
Structural Studies of Pif1 Helicases from Thermophilic Bacteria


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Protein Expression and Purification
2.2. Crystallization of Pif1 Proteins
2.3. Data Collection, Model Building and Structure Refinement
2.4. Small-Angle X-ray Scattering
2.5. Modeling and Molecular Dynamics Simulations
3. Results
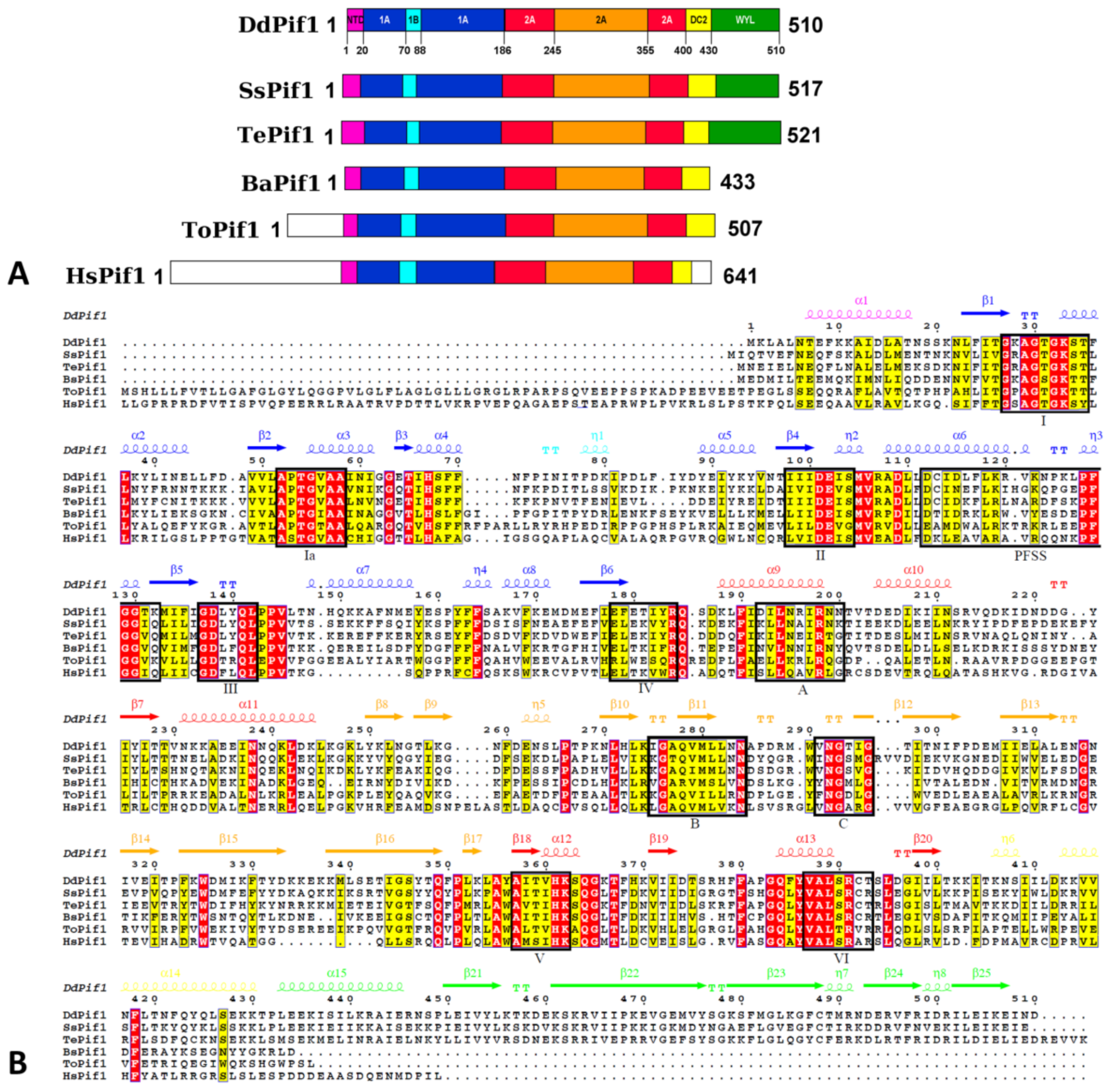
3.1. Structure Determination of DdPif1 and SsPif1
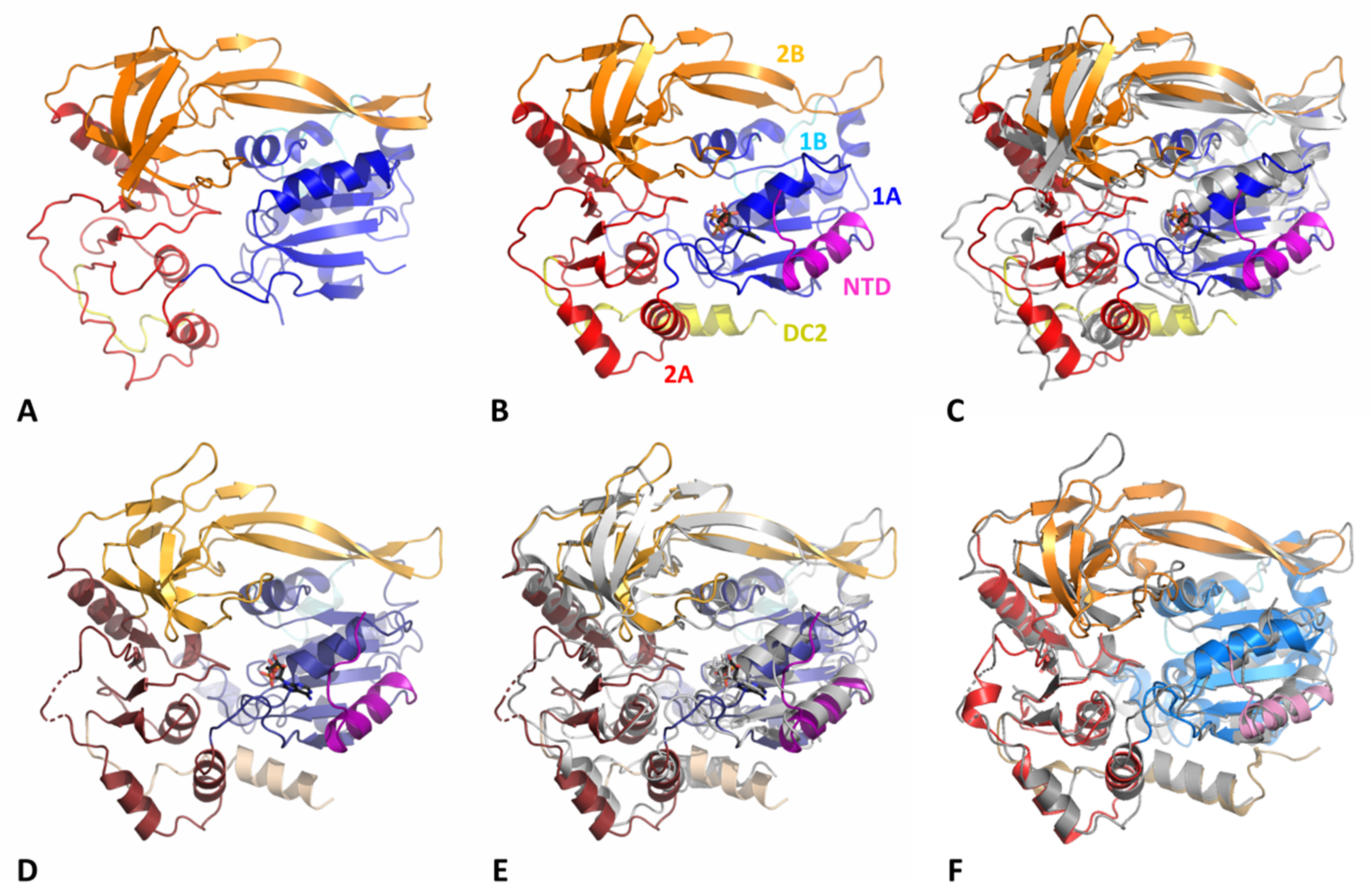
3.2. Overall Structure of DdPif1 and SsPif1
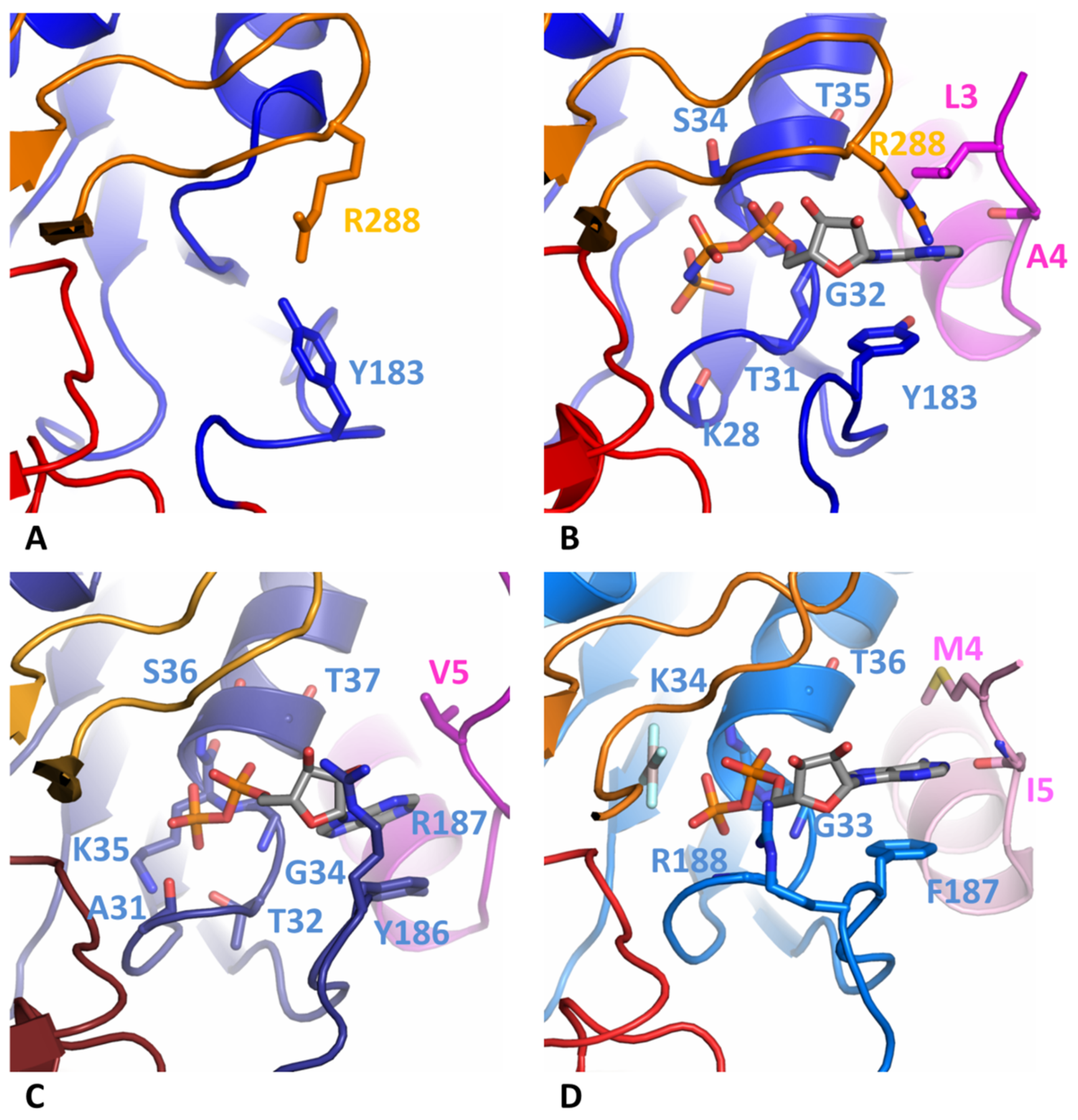
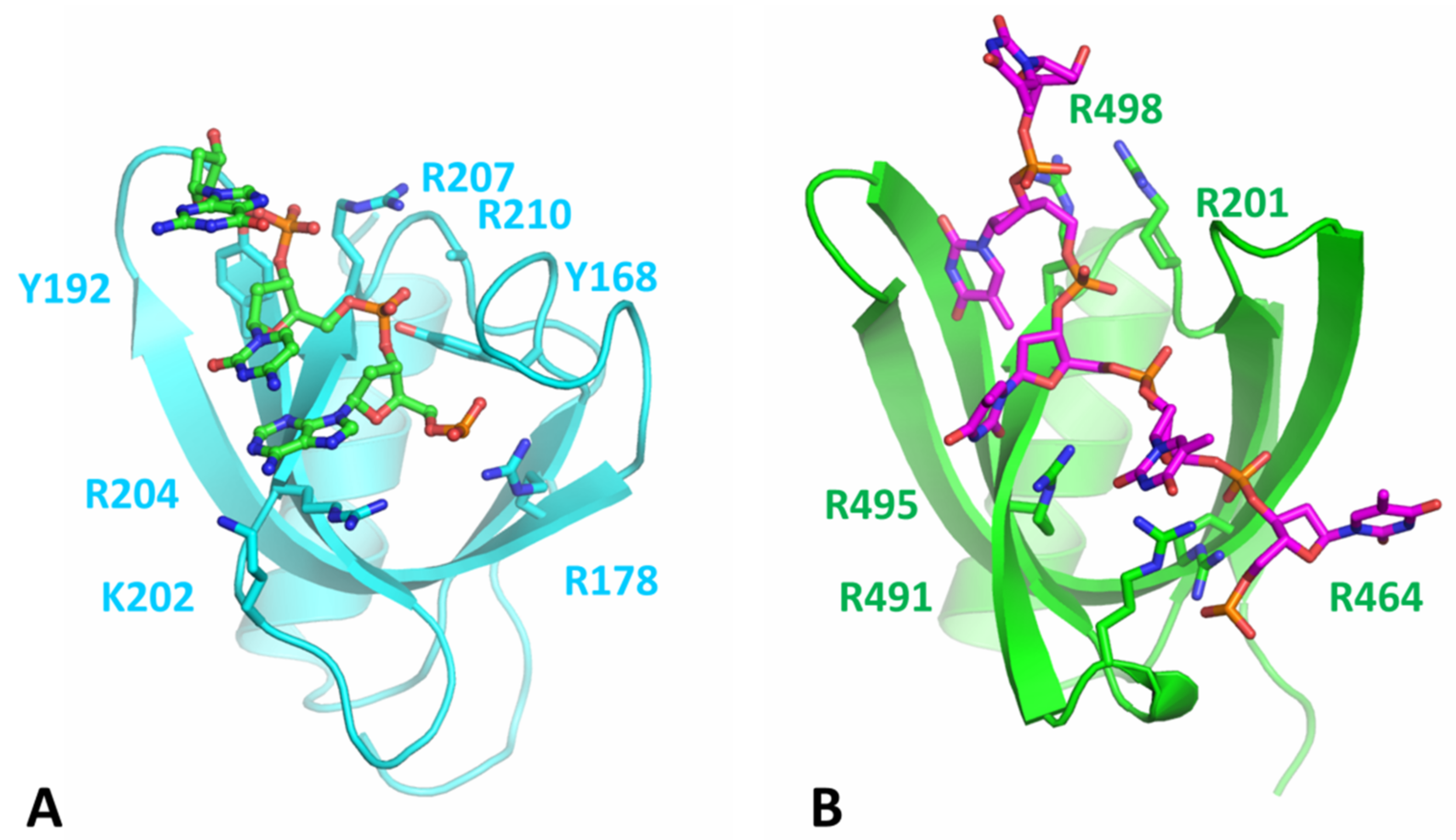
3.3. Comparison of Nucleotide-Binding Site
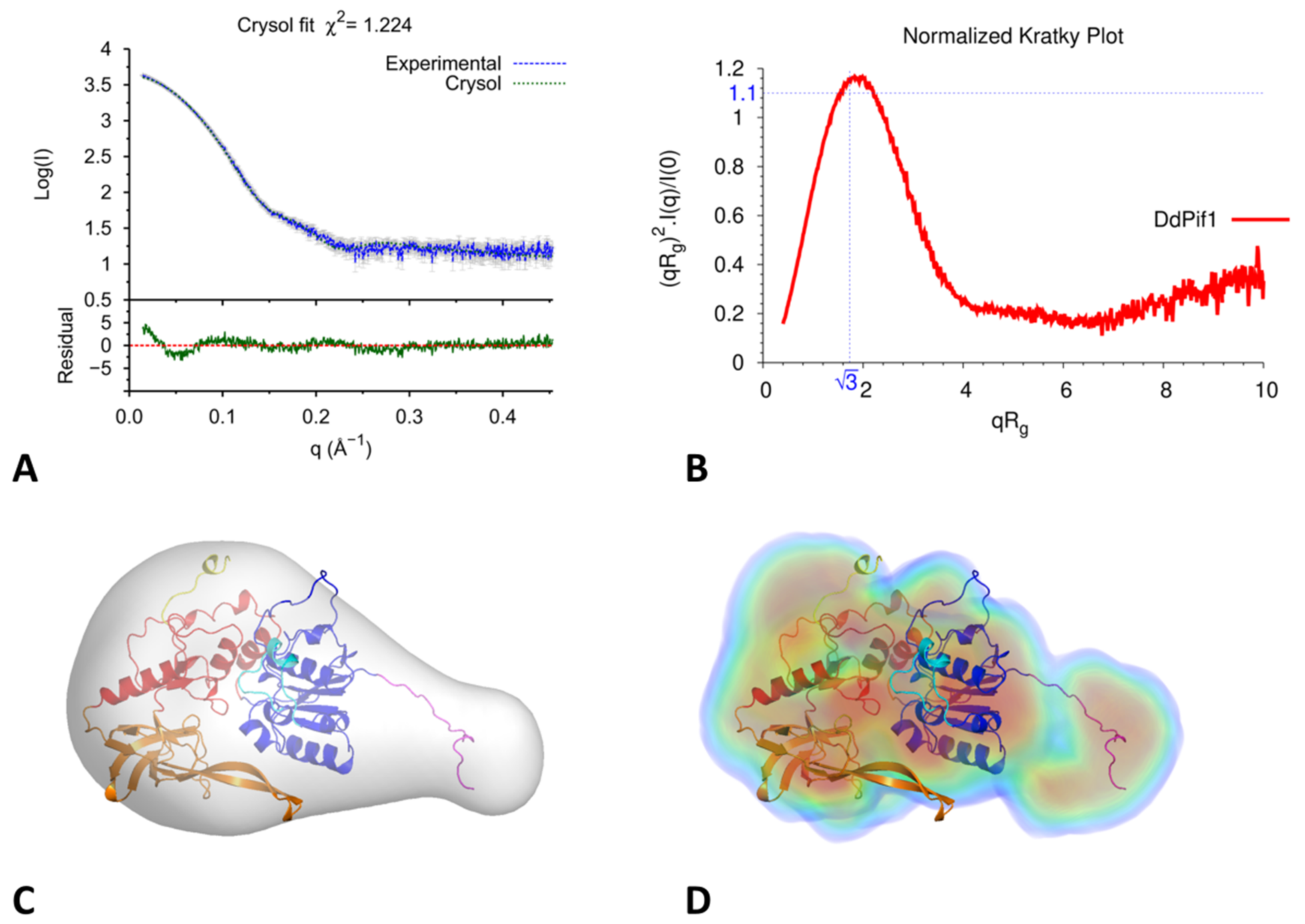
3.4. Analysis of DdPif1-Apo Structure in Solution by SAXS
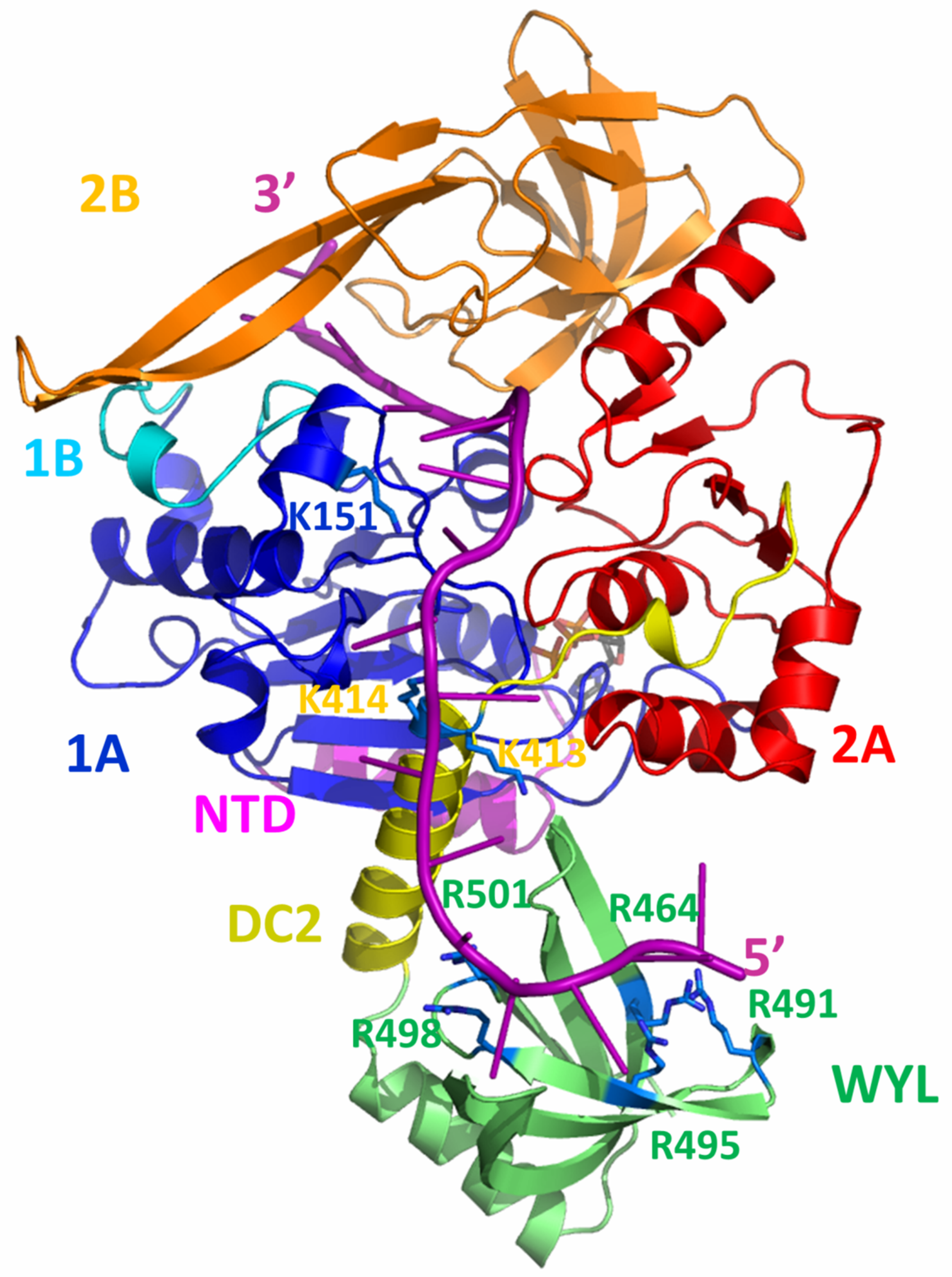
3.5. Modelling of the Structure of the BsPif1-ssDNA Complex
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Singleton, M.R.; Dillingham, M.S.; Wigley, D.B. Structure and mechanism of helicases and nucleic acid translocases. Annu. Rev. Biochem. 2007, 76, 23–50. [Google Scholar] [CrossRef] [PubMed]
- Fairman-Williams, M.E.; Guenther, U.P.; Jankowsky, E. SF1 and SF2 helicases: Family matters. Curr. Opin. Struct. Biol. 2010, 20, 313–324. [Google Scholar] [CrossRef] [PubMed]
- Bochman, M.L.; Sabouri, N.; Zakian, V.A. Unwinding the functions of the Pif1 family helicases. DNA Repair 2010, 9, 237–249. [Google Scholar] [CrossRef] [PubMed]
- Geronimo, C.L.; Zakian, V.A. Getting it done at the ends: Pif1 family DNA helicases and telomeres. DNA Repair 2016, 44, 151–158. [Google Scholar] [CrossRef] [PubMed]
- Byrd, A.K.; Raney, K.D. Structure and function of Pif1 helicase. Biochem. Soc. Trans. 2017, 45, 1159–1171. [Google Scholar] [CrossRef]
- Malone, E.G.; Thompson, M.D.; Byrd, A.K. Role and Regulation of Pif1 Family Helicases at the Replication Fork. Int. J. Mol. Sci. 2022, 23, 3736. [Google Scholar] [CrossRef]
