
Identification of Phytogenic Compounds with Antioxidant Action That Protect Porcine Intestinal Epithelial Cells from Hydrogen Peroxide Induced Oxidative Damage
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Traditional Chinese Herbal Compounds
2.2. Cell Culture
2.3. Oxidative Stress Model Establishment
2.4. Drug Toxicity Assay
2.5. HTS Assay
2.6. Secondary Screening and Validation of the Hit Compounds
2.7. Determination of Malondialdehyde (MDA) Accumulation and ROS Generation
2.8. Detection of Apoptosis and the Mitochondrial Membrane Potential (MMP)
2.9. Determination of Antioxidant Activities
2.10. Total RNA Isolation and Quantitative Reverse Transcription (RT-q) PCR
2.11. Western Blot Analysis
2.12. Establishment of Nrf2-Knockdown IPEC-J2 Cells
2.13. Statistical Analysis
3. Results
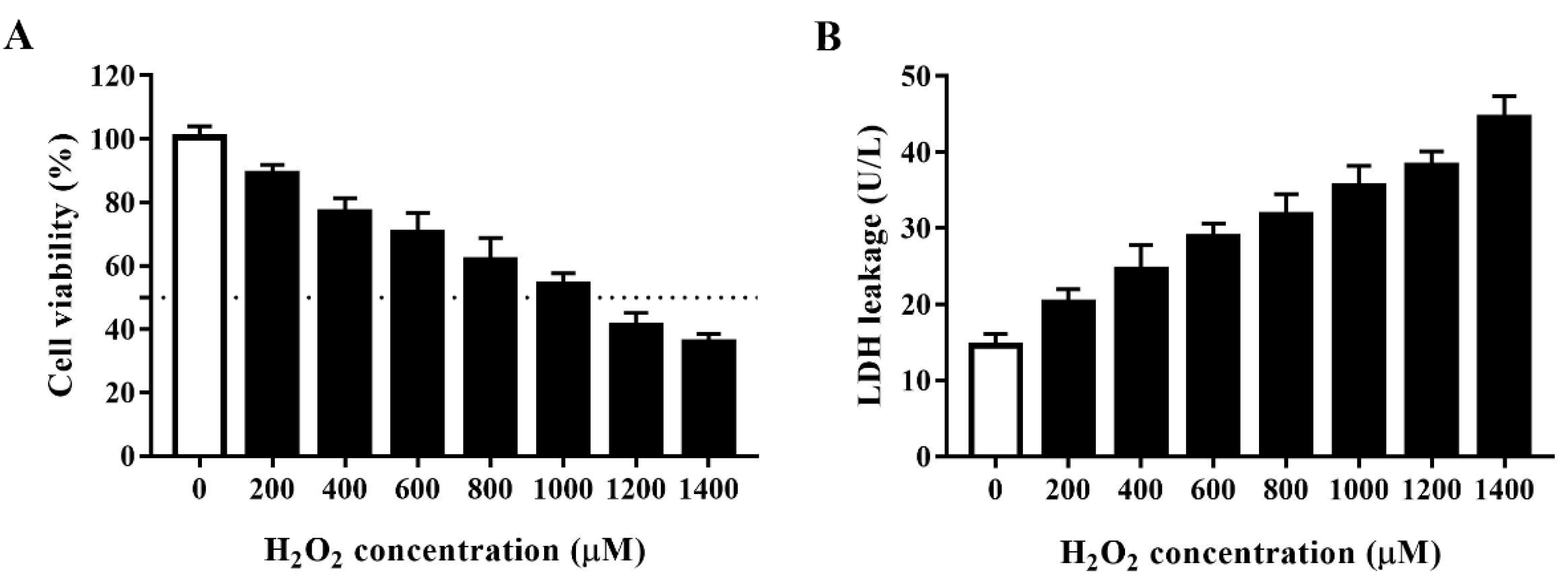
3.1. Establishment of an Oxidative Stress Model for HTS
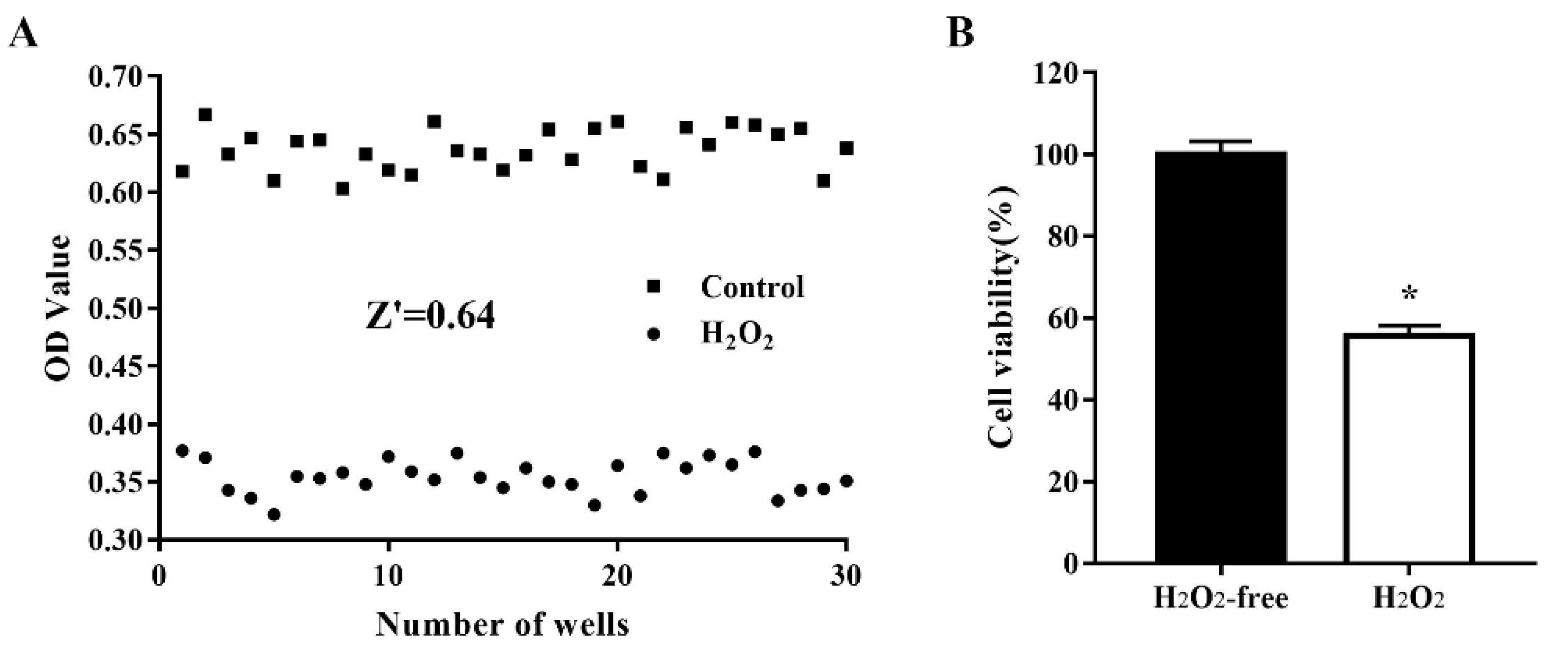
3.2. Variability and Robustness of the Model
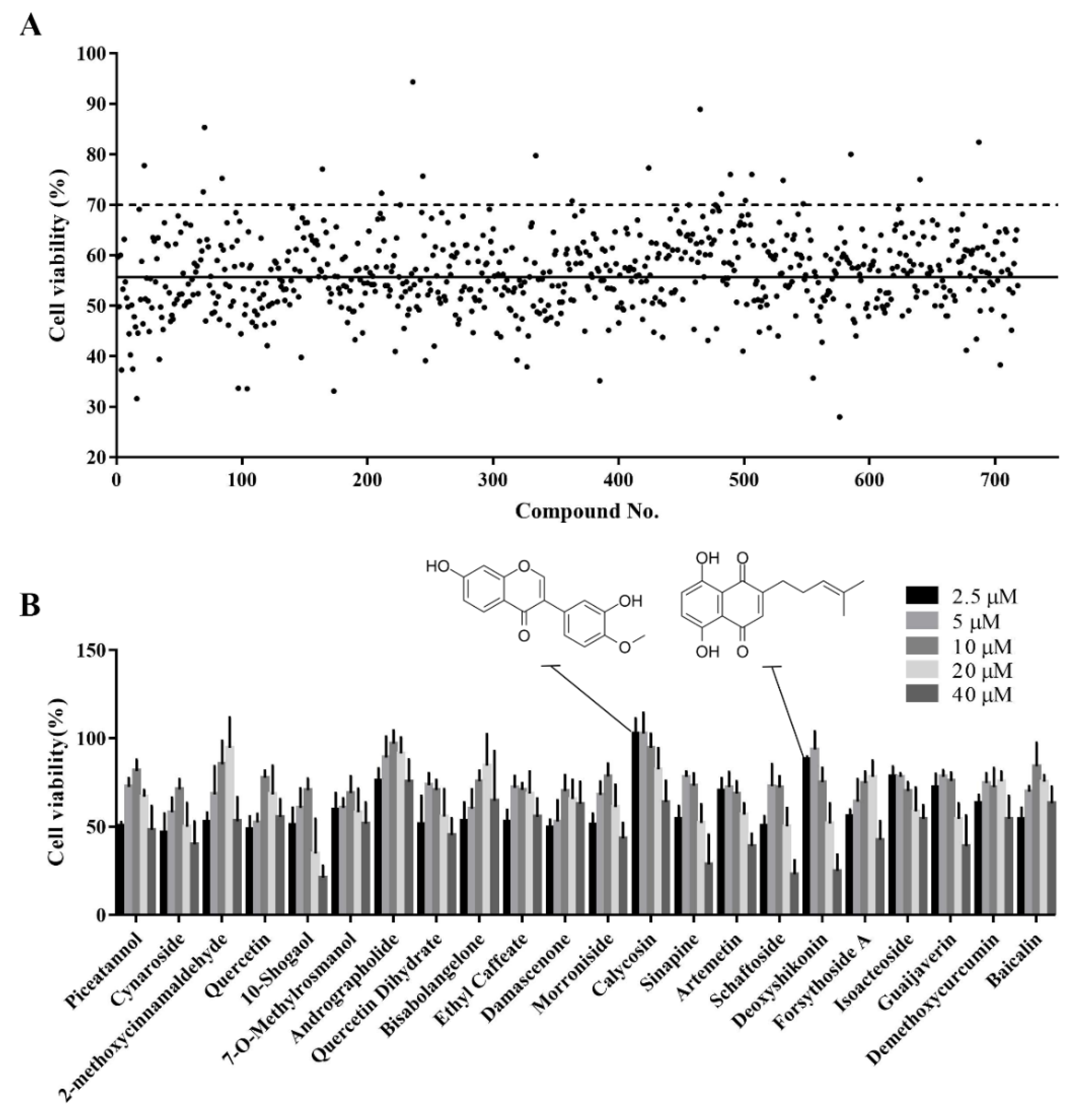
3.3. Identification of Active Monomers in a Chinese Herbal Compound Library
3.4. Secondary Screening of the Hit Compounds
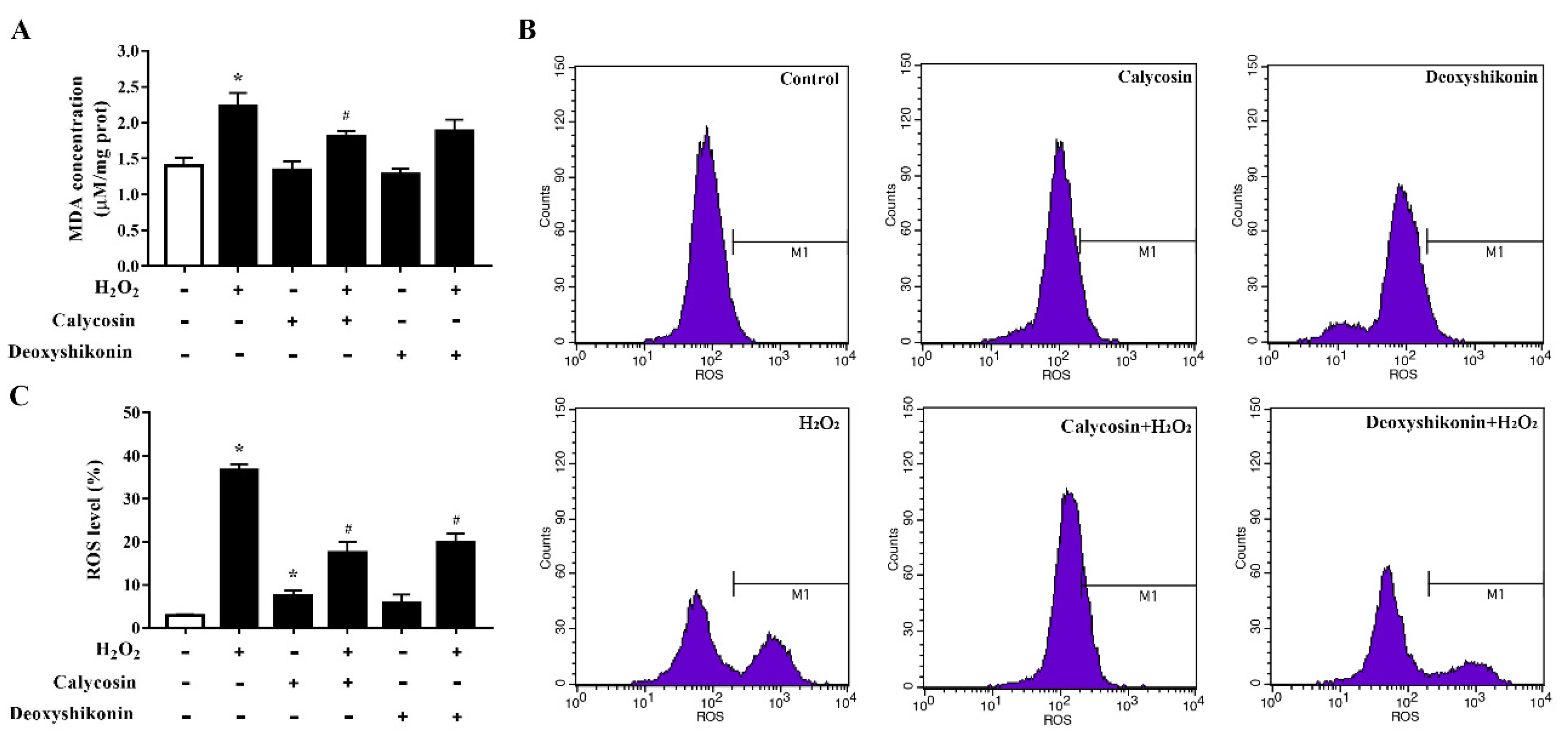
3.5. Characterization of the Antioxidant Activities of Calycosin and Deoxyshikonin
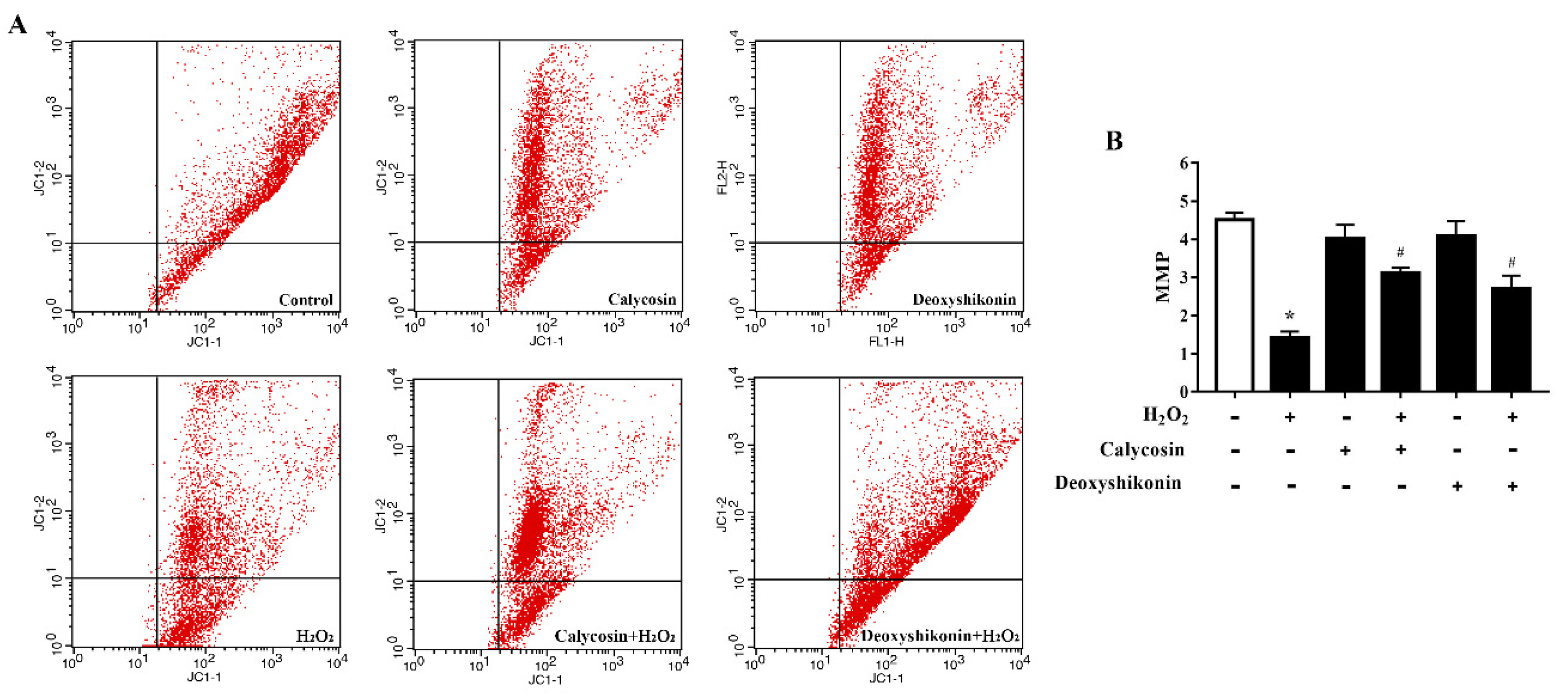
3.6. Effects of Calycosin and Deoxyshikonin on the MMP
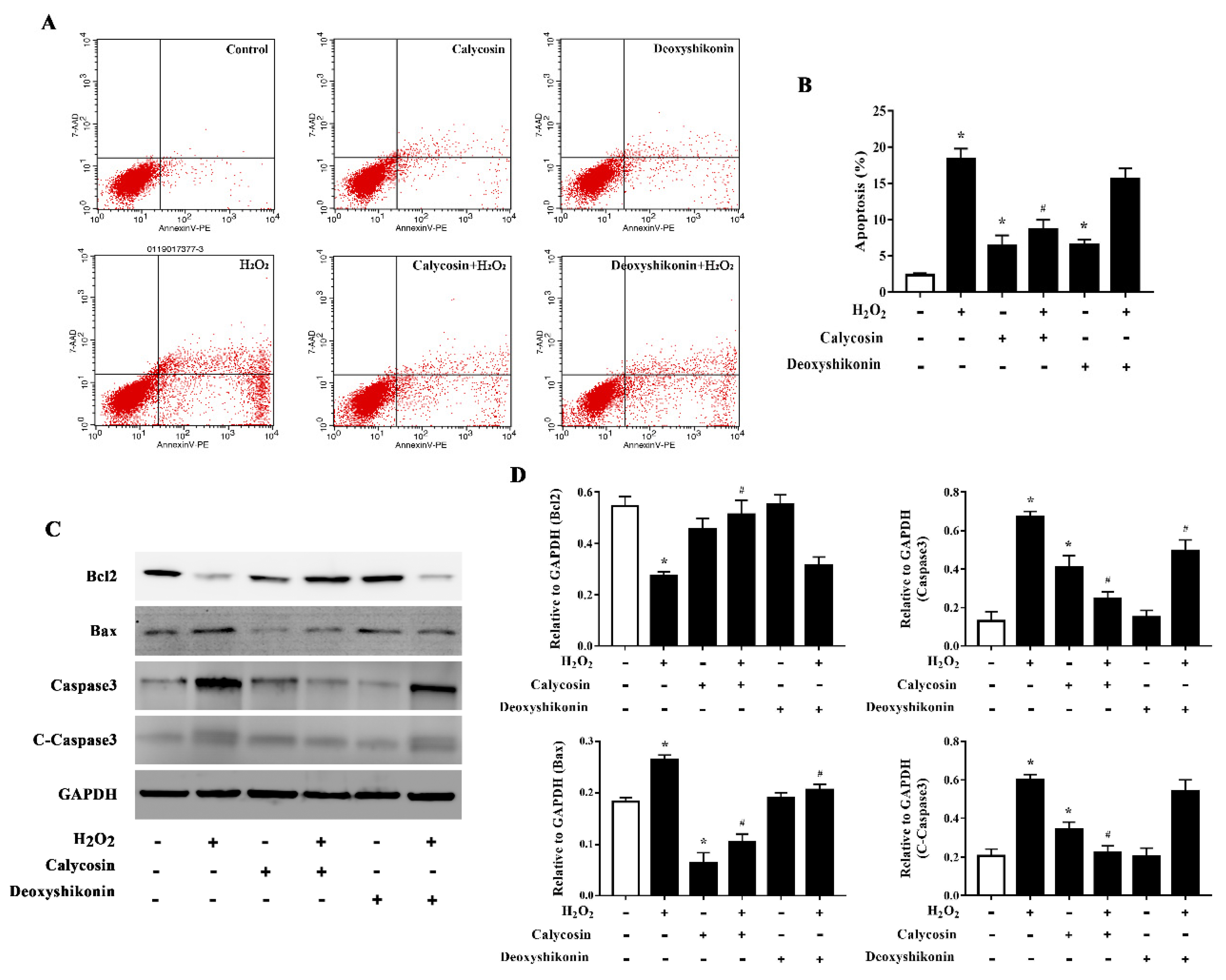
3.7. Effects of Calycosin and Deoxyshikonin on the Apoptosis
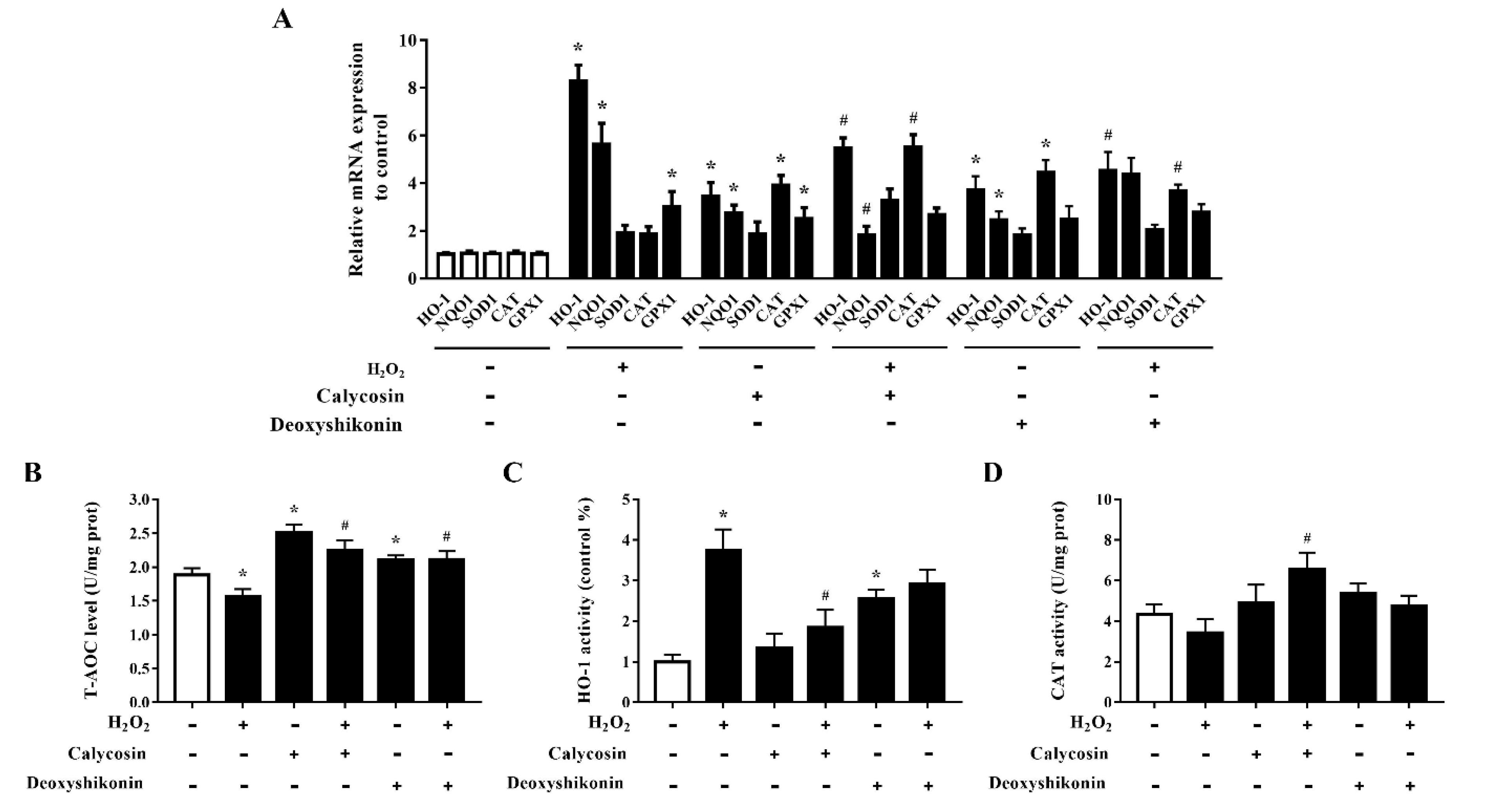
3.8. Effects of Calycosin and Deoxyshikonin on the Antioxidant Defense System
3.9. Activation of the Nrf2 Signaling Pathway
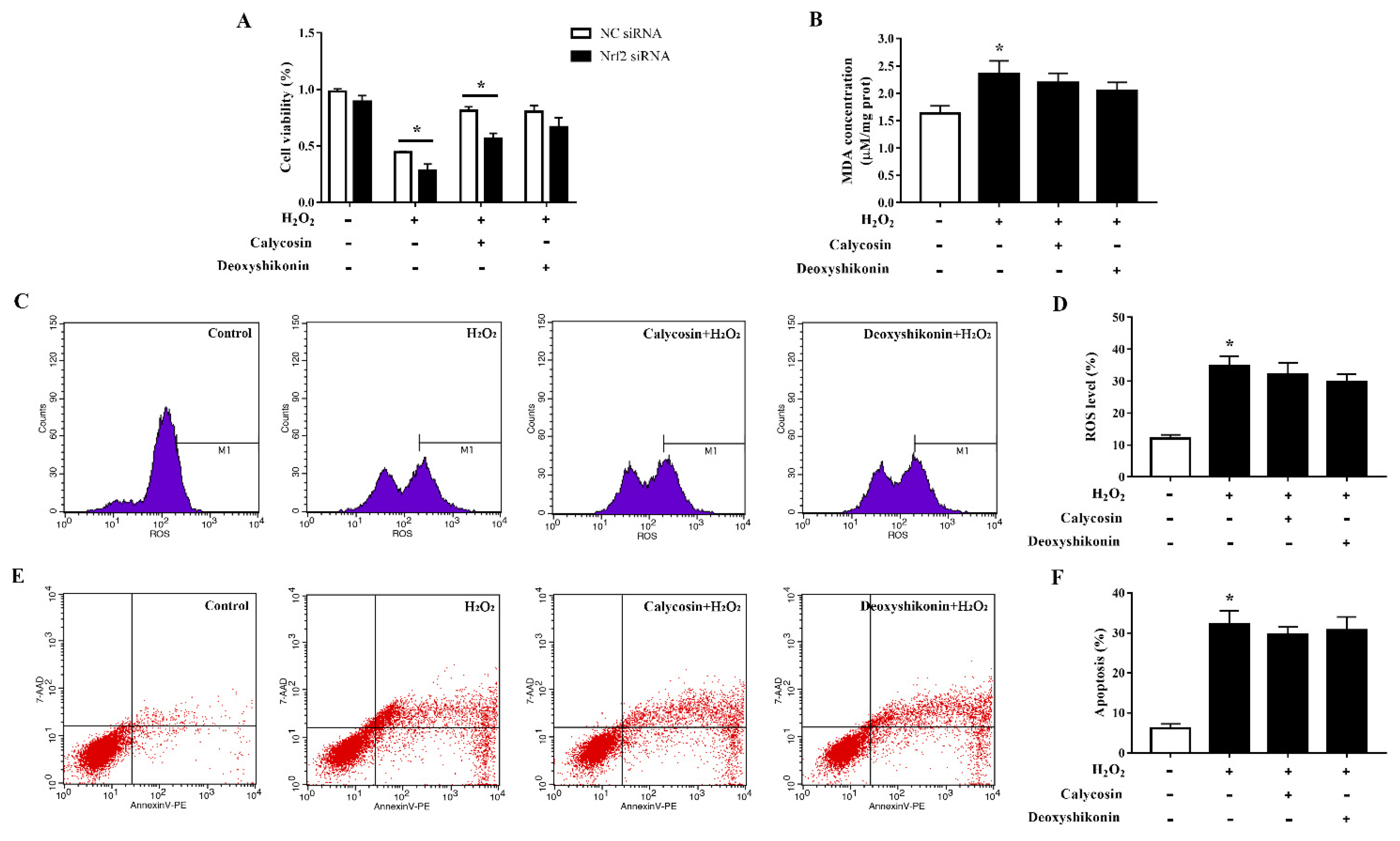
3.10. Effects of Calycosin and Deoxyshikonin on IPEC-J2 Cells Treated with Nrf2 siRNA
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Schieber, M.; Chandel, N.S. ROS function in redox signaling and oxidative stress. Curr. Biol. 2014, 24, R453–R462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manuelian, C.L.; Pitino, R.; Simoni, M.; Mavrommatis, A.; de Marchi, M.; Righi, F.; Tsiplakou, E. Plant feed additives as natural alternatives to the use of synthetic antioxidant vitamins on livestock mammals’ performances, health, and oxidative status: A review of the literature in the last 20 years. Antioxidants 2021, 10, 1461. [Google Scholar] [CrossRef] [PubMed]
