
Structure Prediction and Characterization of Thermostable Aldehyde Dehydrogenase from Newly Isolated Anoxybacillus geothermalis Strain D9

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Culture Conditions
2.2. Nucleotide Sequence Accession Number
2.3. Structure Prediction and Validation of A. geothermalis ALDH
2.4. Cloning and Transformation of A geothemalis ALDH Gene
2.5. Expression and Purification of A geothemalis ALDH
2.6. A. geothermalis ALDH Activity Assay
2.7. Characterization of the Purified A. geothermalis ALDH
2.7.1. Determination of Optimum Temperature and Temperature Stability for A. geothemalis ALDH
2.7.2. Determination of Optimum pH and pH Stability for A. geothermalis ALDH
2.7.3. Determination of the Effects of Metal Ions on A. geothermalis ALDH Stability
2.7.4. Determination of the Effects of Organic Solvents on A. geothermalis ALDH Activity
2.7.5. Determination of the Effects of Substrate on A. geothermalis ALDH Activity
2.7.6. Secondary Structure and Melting Point Estimation Using Circular Dichroism (CD)
3. Results
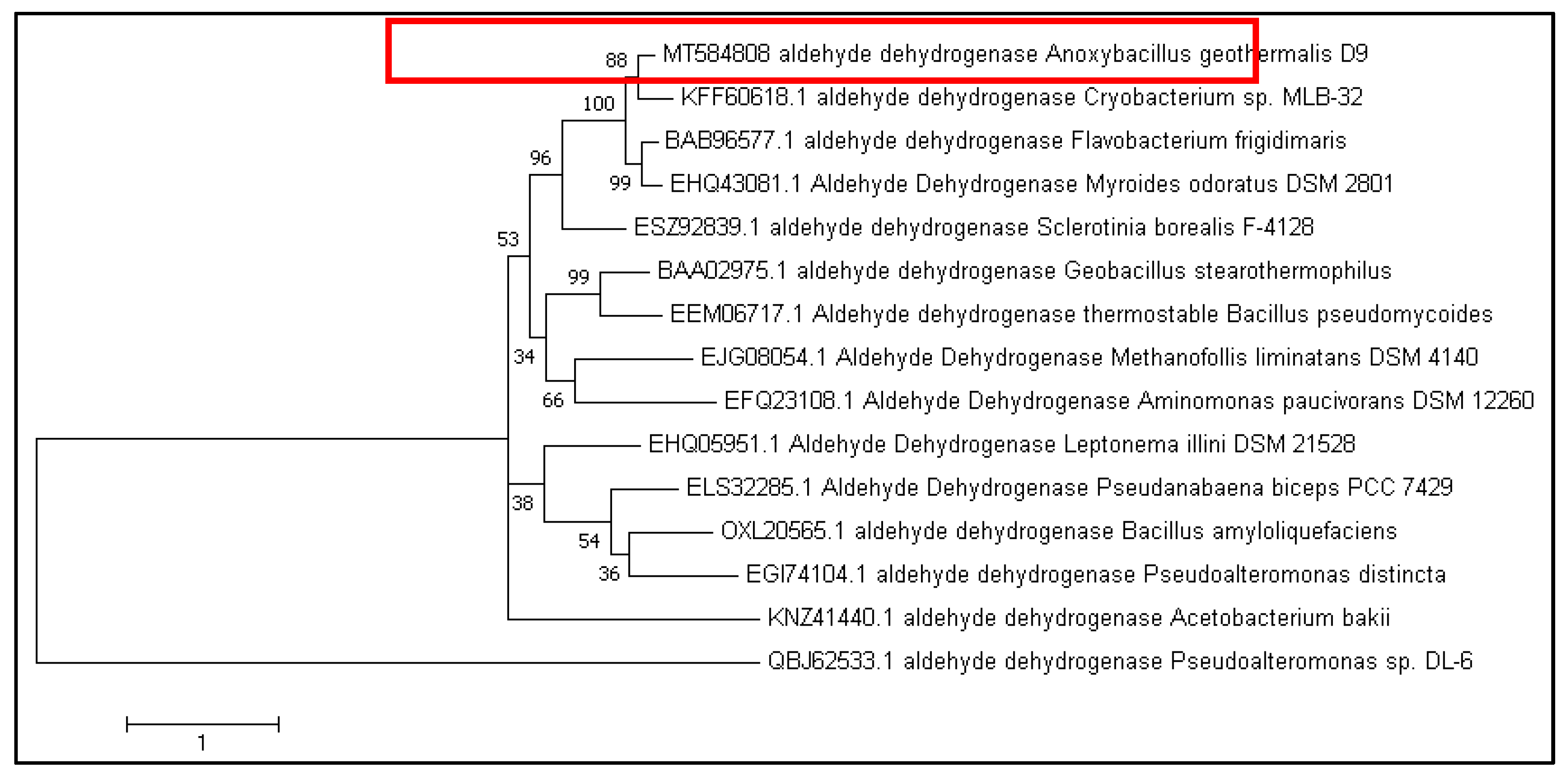
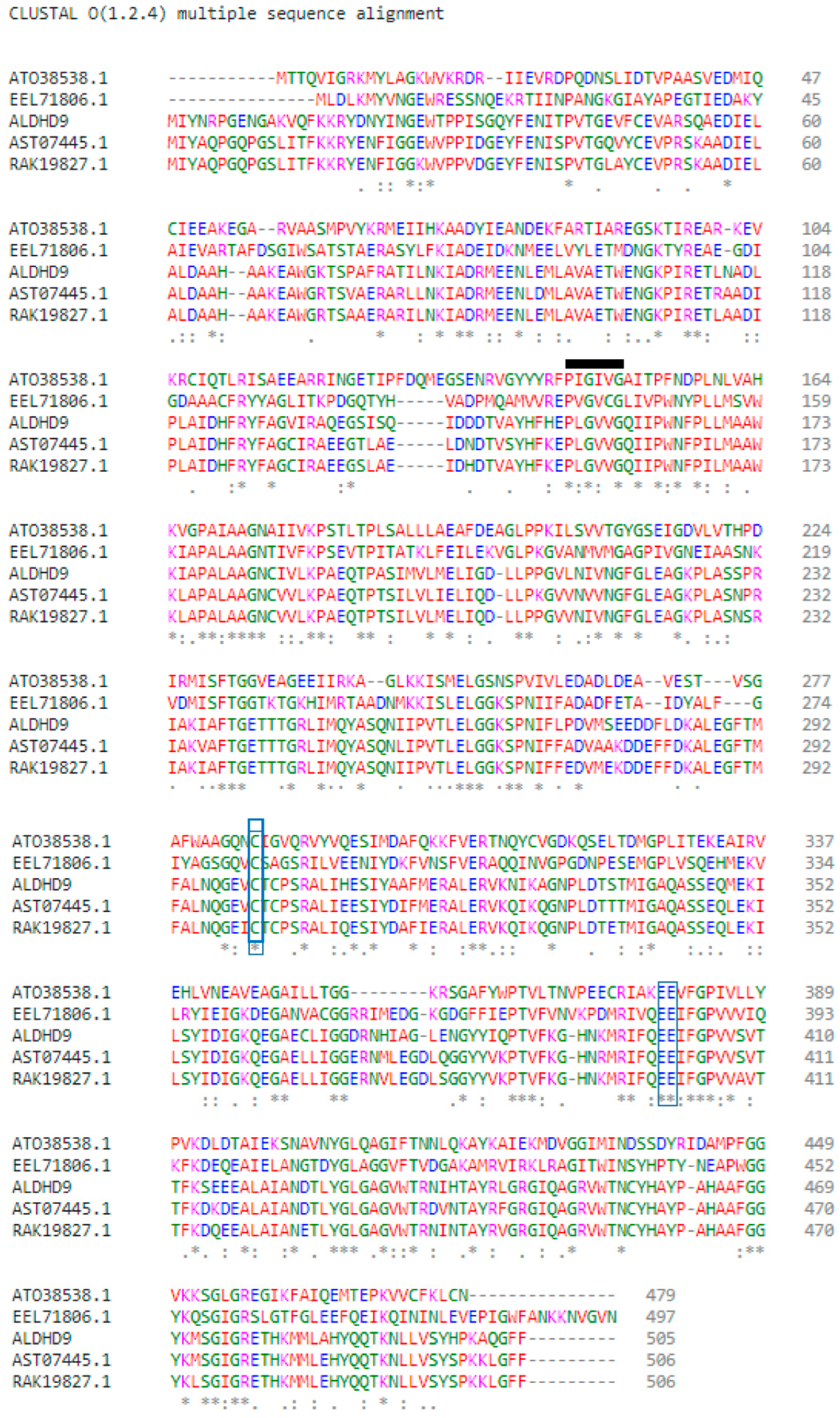
3.1. Sequence Analysis of A. geothermalis ALDH
3.2. Structure Prediction and Validation of A. geothermalis ALDH
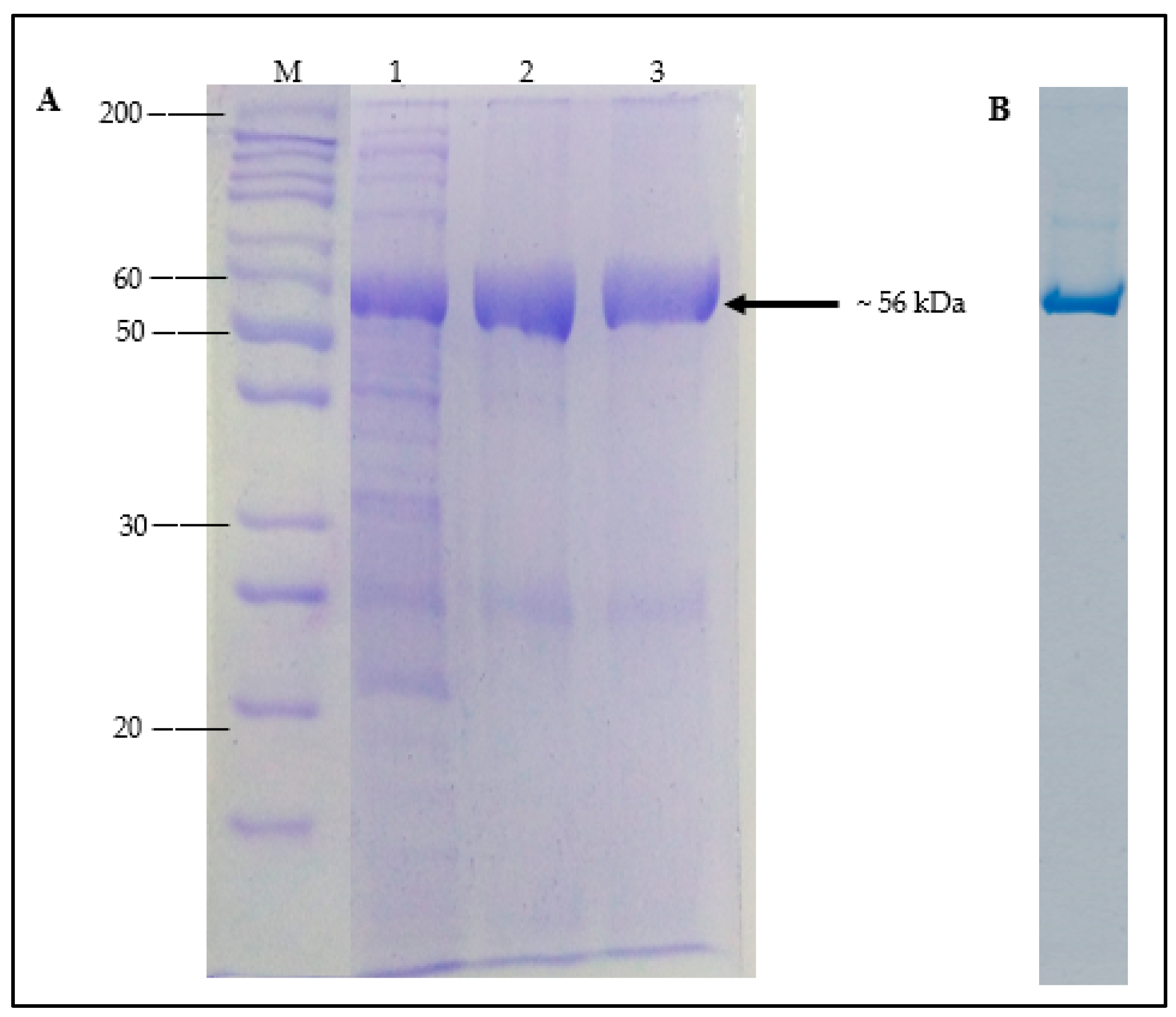
3.3. Expression and Purification of Recombinant A. geothermalis ALDH
3.4. Characterization of Purified A. geothermalis ALDH
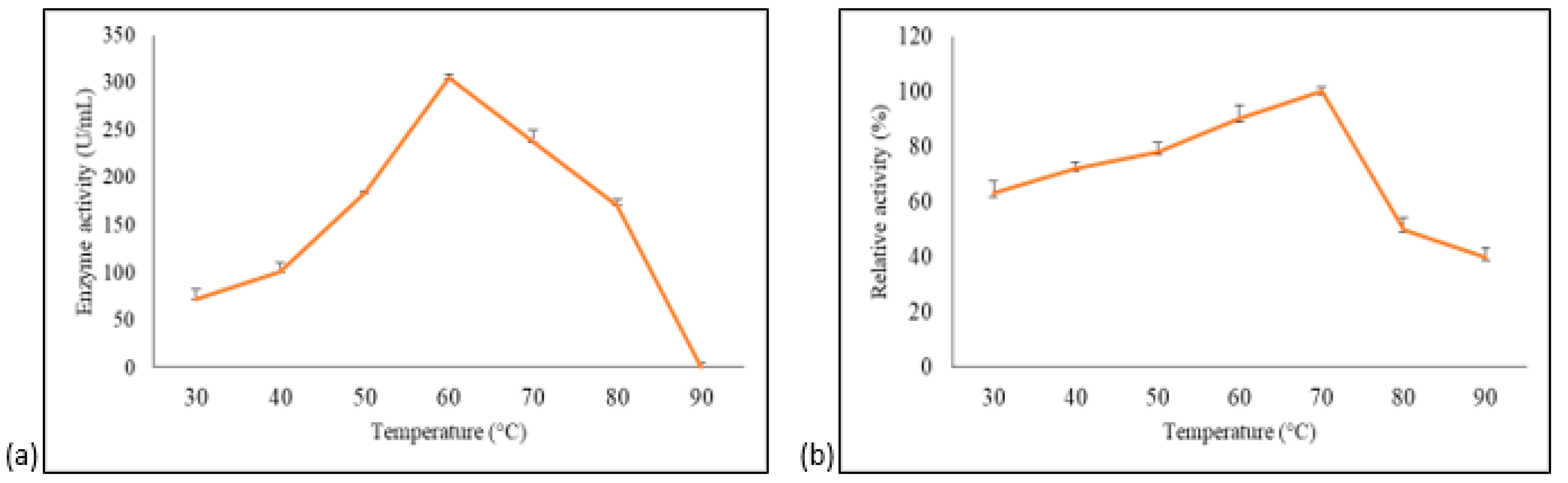
