
Hydrogen Sulfide Biomedical Research in China—20 Years of Hindsight

Abstract
:1. Introduction
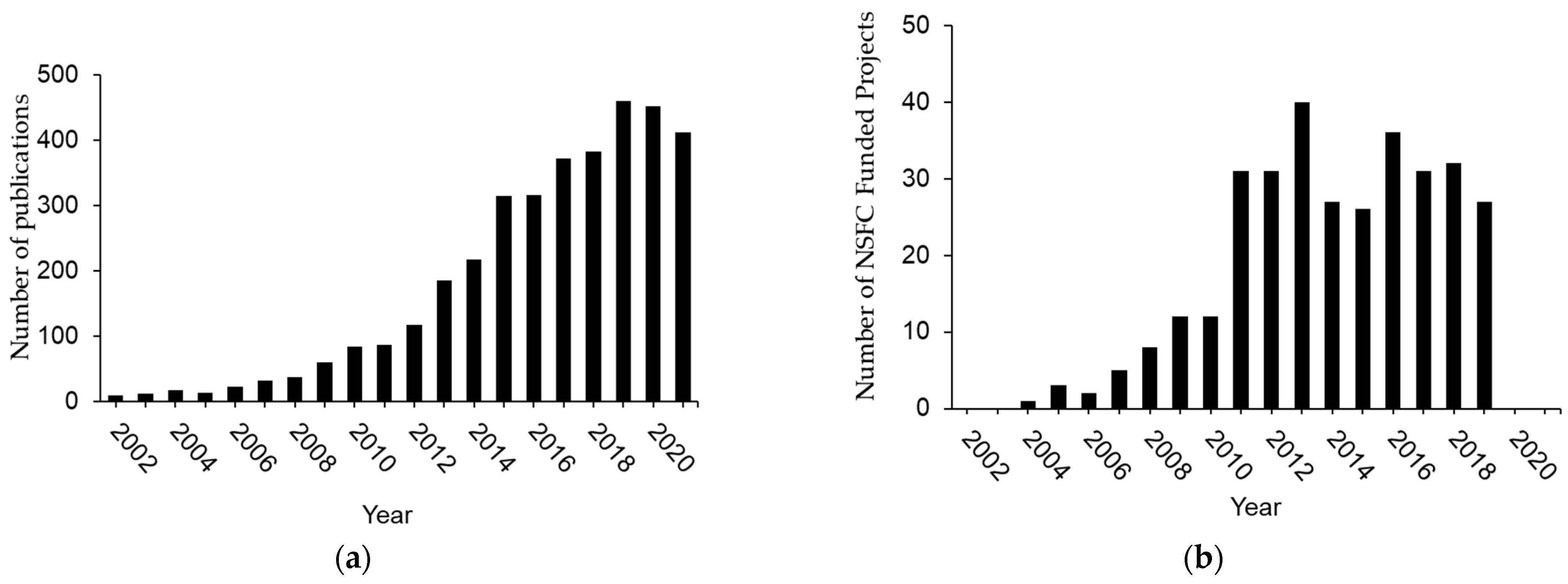
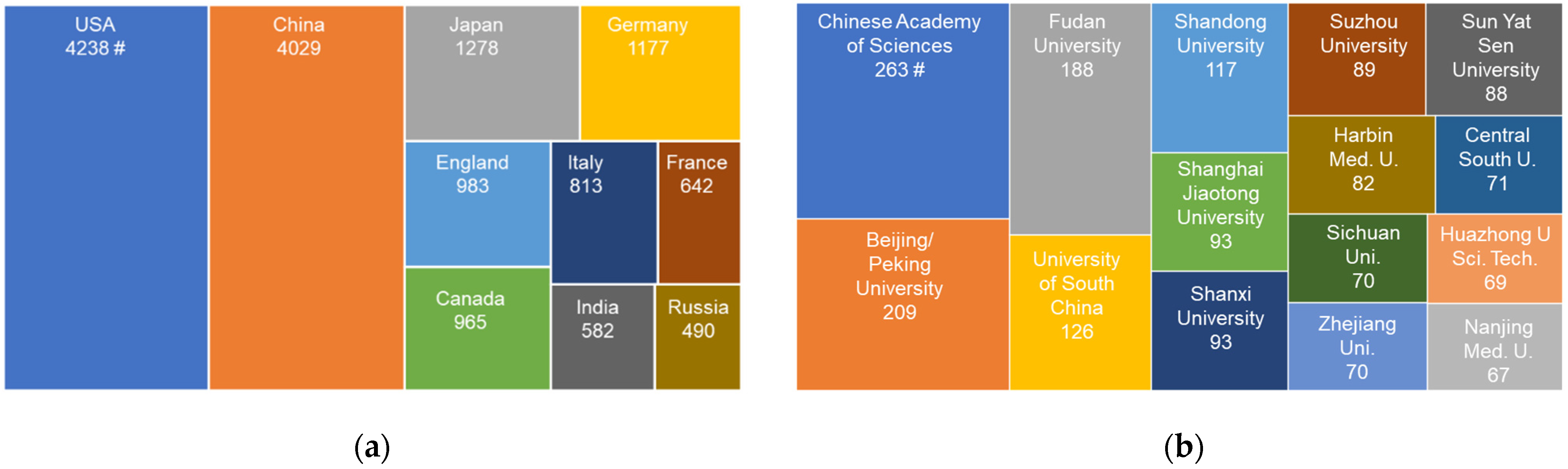
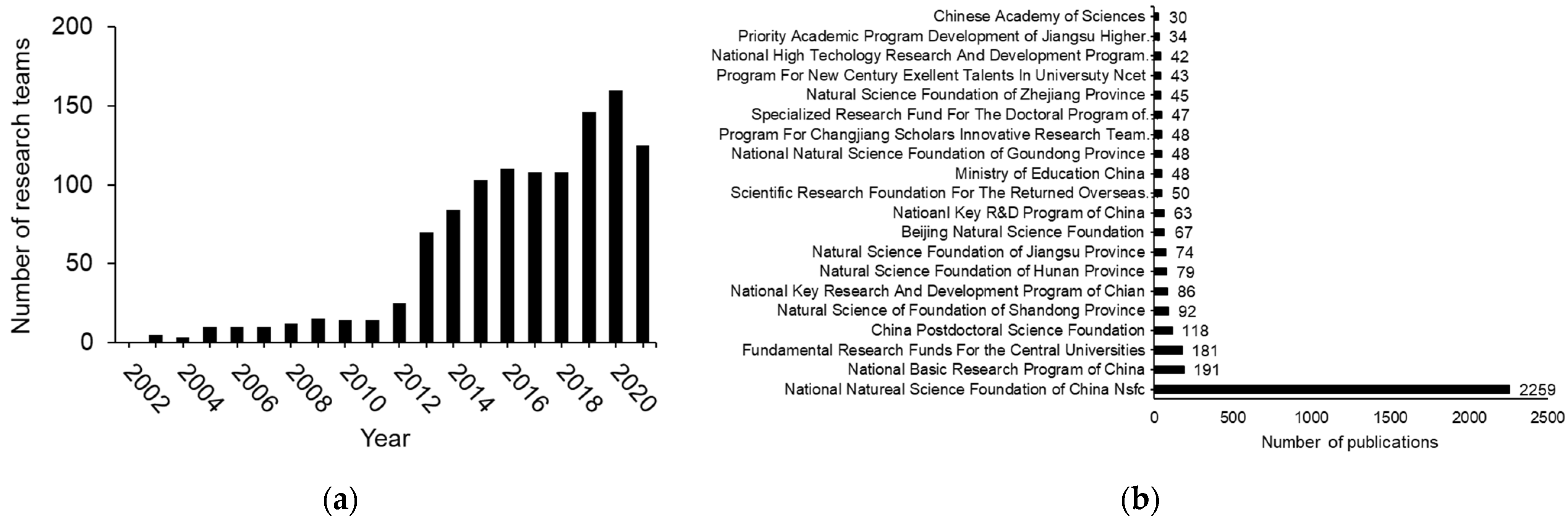
2. Three Phases of H2S Research in China
3. Major Discoveries Made by H2S Researchers in China
3.1. The Correlation of H2S Metabolism with Cardiovascular Diseases
3.1.1. Blood Pressure Regulation by H2S and the Underlying Mechanisms
3.1.2. H2S-Inhibited Vascular Smooth Muscle Proliferation
3.1.3. H2S Regulation of Angiogenesis and Atherosclerosis
3.1.4. The Role of H2S in Heart Failure and Myocardial Pathogeneses
3.2. The Neurobiological Targets of H2S
3.3. H2S and Respiratory Diseases
3.4. Other Important Discoveries of H2S Biomedical Function
3.4.1. Sulfur and Iron Interaction
3.4.2. Regulation of the Reproductive System by H2S
4. Development of H2S-Based Novel Therapeutics
5. Foresight
- As a nation, China needs to have a strategic and systemic plan for efficient and accelerated advancement of H2S biomedical research. As the organizations, China universities and research institutes need to identify and strategically support H2S biomedical research clusters to extend the frontiers of knowledge.
- National and international collaboration and coordinated and networking effort should be continued and strengthened. Persistent but not momentary in the fashion, and organized but not spontaneous or sporadic, are the formula of success for bigger bloom of H2S biomedical research in China.