- Righi, F.; Pitino, R.; Manuelian, C.L.; Simoni, M.; Quarantelli, A.; de Marchi, M.; Tsiplakou, E. Plant feed additives as natural alternatives to the use of synthetic antioxidant vitamins on poultry performances, health, and oxidative status: A review of the literature in the last 20 years. Antioxidants 2021, 10, 659. [Google Scholar] [CrossRef]
- Kuralkar, P.; Kuralkar, S.V. Role of herbal products in animal production—An updated review. J. Ethnopharmacol. 2021, 278, 114246. [Google Scholar] [CrossRef] [PubMed]
- Lillehoj, H.; Liu, Y.; Calsamiglia, S.; Fernandez-Miyakawa, M.E.; Chi, F.; Cravens, R.L.; Oh, S.; Gay, C.G. Phytochemicals as antibiotic alternatives to promote growth and enhance host health. Vet. Res. 2018, 49, 76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rossi, B.; Toschi, A.; Piva, A.; Grilli, E. Single components of botanicals and nature-identical compounds as a non-antibiotic strategy to ameliorate health status and improve performance in poultry and pigs. Nutr. Res. Rev. 2020, 33, 218–234. [Google Scholar] [CrossRef]
- Brewer, M.S. Natural antioxidants: Sources, compounds, mechanisms of action, and potential applications. Compr. Rev. Food Sci. Food Saf. 2011, 10, 221–247. [Google Scholar] [CrossRef]
- Bonilla, J.; Sobral, P.J.d.A. Antioxidant and antimicrobial properties of ethanolic extracts of guarana, boldo, rosemary and cinnamon. Braz. J. Food Technol. 2017, 20, e2016024. [Google Scholar] [CrossRef] [Green Version]
- Mekinić, I.G.; Skroza, D.; Ljubenkov, I.; Katalinić, V.; Šimat, V. Antioxidant and antimicrobial potential of phenolic metabolites from traditionally used mediterranean herbs and spices. Foods 2019, 8, 579. [Google Scholar] [CrossRef] [Green Version]
- Mahfuz, S.; Shang, Q.; Piao, X. Phenolic compounds as natural feed additives in poultry and swine diets: A review. J. Anim. Sci. Biotechnol. 2021, 12, 48. [Google Scholar] [CrossRef]
- De Castilho, T.S.; Matias, T.B.; Nicolini, K.P.; Nicolini, J. Study of interaction between metal ions and quercetin. Food Sci. Hum. Well. 2018, 7, 215–219. [Google Scholar] [CrossRef]
- Zhuang, Y.; Wu, H.; Wang, X.; He, J.; He, S.; Yin, Y. Resveratrol attenuates oxidative stress-induced intestinal barrier injury through PI3K/Akt-mediated Nrf2 signaling pathway. Oxid. Med. Cell. Longev. 2019, 2019, 7591840. [Google Scholar] [CrossRef] [Green Version]