- Varshney, D.; Spiegel, J.; Zyner, K.; Tannahill, D.; Balasubramanian, S. The regulation and functions of DNA and RNA G-quadruplexes. Nat. Rev. Mol. Cell Biol. 2020, 21, 459–474. [Google Scholar] [CrossRef]
- Sauer, M.; Paeschke, K. G-quadruplex unwinding helicases and their function in vivo. Biochem. Soc. Trans. 2017, 45, 1173–1182. [Google Scholar] [CrossRef]
- Paeschke, K.; Bochman, M.L.; Garcia, P.D.; Cejka, P.; Friedman, K.L.; Kowalczykowski, S.C.; Zakian, V.A. Pif1 family helicases suppress genome instability at G-quadruplex motifs. Nature 2013, 497, 458–462. [Google Scholar] [CrossRef]
- Muellner, J.; Schmidt, K.H. Yeast Genome Maintenance by the Multifunctional PIF1 DNA Helicase Family. Genes 2020, 11, 224. [Google Scholar] [CrossRef] [Green Version]
- Sabouri, N. The functions of the multi-tasking Pfh1(Pif1) helicase. Curr. Genet. 2017, 63, 621–626. [Google Scholar] [CrossRef] [PubMed]
- Snow, B.E.; Mateyak, M.; Paderova, J.; Wakeham, A.; Iorio, C.; Zakian, V.; Squire, J.; Harrington, L. Murine Pif1 interacts with telomerase and is dispensable for telomere function in vivo. Mol. Cell. Biol. 2007, 27, 1017–1026. [Google Scholar] [CrossRef] [PubMed]
- Bannwarth, S.; Berg-Alonso, L.; Auge, G.; Fragaki, K.; Kolesar, J.E.; Lespinasse, F.; Lacas-Gervais, S.; Burel-Vandenbos, F.; Villa, E.; Belmonte, F.; et al. Inactivation of Pif1 helicase causes a mitochondrial myopathy in mice. Mitochondrion 2016, 30, 126–137. [Google Scholar] [CrossRef] [PubMed]
- Gagou, M.E.; Ganesh, A.; Thompson, R.; Phear, G.; Sanders, C.; Meuth, M. Suppression of apoptosis by PIF1 helicase in human tumor cells. Cancer Res. 2011, 71, 4998–5008. [Google Scholar] [CrossRef]
- Gagou, M.E.; Ganesh, A.; Phear, G.; Robinson, D.; Petermann, E.; Cox, A.; Meuth, M. Human PIF1 helicase supports DNA replication and cell growth under oncogenic-stress. Oncotarget 2014, 5, 11381–11398. [Google Scholar] [CrossRef]
- Wang, J.; Zhu, X.; Ying, P.; Zhu, Y. PIF1 Affects the Proliferation and Apoptosis of Cervical Cancer Cells by Influencing TERT. Cancer Manag. Res. 2020, 12, 7827–7835. [Google Scholar] [CrossRef]