3.4.1. Effect of Temperature on A. geothermalis ALDH Activity and Stability
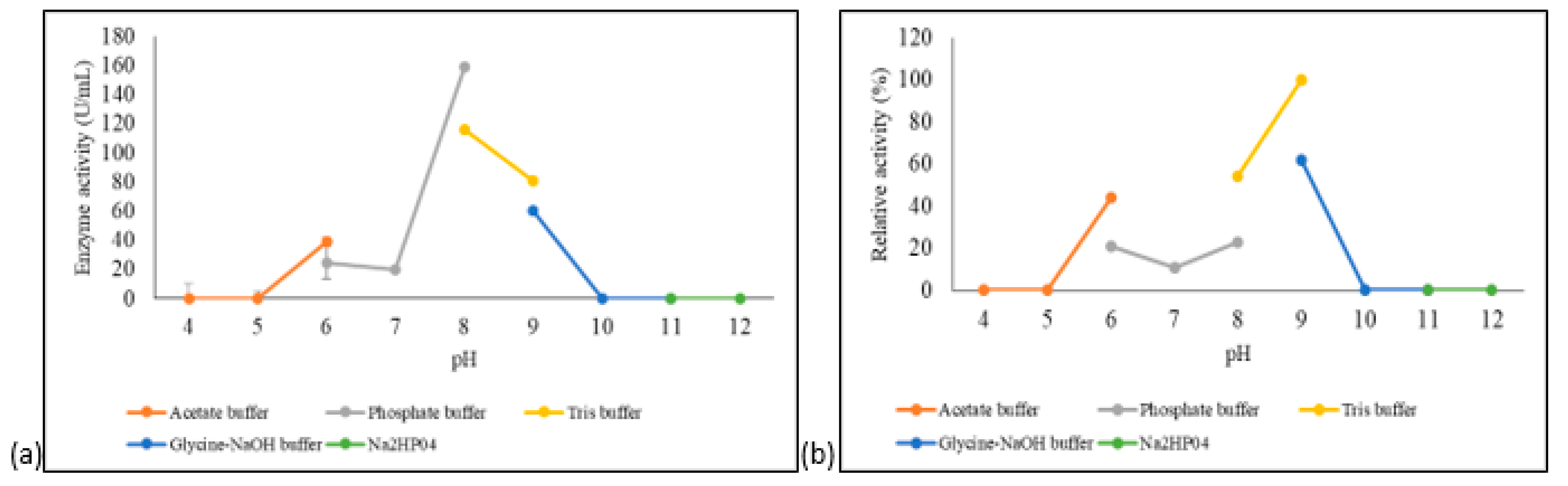
3.4.2. Effect of pH on A. geothermalis ALDH Activity and Stability
3.4.3. Effects of Metal Ions on A. geothermalis ALDH Stability
3.4.4. Effects of Organic Solvents on A. geothermalis ALDH Activity
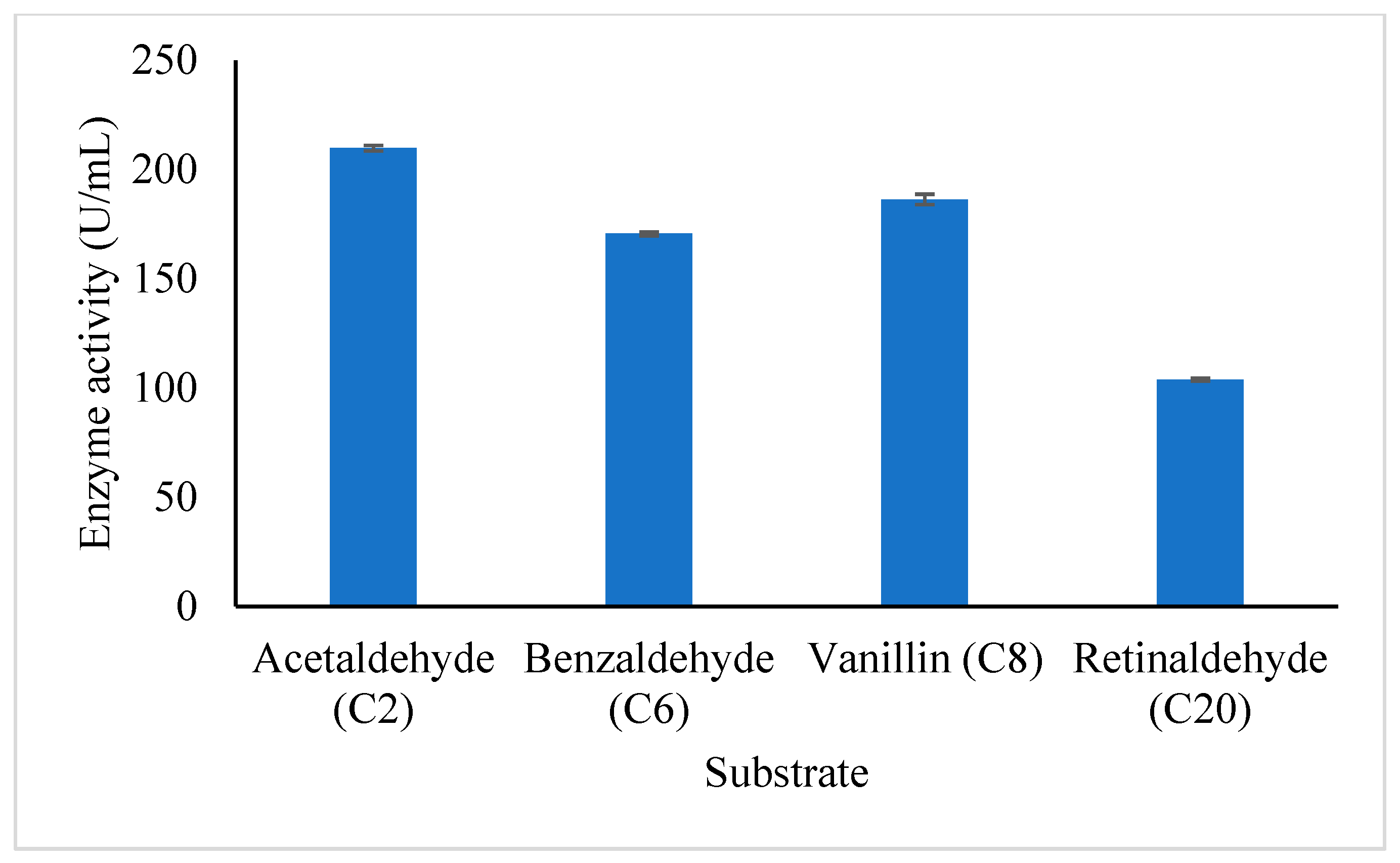
3.4.5. Substrate Specificity of Purified A. geothermalis ALDH
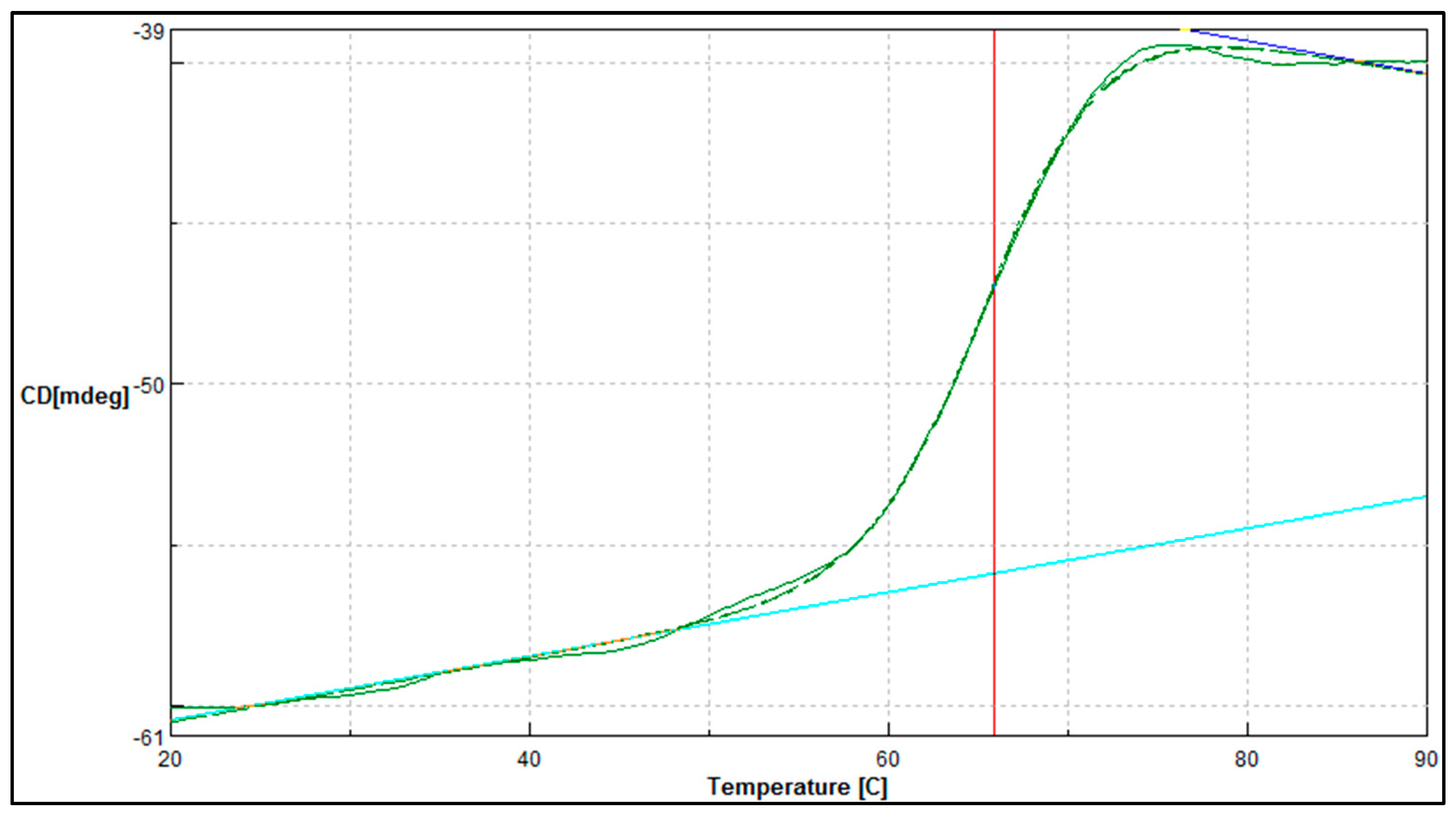
3.4.6. Thermal Denaturation and Secondary Structure Analysis of Purified A. geothermalis ALDH
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Robinson, P.K. Enzymes: Principles and biotechnological applications. Essays Biochem. 2015, 59, 75. [Google Scholar] [CrossRef]
- Haki, G.D.; Rakshit, S.K. Developments in industrially important thermostable enzymes: A review. Bioresour. Technol. 2003, 89, 17–34. [Google Scholar] [CrossRef]
- Kumar, S.; Tsai, C.J.; Nussinov, R. Factors enhancing protein thermostability. Protein Eng. 2000, 13, 179–191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayes, K.; Noor, M.; Djeghader, A.; Armshaw, P.; Pembroke, T.; Tofail, S.; Soulimane, T. The quaternary structure of Thermus thermophilus aldehyde dehydrogenase is stabilized by an evolutionary distinct C-terminal arm extension. Sci. Rep. 2018, 8, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Li, Y.; Wei, D.; Li, P.; Wang, L.; Feng, L. Characterization of a broad-range aldehyde dehydrogenase involved in alkane degradation in Geobacillus thermodenitrificans NG80-2. Microbiol. Res. 2010, 165, 706–712. [Google Scholar] [CrossRef]
