
Cross-Linked Enzyme Aggregate (CLEA) Preparation from Waste Activated Sludge


Abstract
:
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. CLEA Preparation
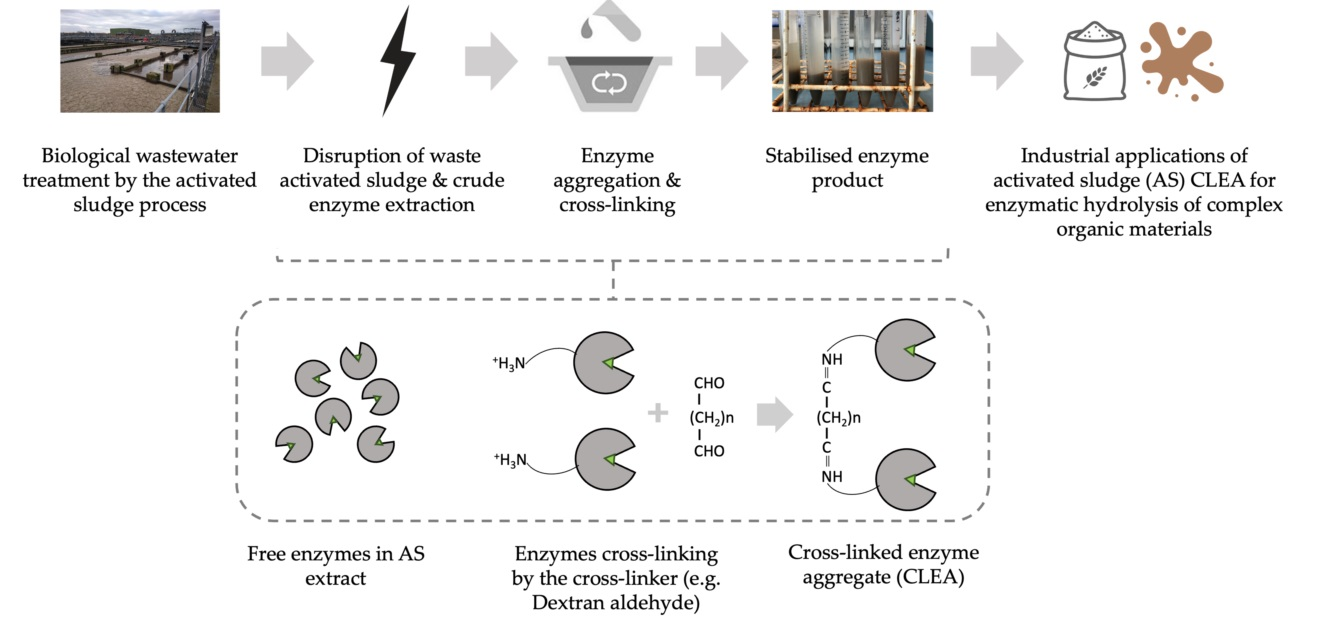
2.2.1. General Processes
2.2.2. Chemical Precipitation of Soluble Enzymes
2.2.3. Cross-Linking of Precipitated Enzymes
2.3. CLEA Preparation from Crude AS Enzyme Extracts
2.3.1. Enzyme Cross-Linking with Glutaraldehyde
2.3.2. Enzyme Cross-Linking with Dextran Aldehyde
2.4. Enzyme Activity Assays
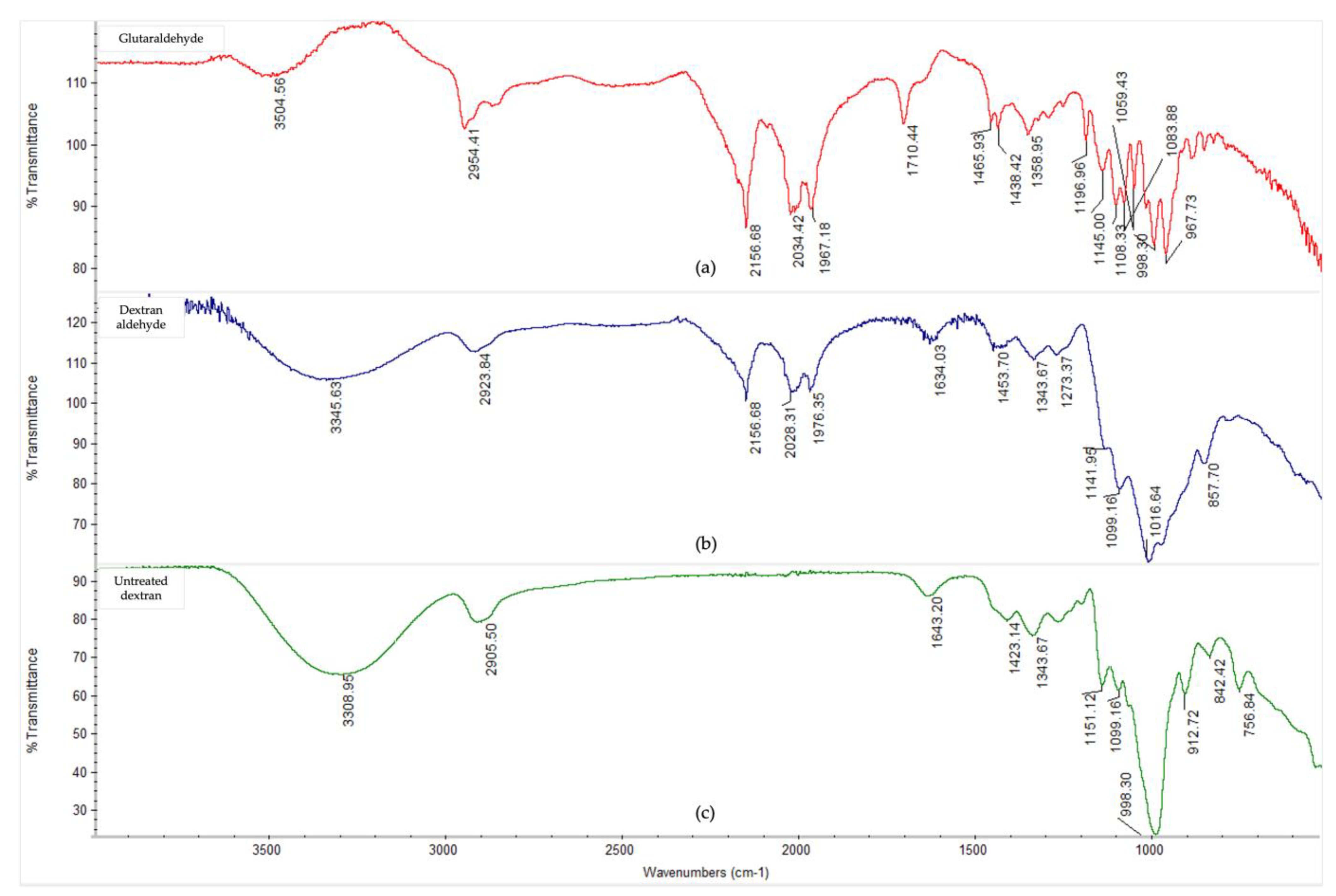
2.5. Fourier-Transform Infrared Spectroscopy
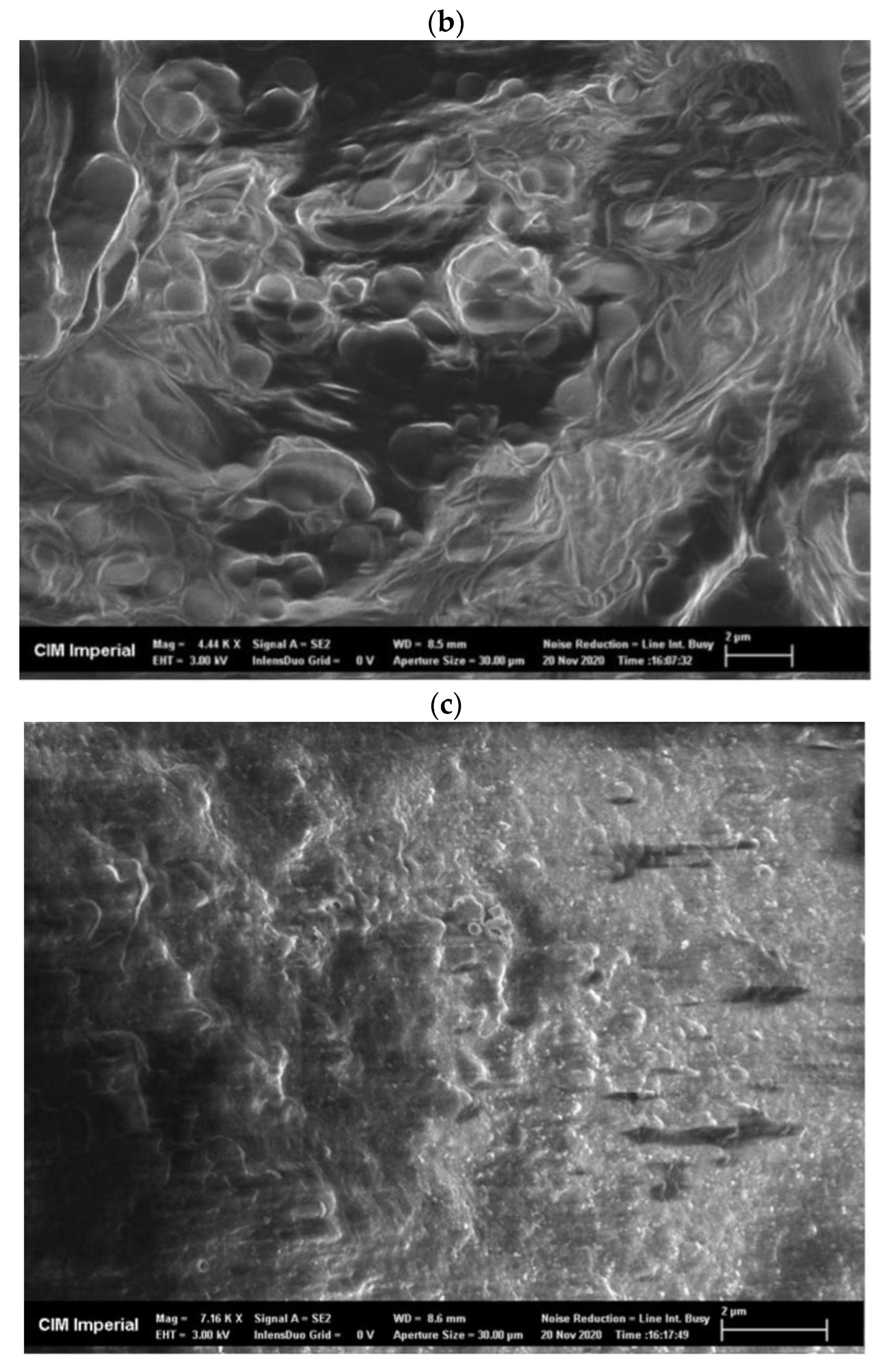
2.6. CLEA Morphology
2.7. Hydrolysis of Organic Materials by CLEA
2.7.1. Substrate Preparation
- Gamma (γ) irradiation: diluted WAS was exposed to a cesium (Cs) 137 radiator (Gammacell 3000 ELAN, Best Theratronics Ltd., Canada). The WAS suspension (450 mL) was placed in a 500 mL HDPE bottle and was exposed to the radiator for 10 h at room temperature; the total radiation dose was 1944 Gy at a dose rate of 3.24 Gy/min.
- Sonication: diluted WAS was treated by sonication probe (VCX130, Sonics & Materials Inc., Newtown, CT, USA) for 10 min duration at 0 °C, 40% amplitude, in pulse cycle mode with 1 min on followed by 1 min off.
2.7.2. Hydrolysis of Organic Substrates
2.7.3. Indicators of the Progress of Enzymatic Hydrolysis
- Soluble Total Organic Carbon
- Reducing Sugar and Tyrosine
2.7.4. Progress Curve of Enzymatic Hydrolysis
- CTotal for sTOC was derived from the specific VS content (0.989 g/g TS) and the element composition ratio of wheat flour (C:H:O:N:S = 1:2.004:0.625:0.249:0.01) reported by [41], and it was equivalent to 0.568 g/g TS.
- CTotal for reducing sugar was derived by dividing the total carbohydrate content in the flour (0.771 g/g TS, which was assumed to be in the form of starch) by the coefficient, 0.9 [42], and it was equivalent to 0.857 g/g TS.
- CTotal for tyrosine was estimated based on the value reported by Siddiqi et al. [43], and it was 5.6 mg/g TS.
