
Carica papaya Reduces Muscle Insulin Resistance via IR/GLUT4 Mediated Signaling Mechanisms in High Fat Diet and Streptozotocin-Induced Type-2 Diabetic Rats
 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Collection of Plant Material
2.3. Phytochemical Qualitative Analysis of C. papaya
2.4. In Vitro Antioxidant Analysis
2.4.1. DPPH Radical Scavenging Activity of C. papaya
2.4.2. Nitric Oxide Radical Scavenging Activity of C. papaya
2.4.3. Superoxide Anion Scavenging Activity of C. papaya
2.5. In Vivo Study
Animals
2.6. Induction of T2DM
2.7. Experimental Design
2.8. FBG
2.9. OGTT
2.10. Serum Insulin
2.11. Homeostasis Model Assessment for Insulin Resistance (HOMA-IR)
2.12. Lipid Profile
2.13. Glycogen Level
2.14. Oxidation Stress Marker
2.15. Enzymatic Antioxidants
2.16. mRNA Expression Analysis
Total RNA Isolation, cDNA Conversion and Real-Time PCR
2.17. Histopathology
2.18. Statistical Analysis
2.19. Molecular Docking
2.19.1. Compound/Ligand Preparation
- Hydrogen bond donors (OH, NH and SH atoms) less than or equal to 5;
- Hydrogen bond acceptors (N, O, and S atoms) 10;
- Molecular weight 500;
- log P;
- Compounds that violate any of the above principles are dubious to own a high rate of oral bioavailability [22].

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| S.No | Compound Name |
|---|---|
| 1 | Caffeic acid |
| 2 | Chlorogenic acid |
| 3 | Kaempferol |
| 4 | Quercetin |
| 5 | Rutin |
| 6 | p-coumaric acid |
| 7 | Trans-Ferulic acid |
| 8 | Protocatechuic acid |
2.19.2. Protein Preparation
2.19.3. Molecular Docking Procedure
2.20. Molecular Simulation and Dynamics
2.20.1. Molecular Dynamic Simulation for IR Complex
2.20.2. Molecular Dynamics Simulation for GLUT-4
3. Results
3.1. Phytochemical Analysis of Ethanolic Extract of C. papaya
3.2. In Vitro Antioxidant Analysis
3.2.1. DPPH Free Radical Scavenging Activity of C. papaya
3.2.2. Nitric Oxide Radical Scavenging Activity of C. papaya
3.2.3. Super Oxide Radical Scavenging Activity of C. papaya
3.3. Effect of C. papaya on Fasting Blood Glucose
3.4. Effect of C. papaya on Oral Glucose Tolerance (OGT)
3.5. Effect of C. papaya on Serum Insulin and HOMA-IR
3.6. Effect of C. papaya on Lipid Profile
3.7. Evaluation of C. papaya on Glycogen Level
3.8. Effect of C. papaya on Oxidation Stress Marker
3.9. Effect of C. papaya on Tissue Antioxidants Enzymes
3.10. Effect of C. papaya mRNA Expression on IR and GLUT4
3.11. Effect of C. papaya on the Histopathological Changes in the Skeletal Muscle
3.12. In Silico Study
3.12.1. Study of Pharmacokinetic Profiling
3.12.2. Molecular Docking Based on Dock Score Proposed Natural Compounds and Selected Target Proteins (IR and GLUT4)