- Perozich, J.; Nicholas, H.; Wang, B.C.; Lindahl, R.; Hempel, J. Relationships within the aldehyde dehydrogenase extended family. Protein Sci. 1999, 8, 137–146. [Google Scholar] [CrossRef] [Green Version]
- Stagos, D.; Chen, Y.; Brocker, C.; Donald, E.; Jackson, B.C.; Orlicky, D.J.; Thompson, D.C.; Vasiliou, V. Aldehyde dehydrogenase 1B1: Molecular cloning and characterization of a novel mitochondrial acetaldehyde-metabolizing enzyme. Drug Metab. Dispos. 2010, 38, 1679–1687. [Google Scholar] [CrossRef] [Green Version]
- Ho, K.K.; Weiner, H. Isolation and characterization of an aldehyde dehydrogenase encoded by the aldB gene of Escherichia coli. J. Bacteriol. 2005, 187, 1067–1073. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.J.; Joo, W.A.; Cho, C.W.; Kim, C.W. Halophile Aldehyde Dehydrogenase from Halobacterium salinarium. J. Proteome Res. 2006, 5, 192–195. [Google Scholar] [CrossRef]
- Liu, T.; Hao, L.; Wang, R.; Liu, B. Molecular characterization of a thermostable aldehyde dehydrogenase (ALDH) from the hyperthermophilic archaeon Sulfolobus tokodaii strain 7. Extremophiles 2013, 17, 181–190. [Google Scholar] [CrossRef]