- Jia, P.; Ji, S.; Zhang, H.; Chen, Y.; Wang, T. Piceatannol ameliorates hepatic oxidative damage and mitochondrial dysfunction of weaned piglets challenged with diquat. Animals 2020, 10, 1239. [Google Scholar] [CrossRef] [PubMed]
- Vallejo, M.J.; Salazar, L.; Grijalva, M. Oxidative stress modulation and ROS-mediated toxicity in cancer: A review on in vitro models for plant-derived compounds. Oxid. Med. Cell. Longev. 2017, 2017, 4586068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, J.M.; Xu, Z.L.; Li, N.; Zhao, Y.W.; Wang, Z.Z.; Xiao, W. Identification of cardioprotective agents from traditional Chinese medicine against oxidative damage. Mol. Med. Rep. 2016, 14, 77–88. [Google Scholar] [CrossRef] [Green Version]
- Ayuso, M.; van Cruchten, S.; van Ginneken, C. Medium-throughput system for in vitro oxidative stress assessment in IPEC-J2 cells. Int. J. Mol. Sci. 2020, 21, 7263. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.H.; Chung, T.D.; Oldenburg, K.R. A simple statistical parameter for use in evaluation and validation of high throughput screening assays. J. Biomol. Screen. 1999, 4, 67–73. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔC T method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Lyu, W.; Zhang, W.; Chen, Y.; Luo, F.; Wang, Y.; Ji, H.; Zhang, G. Discovery of natural products capable of inducing porcine host defense peptide gene expression using cell-based high throughput screening. J. Anim. Sci. Biotechnol. 2021, 12, 14. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, W.; Chu, X.; Wang, S.; Wang, Y.; Ji, H. Deoxyshikonin-induced gene expression profile in porcine epithelial cells. Front. Vet. Sci. 2022, 8, 711721. [Google Scholar] [CrossRef]
- Vergauwen, H.; Tambuyzer, B.; Jennes, K.; Degroote, J.; Wang, W.; de Smet, S.; Michiels, J.; van Ginneken, C. Trolox and ascorbic acid reduce direct and indirect oxidative stress in the IPEC-J2 cells, an in vitro model for the porcine gastrointestinal tract. PLoS ONE 2015, 10, e0120485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wan, H.; Liu, D.; Yu, X.; Sun, H.; Li, Y. A caco-2 cell-based quantitative antioxidant activity assay for antioxidants. Food Chem. 2015, 175, 601–608. [Google Scholar] [CrossRef] [PubMed]
- Vergauwen, H.; Prims, S.; Degroote, J.; Wang, W.; Casteleyn, C.; van Cruchten, S.; de Smet, S.; Michiels, J.; van Ginneken, C. In vitro investigation of six antioxidants for pig diets. Antioxidants 2016, 5, 41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Halliwell, B. Cell culture, oxidative stress, and antioxidants: Avoiding pitfalls. Biomed. J. 2014, 37, 99–105. [Google Scholar] [CrossRef] [PubMed]