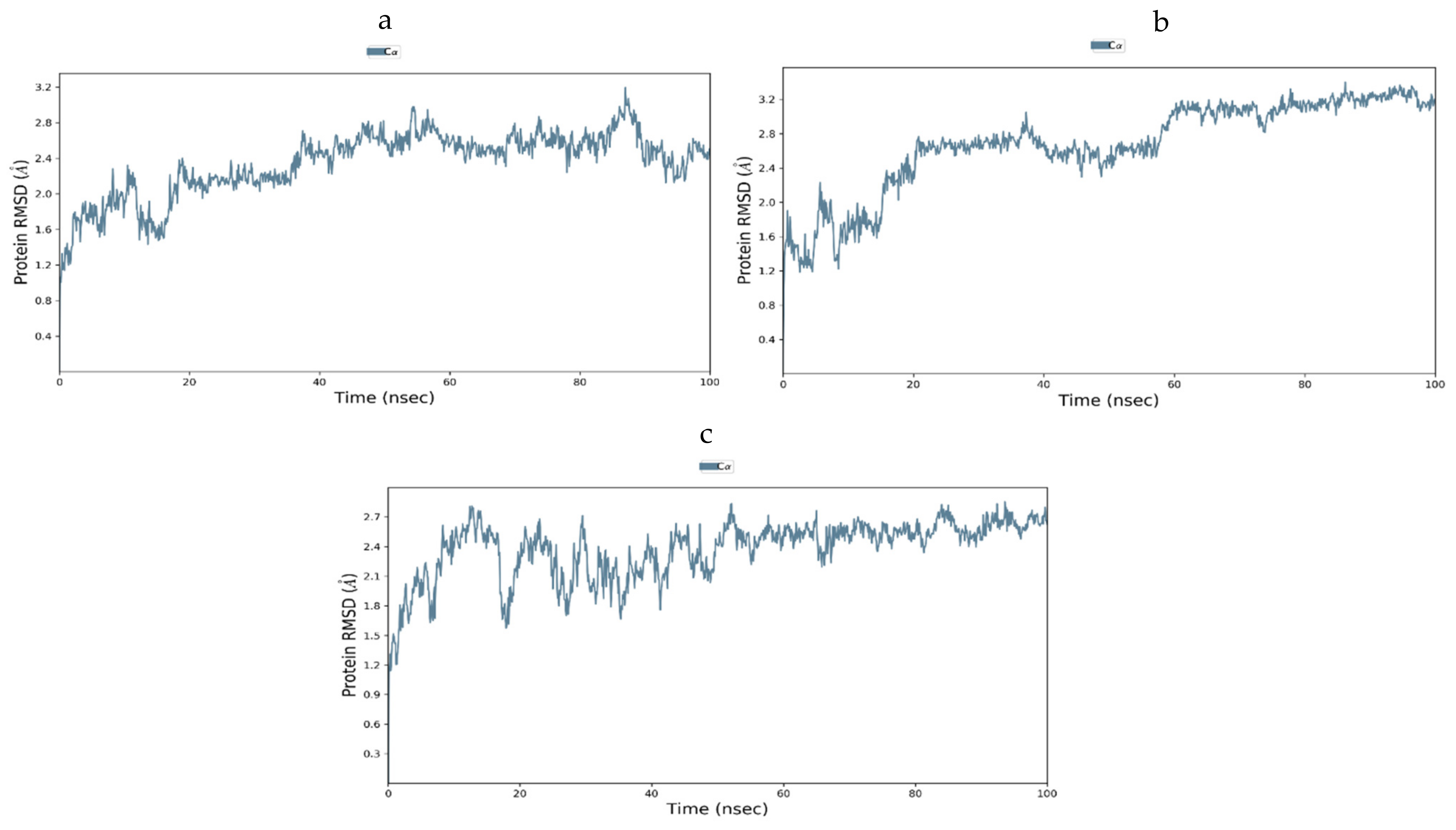
3.12.3. Molecular Dynamic Simulation of IR
3.12.4. Molecular Dynamics of GLUT-4
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Forbes, J.M.; Cooper, M.E. Mechanisms of Diabetic Complications. Physiol. Rev. 2013, 93, 137–188. [Google Scholar] [CrossRef] [PubMed]
- Kallikazaros, I.E. Diabetes mellitus: A sweet-and-sour disease. Hell. J. Cardiol. 2013, 5, 153–154. [Google Scholar]
- Talukder, A.; Hossain, M.Z. Prevalence of Diabetes Mellitus and Its Associated Factors in Bangladesh: Application of Two-level Logistic Regression Model. Sci. Rep. 2020, 10, 10237. [Google Scholar] [CrossRef] [PubMed]
- Animaw, W.; Seyoum, Y. Increasing prevalence of diabetes mellitus in a developing country and its related factors. PLoS ONE 2017, 12, e0187670. [Google Scholar] [CrossRef]
- Kharroubi, A.T.; Darwish, H.M. Diabetes mellitus: The epidemic of the century. World J. Diabetes 2015, 6, 850–867. [Google Scholar] [CrossRef]
- Mason, S.; Wadley, G.D. Skeletal muscle reactive oxygen species: A target of good cop/bad cop for exercise and disease. Redox Rep. 2014, 19, 97–106. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Keung, W.; Samokhvalov, V.; Wang, W.; Lopaschuk, G.D. Role of fatty acid uptake and fatty acid beta-oxidation in mediating insulin resistance in heart and skeletal muscle. Biochim. Biophys. Acta 2010, 1801, 1–22. [Google Scholar] [CrossRef]
- Dresner, A.; Laurent, D.; Marcucci, M.; Griffin, M.E.; Dufour, S.; Cline, G.W.; Slezak, L.A.; Andersen, D.K.; Hundal, R.S.; Rothman, D.L.; et al. Effects of free fatty acids on glucose transport and IRS-1-associated phosphatidylinositol 3-kinase activity. J. Clin. Investig. 1999, 103, 253–259. [Google Scholar] [CrossRef] [Green Version]
- Randle, P.J.; Garland, P.B.; Hales, C.N.; Newsholme, E.A. The glucose fatty-acid cycle. Its role in insulin sensitivity and the metabolic disturbances of diabetes mellitus. Lancet 1963, 1, 785–789. [Google Scholar] [CrossRef]
- Singh, S.P.; Kumar, S.; Mathan, S.V.; Tomar, M.S.; Singh, R.K.; Verma, P.K.; Kumar, A.; Kumar, S.; Singh, R.P.; Acharya, A. Therapeutic application of Carica papaya leaf extract in the management of human diseases. DARU 2020, 28, 735–744. [Google Scholar] [CrossRef]
- Balamurugan, V.; Fatima, M.A.S.; Velurajan, S. A guide to phytochemical analysis. Int. J. Adv. Res. Innov. Ideas Educ. 2019, 5, 236–245. [Google Scholar]
- Hatano, T.; Yasuhara, T.; Yoshihara, R.; Agata, I.; Noro, T.; Okuda, T. Effects of interaction of tannins with co-existing substances. VII. Inhibitory effects of tannins and related polyphenols on xanthine oxidase. Chem. Pharm. Bull. 1990, 38, 1224–1229. [Google Scholar] [CrossRef]