2.8. Statistical Analysis
3. Results and Discussion
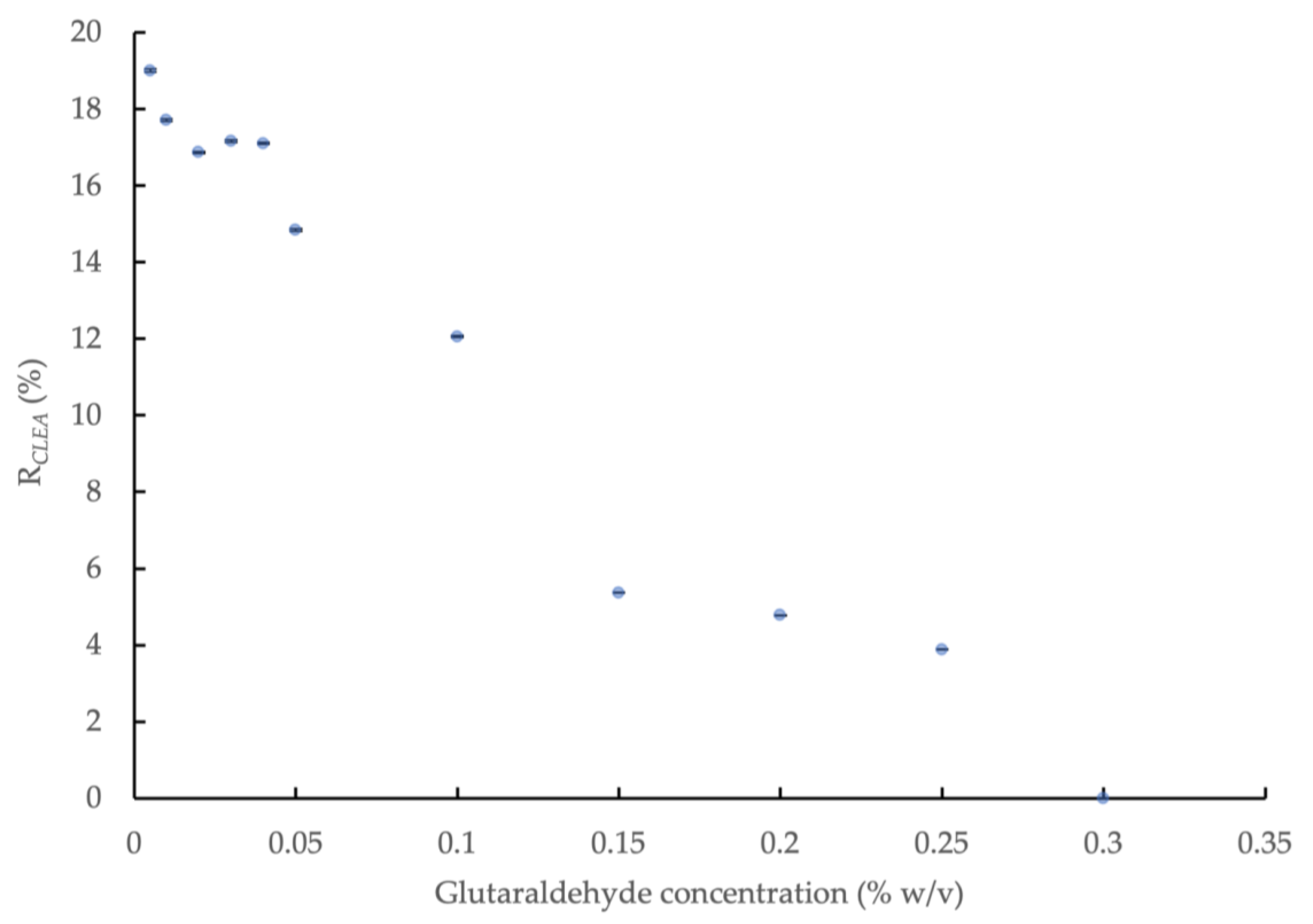
3.1. Cross-Linking AS Enzymes with Glutaraldehyde
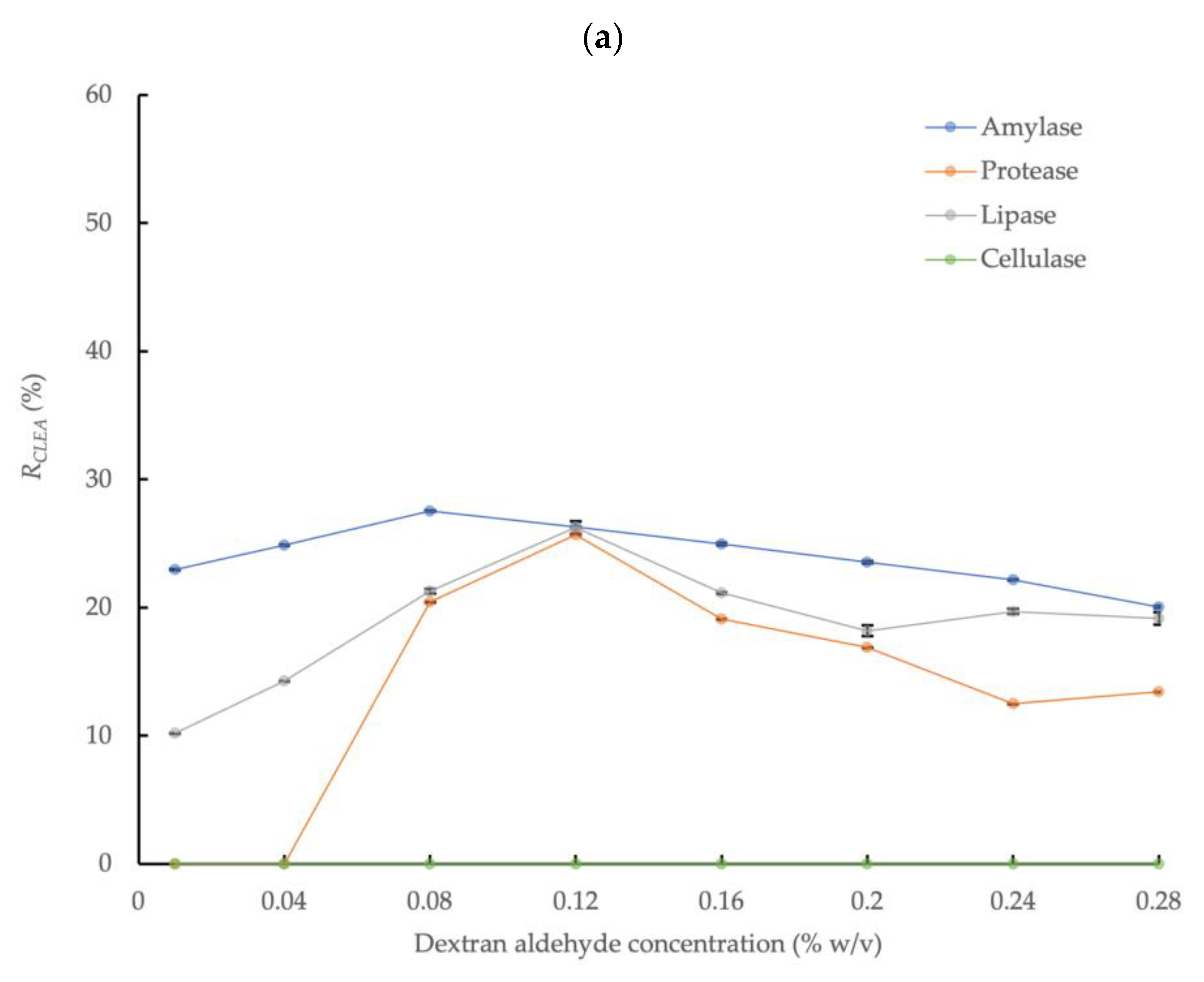
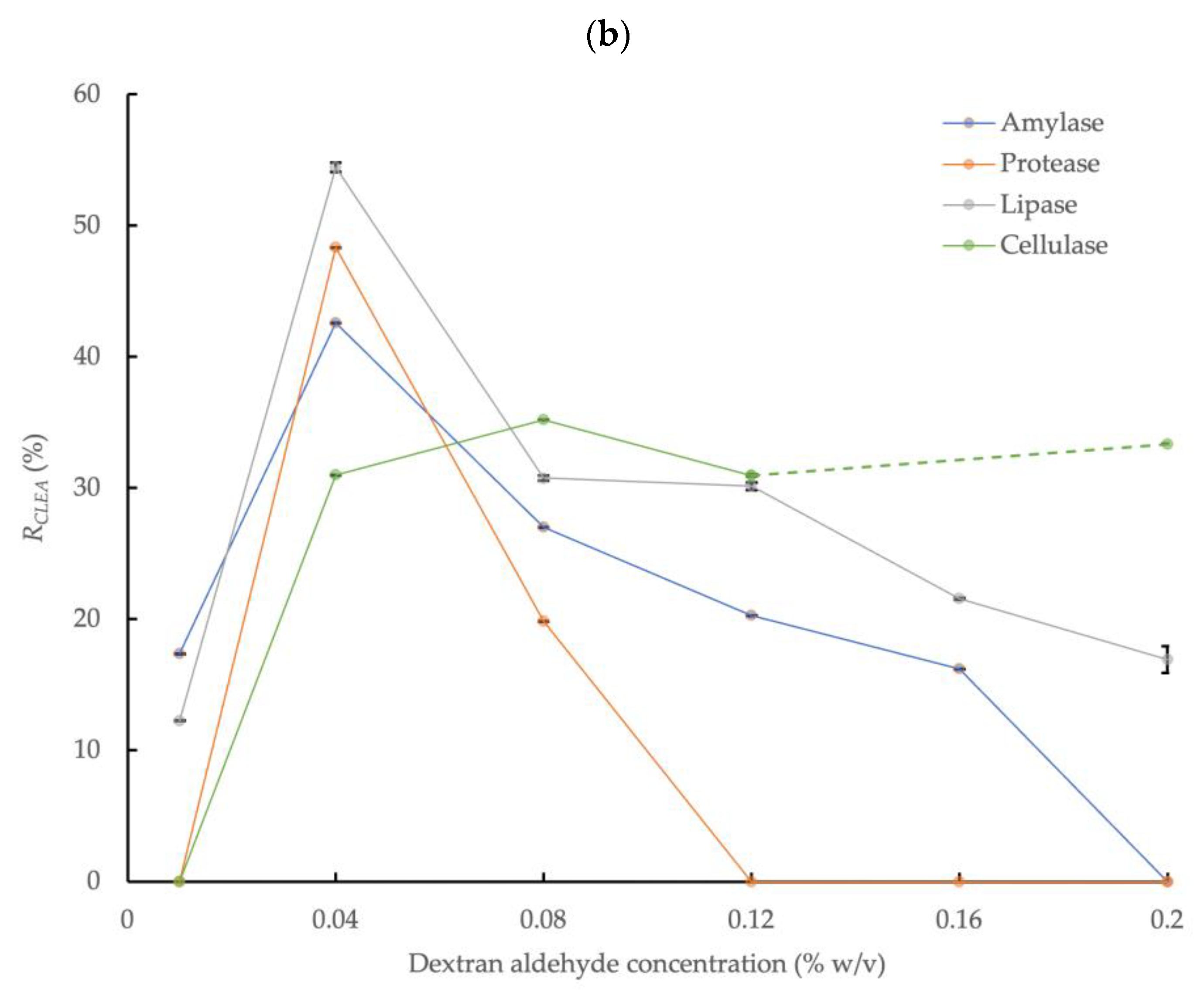
3.2. Cross-Linking AS Enzymes with Dextran Aldehyde
3.2.1. Ammonium Sulfate Precipitation
3.2.2. Acetone Precipitation
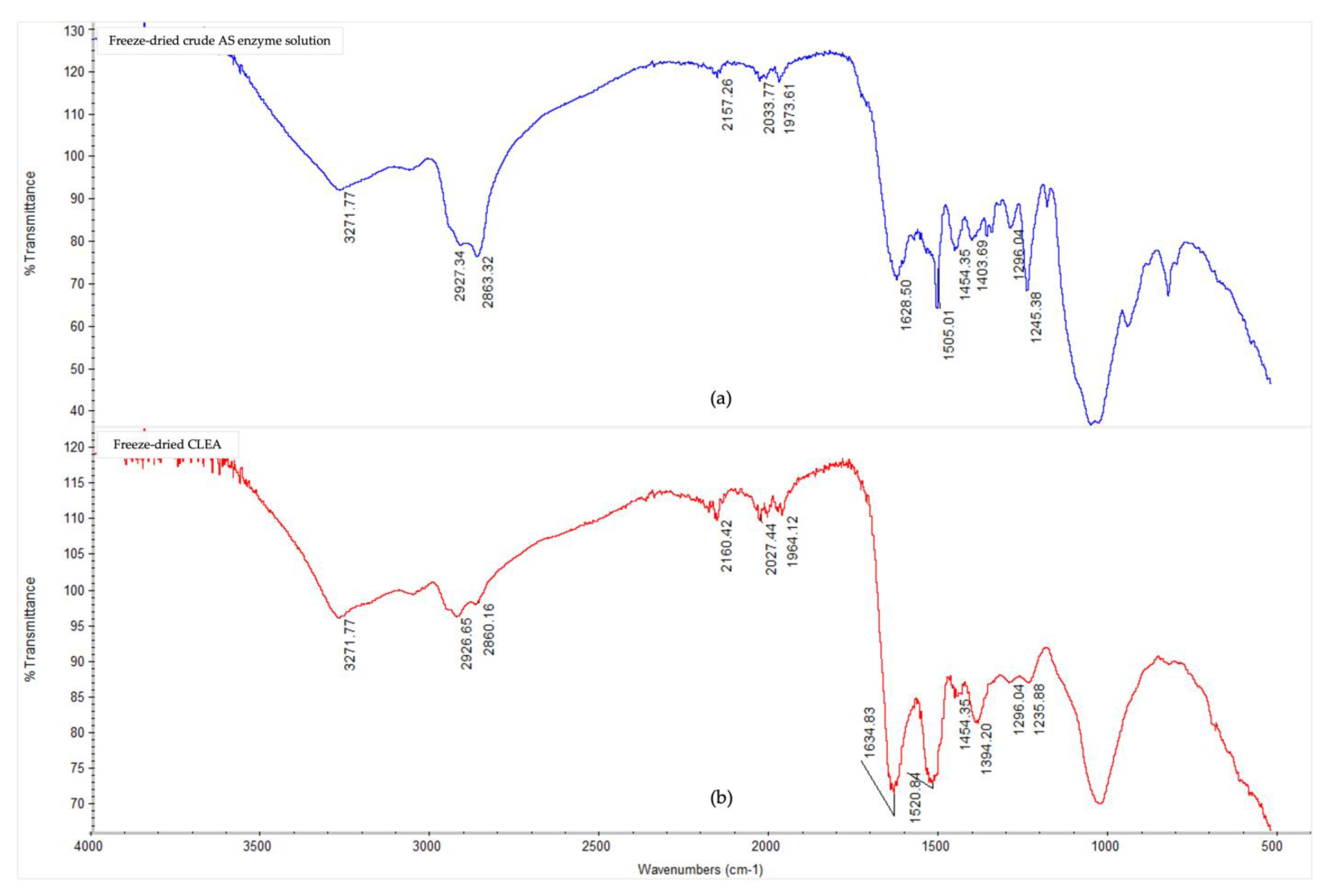
3.3. Fourier-Transform Infrared Spectroscopy Analysis
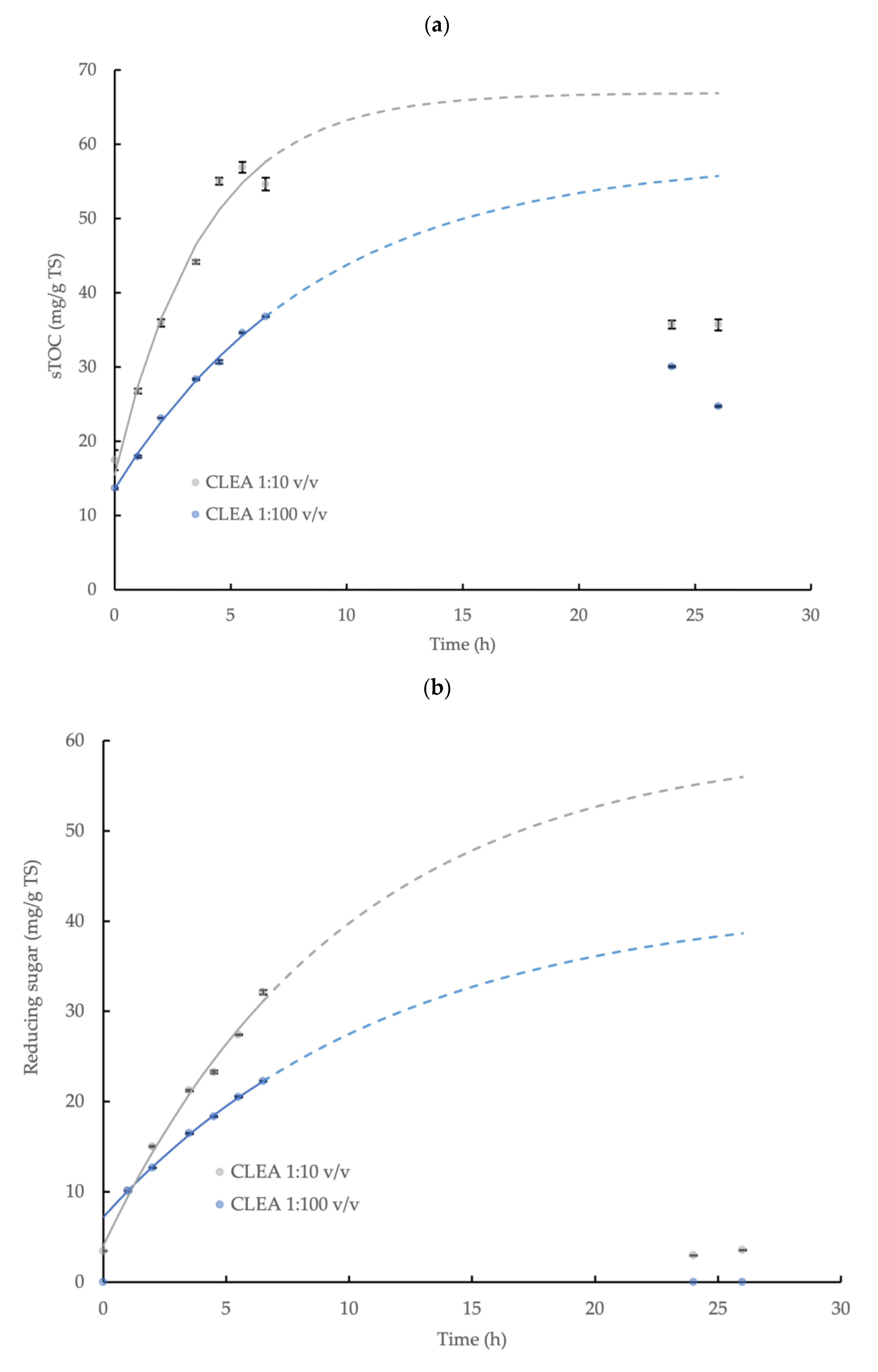
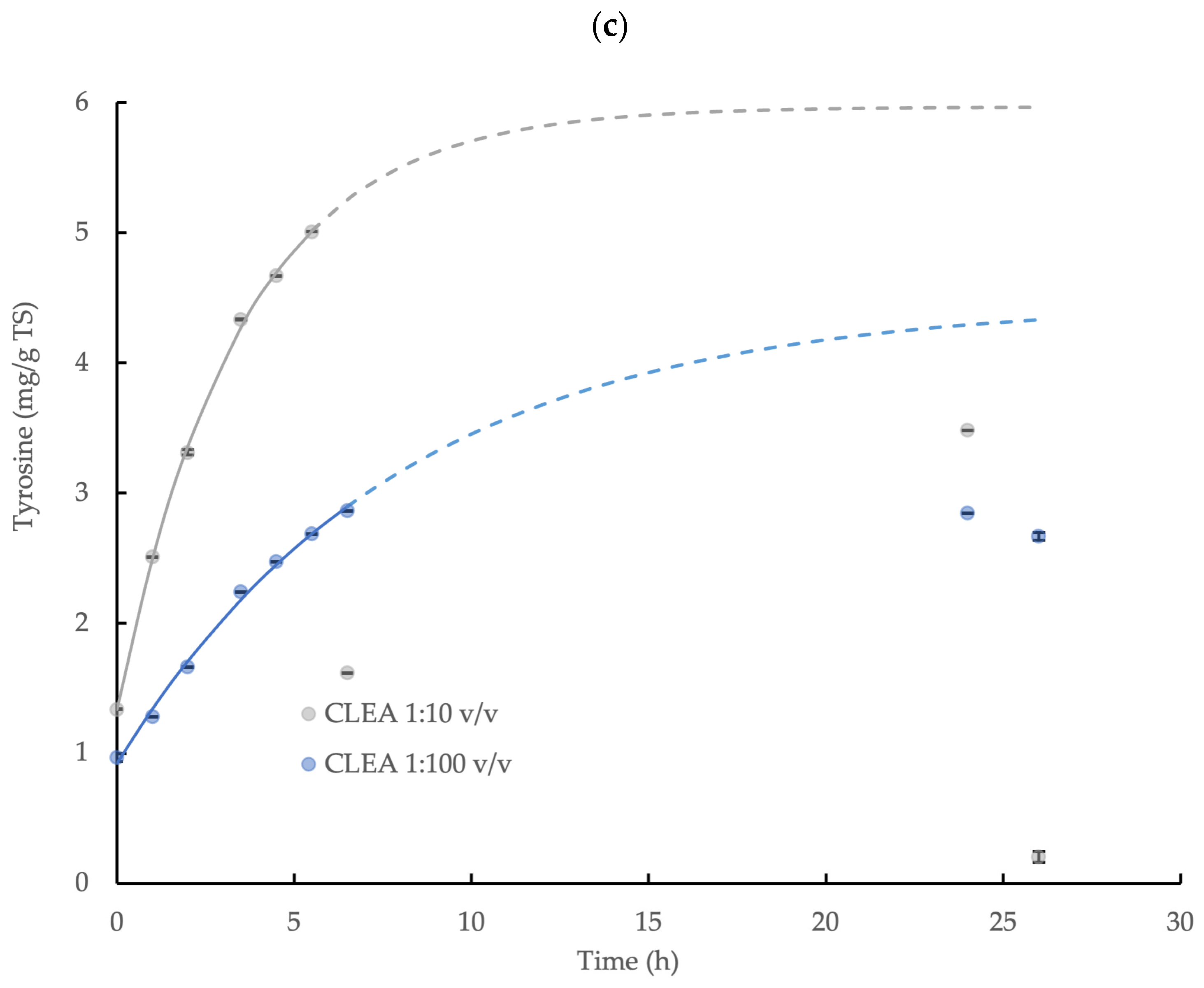
3.4. Hydrolysis Kinetics of Wheat Flour
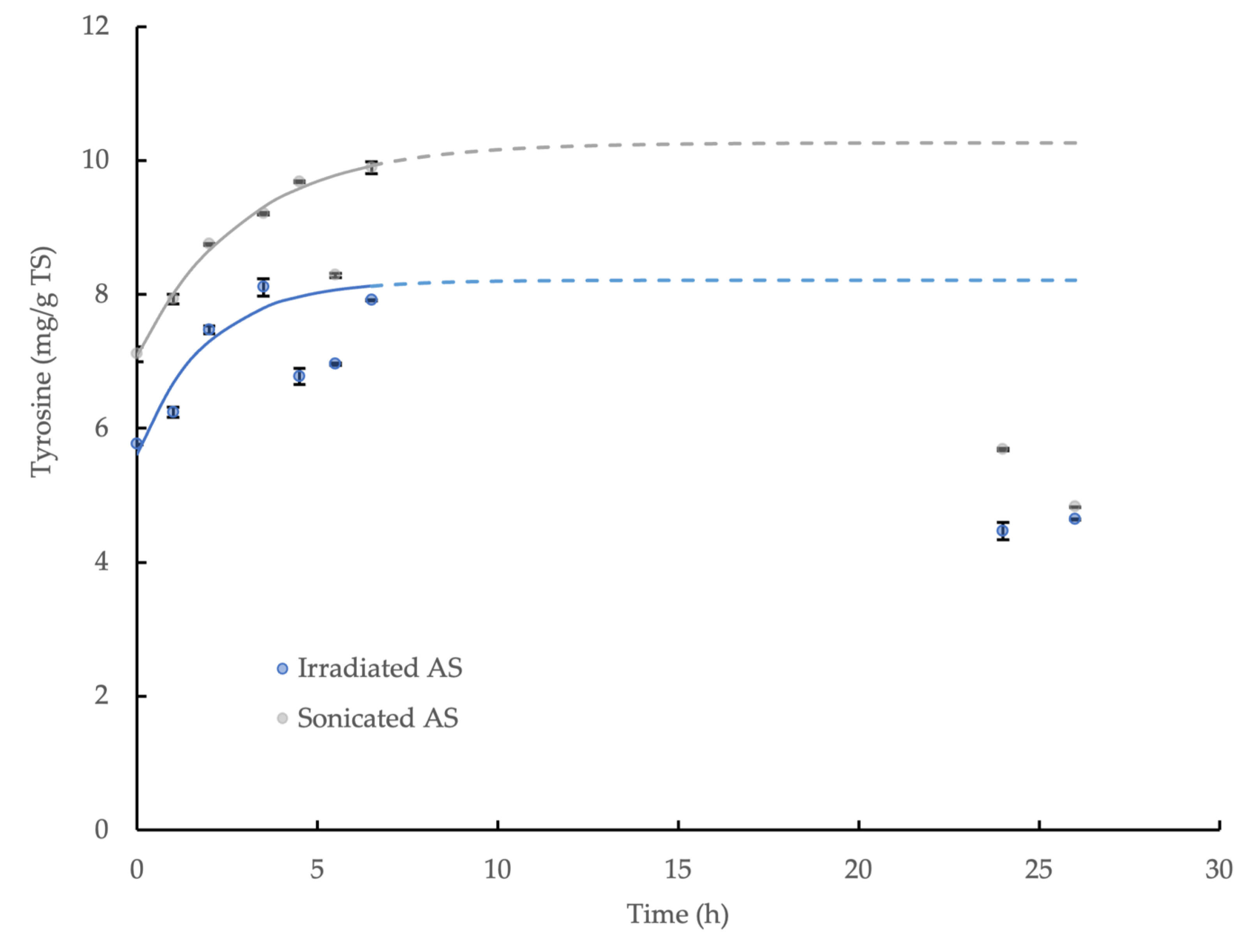
3.5. Hydrolysis Kinetics of γ-Irradiated WAS and Sonicated WAS
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A. Fourier-Transform Infrared Spectroscopy


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Class and Groups | Assignments | Band/Peak Position (Wavenumber, cm−1) | |
|---|---|---|---|
| Free Enzyme | CLEA | ||
| Amide | N-H stretching | 3271 | 3271 |
| Aliphatic; amine salt | C-H stretching; N-H stretching | 2863–2927 | 2860–2926 |
| Amide | C=O stretching, N-H bending and C-N stretching | 1628 | 1634 |
| Imine (Schiff’s Base) | C=N stretching | - | 1634 |
| Amide | N-H bending and C-N stretching | 1505 | 1520 |
| Amide/amine | N-H Bending and C-N stretching, along with C-H and N-H deformation | 1245 | 1235 |
Appendix B. Statistical Validation Protocol
Appendix C. Morphology of AS CLEAs


References
- Liu, Z.; Smith, S.R. Enzyme recovery from biological wastewater treatment. Waste Biomass Valorization 2021, 12, 4185–4211. [Google Scholar]
