
Laccase Production from Agrocybe pediades: Purification and Functional Characterization of a Consistent Laccase Isoenzyme in Liquid Culture

 , , , , , and
, , , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Microorganism and Culture Conditions
2.2. Enzyme Activity and Zymogram Assay
2.3. Partial Purification of Laccase
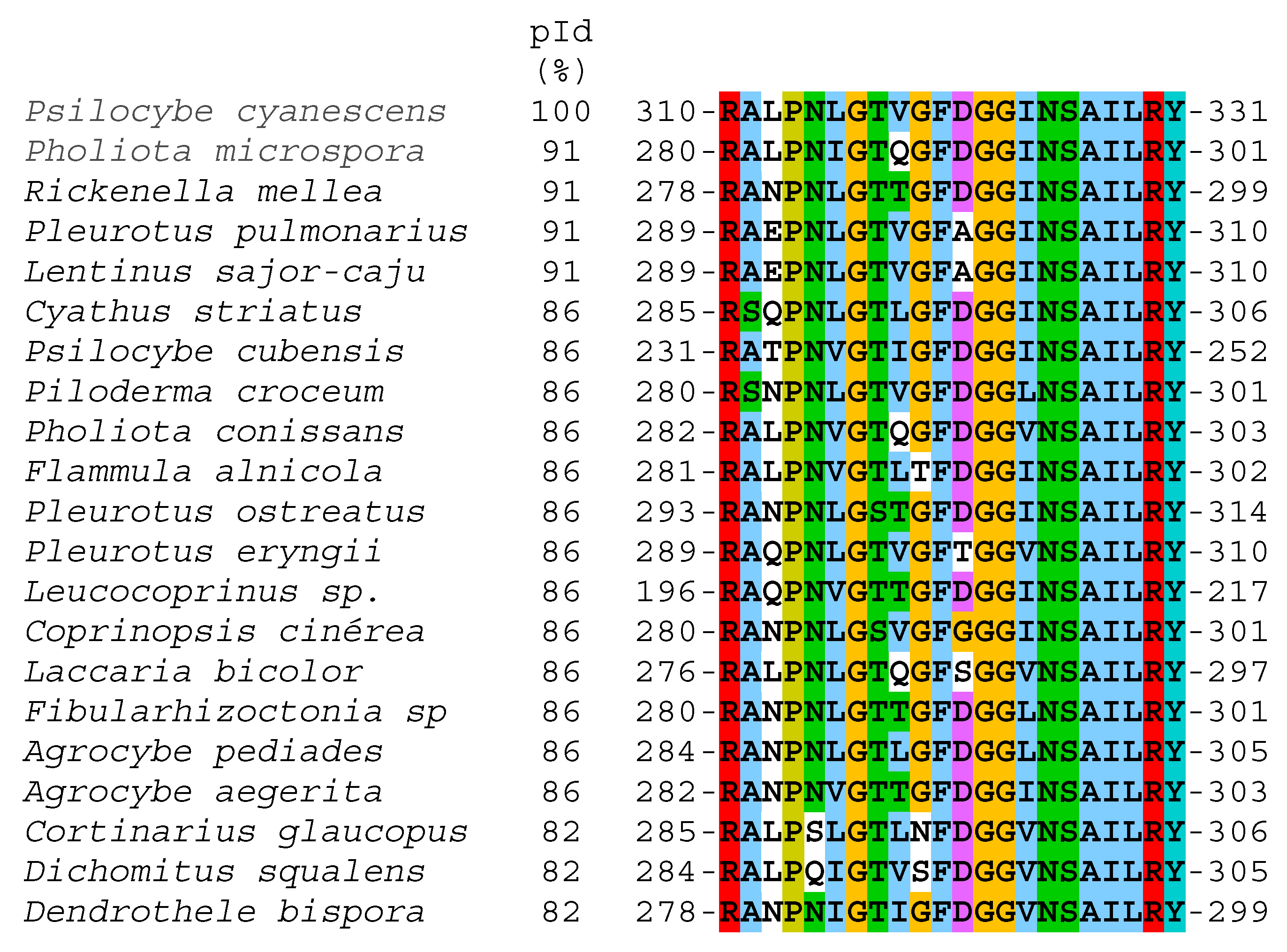
2.4. LC−MS/MS Analysis
2.5. Functional Characterization of Laccase Protein
2.5.1. Determination of Native Status
2.5.2. Effect of pH and Temperature on Laccase Activity
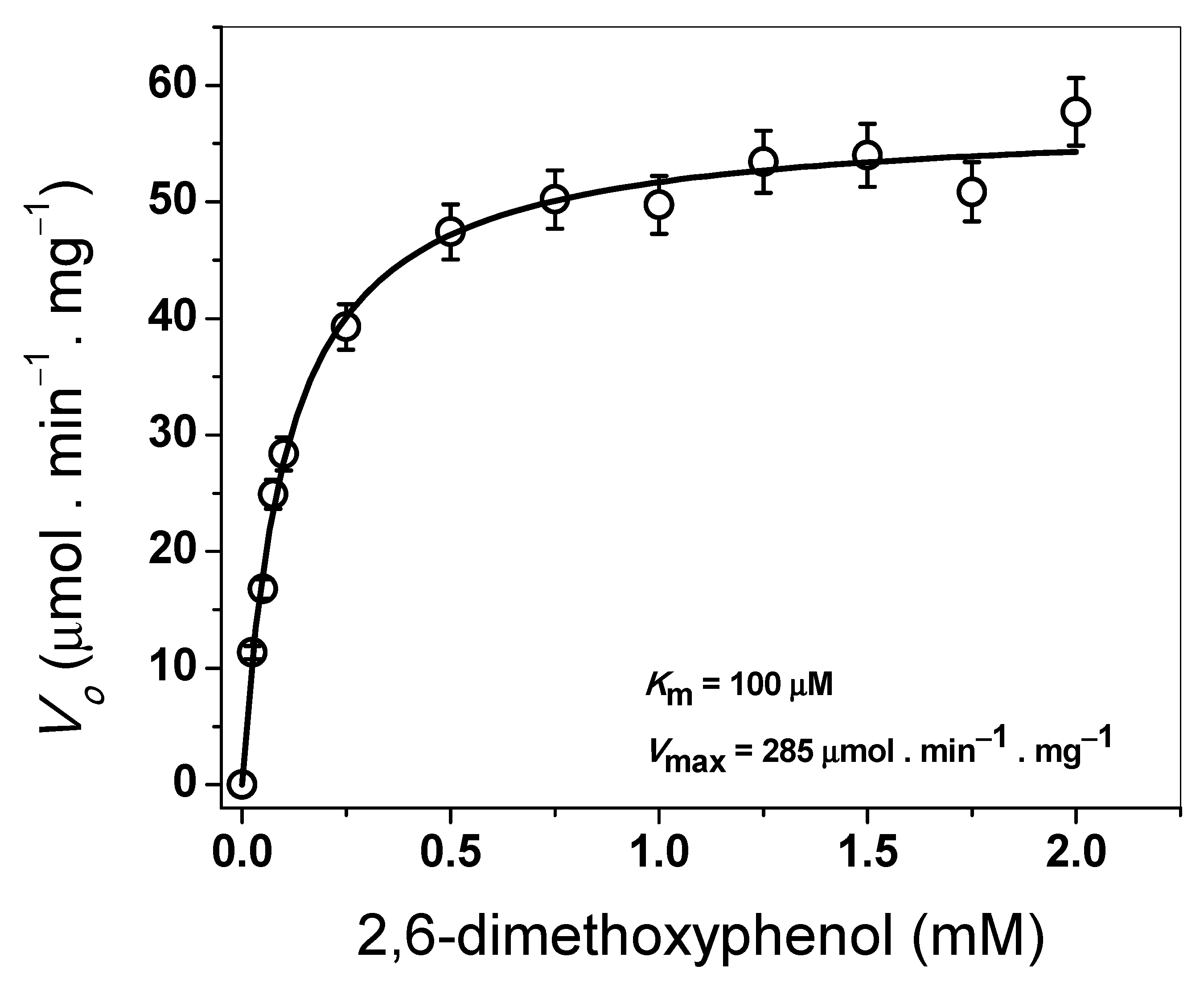
2.5.3. Determination of Steady-State Kinetic Parameters
2.6. Statistical Analysis
3. Results
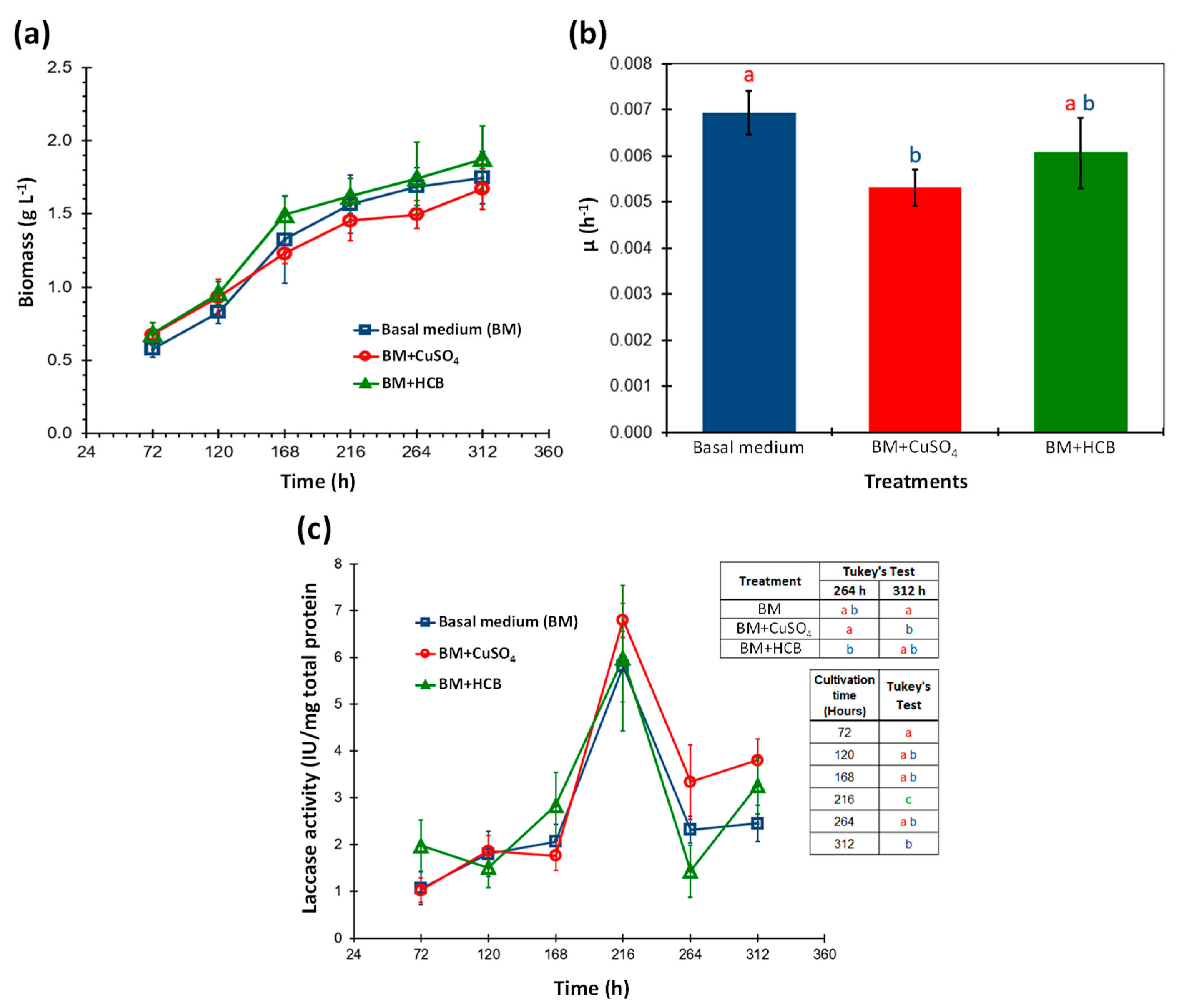
3.1. Effect of CuSO4 and HCB on the Growth and Laccase Activity of Agrocybe pediades
3.2. Analysis of Enzymatic Activity by Zymogram
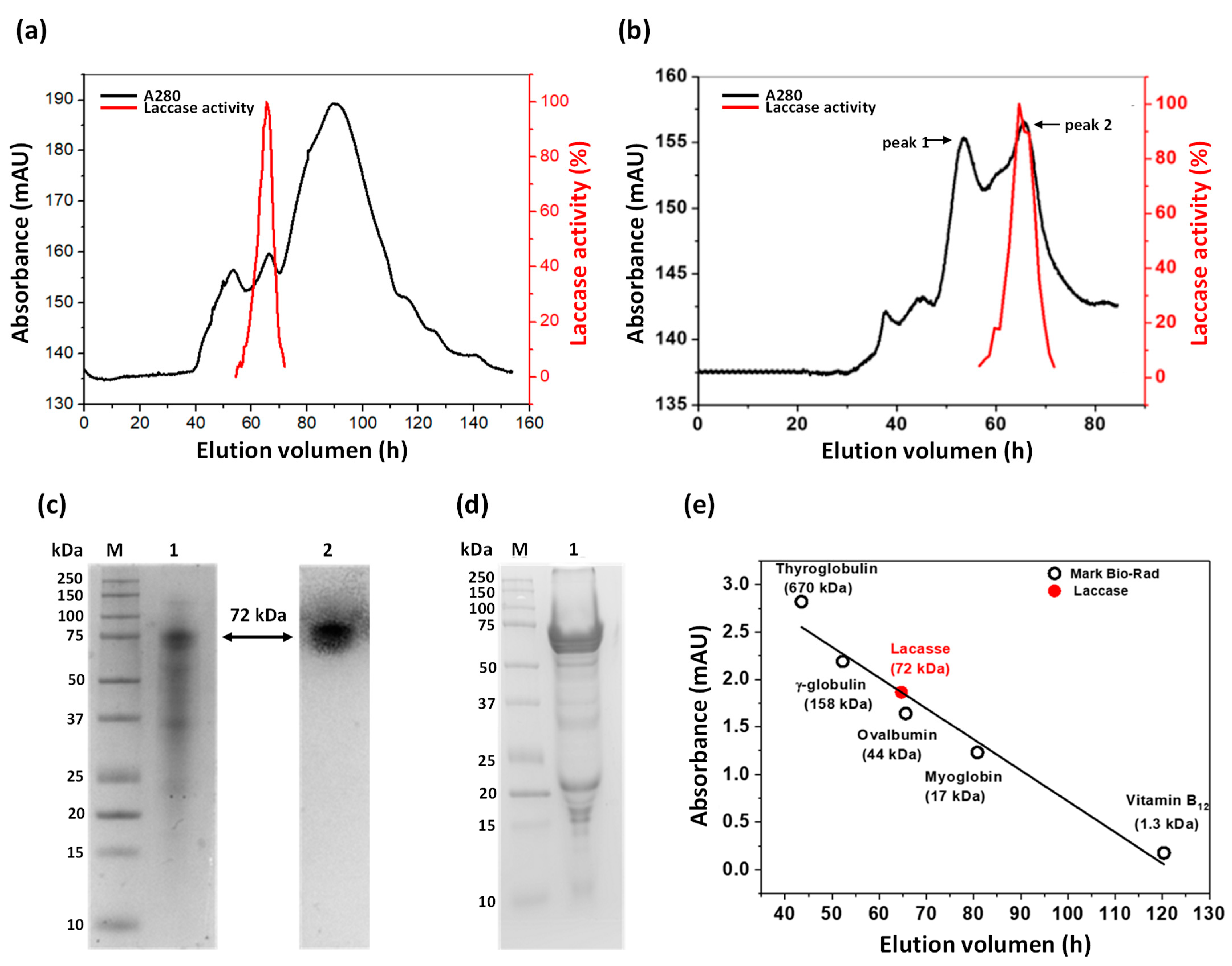
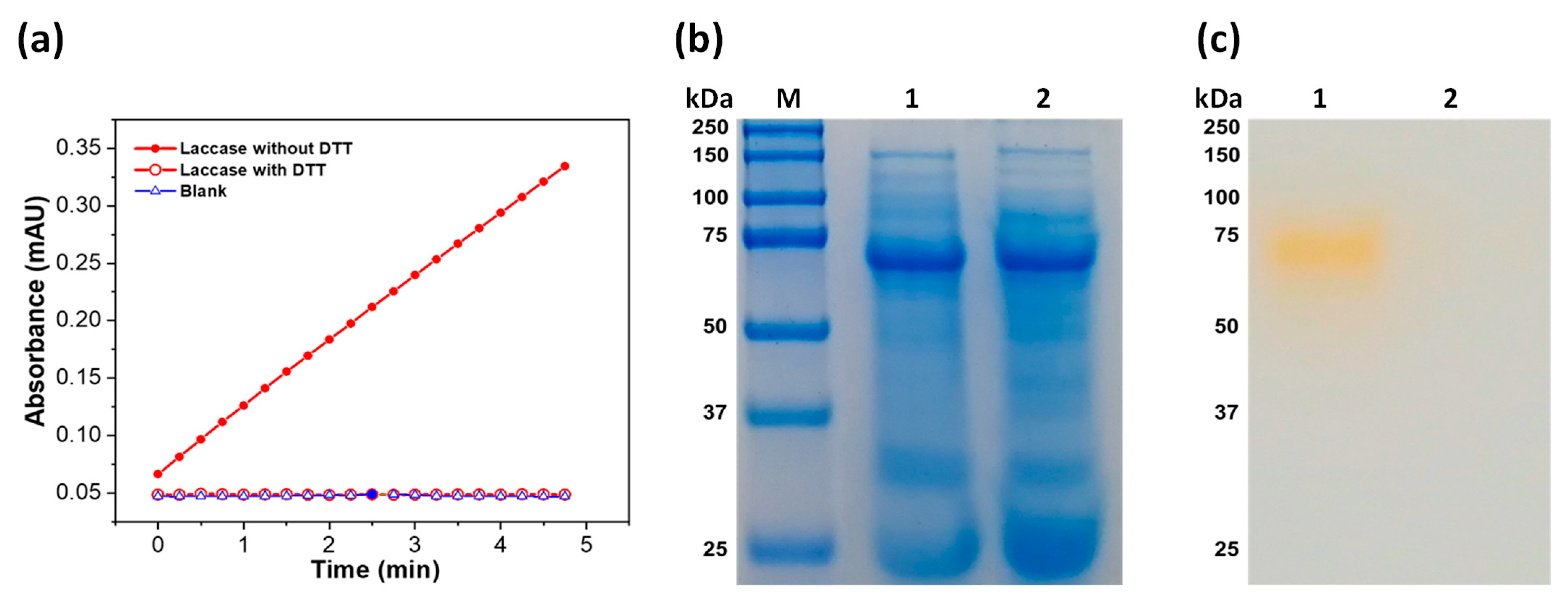
3.3. Partial Purification of Laccase Protein
3.4. Functional Characterization of Laccase Protein
3.4.1. Determination of Native Status
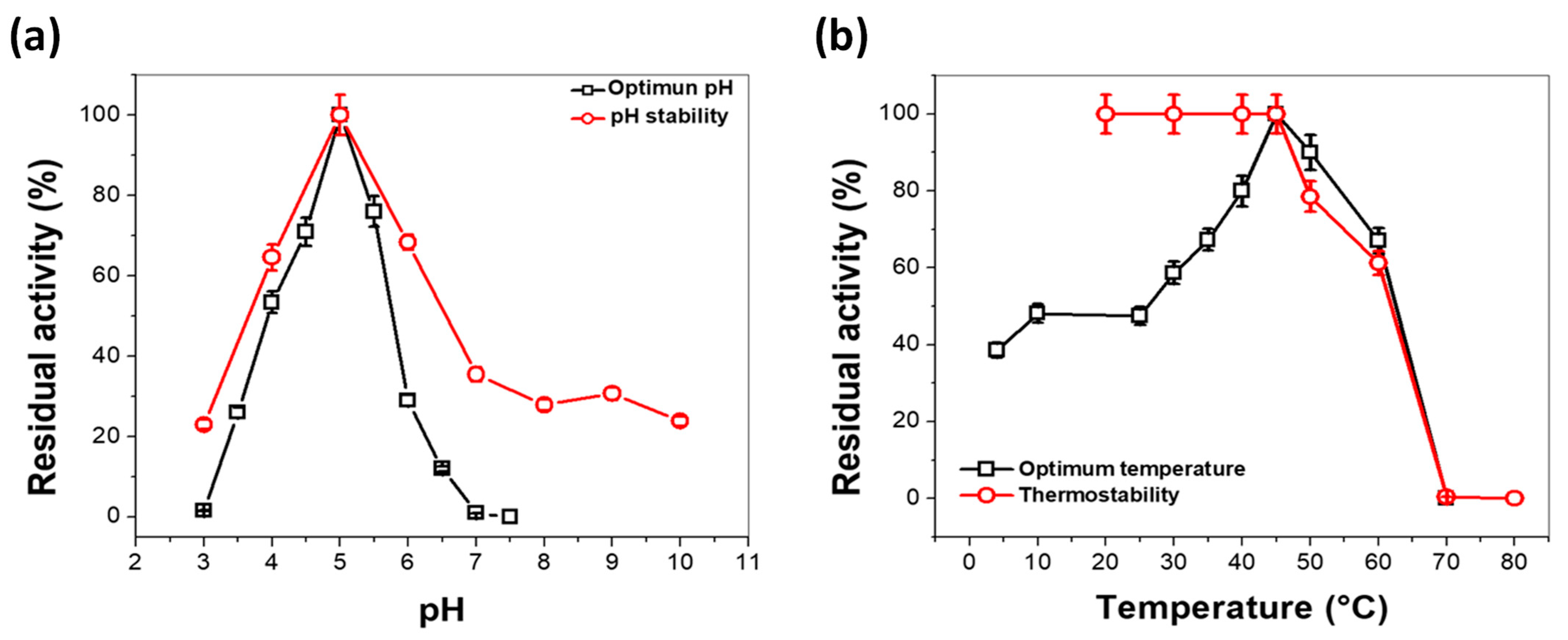
3.4.2. Effect of pH and Temperature on Laccase Activity
3.4.3. Determination of Steady-State Kinetic Parameters
3.4.4. Effect of DL-Dithiothreitol (DTT) on Laccase Activity
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Eichlerová, I.; Baldrian, P. Ligninolytic Enzyme Production and Decolorization Capacity of Synthetic Dyes by Saprotrophic White Rot, Brown Rot, and Litter Decomposing Basidiomycetes. J. Fungi 2020, 6, 301. [Google Scholar] [CrossRef]
- Li, X.; Qu, Z.; Zhang, Y.; Ge, Y.; Sun, H. Soil Fungal Community and Potential Function in Different Forest Ecosystems. Diversity 2022, 14, 520. [Google Scholar] [CrossRef]
- Kumar, A.; Chandra, R. Ligninolytic enzymes and its mechanisms for degradation of lignocellulosic waste in environment. Heliyon 2020, 6, e03170. [Google Scholar] [CrossRef] [PubMed]
- Nargotra, P.; Sharma, V.; Lee, Y.-C.; Tsai, Y.-H.; Liu, Y.-C.; Shieh, C.-J.; Tsai, M.-L.; Dong, C.-D.; Kuo, C.-H. Microbial Lignocellulolytic Enzymes for the Effective Valorization of Lignocellulosic Biomass: A Review. Catalysts 2023, 13, 83. [Google Scholar] [CrossRef]