- Ground-breaking fundamental H2S biomedical research, rather than patching and catching studies, should be encouraged and supported. National and institutional scientific goals in this field should be in place to assure the resource sufficient and metrics laid out.
- Knowledge translation efforts from bench to bedside should be identified and supported. It offers the most tangible benefits to human beings by targeting at the most relevant diseases with high morbidity and mortality/prevalence which are closely related to H2S metabolism and functions.
- Specifically designed training programs should be launched for next generation of H2S scientists, graduate students, post-doc fellows and young scientists.
- H2S biomedical research in China should seek integration and collaboration with other disciplines, such as physics, chemistry, engineering, agriculture, plant science, natural resource management, nutrition, etc.
- The immersion of H2S biomedical study with the Traditional Chinese Medicine (TCM) should be explored. H2S as well as other gasotransmitters may underly the chemical nature of “Qi” in TCM, of which the straight translation is “gas”. From this lens, intangible Qi would become tangible gasotransmitters [84]. As acupuncture in TCM may stimulate the balance and flow of Qi, whether H2S metabolism is regulated by acupuncture becomes intriguing. Furthermore, many Chinese herbs are known to affect microbial production of H2S, such as Dang-shen (Codonopsis pilosula) [85] and Gan-cao (Licorice) [86]. It would be of significant importance to explore the use of Chinese medicinal herbs for providing exogenous source of H2S and for regulating endogenous H2S metabolism.