- Yusoff, D.F.; Raja Abd Rahman, R.N.; Masomian, M.; Ali, M.S.; Leow, T.C. Newly isolated alkane hydroxylase and lipase producing geobacillus and anoxybacillus species involved in crude oil degradation. Catalysts 2020, 10, 851. [Google Scholar] [CrossRef]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Overbeek, R.; Olson, R.; Pusch, G.D.; Olsen, G.J.; Davis, J.J.; Disz, T.; Edwards, R.A.; Gerdes, S.; Parrello, B.; Shukla, M.; et al. The SEED and the Rapid Annotation of microbial genomes using Subsystems Technology (RAST). Nucleic Acids Res. 2014, 42, D206–D214. [Google Scholar] [CrossRef]
- Yamanaka, Y.; Kazuoka, T.; Yoshida, M.; Yamanaka, K.; Oikawa, T.; Soda, K. Thermostable aldehyde dehydrogenase from psychrophile, Cytophaga sp. KUC-1: Enzymological characteristics and functional properties. Biochem. Biophys. Res. Commun. 2002, 298, 632–637. [Google Scholar] [CrossRef]
- Tsybovsky, Y.; Krupenko, S.A. Conserved catalytic residues of the ALDH1L1 aldehyde dehydrogenase domain control binding and discharging of the coenzyme. J. Biol. Chem. 2011, 286, 23357–23367. [Google Scholar] [CrossRef] [Green Version]
- Maejima, R.; Iijima, K.; Kaihovaara, P.; Hatta, W.; Koike, T.; Imatani, A.; Salaspuro, M. Effects of ALDH2 genotype, PPI treatment and L-cysteine on carcinogenic acetaldehyde in gastric juice and saliva after intragastric alcohol administration. PLoS ONE 2015, 10, e0120397. [Google Scholar] [CrossRef] [PubMed]
- Shortall, K.; Djeghader, A.; Magner, E.; Soulimane, T. Insights into aldehyde dehydrogenase enzymes: A structural perspective. Front. Mol. Biosci. 2021, 8, 410. [Google Scholar] [CrossRef]