- Bilal, M.; Bagheri, A.R.; Vilar, D.S.; Aramesh, N.; Eguiluz, K.I.B.; Romanholo Ferreira, L.F.; Ashraf, S.S.; Iqbal, H.M.N. Oxidoreductases as a versatile biocatalytic tool to tackle pollutants for clean environment—A review. J. Chem. Technol. Biotechnol. 2022, 97, 420–435. [Google Scholar] [CrossRef]
- Leynaud Kieffer Curran, L.M.C.; Pham, L.T.M.; Sale, K.L.; Simmons, B.A. Review of advances in the development of laccases for the valorization of lignin to enable the production of lignocellulosic biofuels and bioproducts. Biotechnol. Adv. 2022, 54, 107809. [Google Scholar] [CrossRef]
- Bilal, M.; Rasheed, T.; Nabeel, F.; Iqbal, H.M.N.; Zhao, Y. Hazardous contaminants in the environment and their laccase-assisted degradation-A review. J. Environ. Manag. 2019, 234, 253–264. [Google Scholar] [CrossRef]
- Junior, J.A.; Vieira, Y.A.; Cruz, I.A.; da Silva Vilar, D.; Aguiar, M.M.; Torres, N.H.; Bharagava, R.N.; Lima, Á.S.; de Souza, R.L.; Romanholo Ferreira, L.F. Sequential degradation of raw vinasse by a laccase enzyme producing fungus Pleurotus sajor-caju and its ATPS purification. Biotechnol. Rep. 2019, 25, e00411. [Google Scholar] [CrossRef]
- Loi, M.; Glazunova, O.; Fedorova, T.; Logrieco, A.F.; Mulè, G. Fungal Laccases: The Forefront of Enzymes for Sustainability. J. Fungi 2021, 7, 1048. [Google Scholar] [CrossRef]
- Janusz, G.; Pawlik, A.; Swiderska-Burek, U.; Polak, J.; Sulej, J.; Jarosz-Wilkolazka, A.; Paszczynski, A. Laccase Properties, Physiological Functions, and Evolution. Int. J. Mol. Sci 2020, 21, 966. [Google Scholar] [CrossRef] [Green Version]
- Montiel-González, A.M.; Marcial, J. Phenoloxidases of Fungi and Bioremediation. In Fungal Bioremediation, Fundamentals and Applications; Tomasini, A., León-Santiesteban, H., Eds.; CRC Press: London, UK; New York, NY, USA, 2019; pp. 62–90. [Google Scholar]
- Rodríguez-Couto, S. Fungal Laccase: A Versatile Enzyme for Biotechnological Applications. In Recent Advancement in White Biotechnology Through Fungi. Fungal Biology; Yadav, A., Mishra, S., Singh, S., Gupta, A., Eds.; Springer: Cham, Switzerland, 2019; pp. 429–457. [Google Scholar]