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Zhang, J.; Lei, D.; Feng, W. Analysis of chemical disasters caused by release of hydrogen sulfide-bearing natural gas. Procedia Eng. 2011, 26, 1878–1890. [Google Scholar]
- Wang, R. Two’s company, three’s a crowd—Can H2S be the third endogenous gaseous transmitter? FASEB J. 2002, 16, 1792–1798. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, R. Physiological implications of hydrogen sulfide: A whiff exploration that blossomed. Physiol. Rev. 2012, 92, 791–896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.; Minkler, P.; Grove, D.; Wang, R.; Willard, B.; Dweik, R.; Hine, C. Non-enzymatic hydrogen sulfide production from cysteine in blood is catalyzed by iron and vitamin B6. Commun. Biol. 2019, 2, 194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Searcy, D.G.; Lee, S.H. Sulfur reduction by human erythrocytes. J. Exp. Zool. 1998, 282, 310–322. [Google Scholar] [CrossRef]
- Yan, H.; Du, J.; Tang, C.; Bin, G.; Jiang, H. Changes in arterial hydrogen sulfide (H2S) content during septic shock and endotoxin shock in rats. J. Infect. 2003, 47, 155–160. [Google Scholar]
- Du, J.; Yan, H.; Tang, C. Endogenous H2S is involved in the development of spontaneous hypertension. Beijing Da Xue Xue Bao Yi Xue Ban 2003, 35, 102. (In Chinese) [Google Scholar] [PubMed]
- Zhang, C.; Du, J.; Bu, D.; Yan, H.; Tang, X.; Tang, C. The regulatory effect of hydrogen sulfide on hypoxic pulmonary hypertension in rats. Biochem. Biophys. Res. Commun. 2003, 302, 810–816. [Google Scholar]
- Du, J.; Chen, X.; Geng, B.; Jiang, H.F.; Tang, C. Hydrogen sulfide as a messenger molecule in cardiovascular system. Beijing Da Xue Xue Bao Yi Xue Ban 2002, 34, 187. (In Chinese) [Google Scholar]
- Wang, R. Toxic gas, lifesaver. Sci. Am. 2010, 302, 66–71. [Google Scholar] [CrossRef] [PubMed]
- Cai, W.J.; Wang, M.J.; Moore, P.K.; Jin, H.M.; Yao, T.; Zhu, Y.C. The novel proangiogenic effect of hydrogen sulfide is dependent on Akt phosphorylation. Cardiovasc. Res. 2007, 76, 29–40. [Google Scholar] [CrossRef] [PubMed]
- Chuah, S.C.; Moore, P.K.; Zhu, Y.Z. S-allylcysteine mediates cardioprotection in an acute myocardial infarction rat model via a hydrogen sulfide-mediated pathway. Am. J. Physiol. Heart Circ. Physiol. 2007, 293, H2693–H2701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, Y.X.; Chen, Y.; Zhu, Y.Z.; Huang, G.Y.; Moore, P.K.; Huang, S.H.; Yao, T.; Zhu, Y.C. Chronic sodium hydrosulfide treatment decreases medial thickening of intramyocardial coronary arterioles, interstitial fibrosis, and ROS production in spontaneously hypertensive rats. Am. J. Physiol. Heart Circ. Physiol. 2007, 293, H2093–H2100. [Google Scholar] [CrossRef]
- Zhao, W.; Zhang, J.; Lu, Y.; Wang, R. The vasorelaxant effect of H2S as a novel endogenous gaseous KATP channel opener. EMBO J. 2001, 20, 6008–6016. [Google Scholar] [CrossRef]
- Zhong, G.; Chen, F.R.; Cheng, Y.Q.; Tang, C.S.; Du, J. The role of hydrogen sulfide generation in the pathogenesis of hypertension in rats induced by inhibition of nitric oxide synthase. J. Hypertens. 2003, 21, 1879–1885. [Google Scholar] [CrossRef]
- Huang, P.; Chen, S.; Wang, Y.; Liu, J.; Yao, Q.; Huang, Y.; Li, H.; Zhu, M.; Wang, S.; Li, L.; et al. Down-regulated CBS/H2S pathway is involved in high-salt-induced hypertension in Dahl rats. Nitric Oxide 2015, 6, 192–203. [Google Scholar] [CrossRef]
- Zong, Y.; Huang, Y.; Chen, S.; Zhu, M.; Chen, Q.; Feng, S.; Sun, Y.; Zhang, Q.; Tang, C.; Du, J.; et al. Downregulation of endogenous hydrogen sulfide pathway is involved in mitochondrion-related endothelial cell apoptosis induced by high salt. Oxid. Med. Cell. Longev. 2015, 2015, 754670. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Du, J.; Zhou, W.; Yan, H.; Tang, C.; Zhang, C. Impact of hydrogen sulfide on carbon monoxide oxygenase pathway in the pathogenesis of hypoxic pulmonary hypertension. Biochem. Biophys. Res. Commun. 2004, 317, 30–37. [Google Scholar]