- Liu, Z.; Smith, S.R. Enzyme activity of waste activated sludge extracts. Water Sci. Technol. 2019, 80, 1861–1869. [Google Scholar]
- Mallick, P.; Akunna, J.C.; Walker, G.M. Anaerobic digestion of distillery spent wash: Influence of enzymatic pre-treatment of intact yeast cells. Bioresour. Technol. 2010, 101, 1681–1685. [Google Scholar]
- Sahutoglu, A.S.; Akgul, C. Immobilisation of Aspergillus oryzae alpha-amylase and Aspergillus niger glucoamylase enzymes as cross-linked enzyme aggregates. Chem. Pap. 2015, 69, 433–439. [Google Scholar]
- Asgher, M.; Bashir, F.; Iqbal, H.M.N. Protease-based cross-linked enzyme aggregates with improved catalytic stability, silver removal, and dehairing potentials. Int. J. Biol. Macromol. 2018, 118, 1247–1256. [Google Scholar] [PubMed]
- Okuda, K.; Urabe, I.; Yamada, Y.; Okada, H. Reaction of glutaraldehyde with amino and thiol compounds. J. Ferment. Bioeng. 1991, 71, 100–105. [Google Scholar]
- Migneault, I.; Dartiguenave, C.; Bertrand, M.J.; Waldron, K.C. Glutaraldehyde: Behavior in aqueous solution, reaction with proteins, and application to enzyme crosslinking. Biotechniques 2004, 37, 790–802. [Google Scholar]
- Amaral-Fonseca, M.; Kopp, W.; Giordano, R.D.C.; Fernandez-Lafuente, R.; Tardioli, P.W. Preparation of magnetic cross-linked amyloglucosidase aggregates: Solving some activity problems. Catalysts 2018, 8, 496–516. [Google Scholar]