- Cruz-Vázquez, A.; Tomasini, A.; Armas-Tizapantzi, A.; Marcial-Quino, J.; Montiel-González, A.M. Extracellular proteases and laccases produced by Pleurotus ostreatus PoB: The effects of proteases on laccase activity. Int. Microbiol. 2022, 25, 495–502. [Google Scholar] [CrossRef] [PubMed]
- Nagai, M.; Sato, T.; Watanabe, H.; Saito, K.; Kawata, M.; Enei, H. Purification and characterization of an extracellular laccase from the edible mushroom Lentinula edodes, and decolorization of chemically different dyes. Appl. Microbiol. Biotechnol. 2002, 60, 327–335. [Google Scholar] [PubMed]
- Wu, Y.R.; Luo, Z.H.; Chow, R.K.K.; Vrijmoed, L.P.P. Purification and characterization of an extracellular laccase from the anthracene-degrading fungus Fusarium solani MAS2. Bioresour. Technol. 2010, 101, 9772–9777. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.Y.; Huang, Y.C.; Wei, C.M.; Meng, M.; Liu, W.H.; Yang, C.H. Properties of the newly isolated extracellular thermo-alkali-stable laccase from thermophilic actinomycetes, Thermobifida fusca and its application in dye intermediates oxidation. AMB Express 2013, 3, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sajben-Nagy, E.; Manczinger, L.; Škrbić, B.; Živančev, J.; Antić, I.; Krisch, J.; Vágvölgyi, C. Characterization of an extracellular laccase of Leptosphaerulina chartarum. World J. Microbiol. Biotechnol. 2014, 30, 2449–2458. [Google Scholar] [CrossRef] [PubMed]
- Ling, Z.R.; Wang, S.S.; Zhu, M.J.; Ning, Y.J.; Wang, S.N.; Li, B.; Yang, A.Z.; Zhang, G.Q.; Zhao, X.M. An extracellular laccase with potent dye decolorizing ability from white rot fungus Trametes sp. LAC-01. Int. J. Biol. Macromol. 2015, 81, 785–793. [Google Scholar] [CrossRef]
- Campos, P.A.; Levin, L.N.; Wirth, S.A. Heterologous production, characterization and dye decolorization ability of a novel thermostable laccase isoenzyme from Trametes trogii BAFC 463. Process. Biochem. 2016, 51, 895–903. [Google Scholar] [CrossRef]
- Wang, S.S.; Ning, Y.J.; Wang, S.N.; Zhang, J.; Zhang, G.Q.; Chen, Q.J. Purification, characterization, and cloning of an extracellular laccase with potent dye decolorizing ability from white rot fungus Cerrena unicolor GSM-01. Int. J. Biol. Macromol. 2017, 95, 920–927. [Google Scholar] [CrossRef]