- Feng, S.; Chen, S.; Wen, Y.; Zheng, D.; Zhang, C.; Tang, C.; Du, J.; Jin, H. H2S inhibits pulmonary arterial endothelial cell inflammation in rats with monocrotaline-induced pulmonary hypertension. Lab. Investig. 2017, 97, 268–278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, H.; Cong, B.; Zhao, B.; Zhang, C.; Liu, X.; Zhou, W.; Shi, Y.; Tang, C.; Du, J. Effects of hydrogen sulfide on hypoxic pulmonary vascular structural remodeling. Life Sci. 2006, 78, 1299–1309. [Google Scholar]
- Zhang, D.; Wang, X.; Chen, S.; Chen, S.; Yu, W.; Liu, X.; Yang, G.; Tao, Y.; Tang, X.; Bu, D.; et al. Endogenous hydrogen sulfide sulfhydrates IKKβ at cysteine 179 to control pulmonary artery endothelial cell inflammation. Clin. Sci. 2019, 133, 2045–2059. [Google Scholar] [CrossRef] [PubMed]
- Tian, X.; Zhou, D.; Zhang, Y.; Song, Y.; Zhang, Q.; Bu, D.; Sun, Y.; Wu, L.; Long, Y.; Tang, C.; et al. Persulfidation of transcription factor FOXO1 at cysteine 457, A novel mechanism by which H2S inhibits vascular smooth muscle cell proliferation. J. Adv. Res. 2021, 27, 155–164. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.; Wu, L.; Wang, R. Pro-apoptotic effect of endogenous H2S on human aorta smooth muscle cells. FASEB J. 2006, 20, 553–555. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.; Wu, L.; Jiang, B.; Yang, W.; Qi, J.; Cao, K.; Meng, Q.; Mustafa, A.K.; Mu, W.; Zhang, S.; et al. H2S as a physiologic vasorelaxant: Hypertension in mice with deletion of cystathionine gamma-lyase. Science 2008, 322, 587–590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, L.; Dong, J.H.; Teng, X.; Jin, S.; Xue, H.M.; Liu, S.Y.; Guo, Q.; Shen, W.; Ni, X.C.; Wu, Y.M. Hydrogen sulfide improves endothelial dysfunction in hypertension by activating peroxisome proliferator-activated receptor delta/endothelial nitric oxide synthase signaling. J. Hypertens. 2018, 36, 651–665. [Google Scholar] [CrossRef] [PubMed]
- Tian, D.Y.; Dong, J.H.; Jin, S.; Teng, X.; Wu, Y.M. Endogenous hydrogen sulfide-mediated MAPK inhibition preserves endothelial function through TXNIP signaling. Free Radic. Biol. Med. 2017, 110, 291–299. [Google Scholar] [CrossRef]
- Teng, X.; Li, H.; Xue, H.M.; Jin, S.; Xiao, L.; Guo, Q.; Wu, Y.M. GABAA receptor, KATP channel and L-type Ca2+ channel is associated with facilitation effect of H2S on the baroreceptor reflex in spontaneous hypertensive rats. Pharmacol. Rep. 2019, 71, 968–975. [Google Scholar] [CrossRef]
- Duan, X.C.; Guo, R.; Liu, S.Y.; Xiao, L.; Xue, H.M.; Guo, Q.; Jin, S.; Wu, Y.M. Gene transfer of cystathionine β-synthase into RVLM increases hydrogen sulfide-mediated suppression of sympathetic outflow via KATP channel in normotensive rats. Am. J. Physiol. Heart Circ. Physiol. 2015, 308, H603–H611. [Google Scholar] [CrossRef] [Green Version]
- Du, J.; Hui, Y.; Cheung, Y.; Bin, G.; Jiang, H.; Chen, X.; Tang, C. The possible role of hydrogen sulfide as a smooth muscle cell proliferation inhibitor in rat cultured cells. Heart Vessel. 2004, 19, 75–80. [Google Scholar] [CrossRef]
- Yang, G.; Sun, X.; Wang, R. Hydrogen sulfide-induced apoptosis of human aorta smooth muscle cells via the activation of mitogen-activated protein kinases and caspase-3. FASEB J. 2004, 18, 1782–1784. [Google Scholar] [CrossRef]
- Li, L.; Liu, D.; Bu, D.F.; Chen, S.; Wu, J.; Tang, C.S.; Du, J.; Jin, H.F. Brg1-dependent epigenetic control of vascular smooth muscle cell proliferation by hydrogen sulfide. Biochim. Biophys. Acta—Mol. Cell Res. 2013, 1833, 1347–1355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Zhao, X.; Jin, H.; Wei, H.; Li, W.; Bu, D.; Tang, X.; Ren, Y.; Tang, C.; Du, J. Role of hydrogen sulfide in the development of atherosclerotic lesions in apolipoprotein E knockout mice. Arterioscler. Thromb. Vasc. Biol. 2009, 29, 173–179. [Google Scholar] [CrossRef] [PubMed]
- Xie, L.; Gu, Y.; Wen, M.; Zhao, S.; Wang, W.; Ma, Y.; Meng, G.; Han, Y.; Wang, Y.; Liu, G.; et al. Hydrogen sulfide induces keap1 S-sulfhydration and suppresses diabetes-accelerated atherosclerosis via Nrf2 activation. Diabetes 2016, 65, 3171–3184. [Google Scholar] [CrossRef] [Green Version]