- Numata, K.; Cebe, P.; Kaplan, D.L. Mechanism of enzymatic degradation of beta-sheet crystals. Biomaterials 2010, 31, 2926–2933. [Google Scholar] [CrossRef] [Green Version]
- Daniel, R.M.; Dines, M.; Petach, H.H. The denaturation and degradation of stable enzymes at high temperatures. Biochem. J. 1996, 317, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Meaden, P.G.; Dickinson, F.M.; Mifsud, A.; Tessier, W.; Westwater, J.; Bussey, H.; Midgley, M. The ALD6 gene of Saccharomyces cerevisiae encodes a cytosolic, Mg2+-activated acetaldehyde dehydrogenase. Yeast 1997, 13, 1319–1327. [Google Scholar] [CrossRef]
- Zhao, Y.; Lei, M.; Wu, Y.; Wang, C.; Zhang, Z.; Deng, F.; Wang, H. Molecular cloning and expression of the complete DNA sequence encoding NAD+-dependent acetaldehyde dehydrogenase from Acinetobacter sp. strain HBS-2. Ann. Microbiol. 2009, 59, 97–104. [Google Scholar] [CrossRef]
- Karplus, M.; McCammon, J.A.; Peticolas, W.L. The internal dynamics of globular protein. Crit. Rev. Biochem. 1981, 9, 293–349. [Google Scholar] [CrossRef] [PubMed]
- Shehata, M.; Timucin, E.; Venturini, A.; Sezerman, O.U. Understanding thermal and organic solvent stability of thermoalkalophilic lipases: Insights from computational predictions and experiments. J. Mol. Model. 2020, 26, 122. [Google Scholar] [CrossRef] [PubMed]