- Orrego, A.H.; Ghobadi, R.; Moreno-Perez, S.; Mendoza, A.J.; Fernandez-Lorente, G.; Guisan, J.M.; Rocha-Martin, J. Stabilization of immobilized lipases by intense intramolecular cross-linking of their surfaces by using aldehyde-dextran polymers. Int. J. Mol. Sci. 2018, 19, 553–568. [Google Scholar]
- Dong, T.; Zhao, L.; Huang, Y.; Tan, X. Preparation of cross-linked aggregates of aminoacylase from Aspergillus melleus by using bovine serum albumin as an inert additive. Bioresour. Technol. 2010, 101, 6569–6571. [Google Scholar]
- Zhu, R.; Li, C.; Chen, C.; Xing, S.; Cai, Y.; Zeng, X.; He, L. Effect of cross-linked enzyme aggregate strategy on characterization of sn-1,3 extracellular lipase from Aspergillus niger GZUF36. Appl. Microbiol. Biotechnol. 2021, 105, 1925–1941. [Google Scholar] [PubMed]
- Shah, S.; Sharma, A.; Gupta, M.N. Preparation of cross-linked enzyme aggregates by using bovine serum albumin as a proteic feeder. Anal. Biochem. 2006, 351, 207–213. [Google Scholar]
- Wang, M.; Qi, W.; Jia, C.; Ren, Y.; Su, R.; He, Z. Enhancement of activity of cross-linked enzyme aggregates by a sugar-assisted precipitation strategy: Technical development and molecular mechanism. J. Biotechnol. 2011, 156, 30–38. [Google Scholar] [PubMed]
- Torabizadeh, H.; Tavakoli, M.; Safari, M. Immobilization of thermostable α-amylase from Bacillus licheniformis by cross-linked enzyme aggregates method using calcium and sodium ions as additives. J. Mol. Catal. B Enzym. 2014, 108, 13–20. [Google Scholar]