- Mani, S.; Li, H.; Untereiner, A.; Wu, L.; Yang, G.; Austin, R.C.; Dickhout, J.G.; Lhoták, Š.; Meng, Q.H.; Wang, R. Decreased endogenous production of hydrogen sulfide accelerates atherosclerosis. Circulation 2013, 127, 2523–2534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, J.; Huang, Y.; Yan, H.; Zhang, Q.; Zhao, M.; Zhu, M.; Liu, J.; Chen, S.X.; Bu, D.; Tang, C.; et al. Hydrogen sulfide suppresses oxidized low-density lipoprotein (Ox-LDL)-stimulated monocyte chemoattractant protein 1 generation from macrophages via the nuclear factor κB (NF-κB) pathway. J. Biol. Chem. 2014, 289, 9741–9753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, H.; Zhang, R.; Jin, H.; Liu, D.; Tang, X.; Tang, C.; Du, J. Hydrogen sulfide attenuates hyperhomocysteinemia-induced cardiomyocytic endoplasmic reticulum stress in rats. Antioxid. Redox Signal. 2010, 12, 1079–1091. [Google Scholar] [CrossRef]
- Meng, G.; Liu, J.; Liu, S.; Song, Q.; Liu, L.; Xie, L.; Han, Y.; Ji, Y. Hydrogen sulfide pretreatment improves mitochondrial function in myocardial hypertrophy via a SIRT3-dependent manner. Br. J. Pharmacol. 2018, 175, 1126–1145. [Google Scholar] [CrossRef] [Green Version]
- Meng, G.; Xiao, Y.; Ma, Y.; Tang, X.; Xie, L.; Liu, J.; Gu, Y.; Yu, Y.; Park, C.M.; Xian, M.; et al. Hydrogen sulfide regulates Krüppel-like factor 5 transcription activity via specificity protein 1 S-sulfhydration at Cys664 to prevent myocardial hypertrophy. J. Am. Heart Assoc. 2016, 5, e004160. [Google Scholar] [CrossRef]
- Pan, L.L.; Liu, X.H.; Shen, Y.Q.; Wang, N.Z.; Xu, J.; Wu, D.; Xiong, Q.H.; Deng, H.Y.; Huang, G.Y.; Zhu, Y.Z. Inhibition of NADPH oxidase 4-related signaling by sodium hydrosulfide attenuates myocardial fibrotic response. Int. J. Cardiol. 2013, 168, 3770–3778. [Google Scholar] [CrossRef]
- Tang, X.Q.; Yang, C.T.; Chen, J.; Yin, W.L.; Tian, S.W.; Hu, B.; Feng, J.Q.; Li, Y.J. Effect of hydrogen sulphide on beta-amyloid-induced damage in PC12 cells. Clin. Exp. Pharmacol. Physiol. 2008, 35, 180–186. [Google Scholar]
- Yin, W.L.; He, J.Q.; Hu, B.; Jiang, Z.S.; Tang, X.Q. Hydrogen sulfide inhibits MPP(+)-induced apoptosis in PC12 cells. Life Sci. 2009, 85, 269–275. [Google Scholar] [CrossRef]
- Tang, X.Q.; Zhuang, Y.Y.; Zhang, P.; Fang, H.R.; Zhou, C.F.; Gu, H.F.; Zhang, H.; Wang, C.Y. Formaldehyde impairs learning and memory involving the disturbance of hydrogen sulfide generation in the hippocampus of rats. J. Mol. Neurosci. 2013, 49, 140–149. [Google Scholar] [CrossRef] [PubMed]
- Wei, L.; Kan, L.Y.; Zeng, H.Y.; Tang, Y.Y.; Huang, H.L.; Xie, M.; Zou, W.; Wang, C.Y.; Zhang, P.; Tang, X.Q. BDNF/TrkB pathway mediates the antidepressant-like role of H2S in CUMS-exposed rats by inhibition of hippocampal ER stress. Neuromolecular Med. 2018, 20, 252–261. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.Y.; Wei, H.J.; Wu, L.; Liu, S.M.; Tang, Y.Y.; Zou, W.; Wang, C.Y.; Zhang, P.; Tang, X.Q. BDNF-TrkB pathway mediates antidepressant-like roles of H2S in diabetic rats via promoting hippocampal autophagy. Clin. Exp. Pharmacol. Physiol. 2020, 47, 302–312. [Google Scholar] [CrossRef]
- Wang, Y.; Qu, R.; Hu, S.; Xiao, Y.; Jiang, X.; Xu, G.-Y. Upregulation of cystathionine β-synthetase expression contributes to visceral hyperalgesia induced by heterotypic intermittent stress in rats. PLoS ONE 2012, 7, e53165. [Google Scholar]
- Li, L.; Xie, R.; Hu, S.; Wang, Y.; Yu, T.; Xiao, Y.; Jiang, X.; Gu, J.; Hu, C.Y.; Xu, G.Y. Upregulation of cystathionine beta-synthetase expression by nuclear factor-kappa B activation contributes to visceral hypersensitivity in adult rats with neonatal maternal deprivation. Mol. Pain 2012, 8, 89. [Google Scholar] [CrossRef] [Green Version]
- Hu, S.; Xu, W.; Miao, X.; Gao, Y.; Zhu, L.; Zhou, Y.; Xiao, Y.; Xu, G.Y. Sensitization of sodium channels by cystathionine β-synthetase activation in colon sensory neurons in adult rats with neonatal maternal deprivation. Exp. Neurol. 2013, 248, 275–285. [Google Scholar] [CrossRef]
- Feng, X.; Zhou, Y.L.; Meng, X.; Qi, F.-H.; Jiang, X.; Xu, G.-Y. Hydrogen sulfide increases excitability through suppression of sustained potassium channel currents of rat trigeminal ganglion neurons. Mol Pain 2013, 9, 4. [Google Scholar] [CrossRef]
- Qi, F.; Zhou, Y.; Xiao, Y.; Tao, J.; Gu, J.; Jiang, X.; Xu, G.Y. Promoter demethylation of cystathionine-β-synthetase gene contributes to inflammatory pain in rats. Pain 2013, 154, 34–45. [Google Scholar] [CrossRef]