- Mohamad, N.R.; Marzuki, N.H.C.; Buang, N.A.; Huyop, F.; Wahab, R.A. An overview of technologies for immobilization of enzymes and surface analysis techniques for immobilized enzymes. Biotechnol. Biotechnol. Equip. 2015, 29, 205–220. [Google Scholar] [CrossRef]
- Masomian, M.; Rahman, R.N.; Salleh, A.B. A Novel Method of Affinity Tag Cleavage in the Purification of a Recombinant Thermostable Lipase from Aneurinibacillus thermoaerophilus Strain HZ. Catalysts 2018, 8, 479. [Google Scholar] [CrossRef] [Green Version]
- Ajsuvakova, O.P.; Tinkov, A.A.; Aschner, M.; Rocha, J.B.; Michalke, B.; Skalnaya, M.G.; Skalny, A.V.; Butnariu, M.; Dadar, M.; Sarac, I.; et al. Sulfhydryl groups as targets of mercury toxicity. Coord. Chem. Rev. 2020, 417, 213343. [Google Scholar] [CrossRef]
- Wang, S.; Meng, X.; Zhou, H.; Liu, Y.; Secundo, F.; Liu, Y. Enzyme stability and activity in non-aqueous reaction systems: A mini review. Catalysts 2016, 6, 32. [Google Scholar] [CrossRef] [Green Version]
- Corrêa, D.H.; Ramos, C.H. The use of circular dichroism spectroscopy to study protein folding, form and function. Afr. J. Biochem. Res. 2009, 3, 164–173. [Google Scholar]
- Singh, K.; Shandilya, M.; Kundu, S.; Kayastha, A.M. Heat, acid and chemically induced unfolding pathways, conformational stability and structure-function relationship in wheat α-amylase. PLoS ONE 2015, 10, e0129203. [Google Scholar] [CrossRef]
- Rogers, D.M.; Jasim, S.B.; Dyer, N.T.; Auvray, F.; Réfrégiers, M.; Hirst, J.D. Electronic circular dichroism spectroscopy of proteins. Chem 2019, 5, 2751–2774. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Validation Tools | Score (%) | |