- Li, S.; Wang, H.; Jehi, S.; Li, J.; Liu, S.; Wang, Z.; Truong, L.; Chiba, T.; Wu, X. PIF1 helicase promotes break-induced replication in mammalian cells. EMBO J. 2021, 40, e104509. [Google Scholar] [CrossRef]
- Kosek, D.; Grabundzija, I.; Lei, H.; Bilic, I.; Wang, H.; Jin, Y.; Peaslee, G.F.; Hickman, A.B.; Dyda, F. The large bat Helitron DNA transposase forms a compact monomeric assembly that buries and protects its covalently bound 5′-transposon end. Mol. Cell 2021, 81, 4271–4286.e4. [Google Scholar] [CrossRef]
- Heringer, P.; Kuhn, G.C.S. Pif1 Helicases and the Evidence for a Prokaryotic Origin of Helitrons. Mol. Biol. Evol. 2022, 39, msab334. [Google Scholar] [CrossRef]
- Chen, W.F.; Dai, Y.X.; Duan, X.L.; Liu, N.N.; Shi, W.; Li, N.; Li, M.; Dou, S.X.; Dong, Y.H.; Rety, S.; et al. Crystal structures of the BsPif1 helicase reveal that a major movement of the 2B SH3 domain is required for DNA unwinding. Nucleic Acids Res. 2016, 44, 2949–2961. [Google Scholar] [CrossRef] [Green Version]
- Zhou, X.; Ren, W.; Bharath, S.R.; Tang, X.; He, Y.; Chen, C.; Liu, Z.; Li, D.; Song, H. Structural and Functional Insights into the Unwinding Mechanism of Bacteroides sp Pif1. Cell Rep. 2016, 14, 2030–2039. [Google Scholar] [CrossRef] [PubMed]
- Dai, Y.X.; Chen, W.F.; Liu, N.N.; Teng, F.Y.; Guo, H.L.; Hou, X.M.; Dou, S.X.; Rety, S.; Xi, X.G. Structural and functional studies of SF1B Pif1 from Thermus oshimai reveal dimerization-induced helicase inhibition. Nucleic Acids Res. 2021, 49, 4129–4143. [Google Scholar] [CrossRef] [PubMed]
- Su, N.; Byrd, A.K.; Bharath, S.R.; Yang, O.; Jia, Y.; Tang, X.; Ha, T.; Raney, K.D.; Song, H. Structural basis for DNA unwinding at forked dsDNA by two coordinating Pif1 helicases. Nat. Commun. 2019, 10, 5375. [Google Scholar] [CrossRef]
- Dai, Y.X.; Guo, H.L.; Liu, N.N.; Chen, W.F.; Ai, X.; Li, H.H.; Sun, B.; Hou, X.M.; Rety, S.; Xi, X.G.; et al. Structural mechanism underpinning Thermus oshimai Pif1-mediated G-quadruplex unfolding. EMBO Rep. 2022, 23, e53874. [Google Scholar] [CrossRef] [PubMed]