- Mtibaà, R.; Barriuso, J.; de Eugenio, L.; Aranda, E.; Belbahri, L.; Nasri, M.; Martínez, M.J.; Mechichi, T. Purification and characterization of a fungal laccase from the ascomycete Thielavia sp. and its role in the decolorization of a recalcitrant dye. Int. J. Biol. Macromol. 2018, 120 Pt B, 1744–1751. [Google Scholar] [CrossRef] [Green Version]
- Senthivelan, T.; Kanagaraj, J.; Panda, R.C.; Narayani, T. Screening and production of a potential extracellular fungal laccase from Penicillium chrysogenum: Media optimization by response surface methodology (RSM) and central composite rotatable design (CCRD). Biotechnol. Rep. 2019, 23, e00344. [Google Scholar] [CrossRef]
- Akpinar, M.; Ozturk-Urek, R. Decolorization and degradation potential of enhanced lignocellulolytic enzymes production by Pleurotus eryngii using cherry waste from industry. Biotechnol. Appl. Biochem. 2020, 67, 760–773. [Google Scholar] [CrossRef]
- Maestre-Reyna, M.; Liu, W.C.; Jeng, W.Y.; Lee, C.C.; Hsu, C.A.; Wen, T.N.; Wang, A.H.; Shyur, L.F. Structural and functional roles of glycosylation in fungal laccase from Lentinus sp. PLoS ONE 2015, 10, e0120601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arregui, L.; Ayala, M.; Gómez-Gil, X.; Gutiérrez-Soto, G.; Hernández-Luna, C.E.; de Los Santos, M.H.; Levin, L.; Rojo-Domínguez, A.; Romero-Martínez, D.; Saparrat, M.C.N.; et al. Laccases: Structure, function, and potential application in water bioremediation. Microb. Cell. Fact. 2019, 18, 1–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goudopoulou, A.; Krimitzas, A.; Typas, M.A. Differential gene expression of ligninolytic enzymes in Pleurotus ostreatus grown on olive oil mill wastewater. Appl. Microbiol. Biotechnol. 2010, 88, 541–551. [Google Scholar] [CrossRef] [PubMed]
- Castanera, R.; Pérez, G.; Omarini, A.; Alfaro, M.; Pisabarro, A.G.; Faraco, V.; Amore, A.; Ramírez, L. Transcriptional and enzymatic profiling of Pleurotus ostreatus laccase genes in submerged and solid-state fermentation cultures. Appl. Environ. Microbiol. 2012, 78, 4037–4045. [Google Scholar] [CrossRef] [Green Version]