- Chen, Y.H.; Yao, W.Z.; Geng, B.; Ding, Y.L.; Lu, M.; Zhao, M.W.; Tang, C.S. Endogenous hydrogen sulfide in patients with COPD. Chest 2005, 128, 3205–3211. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, X.M.; Chen, Y.H.; Yao, W.Z. Correlation between Levels of exhaled hydrogen sulfide and airway inflammatory phenotype in patients with chronic persistent asthma. Respirology 2014, 19, 1165–1169. [Google Scholar] [CrossRef]
- Sun, Y.; Wang, K.Y.; Li, M.X.; He, W.; Chang, J.R.; Liao, C.C.; Lin, F.; Qi, Y.F.; Wang, R.; Chen, Y.H. Metabolic changes of H2S in smokers and patients of COPD which might involve in inflammation, oxidative stress and steroid sensitivity. Sci. Rep. 2015, 5, 14971. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.H.; Wang, P.P.; Wang, X.M.; He, Y.J.; Yao, W.Z.; Qi, Y.F.; Tang, C.S. Involvement of endogenous hydrogen sulfide in cigarette smoke-induced changes in airway responsiveness and inflammation of rat lung. Cytokine 2011, 53, 334–341. [Google Scholar] [CrossRef] [PubMed]
- Jia, G.H.; Yu, S.W.; Sun, W.L.; Yang, J.; Wang, Y.; Qi, Y.F.; Chen, Y.H. Hydrogen sulfide attenuates particulate matter-induced emphysema and airway inflammation through Nrf2-dependent manner. Front. Pharmacol. 2020, 11, 29. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M.W.; Yang, G.; Zhou, Y.F.; Qian, C.; Mu, M.D.; Ke, Y.; Qian, Z.M. Regulating ferroportin-1 and transferrin receptor-1 expression: A novel function of hydrogen sulfide. J. Cell. Physiol. 2019, 234, 3158–3169. [Google Scholar] [CrossRef]
- Zhou, Y.F.; Wu, X.M.; Zhou, G.; Mu, M.D.; Zhang, F.L.; Li, F.M.; Qian, C.; Du, F.; Yung, W.H.; Qian, Z.M.; et al. Cystathionine β-synthase is required for body iron homeostasis. Hepatology 2018, 67, 21–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, P.; Qian, C.; Chen, Y.J.; Sheng, Y.; Ke, Y.; Qian, Z.M. Cystathionine β-synthase (CBS) deficiency suppresses erythropoiesis by disrupting expression of heme biosynthetic enzymes and transporter. Cell Death Dis. 2019, 10, 708. [Google Scholar] [CrossRef] [Green Version]
- Módis, K.; Ramanujam, V.S.; Govar, A.A.; Lopez, E.; Anderson, K.E.; Wang, R.; Szabo, C. Cystathionine-γ-lyase (CSE) deficiency increases erythropoiesis and promotes mitochondrial electron transport via the upregulation of coproporphyrinogen III oxidase and consequent stimulation of heme biosynthesis. Biochem. Pharmacol. 2019, 169, 113604. [Google Scholar] [CrossRef]
- Patel, P.; Vatish, M.; Heptinstall, J.; Wang, R.; Carson, R.J. The endogenous production of hydrogen sulphide in intrauterine tissues. Reprod. Biol. Endocrinol. 2009, 7, 10. [Google Scholar] [CrossRef] [Green Version]
- D’Emmanuele di Villa Bianca, R.; Sorrentino, R.; Maffia, P.; Mirone, V.; Imbimbo, C.; Fusco, F.; De Palma, R.; Ignarro, L.J.; Cirino, G. Hydrogen sulfide as a mediator of human corpus cavernosum smooth-muscle relaxation. Proc. Natl. Acad. Sci. USA 2009, 106, 4513–4518. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Yang, J.; Wang, T.; Wang, S.G.; Liu, J.H.; Yin, C.P.; Ye, Z.Q. Decreased endogenous hydrogen sulfide generation in penile tissues of diabetic rats with erectile dysfunction. J. Sex. Med. 2016, 13, 350–360. [Google Scholar] [CrossRef]
- Zuo, C.; Huang, Y.M.; Jiang, R.; Yang, H.F.; Cheng, B.; Chen, F. Endogenous hydrogen sulfide and androgen deficiency-induced erectile dysfunction in rats. Zhonghua Nan Ke Xue 2014, 20, 605–612. (In Chinese) [Google Scholar]
- Zhu, X.B.; Jiang, J.; Jiang, R.; Chen, F. Expressions of CSE and CBS in the corpus cavernosum of spontaneous hypertensive rats. Zhonghua Nan Ke Xue 2014, 20, 4–9. (In Chinese) [Google Scholar] [PubMed]
- Huang, Y.M.; Xia, J.Y.; Jiang, R. Expressions of CSE and CBS in the penile corpus cavernosum of hyperglycemia rats and their implications. Zhonghua Nan Ke Xue 2014, 20, 299–303. (In Chinese) [Google Scholar] [PubMed]
- Wang, J.; Wang, W.; Li, S.; Han, Y.; Zhang, P.; Meng, G.; Xiao, Y.; Xie, L.; Wang, X.; Sha, J.; et al. Hydrogen sulfide as a potential target in preventing spermatogenic failure and testicular dysfunction. Antioxid. Redox Signal. 2018, 28, 1447–1462. [Google Scholar] [CrossRef] [PubMed]