- Dehghani-Tafti, S.; Levdikov, V.; Antson, A.A.; Bax, B.; Sanders, C.M. Structural and functional analysis of the nucleotide and DNA binding activities of the human PIF1 helicase. Nucleic Acids Res. 2019, 47, 3208–3222. [Google Scholar] [CrossRef]
- Lu, K.Y.; Chen, W.F.; Rety, S.; Liu, N.N.; Wu, W.Q.; Dai, Y.X.; Li, D.; Ma, H.Y.; Dou, S.X.; Xi, X.G.; et al. Insights into the structural and mechanistic basis of multifunctional S. cerevisiae Pif1p helicase. Nucleic Acids Res. 2018, 46, 1486–1500. [Google Scholar] [CrossRef]
- Lu, K.Y.; Xin, B.G.; Zhang, T.; Liu, N.N.; Li, D.; Rety, S.; Xi, X.G. Structural study of the function of Candida Albicans Pif1. Biochem. Biophys. Res. Commun. 2021, 567, 190–194. [Google Scholar] [CrossRef]
- Bochman, M.L.; Judge, C.P.; Zakian, V.A. The Pif1 family in prokaryotes: What are our helicases doing in your bacteria? Mol. Biol. Cell 2011, 22, 1955–1959. [Google Scholar] [CrossRef]
- Dillingham, M.S.; Kowalczykowski, S.C. RecBCD enzyme and the repair of double-stranded DNA breaks. Microbiol. Mol. Biol. Rev. 2008, 72, 642–671. [Google Scholar] [CrossRef]
- Andis, N.M.; Sausen, C.W.; Alladin, A.; Bochman, M.L. The WYL Domain of the PIF1 Helicase from the Thermophilic Bacterium Thermotoga elfii is an Accessory Single-Stranded DNA Binding Module. Biochemistry 2018, 57, 1108–1118. [Google Scholar] [CrossRef]
- Takaki, Y.; Shimamura, S.; Nakagawa, S.; Fukuhara, Y.; Horikawa, H.; Ankai, A.; Harada, T.; Hosoyama, A.; Oguchi, A.; Fukui, S.; et al. Bacterial lifestyle in a deep-sea hydrothermal vent chimney revealed by the genome sequence of the thermophilic bacterium Deferribacter desulfuricans SSM1. DNA Res. 2010, 17, 123–137. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, S.; Shtaih, Z.; Banta, A.; Beveridge, T.J.; Sako, Y.; Reysenbach, A.L. Sulfurihydrogenibium yellowstonense sp. nov., an extremely thermophilic, facultatively heterotrophic, sulfur-oxidizing bacterium from Yellowstone National Park, and emended descriptions of the genus Sulfurihydrogenibium, Sulfurihydrogenibium subterraneum and Sulfurihydrogenibium azorense. Int. J. Syst. Evol. Microbiol. 2005, 55, 2263–2268. [Google Scholar]