- Vilar, D.D.S.; Bilal, M.; Bharagava, R.N.; Kumar, A.; Nadda, A.K.; Salazar-Banda, G.R.; Eguiluz, K.I.B.; Ferreira, L.F.R. Lignin-modifying enzymes: A green and environmental responsive technology for organic compound degradation. J. Chem. Technol. Biotechnol. 2021, 96, 6751. [Google Scholar]
- Piscitelli, A.; Giardina, P.; Lettera, V.; Pezzella, C.; Sannia, G.; Faraco, V. Induction and transcriptional regulation of laccases in fungi. Curr. Genomics 2011, 12, 104–112. [Google Scholar] [CrossRef] [Green Version]
- Palmieri, G.; Giardina, P.; Bianco, C.; Fontanella, B.; Sannia, G. Copper Induction of Laccase Isoenzymes in the Ligninolytic Fungus Pleurotus ostreatus. Appl. Environ. Microbiol. 2000, 66, 920–924. [Google Scholar] [CrossRef] [Green Version]
- Palmieri, G.; Cennamo, G.; Faraco, V.; Amoresano, A.; Sannia, G.; Giardina, P. Atypical laccase isoenzymes from copper supplemented Pleurotus ostreatus cultures. Enzyme Microb. Technol. 2003, 33, 220–230. [Google Scholar] [CrossRef]
- Lu, X.; Ding. S. Effect of Cu2+, Mn2+ and aromatic compounds on the production of laccase isoforms by Coprinus comatus. Mycoscience 2010, 51, 68–74. [Google Scholar] [CrossRef]
- Bertrand, B.; Martínez-Morales, F.; Trejo-Hernández, M.R. Fungal laccases: Induction and production. Rev. Mex. Ing. Quim. 2013, 12, 473–488. [Google Scholar]
- Yang, J.; Wang, G.; Ng, T.B.; Lin, J.; Ye, X. Laccase Production and Differential Transcription of Laccase Genes in Cerrena sp. in Response to Metal Ions, Aromatic Compounds, and Nutrients. Front. Microbiol. 2016, 6, 1558. [Google Scholar] [CrossRef] [PubMed]
- Kandasamy, S.; Muniraj, I.K.; Purushothaman, N.; Sekar, A.; Sharmila, D.J.S.; Kumarasamy, R.; Uthandi, S. High level secretion of Laccase (LccH) from a newly isolated white-rot Basidiomycete, Hexagonia hirta MSF2. Front. Microbiol. 2016, 7, 707. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glazunova, O.A.; Shakhova, N.V.; Psurtseva, N.V.; Moiseenko, K.V.; Kleimenov, S.Y.; Fedorova, T.V. White-rot basidiomycetes Junghuhnia nitida and Steccherinum bourdotii: Oxidative potential and laccase properties in comparison with Trametes hirsuta and Coriolopsis caperata. PLoS ONE 2018, 13, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Calaça, F.J.S.; Cortez, V.G.; Xavier-Santos, S. Dung fungi from Brazil: Agrocybe pediades (Fr.) Fayod (Basidiomycota) in Cerrado. Sci. Plena 2020, 16, 066201. [Google Scholar] [CrossRef]
- Téllez-Téllez, M.; Fernández, J.F.; Montiel-González, A.M.; Sánchez, C.; Díaz-Godínez, G. Growth and laccase production by Pleurotus ostreatus in submerged and solid-state fermentation. Appl Microbiol. Biotechnol. 2008, 81, 675–679. [Google Scholar] [CrossRef]
- Madigan, M.T.; Martinko, J.M.; Bender, K.S.; Buckley, D.H.; Stahl, D.A. BROCK: Biología de los Microorganismos; Pearson: Madrid, Spain, 2005. [Google Scholar]
- Sharma, M.; Chaurasia, P.K.; Yadav, A.; Yadav, R.S.S.; Yadava, S.; Yadav, K.D.S. Purification and characterization of a thermally stable yellow laccase from Daedalea flavida MTCC-145 with higher catalytic performance towards selective synthesis of substituted benzaldehydes. Russian. J. Bioorg. Chem. 2016, 42, 59–68. [Google Scholar] [CrossRef]
- Armas-Tizapantzi, A.; Marcial, J.; Fernández, F.J.; Estrada-Torres, A.; Pérez-Godínez, E.A.; Montiel-González, A.M. Laccase gene silencing negatively effects growth and development in Pleurotus ostreatus. Indian J. Biotechnol. 2019, 18, 42–51. [Google Scholar]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Si, J.; Peng, F.; Cui, B. Purification, biochemical characterization and dye decolorization capacity of an alkali-resistant and metal-tolerant laccase from Trametes pubescens. Bioresour. Technol. 2013, 128, 49–57. [Google Scholar] [CrossRef]
- Si, J.; Ma, H.; Cao, Y.; Cui, B.; Dai, Y. Introducing a Thermo-Alkali-Stable, Metallic Ion-Tolerant Laccase Purified From White Rot Fungus Trametes hirsuta. Front. Microbiol. 2021, 12, 670163. [Google Scholar] [CrossRef] [PubMed]
- Si, J.; Wu, Y.; Ma, H.F.; Cao, Y.J.; Sun, Y.F.; Cui, B.K. Selection of a pH- and temperature-stable laccase from Ganoderma australe and its application for bioremediation of textile dyes. J. Environ. Manag. 2021, 299, 113619. [Google Scholar] [CrossRef] [PubMed]