|---|---|---|
| A | Verify 3D | 96.23 |
| B | Errat | 95.18 |
| C | Ramacandran plot | |
| Most favoured region | 90.4 | |
| Additional allowed region | 9.1 | |
| Generously allowed region | 0.3 | |
| Disallowed region | 0.2 | |
| Purification Step | Fraction Volume (mL) | Protein Content (mg/mL) | Protein Activity (U/mL) | Total Protein (mg) | Total Activity (U) | Specific Activity (U/mg) | Yield (%) | Fold-Purification |
|---|---|---|---|---|---|---|---|---|
| Crude extract | 15 | 10.53 | 717.65 | 157.95 | 7764.75 | 49.18 | 100 | 1 |
| Affinity chromatography | 8 | 2.65 | 368.63 | 21.2 | 2949.04 | 139.11 | 37.98 | 2.83 |
| Metal Ions/Inhibitors | Concentration (mM) | Relative Activity (%) ± SE |
|---|---|---|
| Control | - | 100 |
| Li+ | 1 | 138.83 ± 0.4 |
| 5 | 34.95 ± 0.9 | |
| Na+ | 1 | 74.55 ± 0.3 |
| 5 | 65.05 ± 0.6 | |
| K+ | 1 | 77.67 ± 0.2 |
| 5 | 63.11 ± 0.6 | |
| Ca2+ | 1 | 97.09 ± 0.2 |
| 5 | 72.82 ± 0.3 | |
| Mg2+ | 1 | 205.83 ± 0.8 |
| 5 | 100.98 ± 0.9 | |
| Mn2+ | 1 | 69.9 ± 0.6 |
| 5 | 45.63 ± 0.4 | |
| Fe3+ | 1 | 6.3 ± 0.1 |
| 5 | 4.1 ± 0.5 | |
| Ni2+ | 1 | 63.11 ± 0.5 |
| 5 | 49.69 ± 0.9 | |
| Cu2+ | 1 | 65.04 ± 0.7 |
| 5 | 44.79 ± 0.8 |
| Solvents | Log p | Relative Activity (%) ± SE |
|---|---|---|
| Control | - | 100 |
| DMSO | −1.3 | 55.21 ± 1.8 |
| Methanol | −0.76 | 65. 41 ± 2.7 |
| 1-Propanol | 0.28 | 160.26 ± 3.2 |
| 2-Propanol | 1.2 | 102.00 ± 0.6 |