- Benavides, G.A.; Squadrito, G.L.; Mills, R.W.; Patel, H.D.; Isbell, T.S.; Patel, R.P.; Darley-Usmar, V.M.; Doeller, J.E.; Kraus, D.W. Hydrogen sulfide mediates the vasoactivity of garlic. Proc. Natl. Acad. Sci. USA 2007, 104, 17977–17982. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gong, Q.H.; Wang, Q.; Pan, L.L.; Liu, X.H.; Xin, H.; Zhu, Y.Z. S-propargyl-cysteine, a novel hydrogen sulfide-modulated agent, attenuates lipopolysaccharide-induced spatial learning and memory impairment: Involvement of TNF signaling and NF-κB pathway in rats. Brain Behav. Immun. 2011, 25, 110–119. [Google Scholar] [CrossRef]
- Hu, Q.; Wu, D.; Ma, F.; Yang, S.; Tan, B.; Xin, H.; Gu, X.; Chen, X.; Chen, S.; Mao, Y.; et al. Novel angiogenic activity and molecular mechanisms of ZYZ-803, a slow-releasing hydrogen sulfide-nitric oxide hybrid molecule. Antioxid. Redox Signal. 2016, 25, 498–514. [Google Scholar] [CrossRef]
- Fang, L.; Zhao, J.; Chen, Y.; Ma, T.; Xu, G.; Tang, C.; Liu, X.; Geng, B. Hydrogen sulfide derived from periadventitial adipose tissue is a vasodilator. J. Hypertens. 2009, 27, 2174–2185. [Google Scholar] [CrossRef]
- Fan, J.; Zheng, F.; Li, S.; Cui, C.; Jiang, S.; Zhang, J.; Cai, J.; Cui, Q.; Yang, J.; Tang, X.; et al. Hydrogen sulfide lowers hyperhomocysteinemia dependent on cystathionine gamma lyase S-sulfhydration in ApoE-knockout atherosclerotic mice. Br. J. Pharmacol. 2019, 176, 3180–3192. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Gao, J.; Sun, W.; Li, L.; Wang, Y.; Bai, S.; Li, X.; Wang, R.; Wu, L.; Li, H.; et al. Involvement of exogenous H2S in recovery of cardioprotection from ischemic post-conditioning via increase of autophagy in the aged hearts. Int. J. Cardiol. 2016, 220, 681–692. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Gao, J.; Sun, W.; Wen, X.; Xi, Y.; Wang, Y.; Wei, C.; Xu, C.; Li, H. H2S restores the cardioprotective effects of ischemic post-conditioning by upregulating HB-EGF/EGFR signaling. Aging. 2019, 11, 1745–1758. [Google Scholar] [CrossRef] [PubMed]
- Du, J.; Chen, X.; Geng, B.; Jiang, H.; Tang, C. Hydrogen sulfide as a messenger molecule in cardiovascular system. J. Peking Univ. (Health Sci.) 2002, 34, 1871. [Google Scholar]
- Jin, H.; Du, S.X.; Zhao, X.; Wei, H.L.; Wang, Y.F.; Liang, Y.F.; Tang, C.; Du, J. Effects of endogenous sulfur dioxide on monocrotaline-induced pulmonary hypertension in rats. Acta Pharmacol. Sin. 2008, 29, 1157–1166. [Google Scholar] [CrossRef]
- Qu, R.; Tao, J.; Wang, Y.; Zhou, Y.; Wu, G.; Xiao, Y.; Hu, C.; Jiang, X.; Xu, G.-Y. Neonatal colonic inflammation sensitizes voltage-gated Na(+) channels via upregulation of cystathionine beta-synthetase expression in rat primary sensory neurons. Am. J. Physiol. Gastrointest. Liver Physiol. 2013, 304, G763–G772. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; Tian, Z.; Liu, N.; Zhang, L.; Gao, Z.; Sun, X.; Yu, M.; Wu, J.; Yang, F.; Zhao, Y.; et al. Exogenous H2S switches cardiac energy substrate metabolism by regulating SIRT3 expression in db/db mice. J. Mol. Med. (Berl.) 2018, 96, 281–299. [Google Scholar] [CrossRef]
- Sun, X.; Zhao, D.; Lu, F.; Peng, S.; Yu, M.; Liu, N.; Sun, Y.; Du, H.; Wang, B.; Chen, J.; et al. Hydrogen sulfide regulates muscle RING finger-1 protein S-sulfhydration at Cys44 to prevent cardiac structural damage in diabetic cardiomyopathy. Br. J. Pharmacol. 2020, 177, 836–856. [Google Scholar] [CrossRef]
- Yu, M.; Du, H.; Wang, B.; Chen, J.; Lu, F.; Peng, S.; Sun, Y.; Liu, N.; Sun, X.; Zhao, Y.; et al. Exogenous H2S induces Hrd1 S-sulfhydration and prevents CD36 translocation via VAMP3 ubiquitylation in diabetic hearts. Aging Dis. 2020, 11, 286–300. [Google Scholar] [CrossRef] [Green Version]
- Tao, B.B.; Liu, S.Y.; Zhang, C.C.; Fu, W.; Cai, W.J.; Wang, Y.; Shen, Q.; Wang, M.J.; Chen, Y.; Zhang, L.J.; et al. VEGFR2 functions as an H2S-targeting receptor protein kinase with its novel Cys1045-Cys1024 disulfide bond serving as a specific molecular switch for hydrogen sulfide actions in vascular endothelial cells. Antioxid. Redox Signal. 2013, 19, 448–464. [Google Scholar] [CrossRef] [Green Version]