- Foss, K.; Przybyłowicz, K.E.; Sawicki, T. Antioxidant activity and profile of phenolic compounds in selected herbal plants. Plant Foods Hum. Nutr. 2022, 77, 383–389. [Google Scholar] [CrossRef] [PubMed]
- Shen, N.; Wang, T.; Gan, Q.; Liu, S.; Wang, L.; Jin, B. Plant flavonoids: Classification, distribution, biosynthesis, and antioxidant activity. Food Chem. 2022, 383, 132531. [Google Scholar] [CrossRef]
- Park, C.; Cha, H.J.; Lee, H.; Kim, G.Y.; Choi, Y.H. The regulation of the TLR4/NF-κB and Nrf2/HO-1 signaling pathways is involved in the inhibition of lipopolysaccharide-induced inflammation and oxidative reactions by morroniside in RAW 264.7 macrophages. Arch. Biochem. Biophys. 2021, 706, 108926. [Google Scholar] [CrossRef] [PubMed]
- Lu, C.Y.; Day, C.H.; Kuo, C.H.; Wang, T.F.; Ho, T.J.; Lai, P.F.; Chen, R.J.; Yao, C.H.; Viswanadha, V.P.; Kuo, W.W.; et al. Calycosin alleviates H2O2-induced astrocyte injury by restricting oxidative stress through the Akt/Nrf2/HO-1 signaling pathway. Environ. Toxicol. 2022, 37, 858–867. [Google Scholar] [CrossRef] [PubMed]
- Yates, K.; Pohl, F.; Busch, M.; Mozer, A.; Watters, L.; Shiryaev, A.; Lin, P.K.T. Determination of sinapine in rapeseed pomace extract: Its antioxidant and acetylcholinesterase inhibition properties. Food Chem. 2019, 276, 768–775. [Google Scholar] [CrossRef]
- Paudel, K.R.; Kim, D.-W. Microparticles-mediated vascular inflammation and its amelioration by antioxidant activity of baicalin. Antioxidants 2020, 9, 890. [Google Scholar] [CrossRef]
- Gong, L.; Wang, C.; Zhou, H.; Ma, C.; Zhang, Y.; Peng, C.; Li, Y. A review of pharmacological and pharmacokinetic properties of Forsythiaside A. Pharmacol. Res. 2021, 169, 105690. [Google Scholar] [CrossRef] [PubMed]
- Deng, M.; Chen, H.; Long, J.; Song, J.; Xie, L.; Li, X. Calycosin: A review of its pharmacological effects and application prospects. Expert. Rev. Anti. Infect. Ther. 2021, 19, 911–925. [Google Scholar] [CrossRef]