- Garrat, D.C. The Quantitative Analysis of Drugs Japan, 3rd ed.; Chapman and Hall: Tokyo, Japan, 1964; pp. 456–458. [Google Scholar]
- Liu, F.; Ooi, V.E.; Chang, S.T. Free radical scavenging activities of mushroom polysaccharide extracts. Life Sci. 1997, 60, 763–771. [Google Scholar] [CrossRef]
- Chao, P.C.; Li, Y.; Chang, C.H.; Shieh, J.P.; Cheng, J.T.; Cheng, K.C. Investigation of insulin resistance in the popularly used four rat models of type-2 diabetes. Biomed. Pharmacother. 2018, 101, 155–161. [Google Scholar] [CrossRef]
- Matthews, D.R.; Hosker, J.P.; Rudenski, A.S.; Naylor, B.A.; Treacher, D.F.; Turner, R.C. Homeostasis model assessment: Insulin resistance and beta-cell function from fasting plasma glucose and insulin concentrations in man. Diabetologia 1985, 28, 412–419. [Google Scholar] [CrossRef] [Green Version]
- Hassid, W.Z.; Abraham, S. Determination of glycogen with anthrone reagent. Methods Enzym. 1975, 3, 34–37. [Google Scholar]
- Prasad, M.; Jayaraman, S.; Rajagopal, P.; Veeraraghavan, V.P.; Kumar, P.K.; Piramanayagam, S.; Pari, L. Diosgenin inhibits ER stress-induced inflammation in aorta via iRhom2/TACE mediated signaling in experimental diabetic rats: An in vivo and in silico approach. Chem. Biol. Interact. 2022, 358, 109885. [Google Scholar] [CrossRef]
- Liu, Y.; Wan, Q.; Guan, Q.; Gao, L.; Zhao, J. High-fat diet feeding impairs both the expression and activity of AMPKa in rats’ skeletal muscle. Biochem. Biophys. Res. Commun. 2006, 339, 701–707. [Google Scholar] [CrossRef]
- Gonzalez, C.; Alonso, A.; Fernandez, R.; Patterson, A.M. Regulation of insulin receptor substrate-1 in the liver, skeletal muscle and adipose tissue of rats throughout pregnancy. Gynecol. Endocrinol. 2003, 17, 187–197. [Google Scholar]
- Gabe, M. Techniques Histologiques, 6th ed.; Massie e Cie: Paris, France, 1968; 1113p. [Google Scholar]
- Lipinski, C.A.; Lombardo, F.; Dominy, B.W.; Feeney, P.J. Experimental and computational approaches to estimate solubility and permeability in drug discovery and development settings. Adv. Drug Deliv. Rev. 1997, 23, 3–25. [Google Scholar] [CrossRef]
- BIOVIA. Discovery Studio Visualizer; BIOVIA: San Diego, CA, USA, 2012. [Google Scholar]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Žídek, A.; Potapenko, A.; et al. Highly accurate protein structure prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef]
- Christy, J.; Shankari, S.; Majeed, I.; Anand, D.A. Deciphering the Synergistic Mechanism of Cortistatin towards Cancer Targets using Network Pharmacology Approach. Indian J. Pharm. Educ. Res. 2021, 55, 1017–1027. [Google Scholar] [CrossRef]