- Sinirlioglu, Z.A.; Sinirlioglu, D.; Akbas, F. Preparation and characterization of stable cross-linked enzyme aggregates of novel laccase enzyme from Shewanella putrefaciens and using malachite green decolorization. Bioresour. Technol. 2013, 146, 807–811. [Google Scholar]
- Bashir, F.; Asgher, M.; Hussain, F.; Randhawa, M.A. Development and characterization of cross-linked enzyme aggregates of thermotolerant alkaline protease from Bacillus licheniformis. Int. J. Biol. Macromol. 2018, 113, 944–951. [Google Scholar]
- Yu, C.Y.; Li, X.F.; Lou, W.Y.; Zong, M.H. Cross-linked enzyme aggregates of Mung bean epoxide hydrolases: A highly active, stable and recyclable biocatalyst for asymmetric hydrolysis of epoxides. J. Biotechnol. 2013, 166, 12–19. [Google Scholar]
- Tandjaoui, N.; Tassist, A.; Abouseoud, M.; Couvert, A.; Amrane, A. Preparation and characterization of cross-linked enzyme aggregates (CLEAs) of Brassica rapa peroxidase. Biocatal. Agric. Biotechnol. 2015, 4, 208–213. [Google Scholar]
- Ozacar, M.; Mehde, A.A.; Mehdi, W.A.; Ozacar, Z.Z.; Severgun, O. The novel multi cross-linked enzyme aggregates of protease, lipase, and catalase production from the sunflower seeds, characterization and application. Colloids Surf. B-Biointerfaces 2019, 173, 58–68. [Google Scholar]
- Kim, T.H.; Song, D.; Lee, J.S.; Yun, Y.M. Enhanced methane production from pretreatment of waste activated sludge by economically feasible biocatalysts. Energies 2023, 16, 552–562. [Google Scholar]
- Volschan Junior, I.; de Almeida, R.; Cammarota, M.C. A review of sludge pretreatment methods and co-digestion to boost biogas production and energy self-sufficiency in wastewater treatment plants. J. Water Process Eng. 2021, 40, 101857. [Google Scholar]