- Zheng, F.; An, Q.; Meng, G.; Wu, X.J.; Dai, Y.C.; Si, J.; Cui, B.K. A novel laccase from white rot fungus Trametes orientalis: Purification, characterization, and application. Int. J. Biol. Macromol. 2017, 102, 758–770. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Manzo, S.; Terrón-Hernández, J.; De la Mora-De la Mora, I.; González-Valdez, A.; Marcial-Quino, J.; García-Torres, I.; Vanoye-Carlo, A.; López-Velázquez, G.; Hernández-Alcántara, G.; Oria-Hernández, J.; et al. The Stability of G6PD Is Affected by Mutations with Different Clinical Phenotypes. Int. J. Mol. Sci. 2014, 15, 21179. [Google Scholar] [CrossRef] [Green Version]
- Ullrich, R.; Huong, L.M.; Dung, N.L.; Hofrichter, M. Laccase from the medicinal mushroom Agaricus blazei: Production, purification and characterization. Appl. Microbiol. Biotechnol. 2005, 67, 357–363. [Google Scholar] [CrossRef]
- Liu, H.; Tong, C.; Du, B.; Liang, S.; Lin, Y. Expression and characterization of LacMP, a novel fungal laccase of Moniliophthora perniciosa FA553. Biotechnol. Lett. 2015, 37, 1829–1835. [Google Scholar] [CrossRef]
- Chen, S.; Ge, W.; Buswell, J.A. Biochemical and molecular characterization of a laccase from the edible straw mushroom, Volvariella volvacea. Eur. J. Biochem. 2004, 271, 318–328. [Google Scholar] [CrossRef] [Green Version]
- Aza, P.; Molpeceres, G.; Ruiz-Dueñas, F.J.; and Camarero, S. Heterologous Expression, Engineering and Characterization of a Novel Laccase of Agrocybe pediades with Promising Properties as Biocatalyst. J. Fungi 2021, 7, 359. [Google Scholar] [CrossRef]
- Daroch, M.; Houghton, C.A.; Moore, J.K.; Wilkinson, M.C.; Carnell, A.J.; Bates, A.D.; Iwanejko, L.A. Glycosylated yellow laccases of the basidiomycete Stropharia aeruginosa. Enzyme Microb. Technol. 2014, 58–59, 1–7. [Google Scholar] [CrossRef]
- Dwivedi, U.N.; Singh, P.; Pandey, V.P.; Kumar, A. Structure–function relationship among bacterial, fungal and plant laccases. J. Mol. Catal. B Enzym. 2011, 68, 117–128. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Purification Steps | Total Protein (mg) | Specific Activity (µmol∙min−1∙mg−1) | Total Activity (µmol∙min−1) | Yield (%) |