- Christy, J.; Harini; Vasudevan, S.; Lingesan, P.; Anand, D.A. Deciphering the molecular interplay between pelvic inflammatory disease (PID) and ovarian cancer (OC)—A network biology approach. Gene Rep. 2021, 25, 101405. [Google Scholar] [CrossRef]
- Schüttelkopf, A.W.; van Aalten, D.M. PRODRG: A tool for high-throughput crystallography of protein-ligand complexes. Acta Crystallogr. Sect. D Biol. Crystallogr. 2004, 60, 1355–1363. [Google Scholar] [CrossRef]
- Galeazzi, R. Molecular Dynamics as a Tool in Rational Drug Design: Current Status and Some Major Applications. Curr. Comput. -Aided Drug Des. 2009, 5, 225–240. [Google Scholar] [CrossRef]
- Maragakis, P.; Larsen, K.L.; Eastwood, M.P.; Dror, R.O.; Klepeiss, J.L.; Arkin, I.T.; Jensen, M.; Xu, H.; Trbovic, N.; Friesner, R.A.; et al. Microsecond molecular dynamics simulation shows elect of slow loop dynamics on backbone amide order parameters of proteins. J. Phys. Chem. B 2008, 112, 6155–6158. [Google Scholar] [CrossRef] [Green Version]
- Kansal, N.; Silakari, O.; Ravikumar, M. Three dimensional pharmacophore modelling for c-Kit receptor tyrosine kinase inhibitors. Eur. J. Med. Chem. 2010, 45, 393–404. [Google Scholar] [CrossRef]
- Rizvi, S.I.; Matteucci, E.; Atukeren, P. Traditional medicine in management of type 2 diabetes mellitus. J. Diabetes Res. 2013, 2013, 580823. [Google Scholar] [CrossRef]
- Ghani, M.A.A.; DeFronzo, R.A. Pathogenesis of insulin resistance in skeletal muscle. J. Biomed. Biotechnol. 2010, 2010, 476279. [Google Scholar]
- Kong, Y.R.; Jong, Y.X.; Balakrishnan, M.; Bok, Z.K.; Weng, J.K.K.; Tay, K.C.; Goh, B.H.; Ong, Y.S.; Chan, K.G.; Lee, L.H.; et al. Beneficial Role of Carica papaya Extracts and Phytochemicals on Oxidative Stress and Related Diseases: A Mini Review. Biology 2021, 10, 287. [Google Scholar] [CrossRef]
- Rojop, I.E.J.; Zarate, C.A.T.; Domínguez, D.E.A.; Fuente, L.F.R.; Lobato, C.; Ble, J.; Meraz, M.L.L.; Zagoya, J.C.D.; Bermúdez, D. Phytochemical screening and hypoglycemic activity of Carica papaya leaf in streptozotocin-induced diabetic rats. Rev. Bras. De Farmacogn. 2014, 24, 341–347. [Google Scholar] [CrossRef] [Green Version]
- Ilham, R.; Lelo, A.; Harahap, U.; Widyawati, T.; Siahaan, L. The Effectivity of Ethanolic Extract from Papaya Leaves (Carica papaya L.) as an Alternative Larvacide to Aedes spp. Open Access Maced. J. Med. Sci. 2019, 7, 3395–3399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adachukwu, I.P.; Ogbonna, A.O.; Faith, E.U. Phytochemical analysis of Paw-paw (Carica papaya) leaves. Int. J. Life Sci. Biotechnol. Pharma Res. 2013, 2, 347–351. [Google Scholar]