- Wang, F.; Zhao, S.; Li, F.; Zhang, B.; Qu, Y.; Sun, T.; Luo, T.; Li, D. Investigation of antioxidant interactions between Radix Astragali and Cimicifuga foetida and identification of synergistic antioxidant compounds. PLoS ONE 2014, 9, e87221. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhao, L.H. Calycosin ameliorates diabetes-induced cognitive impairments in rats by reducing oxidative stress via the PI3K/Akt/GSK-3 beta signaling pathway. Biochem. Biophys. Res. Commun. 2016, 473, 428–434. [Google Scholar] [CrossRef]
- Ma, R.; Yuan, F.; Wang, S.; Liu, Y.; Fan, T.; Wang, F. Calycosin alleviates cerulean induced acute pancreatitis by inhibiting the inflammatory response and oxidative stress via the p38 MAPK and NF-κB signal pathways in mice. Biomed. Pharmacother. 2018, 105, 599–605. [Google Scholar] [CrossRef]
- Zhai, J.; Tao, L.; Zhang, S.; Gao, H.; Zhang, Y.; Sun, J.; Song, Y.; Qu, X. Calycosin ameliorates doxorubicin-induced cardiotoxicity by suppressing oxidative stress and inflammation via the sirtuin 1-NOD-like receptor protein 3 pathway. Phytother. Res. 2020, 34, 649–659. [Google Scholar] [CrossRef] [PubMed]
- Phull, P.S.; Green, C.J.; Jacyna, M.R. A radical view of the stomach: The role of oxygen-derived free radicals and anti-oxidants in gastroduodenal disease. Eur. J. Gastroenterol. Hepatol. 1995, 7, 265–274. [Google Scholar]
- Lu, L.; Zhao, X.; Zhang, J.; Li, M.; Qi, Y.; Zhou, L. Calycosin promotes lifespan in Caenorhabditis elegans through insulin signaling pathway via daf-16, age-1 and daf-2. J. Biosci. Bioeng. 2017, 124, 1–7. [Google Scholar] [CrossRef]
- Fu, L.; Wu, W.L.; Jian, Z.; Qiang, S.; Chen, J. Calycosin regulates glucocorticoid-induced apoptosis via Nrf2/ARE signaling in MC3T3-E1 cells. Trop. J. Pharm. Res. 2018, 17, 205–212. [Google Scholar] [CrossRef] [Green Version]
- Park, J.Y.; Shin, M.S.; Hwang, G.S.; Yamabe, N.; Yoo, J.E.; Kang, K.S.; Kim, J.C.; Lee, J.G.; Ham, J.; Lee, H.L. Beneficial effects of deoxyshikonin on delayed wound healing in diabetic mice. Int. J. Mol. Sci. 2018, 19, 3660. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Y.; Zhong, Y.; Long, X.; Zhu, Z.; Zhou, Y.; Ye, H.; Zeng, X.; Zheng, X. Deoxyshikonin isolated from Arnebia euchroma inhibits colorectal cancer by down-regulating the PI3K/Akt/mTOR pathway. Pharm. Biol. 2019, 57, 412–423. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Y.; Qi, A.; Liu, L.; Huang, Q.; Zhang, J.; Cai, K.; Cai, C. Shikonin attenuates H2O2-induced oxidative injury in HT29 cells via antioxidant activities and the inhibition of mitochondrial pathway-mediated apoptosis. Exp. Ther. Med. 2021, 22, 1118. [Google Scholar] [CrossRef]