|---|---|---|---|---|
| EECE | 1518 | 2.8 | 4250 | 100 |
| Sephacryl 200 | 1.3 | 285 | 370.5 | 8.7 |
| Family | Species | Molecular Weight (kDa) | pH | Temperature (°C) | Km (μM) | Reference |
|---|---|---|---|---|---|---|
| Strophariaceae | Agrocybe pediades | 72 | 5.0 | 40 | 100 | This work |
| Agaricaceae | Agaricus blazei | 66 | 5.5 | n.d * | 1026 | [48] |
| Marasmiaceae | Lentinula edodes | 72 | 4.0 | 40 | 557 | [14] |
| Marasmiaceae | Moniliophthora perniciosa | 57 | 6.5 | 55 | n.d. | [49] |
| Pleurotaceae | Pleurotus ostreatus | 83−85 | 5.5 | 35 | 8800 | [31] |
| Pluteaceae | Volvariella volvacea | 58 | 4.6 | 45 | 570 | [50] |
| Chaetomiaceae | Thielavia sp. | 70 | 5.0 | 70 | 24 | [21] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
González-González, P.; Gómez-Manzo, S.; Tomasini, A.; Martínez y Pérez, J.L.; García Nieto, E.; Anaya-Hernández, A.; Ortiz Ortiz, E.; Castillo Rodríguez, R.A.; Marcial-Quino, J.; Montiel-González, A.M. Laccase Production from Agrocybe pediades: Purification and Functional Characterization of a Consistent Laccase Isoenzyme in Liquid Culture. Microorganisms 2023, 11, 568. https://doi.org/10.3390/microorganisms11030568
González-González P, Gómez-Manzo S, Tomasini A, Martínez y Pérez JL, García Nieto E, Anaya-Hernández A, Ortiz Ortiz E, Castillo Rodríguez RA, Marcial-Quino J, Montiel-González AM. Laccase Production from Agrocybe pediades: Purification and Functional Characterization of a Consistent Laccase Isoenzyme in Liquid Culture. Microorganisms. 2023; 11(3):568. https://doi.org/10.3390/microorganisms11030568
Chicago/Turabian StyleGonzález-González, Paulina, Saúl Gómez-Manzo, Araceli Tomasini, José Luis Martínez y Pérez, Edelmira García Nieto, Arely Anaya-Hernández, Elvia Ortiz Ortiz, Rosa Angélica Castillo Rodríguez, Jaime Marcial-Quino, and Alba Mónica Montiel-González. 2023. "Laccase Production from Agrocybe pediades: Purification and Functional Characterization of a Consistent Laccase Isoenzyme in Liquid Culture" Microorganisms 11, no. 3: 568. https://doi.org/10.3390/microorganisms11030568