- Iordăanescu, O.A.; Băla, M.; Gligor, D.; Zippenfening, S.E.; Cugerean, M.I.; Petroman, M.I.; Hădărugă, D.I.; Hădărugă, N.G.; Rivis, M. A DPPH_ Kinetic Approach on the Antioxidant Activity of Various Parts and Ripening Levels of Papaya (Carica papaya L.) Ethanolic Extracts. Plants 2021, 10, 1679. [Google Scholar] [CrossRef]
- Zhou, K.; Wang, H.; Mei, W.; Li, X.; Luo, Y.; Dai, H. Antioxidant activity of papaya seed extracts. Molecules 2011, 16, 6179–6192. [Google Scholar] [CrossRef] [Green Version]
- Mandal, S.D.; Lalmawizuala, R.; Vabeiryureilai, M.; Kumar, N.S.; Lalnunmawii, E. An Investigation of the Antioxidant Property of Carica papaya Leaf Extracts from Mizoram, Northeast India. J. Bot. 2015, 4, 42–45. [Google Scholar]
- Dwivedi, M.K.; Sonter, S.; Mishra, S.; Patel, D.K.; Singh, P.K. Antioxidant, antibacterial activity, and phytochemical characterization of Carica papaya flowers. Beni-Suef Univ. J. Basic Appl. Sci. 2020, 9, 23–34. [Google Scholar] [CrossRef]
- Gudimella, K.K.; Gedda, G.; Kumar, P.S.; Babu, B.K.; Yamajala, B.; Rao, B.V.; Singh, P.P.; Kumar, D.; Sharma, A. Novel synthesis of fluorescent carbon dots from bio-based Carica Papaya Leaves: Optical and structural properties with antioxidant and anti-inflammatory activities. Environ. Res. 2022, 204, 111854. [Google Scholar] [CrossRef]
- Srikanth, G.; Babu, S.M.; Kavitha, C.N.; Rao, M.E.; Vijaykumar, N.S.; Pradeep, C. Studies on in-vitro antioxidant activities of Carica papaya aqueous leaf extract. Res. J. Pharm. Biol. Chem. Sci. 2010, 1, 59–65. [Google Scholar]
- Wijesooriya, A.A.; Deraniyagala, S.A.; Hettiarachchi, C.M. Antioxidant, Anti-Inflammatory and Antibacterial Activities of the Seeds of A Sri Lankan Variety of Carica Papaya. Biomed. Pharmacol. J. 2019, 12, 539–547. [Google Scholar] [CrossRef]
- Raaman, N. Thin Layer Chromatographic Analysis and Antioxidant Activities of Methanol Extract of Leaves of Carica papaya L. Int. J. Adv. Pharm. Biol. Sci. 2015, 4, 414–423. [Google Scholar]
- Nariya, A.; Jhala, D. Pharmacognostic study of Carica papaya leaf extract as inhibitors of reactive oxygen species. Int. Res. J. Pharm. 2017, 8, 13–17. [Google Scholar] [CrossRef]
- Michael, P. Insulin action and resistance in obesity and type 2 diabetes. Czech Nat. Med. 2017, 23, 804–814. [Google Scholar]
- Kahn, S.E. The importance of E-cell failure in the development and progression of type 2 diabetes. J. Clin. Endocrinol. Metab. 2001, 86, 4047–4058. [Google Scholar]
- Prasad, M.; Jayaraman, S.; Eladl, M.A.; El-Sherbiny, M.; Abdelrahman, M.A.E.; Veeraraghavan, V.P.; Vengadassalapathy, S.; Umapathy, V.R.; Hussain, S.F.J.; Krishnamoorthy, K.; et al. A Comprehensive Review on Therapeutic Perspectives of Phytosterols in Insulin Resistance: A Mechanistic Approach. Molecules 2022, 27, 1595. [Google Scholar] [CrossRef]