- Vonrhein, C.; Flensburg, C.; Keller, P.; Sharff, A.; Smart, O.; Paciorek, W.; Womack, T.; Bricogne, G. Data processing and analysis with the autoPROC toolbox. Acta Crystallogr. 2011, 67, 293–302. [Google Scholar]
- Kabsch, W. Acta crystallographica. Biol. Crystallogr. 2010, 66, 125–132. [Google Scholar] [CrossRef] [PubMed]
- Evans, P.R.; Murshudov, G.N. How good are my data and what is the resolution? Acta Crystallogr. 2013, 69, 1204–1214. [Google Scholar] [CrossRef]
- Liebschner, D.; Afonine, P.V.; Baker, M.L.; Bunkoczi, G.; Chen, V.B.; Croll, T.I.; Hintze, B.; Hung, L.W.; Jain, S.; McCoy, A.J.; et al. Macromolecular structure determination using X-rays, neutrons and electrons: Recent developments in Phenix. Acta Crystallogr. 2019, 75, 861–877. [Google Scholar] [CrossRef]
- Emsley, P.; Cowtan, K. Coot: Model-building tools for molecular graphics. Acta Crystallogr. 2004, 60, 2126–2132. [Google Scholar] [CrossRef]
- Hopkins, J.B.; Gillilan, R.E.; Skou, S. BioXTAS RAW: Improvements to a free open-source program for small-angle X-ray scattering data reduction and analysis. J. Appl. Crystallogr. 2017, 50, 1545–1553. [Google Scholar] [CrossRef]
- Manalastas-Cantos, K.; Konarev, P.V.; Hajizadeh, N.R.; Kikhney, A.G.; Petoukhov, M.V.; Molodenskiy, D.S.; Panjkovich, A.; Mertens, H.D.T.; Gruzinov, A.; Borges, C.; et al. ATSAS 3.0: Expanded functionality and new tools for small-angle scattering data analysis. J. Appl. Crystallogr. 2021, 54, 343–355. [Google Scholar] [CrossRef]
- Franke, D.; Svergun, D.I. DAMMIF, a program for rapid ab-initio shape determination in small-angle scattering. J. Appl. Crystallogr. 2009, 42, 342–346. [Google Scholar] [CrossRef]