- Xue, R.; Hao, D.D.; Sun, J.P.; Li, W.W.; Zhao, M.M.; Li, X.H.; Chen, Y.; Zhu, J.H.; Ding, Y.J.; Liu, J.; et al. Hydrogen sulfide treatment promotes glucose uptake by increasing insulin receptor sensitivity and ameliorates kidney lesions in type 2 diabetes. Antioxid. Redox Signal. 2013, 19, 5–23. [Google Scholar] [CrossRef]
- Ge, S.N.; Zhao, M.M.; Wu, D.D.; Chen, Y.; Wang, Y.; Zhu, J.H.; Cai, W.J.; Zhu, Y.Z.; Zhu, Y.C. Hydrogen sulfide targets EGFR Cys797/Cys798 residues to induce Na(+)/K(+)-ATPase endocytosis and inhibition in renal tubular epithelial cells and increase sodium excretion in chronic salt-loaded rats. Antioxid. Redox Signal. 2014, 21, 2061–2082. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Q.; Wang, X.L.; Liu, H.R.; Rose, P.; Zhu, Y.Z. Protective effects of cysteine analogues on acute myocardial ischemia: Novel modulators of endogenous H2S production. Antioxid. Redox Signal. 2010, 12, 1155–1165. [Google Scholar] [CrossRef] [Green Version]
- Wu, W.; Qin, M.; Jia, W.; Huang, Z.; Li, Z.; Yang, D.; Huang, M.; Xiao, C.; Long, F.; Mao, J.; et al. Cystathionine-γ-lyase ameliorates the histone demethylase JMJD3-mediated autoimmune response in rheumatoid arthritis. Cell. Mol. Immunol. 2019, 16, 694–705. [Google Scholar] [CrossRef]
- Li, W.-W.; Guo, H.; Wang, X.-M. Relationship between endogenous hydrogen sulfide and blood stasis syndrome based on the Qi-blood theory of Chinese medicine. Chin. J. Integr. Med. 2013, 19, 701–705. [Google Scholar] [CrossRef]
- Jing, Y.; Li, A.; Liu, Z.; Yang, P.; Wei, J.; Chen, X.; Zhao, T.; Bai, Y.; Zha, L.; Zhang, C. Absorption of Codonopsis pilosula saponins by coexisting polysaccharides alleviates gut microbial dysbiosis with dextran sulfate sodium-induced colitis in model mice. BioMed Res. Int. 2018, 2018, 1781036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, J.; Liu, Y.; Guo, J.; Tao, W.; Chen, Y.; Fan, X.; Shen, J.; Duan, J.-A. Health risk of Licorice-Yuanhua combination through induction of colonic H2S metabolism. J. Ethnopharmacol. 2019, 236, 136–146. [Google Scholar] [CrossRef]
- Wang, R. We are what we smell—Not only rotten eggs. Lab. Investig. 2006, 86, 324–325. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Conferences | Date | Location | Attendees | Organizers |
|---|---|---|---|---|
| The 1st China National symposium on H2S gasotransmitter | 24 November 2007 | Beijing, China | 169 | Tang C, Du J, Zhu YC, Zhu YZ, Yao T |
| The First International Conference of H2S in Biology and Medicine | 26–28 June 2009 | Shanghai, China | ~250 | Wang R (Canada), Zhu YZ (China) |
| The 2nd China National symposium on H2S gasotransmitter | 26 June 2010 | Beijing, China | 150 | Du J, Tang C, Zhu YC, Zhu YZ |
| The 3rd China National symposium on H2S gasotransmitter | 26 October 2013 | Beijing, China | 200 | Tang C, Du J, Zhu YC, Zhu YZ, Jin H, Geng B |
| Name * | Affiliation | Discoveries | References | Future Research Directions |
|---|---|---|---|---|
| Chen, Yahong | Peking University, Third Hospital |
|
| |
| Geng, Bin | Chinese Academy of Medical Sciences and Peking Union Medical College, |
|
| |
| Ji, Yong | Nanjing Medical University |
|
| |
| Li, Hongzhu | Xiamen University |
|
| |
| Tang, Chao-Shu Du, Junbao Jin, Hong-Fang | Peking University |
|
|
|
| Tang, Xiao Qing | University of South China |
|
| |
| Wu, Yuming | Hebei Medical University |
|
| |
| Xu, Guang-Yin | Soochow University |
|
| |
| Zhang, Weihua | Harbin Medical University |
|
| |
| Zhu, Yi-Chun | Fudan University |
|
| |
| Zhu, Yi-Zhun | Marco Science and Technology University |
|
|
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, R.; Tang, C. Hydrogen Sulfide Biomedical Research in China—20 Years of Hindsight. Antioxidants 2022, 11, 2136. https://doi.org/10.3390/antiox11112136
Wang R, Tang C. Hydrogen Sulfide Biomedical Research in China—20 Years of Hindsight. Antioxidants. 2022; 11(11):2136. https://doi.org/10.3390/antiox11112136
Chicago/Turabian StyleWang, Rui, and Chaoshu Tang. 2022. "Hydrogen Sulfide Biomedical Research in China—20 Years of Hindsight" Antioxidants 11, no. 11: 2136. https://doi.org/10.3390/antiox11112136