- Bergsten, P. Pathophysiology of impaired pulsatile insulin release. Diabetes Metab. Res. Rev. 2000, 16, 179–191. [Google Scholar] [CrossRef]
- Padmanabhan, M.; Arumugam, G. Effect of Persea Americana (avocado) fruit extract on the level of expression of adiponectin and PPAR-γ in rats subjected to experimental hyperlipidemia and obesity. J. Complement. Integr. Med. 2014, 11, 107–119. [Google Scholar] [CrossRef]
- Nammi, S.; Kim, M.S.; Gavande, N.S.; Li, G.Q.; Roufogalis, B.D. Regulation of low-density lipoprotein receptor and 3-hydroxy-3-methylglutaryl coenzyme A reductase expression by Zingiber officinale in the liver of high-fat diet-fed rats. Basic Clin. Pharmacol. Toxicol. 2010, 106, 389–395. [Google Scholar] [CrossRef]
- Krishnan, M.; Babu, S.; Rajagopal, P.; Nazar, S.P.; Chinnaiyan, M.; Jayaraman, S. Effect of β-sitosterol on Insulin Receptor, Glucose Transporter 4 Protein Expression and Glucose Oxidation in the Gastrocnemius Muscle of High Fat Diet Induced Type-2 Diabetic Experimental Rats. Int. J. Pharm. Edu. Res. 2021, 55, 479–491. [Google Scholar] [CrossRef]
- Mohamed, A.H.; Ismail, S.; Lithey, B.A. Clinicopathological Studies on The Effect of Papaya on experimentally induced short term hyperglycemia in rats. Zagazig Vet. J. 2019, 47, 45–56. [Google Scholar]
- Lay, S.L.; Simard, G.; Martinez, M.C. Oxidative stress and metabolic pathologies: From an adipocentric point of view. Oxidative Med. Cell. Longev. 2014, 2014, 908539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, J.; Li, C.; Zhang, P.; Cao, X.; Huang, T.; Bai, Y.; Chen, K. Antidiabetic effect of burdock (Arctium lappa L.) root ethanolic extract on streptozotocin-induced diabetic rats. Afr. J. Biotech. 2012, 11, 9079–9085. [Google Scholar]
- Raffaelli, F.; Nanetti, L.; Montecchiani, G.; Borroni, F.; Salvolini, E.; Faloia, E.; Ferretti, G.; Mazzanti, L.; Vignini, A. In vitro effects of fermented papaya (Carica papaya, L.) on platelets obtained from patients with type 2 diabetes. Nutr. Metab. Cardiovasc. Dis. 2015, 25, 224–229. [Google Scholar] [CrossRef] [PubMed]
- Cross, D.A.; Alessi, D.R.; Cohen, P.; Andjelkovich, M.; Hemmings, B.A. Inhibition of glycogen synthase kinase-3 by insulin mediated by protein kinase B. Nature 1995, 378, 785–789. [Google Scholar] [CrossRef] [PubMed]
- Cicero, A.F.; Fiorito, A.; Panourgia, M.P. Effects of a new soy/beta-sitosterol supplement on plasma lipids in moderately hypercholesterolemic subjects. J. Am. Diet. Assoc. 2002, 102, 1807–1811. [Google Scholar] [CrossRef]