- Dorn, G.W., II. Apoptotic and non-apoptotic programmed cardiomyocyte death in ventricular remodelling. Cardiovasc. Res. 2009, 81, 465–473. [Google Scholar] [CrossRef]
- Shih, P.H.; Yeh, C.T.; Yen, G.C. Anthocyanins induce the activation of phase II enzymes through the antioxidant response element pathway against oxidative stress-induced apoptosis. J. Agric. Food Chem. 2007, 55, 9427–9435. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Feng, L.; Wang, S.; Zhu, Q.; Lin, J.; Lou, C.; Xiang, P.; He, B.; Zheng, Z.; Tang, D.; et al. Phytoestrogen calycosin-7-Obeta-D-glucopyranoside ameliorates advanced glycation end products-induced HUVEC damage. J. Cell. Biochem. 2011, 112, 2953–2965. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.H.; Xiong, J.; Wang, S.S.; Tang, D.; Wang, R.S.; Zhu, Q. Calycosin entered HUVECs and ameliorated AGEs-promoted cell apoptosis via the Bcl-2 pathway. J. Nat. Med. 2014, 68, 163–172. [Google Scholar] [CrossRef] [PubMed]
- Tian, J.; Duan, Y.; Bei, C.; Chen, J. Calycosin induces apoptosis by upregulation of rasd1 in human breast cancer cells MCF-7. Horm. Metab. Res. 2013, 45, 593–598. [Google Scholar] [CrossRef]
- Tian, W.; Wang, Z.W.; Yuan, B.M.; Bao, Y.G. Calycosin induces apoptosis via p38-MAPK pathway-mediated activation of the mitochondrial apoptotic pathway in human osteosarcoma 143B cells. Mol. Med. Rep. 2020, 22, 3962–3968. [Google Scholar] [CrossRef]
- Buendia, I.; Michalska, P.; Navarro, E.; Gameiro, I.; Egea, J.; León, R. Nrf2-ARE pathway: An emerging target against oxidative stress and neuroinflammation in neurodegenerative diseases. Pharmacol. Ther. 2016, 157, 84–104. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T.; Yamamoto, M. Molecular basis of the keap1-Nrf2 system. Free Radic. Biol. Med. 2015, 88, 93–100. [Google Scholar] [CrossRef] [Green Version]
- Bhattacharjee, S.; Dashwood, R.H. Epigenetic regulation of NRF2/KEAP1 by phytochemicals. Antioxidants 2020, 9, 865. [Google Scholar] [CrossRef]
- Jayasuriya, R.; Dhamodharan, U.; Ali, D.; Ganesan, K.; Xu, B.; Ramkumar, K.M. Targeting Nrf2/Keap1 signaling pathway by bioactive natural agents: Possible therapeutic strategy to combat liver disease. Phytomedicine 2021, 92, 153755. [Google Scholar] [CrossRef]
- Shahcheraghi, S.H.; Salemi, F.; Peirovi, N.; Ayatollahi, J.; Alam, W.; Khan, H.; Saso, L. Nrf2 regulation by curcumin: Molecular aspects for therapeutic prospects. Molecules 2021, 27, 167. [Google Scholar] [CrossRef] [PubMed]