- Xin, X.; Hong, J.; He, J.; Qiu, W. An integrated approach for waste activated sludge management towards electric energy production/resource reuse. Bioresour. Technol. 2019, 274, 225–231. [Google Scholar] [PubMed]
- Wingfield, P. Protein precipitation using ammonium sulfate. Curr. Protoc. Protein Sci. 2001, Appendix 3, Appendix-3F. [Google Scholar]
- Morris, C.W. Protein Precipitation for the Purification of Therapeutic Proteins. Ph.D. Thesis, The Department of Biochemical Engineering, University College London, London, UK, 2019. [Google Scholar]
- Niu, L.J.; Zhang, H.; Wu, Z.K.; Wang, Y.B.; Liu, H.; Wu, X.L.; Wang, W. Modified TCA/acetone precipitation of plant proteins for proteomic analysis. PLoS ONE 2018, 13, e0202238. [Google Scholar]
- Zhang, Y.; Bottinelli, D.; Lisacek, F.; Luban, J.; Strambio-De-Castillia, C.; Varesio, E.; Hopfgartner, G. Optimization of human dendritic cell sample preparation for mass spectrometry-based proteomic studies. Anal. Biochem. 2015, 484, 40–50. [Google Scholar] [PubMed] [Green Version]
- Mateo, C.; Palomo, J.M.; van Langen, L.M.; van Rantwijk, F.; Sheldon, R.A. A new, mild cross-linking methodology to prepare cross-linked enzyme aggregates. Biotechnol. Bioeng. 2004, 86, 273–276. [Google Scholar]
- Hermanson, G.T. Antibody modification and conjugation. In Bioconjugate Techniques, 3rd ed.; Hermanson, G.T., Ed.; Academic Press: Boston, MA, USA, 2013; pp. 867–920. [Google Scholar]