- Erion, D.M.; Shulman, G.I. Diacylglycerol-mediated Insulin resistance. Nat. Med. 2010, 16, 400–402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anandhi, R.; Annadurai, T.; Anitha, T.S.; Muralidharan, A.R.; Najmunnisha, K.; Nachiappan, V.; Thomas, P.A.; Geraldine, P. Antihypercholesterolemic and antioxidative effects of an extract of the oyster mushroom, pleurotusostreatus and its major constituent, chrysin, in triton WR-1339-induced hypercholesterolemic rats. J. Physiol. Biochem. 2013, 69, 313–323. [Google Scholar] [CrossRef]
- Nie, Y.; Ma, R.C.; Chan, J.C.; Xu, H.; Xu, G. Glucose-dependent insulinotropic peptide impairs insulin signaling via inducing adipocyte inflammation in glucose-dependent insulinotropic peptide receptor-overexpressing adipocytes. FASEB J. 2012, 26, 2383–2393. [Google Scholar] [CrossRef]
- Balaji, V.; Williams, A.A.; Sathish, S.; Mayilvanan, C.; Narasimhan, A.; Balasubramanian, K. Antidiabetic potentials of a novel polyherbal preparation formulated according to principles of Siddha system of medicine. J. Evid.-Based Complement. Altern. Med. 2012, 17, 180–190. [Google Scholar] [CrossRef]
- Samir, S.M.; Abbas, A.M.; Safwat, S.M.; Elserougy, H.G. Effect of L-carnitine on diabetes-induced changes of skeletal muscles in rats. J. Basic Clin. Physiol. Pharmacol. 2018, 29, 47–59. [Google Scholar] [CrossRef]
















| S.No | Gene Name | Primer Sequence | Reference |
|---|---|---|---|
| 1 | Rat βactin | Sense primer: 5′- AAG TCC CTC ACC CTC CCA AAA G-3′ Anti-sense primer: 5′- AAG CAA TGC TGT CAC CTT CCC-3′ | [18] |
| 2 | Glut-4 | Sense primer: 5′- GGG CTG TGA GTG AGT GCT TTC-3′ Anti-sense primer: 5′- CAG CGA GGC AAG GCT AGA-3′ | [19] |
| 3 | IR | Sense primer: 5′- GCC ATC CCG AAA GCG AAG ATC-3’ Anti-sense primer: 5′- TCT GGG TCC TGA TTG CAT-3’ | [20] |
| S.No | Phytoconstituents | Ethanolic Extract of C. papaya |
|---|---|---|
| 1 | Phytosterols | − |
| 2 | Triterpenoids | + |
| 3 | Flavonoids | + |
| 4 | Phenols | + |
| 5 | Tannins | + |
| 6 | Alkaloids | + |
| 7 | Saponin | + |
| 8 | Acid | + |
| 9 | Proteins | + |
| 10 | Carbohydrates | − |
| 11 | Glycosides | + |
| Grouping | Body Weight (in g) | FBG (mg/dL) | Serum Insulin (µIU/dL) | HOMA-IR | Lipid Profile (mg/dL) | ||||
|---|---|---|---|---|---|---|---|---|---|
| O Day | 45th Day | TG | TC | LDL-c | HDL-c | ||||
| Control | 183 ± 5.2 | 210 ± 6.7 | 111 ± 9.5 | 0.275 ± 0.025 | 3.7 ± 0.39 | 109 ± 9 | 96 ± 6.5 | 92 ± 7.5 | 121 ± 10.5 |
| Diabetes | 189 ± 6.9 | 310 ± 9.3 a | 213 ± 14 a | 0.470 ± 0.020 a | 8.9 ± 0.71 a | 246 ± 16.5 a | 264 ± 14 a | 194 ± 5.5 a | 46 ± 8.5 a |
| Diab + C. papaya | 170 ± 4.3 | 235 ± 8.0 ab | 155 ± 13 ab | 0.349 ± 0.020 b | 5.3 ± 0.61 ab | 137 ± 6.5 ab | 135 ± 9.5 b | 127 ± 7.5 ab | 92 ± 10 b |
| Diab + Metformin | 194 ± 6.9 | 222 ± 6.9 abc | 130 ± 9 abc | 0.364 ± 0.014 b | 4.6 ± 0.41 abc | 117 ± 5.5 bc | 109 ± 8 b | 109 ± 12.5 b | 106 ± 6.5 b |