- Grant, T.D. Ab initio electron density determination directly from solution scattering data. Nat. Methods 2018, 15, 191–193. [Google Scholar] [CrossRef]
- Evrard, G.; Mareuil, F.; Bontems, F.; Sizun, C.; Perez, J. DADIMODO: A program for refining the structure of multidomain proteins and complexes against small-angle scattering data and NMR-derived restraints. J. Appl. Crystallogr. 2011, 44, 1264–1271. [Google Scholar] [CrossRef]
- Kozin, M.B.; Svergun, D.I. Automated matching of high- and low-resolution structural models. J. Appl. Crystallogr. 2001, 34, 33–41. [Google Scholar] [CrossRef]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Zidek, A.; Potapenko, A.; et al. Highly accurate protein structure prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef] [PubMed]
- Baek, M.; DiMaio, F.; Anishchenko, I.; Dauparas, J.; Ovchinnikov, S.; Lee, G.R.; Wang, J.; Cong, Q.; Kinch, L.N.; Schaeffer, R.D.; et al. Accurate prediction of protein structures and interactions using a three-track neural network. Science 2021, 373, 871–876. [Google Scholar] [CrossRef] [PubMed]
- Jo, S.; Kim, T.; Iyer, V.G.; Im, W. CHARMM-GUI: A web-based graphical user interface for CHARMM. J. Comput. Chem. 2008, 29, 1859–1865. [Google Scholar] [CrossRef]
- Van der Spoel, D.; Lindahl, E.; Hess, B.; Groenhof, G.; Mark, A.E.; Berendsen, H.J. GROMACS: Fast, flexible, and free. J. Comput. Chem. 2005, 26, 1701–1718. [Google Scholar] [CrossRef] [PubMed]
- Schrodinger, Inc. The PyMOL Molecular Graphics System; Version 1.8.; Schrodinger, Inc.: New York, NY, USA, 2015. [Google Scholar]
- Gouet, P.; Courcelle, E.; Stuart, D.I.; Metoz, F. ESPript: Analysis of multiple sequence alignments in PostScript. Bioinformatics 1999, 15, 305–308. [Google Scholar] [CrossRef] [PubMed]
- Gozzi, K.; Salinas, R.; Nguyen, V.D.; Laub, M.T.; Schumacher, M.A. ssDNA is an allosteric regulator of the C. crescentus SOS-independent DNA damage response transcription activator, DriD. Genes Dev. 2022, 36, 618–633. [Google Scholar] [CrossRef] [PubMed]
