.V.; Predescu, C.N.; Nicorescu, V.; Stefan, G. Plant polyphenols as antioxidant and antibacterial agents for shelf-life extension of meat and meat products: Classification, structures, sources, and action mechanisms. Compr. Rev. Food Sci. Food Saf. 2017, 16, 1243–1268. [Google Scholar] [CrossRef] [Green Version]
- Saracila, M.; Panaite, T.D.; Papuc, C.P.; Criste, R.D. Heat stress in broiler chickens and the effect of dietary polyphenols, with special reference to Willow (Salix spp.) bark supplements—A review. Antioxidants 2021, 10, 686. [Google Scholar] [CrossRef]
- Xiong, Y.L.; Mikel, W.B. Meat Science and Applications; Wai-Kit, N., Rogers, R., Eds.; Marcel Dekker: New York, NY, USA, 2001; pp. 351–369. [Google Scholar]
- Erickson, M.C. Food Lipids–Chemistry, Nutrition, and Biotechnology, 2nd ed.; Akoh, C.C., Min, D.B., Eds.; Marcel Dekker: New York, NY, USA, 2002; pp. 365–411. [Google Scholar]
- Wagh, R.V.; Chatli, M.K. Response surface optimization of extraction protocols to obtain phenolic rich antioxidant from sea buckthorn and their potential application into model meat system. J. Food Sci. Technol. 2017, 54, 1565–1576. [Google Scholar] [CrossRef] [Green Version]
- Surai, P.F. Polyphenol compounds in the chicken/animal diet: From the past to the future. J. Anim. Physiol. Anim. Nutr. 2014, 98, 19–31. [Google Scholar] [CrossRef]
- Jerónimo, E.; Soldado, D.; Sengo, S.; Francisco, A.; Fernandes, F.; Portugal, A.P.; Bessa, R.J. Increasing the α-tocopherol content and lipid oxidative stability of meat through dietary Cistus ladanifer L. in lamb fed increasing levels of polyunsaturated fatty acid rich vegetable oils. Meat Sci. 2020, 164, 108092. [Google Scholar] [CrossRef]
- Serra, V.; Salvatori, G.; Pastorelli, G. Dietary polyphenol supplementation in food producing animals: Effects on the quality of derived products. Animals 2021, 11, 401. [Google Scholar] [CrossRef] [PubMed]
- Reboul, E.; Thap, S.; Perrot, E. Effect of the main dietary antioxidants (carotenoids, γ-tocopherol, polyphenols, and vitamin C) on α-tocopherol absorption. Eur. J. Clin. Nutr. 2007, 61, 1167–1173. [Google Scholar] [CrossRef] [PubMed]
- Samtiya, M.; Aluko, R.E.; Dhewa, T. Plant food anti-nutritional factors and their reduction strategies: An overview. Food Prod. Process. Nutr. 2020, 2, 6. [Google Scholar] [CrossRef] [Green Version]
- Antoine, T.; Georgé, S.; Leca, A.; Desmarchelier, C.; Halimi, C.; Gervais, S.; Aupy, F.; Marconot, G.; Reboul, E. Reduction of pulse “antinutritional” content by optimizing pulse canning process is insufficient to improve fat-soluble vitamin bioavailability. Food Chem. 2022, 15, 131021. [Google Scholar] [CrossRef] [PubMed]