| Control + C. papaya | 179 ± 4.9 | 201 ± 9.2 bcd | 117 ± 8.5 bcd | 0.298 ± 0.018 b | 3.9 ± 0.39 bcd | 107 ± 8.5 bc | 104 ± 6 b | 101 ± 12 bc | 135 ± 5.5 b |
| S.No | Compound Name | Lig Score1_Dreiding | Lig Score2_Dreiding | PLP 1 | PLP 2 | JAIN | PMF | Dock Score |
|---|---|---|---|---|---|---|---|---|
| IR | ||||||||
| 1 | Trans-Ferulic Acid | 2.67 | 3.78 | 72.66 | 63.58 | 1.6 | 61.46 | 67.774 |
| 2 | Quercetin | 3.28 | 3.41 | 86.87 | 91.19 | 1.77 | 104.6 | 81.475 |
| 3 | Kaempferol | 2.87 | 3.53 | 81.13 | 85.28 | 1.99 | 99.7 | 76.777 |
| 4 | Rutin | 3.25 | 1.88 | 122.11 | 113.91 | 2.37 | 142.19 | 114.771 |
| 5 | p-coumaric acid | No interaction | ||||||
| 6 | Chlorogenic acid | No interaction | ||||||
| 7 | Protocatechuic acid | No interaction | ||||||
| 8 | Caffeic acid | 2.78 | 3.63 | 62.17 | 54.35 | 0.55 | 57.25 | 59.454 |
| GLUT4 | ||||||||
| 1 | Trans-Ferulic Acid | 3.18 | 4.48 | 78.29 | 77.17 | −0.77 | 32.06 | 74.475 |
| 2 | Quercetin | 2.69 | 4.3 | 100.91 | 104.57 | 1.23 | 56.06 | 94.607 |
| 3 | Kaempferol | 3.24 | 4.28 | 98.43 | 102.65 | 0.81 | 54.23 | 92.185 |
| 4 | Rutin | 4.84 | 3.17 | 131.02 | 143.21 | 3.98 | 116.67 | 116.012 |
| 5 | p-coumaric acid | 2.74 | 4.35 | 68.22 | 64.13 | −0.47 | 32.16 | 65.275 |
| 6 | Chlorogenic acid | 3.42 | 4.48 | 117.78 | 120.88 | −0.31 | 82.04 | 108.434 |
| 7 | Protocatechuic acid | 1.14 | 3.46 | 69.08 | 73.26 | −0.92 | 30.13 | 64.194 |
| 8 | Caffeic acid | 2.87 | 4.53 | 76.01 | 77.06 | −0.91 | 34.51 | 72.9 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Roy, J.R.; Janaki, C.S.; Jayaraman, S.; Periyasamy, V.; Balaji, T.; Vijayamalathi, M.; Veeraraghavan, V.P. Carica papaya Reduces Muscle Insulin Resistance via IR/GLUT4 Mediated Signaling Mechanisms in High Fat Diet and Streptozotocin-Induced Type-2 Diabetic Rats. Antioxidants 2022, 11, 2081. https://doi.org/10.3390/antiox11102081
Roy JR, Janaki CS, Jayaraman S, Periyasamy V, Balaji T, Vijayamalathi M, Veeraraghavan VP. Carica papaya Reduces Muscle Insulin Resistance via IR/GLUT4 Mediated Signaling Mechanisms in High Fat Diet and Streptozotocin-Induced Type-2 Diabetic Rats. Antioxidants. 2022; 11(10):2081. https://doi.org/10.3390/antiox11102081
Chicago/Turabian StyleRoy, Jeane Rebecca, Coimbatore Sadagopan Janaki, Selvaraj Jayaraman, Vijayalakshmi Periyasamy, Thotakura Balaji, Madhavan Vijayamalathi, and Vishnu Priya Veeraraghavan. 2022. "Carica papaya Reduces Muscle Insulin Resistance via IR/GLUT4 Mediated Signaling Mechanisms in High Fat Diet and Streptozotocin-Induced Type-2 Diabetic Rats" Antioxidants 11, no. 10: 2081. https://doi.org/10.3390/antiox11102081