| Benzene | 2 | 175.11 ± 2.8 |
| Toluene | 2.5 | 25.11 ± 1.2 |
| Octanol | 3 | 82.43 ± 2.3 |
| Xylene | 3.1 | 52.66 ± 3.9 |
| n-Hexane | 3.9 | 55.97 ± 2.8 |
| n-Heptane | 4.66 | 101.26 ± 1.4 |
| n-Tetradecane | 7.2 | 64.47 ± 0.9 |
| Secondary Structure | Amount of Secondary Structure (%) |
|---|---|
| α-helix | 39.3 |
| β-sheet | 21.6 |
| Turn | 19.1 |
| Coil | 20.6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rosli, N.E.; Ali, M.S.M.; Kamarudin, N.H.A.; Masomian, M.; Latip, W.; Saadon, S.; Rahman, R.N.Z.R.A. Structure Prediction and Characterization of Thermostable Aldehyde Dehydrogenase from Newly Isolated Anoxybacillus geothermalis Strain D9. Microorganisms 2022, 10, 1444. https://doi.org/10.3390/microorganisms10071444
Rosli NE, Ali MSM, Kamarudin NHA, Masomian M, Latip W, Saadon S, Rahman RNZRA. Structure Prediction and Characterization of Thermostable Aldehyde Dehydrogenase from Newly Isolated Anoxybacillus geothermalis Strain D9. Microorganisms. 2022; 10(7):1444. https://doi.org/10.3390/microorganisms10071444
Chicago/Turabian StyleRosli, Nur Ezzati, Mohd Shukuri Mohamad Ali, Nor Hafizah Ahmad Kamarudin, Malihe Masomian, Wahhida Latip, Shazleen Saadon, and Raja Noor Zaliha Raja Abd Rahman. 2022. "Structure Prediction and Characterization of Thermostable Aldehyde Dehydrogenase from Newly Isolated Anoxybacillus geothermalis Strain D9" Microorganisms 10, no. 7: 1444. https://doi.org/10.3390/microorganisms10071444