- Muller, A.U.; Leibundgut, M.; Ban, N.; Weber-Ban, E. Structure and functional implications of WYL domain-containing bacterial DNA damage response regulator PafBC. Nat. Commun. 2019, 10, 4653. [Google Scholar] [CrossRef]
- Harman, A.; Manna, S. Identification of Pif1 helicases with novel accessory domains in various amoebae. Mol. Phylogenetics Evol. 2016, 103, 64–74. [Google Scholar] [CrossRef]
- Makarova, K.S.; Anantharaman, V.; Grishin, N.V.; Koonin, E.V.; Aravind, L. CARF and WYL domains: Ligand-binding regulators of prokaryotic defense systems. Front. Genet. 2014, 5, 102. [Google Scholar] [CrossRef] [PubMed]
- Blankenchip, C.L.; Nguyen, J.V.; Lau, R.K.; Ye, Q.; Gu, Y.; Corbett, K.D. Control of bacterial immune signaling by a WYL domain transcription factor. Nucleic Acids Res. 2022, 50, 5239–5250. [Google Scholar] [CrossRef] [PubMed]
- Picton, D.M.; Harling-Lee, J.D.; Duffner, S.J.; Went, S.C.; Morgan, R.D.; Hinton, J.C.D.; Blower, T.R. A widespread family of WYL-domain transcriptional regulators co-localizes with diverse phage defence systems and islands. Nucleic Acids Res. 2022, 50, 5191–5207. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Ma, J.; Kumar, V.; Fu, H.; Xu, C.; Wang, S.; Jia, Q.; Fan, Q.; Xi, X.; Li, M.; et al. Identification of flexible Pif1-DNA interactions and their impacts on enzymatic activities. Nucleic Acids Res. 2022, 50, 7002–7012. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.P.; Koc, K.N.; Stodola, J.L.; Galletto, R. A Monomer of Pif1 Unwinds Double-Stranded DNA and It Is Regulated by the Nature of the Non-Translocating Strand at the 3′-End. J. Mol. Biol. 2016, 428, 1053–1067. [Google Scholar] [CrossRef]
- Valle-Orero, J.; Rieu, M.; Tran, P.L.T.; Joubert, A.; Raj, S.; Allemand, J.F.; Croquette, V.; Boule, J.B. Strand switching mechanism of Pif1 helicase induced by its collision with a G-quadruplex embedded in dsDNA. Nucleic Acids Res. 2022, 50, 8767–8778. [Google Scholar] [CrossRef]