- Timotheo, C.A.; Lauer, C.M. Toxicity of vegetable tannin extract from Acacia mearnsii in Saccharomyces cerevisiae. Int. J. Environ. Sci. Technol. 2018, 15, 659–664. [Google Scholar] [CrossRef]
- Chen, X.; Li, H.; Zhang, B.; Deng, Z. The synergistic and antagonistic antioxidant interactions of dietary phytochemical combinations. Crit. Rev. Food Sci. Nutr. 2021, 62, 5658–5677. [Google Scholar] [CrossRef]





| Ingredient | Grower (14–28 d) | Finisher (28–42 d) | ||||
|---|---|---|---|---|---|---|
| T0 | T1 | T2 | T0 | T1 | T2 | |
| % | ||||||
| Corn | 40.18 | 40.18 | 38.36 | 44.70 | 44.70 | 42.91 |
| Soybean meal | 26.33 | 26.33 | 26.07 | 21.32 | 21.32 | 21.06 |
| Wheat | 20.00 | 20.00 | 20.00 | 20.00 | 20.00 | 20.00 |
| Corn gluten | 5.00 | 5.00 | 5.00 | 5.00 | 5.00 | 5.00 |
| Sea buckthorn leaves (SBL) | - | - | 2.00 | - | - | 2.00 |
| Oil | 3.78 | 3.78 | 3.83 | 4.62 | 4.62 | 4.66 |
| Monocalcium phosphate | 1.36 | 1.36 | 1.36 | 1.19 | 1.19 | 1.19 |
| Calcium carbonate | 1.25 | 1.25 | 1.23 | 1.13 | 1.13 | 1.11 |
| Salt | 0.36 | 0.36 | 0.36 | 0.36 | 0.36 | 0.36 |
| Methionine | 0.30 | 0.30 | 0.31 | 0.26 | 0.26 | 0.27 |
| Lysine | 0.30 | 0.30 | 0.32 | 0.30 | 0.30 | 0.31 |
| Threonine | 0.09 | 0.09 | 0.11 | 0.07 | 0.07 | 0.08 |
| Choline | 0.05 | 0.05 | 0.05 | 0.05 | 0.05 | 0.05 |
| Chromium | - | 0.00002 | 0.00002 | - | 0.00002 | 0.00002 |
| A1 Premix 1 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 |
| Total | 100 | 100 | 100 | 100 | 100 | 100 |
| Chemical analysis- theoretical | ||||||
| ME, Kcal/kg | 3128.99 | 3217.72 | ||||
| CP, % | 21.50 | 20.00 | ||||
| EE, % | 6.01 | 6.49 | ||||
| CF, % | 3.57 | 3.36 | ||||
| Ca., % | 0.87 | 0.81 | ||||
| P, % | 0.70 | 0.65 | ||||
| Available phosphorus, % | 0.43 | 0.41 | ||||
| Analyzed Parameters | Sea Buckthorn Leaves (SBL) |
|---|---|
| Proximate composition (%) | |
| Dry matter | 91.63 |
| Crude protein | 14.48 |
| Ether extractives | 5.12 |
| Crude fiber | 13.68 |
| Ash | 6.37 |
| Fatty acids (g/100 g FAME) | |
| Σ Saturated fatty acids (SFA) | 30.76 |
| Σ Monounsaturated fatty acids (MUFA) | 32.66 |
| Σ Polyunsaturated fatty acids (PUFA) | 35.65 |
| Σ Unsaturated fatty acids (UFA) | 68.31 |
| SFA/UFA | 0.45 |
| PUFA/MUFA | 1.09 |
| Σ n-3 of which: | 24.92 |
| α-linolenic acid | 23.61 |
| Eicosadienoic acid | 0.47 |
| Σ n-6 of which: | 10.72 |
| Arachidonic acid | 0.69 |
| Σ n-6/ Σ n-3 | 0.43 |
| Antioxidant profile | |
| TPC, mg/g GAE | 58.61 |
| TFC, mg/g QE | 9.03 |
| Lutein and zeaxanthin, µg/g | 583.4 |
| Vitamin E, µg/g | 321.29 |
| TAC, µM Trolox | 1147.91 |
| Mineral profile (mg/kg) | |
| Copper | 3.05 |
| Iron | 334.79 |