- Su, X.; Huang, Q.; Chen, J.; Wang, M.; Pan, H.; Wang, R.; Liu, L. Calycosin suppresses expression of pro-inflammatory cytokines via the activation of p62/Nrf2-linked heme oxygenase 1 in rheumatoid arthritis synovial fibroblasts. Pharmacol. Res. 2016, 113, 695–704. [Google Scholar] [CrossRef] [PubMed]
- Zhu, D.; Yu, H.; Liu, P.; Yang, Q.; Chen, Y.; Luo, P.; Zhang, C.; Gao, Y. Calycosin modulates inflammation via suppressing TLR4/NF—κB pathway and promotes bone formation to ameliorate glucocorticoid—Induced osteonecrosis of the femoral head in rat. Phytother. Res. 2021, 35, 2824–2835. [Google Scholar] [CrossRef]
- Du, H.; Zhou, X.; Shi, L.; Xia, M.; Wang, Y.; Guo, N.; Hu, H.; Zhang, P.; Yang, H.; Zhu, F.; et al. Shikonin attenuates cochlear spiral ganglion neuron degeneration by activating Nrf2-ARE signaling pathway. Front. Mol. Neurosci. 2022, 15, 829642. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds | Cell Viability | CAS Number | Molecular Formula | Molecular Weight | Structural Family | Source | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| 2.5 μM | 5 μM | 10 μM | 20 μM | 40 μM | ||||||
| Piceatannol | 50.5 | 72.7 | 81.7 | 67.1 | 48.3 | 10083-24-6 | C14H12O4 | 244.24 | Alcohols | Rheum officinale Baill. |
| Cynaroside | 46.7 | 58.2 | 71.2 | 50.0 | 40.1 | 5373-11-5 | C21H20O11 | 448.38 | Glycosides/Flavonoids | Anthriscus sylvestris |
| 2-methoxycinnamaldehyde | 52.8 | 68.4 | 85.4 | 94.6 | 53.2 | 1504-74-1 | C10H10O2 | 162.18 | Phenols | Cinnamomum cassia |
| Quercetin | 48.9 | 52.6 | 77.8 | 68.4 | 55.5 | 117-39-5 | C15H10O7 | 302.24 | Glycosides/Flavonoids | Fructus sophorae |
| 10-Shogaol | 50.9 | 60.6 | 70.7 | 34.8 | 21.2 | 36752-54-2 | C21H32O3 | 332.47 | Phenols | Zingiber officinale |
| 7-O-Methylrosmanol | 59.6 | 60.7 | 69.1 | 58.3 | 51.8 | 113085-62-4 | C21H28O5 | 360.45 | Diterpenoids | Rosmarinus officinalis |
| Andrographolide | 76.0 | 89.2 | 97.1 | 91.5 | 75.4 | 5508-58-7 | C20H30O5 | 350.44 | Terpenoids | Andrographis paniculata |
| Quercetin Dihydrate | 51.5 | 73.7 | 70.8 | 55.8 | 45.3 | 6151-25-3 | C15H14O9 | 338.27 | Flavonoid | Sophora japonica |
| Bisabolangelone | 53.3 | 60.2 | 75.8 | 84.7 | 64.9 | 30557-81-4 | C15H20O3 | 248.32 | Sesquiterpenoids | Angelica sinensis |
| Ethyl Caffeate | 53.1 | 72.2 | 71.0 | 68.7 | 55.6 | 102-37-4 | C11H12O4 | 208.21 | Polyphenols | Bidens pilosa |