- Ding, Y.; Fleming, A.M.; Burrows, C.J. Case studies on potential G-quadruplex-forming sequences from the bacterial orders Deinococcales and Thermales derived from a survey of published genomes. Sci. Rep. 2018, 8, 15679. [Google Scholar] [CrossRef]
- Vannutelli, A.; Perreault, J.P.; Ouangraoua, A. G-quadruplex occurrence and conservation: More than just a question of guanine-cytosine content. NAR Genom. Bioinform. 2022, 4, lqac010. [Google Scholar] [CrossRef]
- Kumar, S.; Tsai, C.J.; Nussinov, R. Factors enhancing protein thermostability. Protein Eng. 2000, 13, 179–191. [Google Scholar] [CrossRef]
- Sterner, R.; Liebl, W. Thermophilic adaptation of proteins. Crit. Rev. Biochem. Mol. Biol. 2001, 36, 39–106. [Google Scholar] [CrossRef] [PubMed]
- Sawle, L.; Ghosh, K. How do thermophilic proteins and proteomes withstand high temperature? Biophys. J. 2011, 101, 217–227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kreil, D.P.; Ouzounis, C.A. Identification of thermophilic species by the amino acid compositions deduced from their genomes. Nucleic Acids Res. 2001, 29, 1608–1615. [Google Scholar] [CrossRef] [PubMed]
- Schermerhorn, K.M.; Tanner, N.; Kelman, Z.; Gardner, A.F. High-temperature single-molecule kinetic analysis of thermophilic archaeal MCM helicases. Nucleic Acids Res. 2016, 44, 8764–8771. [Google Scholar] [CrossRef] [PubMed] [Green Version]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Réty, S.; Zhang, Y.; Fu, W.; Wang, S.; Chen, W.-F.; Xi, X.-G. Structural Studies of Pif1 Helicases from Thermophilic Bacteria. Microorganisms 2023, 11, 479. https://doi.org/10.3390/microorganisms11020479
Réty S, Zhang Y, Fu W, Wang S, Chen W-F, Xi X-G. Structural Studies of Pif1 Helicases from Thermophilic Bacteria. Microorganisms. 2023; 11(2):479. https://doi.org/10.3390/microorganisms11020479
Chicago/Turabian StyleRéty, Stéphane, Yingzi Zhang, Wentong Fu, Shan Wang, Wei-Fei Chen, and Xu-Guang Xi. 2023. "Structural Studies of Pif1 Helicases from Thermophilic Bacteria" Microorganisms 11, no. 2: 479. https://doi.org/10.3390/microorganisms11020479