| Manganese | 159.59 |
| Zinc | 126.78 |
| Item | Treatment | SEM | p-Value | ||
|---|---|---|---|---|---|
| T0 | T1 | T2 | |||
| Broiler performance | |||||
| Initial BW, g | 521.65 | 521.47 | 521.65 | 3.59 | 0.9994 |
| Final BW, g | 3246.74 | 3244.06 | 3260.00 | 2.68 | 0.9542 |
| ADWG, g/broiler/day | 97.84 | 97.24 | 97.80 | 0.746 | 0.7554 |
| ADFI, g/broiler/day | 158.68 | 149.33 | 157.73 | 4.688 | 0.6747 |
| FCR, g feed/g gain | 1.61 | 1.54 | 1.62 | 0.044 | 0.7108 |
| Carcass and cuts yield | |||||
| Carcass yield, % | 84.79 | 81.80 | 81.17 | 0.891 | 0.2151 |
| Breast yield, % | 26.68 | 27.67 | 26.87 | 0.424 | 0.6446 |
| Thigh yield, % | 21.23 | 22.01 | 21.73 | 0.264 | 0.5118 |
| Gizzard, % | 1.16 | 1.14 | 1.12 | 0.046 | 0.9587 |
| Liver, % | 2.44 | 2.34 | 2.59 | 0.069 | 0.3793 |
| Heart, % | 0.57 | 0.54 | 0.54 | 0.018 | 0.8013 |
| Spleen, % | 0.13 | 0.12 | 0.13 | 0.007 | 0.8766 |
| Bile, % | 0.08 | 0.10 | 0.07 | 0.008 | 0.2576 |
| Item | Treatment | SEM | p-Value | ||
|---|---|---|---|---|---|
| T0 | T1 | T2 | |||
| Breast | |||||
| Dry matter | 25.32 | 25.17 | 25.99 | 0.6549 | 0.561 |
| Crude Protein | 22.78 | 23.12 | 23.39 | 0.5394 | 0.530 |
| Ether extractives | 1.46 | 1.50 | 1.49 | 0.0024 | 0.061 |
| Ash | 1.04 | 1.06 | 1.10 | 0.0015 | 0.188 |
| Thigh | |||||
| Dry matter | 23.47 | 23.89 | 24.47 | 0.8651 | 0.346 |
| Crude Protein | 18.84 | 19.21 | 18.89 | 0.5478 | 0.762 |
| Ether extractives | 3.68 b | 3.70 b | 4.45 a | 0.0197 | 0.0001 |
| Ash | 0.85 | 0.88 | 0.91 | 0.0025 | 0.288 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saracila, M.; Untea, A.E.; Panaite, T.D.; Varzaru, I.; Oancea, A.-G.; Turcu, R.P.; Vlaicu, P.A. Effects of Supplementing Sea Buckthorn Leaves (Hippophae rhamnoides L.) and Chromium (III) in Broiler Diet on the Nutritional Quality and Lipid Oxidative Stability of Meat. Antioxidants 2022, 11, 2220. https://doi.org/10.3390/antiox11112220
Saracila M, Untea AE, Panaite TD, Varzaru I, Oancea A-G, Turcu RP, Vlaicu PA. Effects of Supplementing Sea Buckthorn Leaves (Hippophae rhamnoides L.) and Chromium (III) in Broiler Diet on the Nutritional Quality and Lipid Oxidative Stability of Meat. Antioxidants. 2022; 11(11):2220. https://doi.org/10.3390/antiox11112220
Chicago/Turabian StyleSaracila, Mihaela, Arabela Elena Untea, Tatiana Dumitra Panaite, Iulia Varzaru, Alexandra-Gabriela Oancea, Raluca Paula Turcu, and Petru Alexandru Vlaicu. 2022. "Effects of Supplementing Sea Buckthorn Leaves (Hippophae rhamnoides L.) and Chromium (III) in Broiler Diet on the Nutritional Quality and Lipid Oxidative Stability of Meat" Antioxidants 11, no. 11: 2220. https://doi.org/10.3390/antiox11112220