| Damascenone | 49.4 | 53.0 | 70.1 | 65.8 | 63.1 | 23696-85-7 | C13H18O | 190.28 | Ketone | Rosa rugosa |
| Morroniside | 51.3 | 68.1 | 78.6 | 61.1 | 43.5 | 25406-64-8 | C17H26O11 | 406.38 | Iridoids | Cinchona |
| Calycosin | 102.6 | 102.8 | 94.7 | 82.5 | 63.9 | 20575-57-9 | C16H12O5 | 284.26 | Flavonoids | Astragalus membranaceus |
| Sinapine | 54.6 | 78.4 | 73.4 | 52.2 | 28.7 | 18696-26-9 | C16H24NO5+ | 310.37 | Alkaloids | Raphanus sativus |
| Artemetin | 70.5 | 72.5 | 68.8 | 57.0 | 39.1 | 479-90-3 | C20H20O8 | 388.37 | Flavonoids | Achillea millefolium |
| Schaftoside | 50.5 | 73.1 | 72.4 | 50.2 | 23.1 | 51938-32-0 | C26H28O14 | 564.49 | Flavonoids | Desmodium styracifolium |
| Deoxyshikonin | 88.2 | 93.8 | 75.3 | 51.9 | 25.1 | 43043-74-9 | C16H16O4 | 272.30 | Quinones | Lithosperraum erythrorhizon |
| Forsythoside A | 56.1 | 64.3 | 74.8 | 78.2 | 42.6 | 79916-77-1 | C29H36O15 | 624.59 | Phenylpropanoids | Forsythia suspensa |
| Isoacteoside | 78.4 | 78.1 | 70.2 | 58.7 | 54.6 | 61303-13-7 | C29H36O15 | 624.59 | Phenylpropanoids | Pedicularis striata Pall. |
| Guaijaverin | 72.5 | 78.5 | 76.0 | 54.6 | 39.1 | 22255-13-6 | C20H18O11 | 434.35 | Flavonoids | Psidium guajava Linn. |
| Demethoxycurcumin | 63.4 | 74.8 | 72.6 | 75.8 | 54.5 | 22608-11-3 | C20H18O5 | 338.36 | Phenols | Zingiber officinale Roscoe. |
| Baicalin | 54.3 | 69.9 | 84.3 | 75.6 | 63.3 | 21967-41-9 | C21H18O11 | 446.36 | Flavonoids | Scutellaria baicalensis |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, J.; Chen, M.; Wang, S.; Chu, X.; Ji, H. Identification of Phytogenic Compounds with Antioxidant Action That Protect Porcine Intestinal Epithelial Cells from Hydrogen Peroxide Induced Oxidative Damage. Antioxidants 2022, 11, 2134. https://doi.org/10.3390/antiox11112134
Wang J, Chen M, Wang S, Chu X, Ji H. Identification of Phytogenic Compounds with Antioxidant Action That Protect Porcine Intestinal Epithelial Cells from Hydrogen Peroxide Induced Oxidative Damage. Antioxidants. 2022; 11(11):2134. https://doi.org/10.3390/antiox11112134
Chicago/Turabian StyleWang, Jing, Meixia Chen, Sixin Wang, Xu Chu, and Haifeng Ji. 2022. "Identification of Phytogenic Compounds with Antioxidant Action That Protect Porcine Intestinal Epithelial Cells from Hydrogen Peroxide Induced Oxidative Damage" Antioxidants 11, no. 11: 2134. https://doi.org/10.3390/antiox11112134