- Scopes, R.K. Enzyme activity and assays. In Encyclopedia of Life Sciences; Wiley-Blackwell: Hoboken, NJ, USA, 2002. [Google Scholar]
- Nabarlatz, D.; Vondrysova, J.; Jenicek, P.; Stuber, F.; Font, J.; Fortuny, A.; Fabregat, A.; Bengoa, C. Hydrolytic enzymes in activated sludge: Extraction of protease and lipase by stirring and ultrasonication. Ultrason. Sonochem. 2010, 17, 923–931. [Google Scholar]
- Potty, V.H. Determination of proteins in the presence of phenols and pectins. Anal. Biochem. 1969, 29, 535–539. [Google Scholar]
- Pencreach, G.; Baratti, J.C. Hydrolysis of p-nitrophenyl palmitate in n-heptane by the Pseudomonas cepacia lipase: A simple test for the determination of lipase activity in organic media. Enzym. Microb. Technol. 1996, 18, 417–422. [Google Scholar]
- Ghose, T.K. Measurement of cellulase activities. Pure Appl. Chem. 1987, 59, 257–268. [Google Scholar]
- Juhnevica-Radenkova, K.; Kviesis, J.; Moreno, D.A.; Seglina, D.; Vallejo, F.; Valdovska, A.; Radenkovs, V. Highly-efficient release of ferulic acid from agro-industrial by-products via enzymatic hydrolysis with cellulose-degrading enzymes: Part I-the superiority of hydrolytic enzymes versus conventional hydrolysis. Foods 2021, 10, 782. [Google Scholar]
- Krah, D.; Ghattas, A.-K.; Wick, A.; Bröder, K.; Ternes, T.A. Micropollutant degradation via extracted native enzymes from activated sludge. Water Res. 2016, 95, 348–360. [Google Scholar]
- Asgari Lajayer, B.; Najafi, N.; Moghiseh, E.; Mosaferi, M.; Hadian, J. Effects of gamma irradiation on physicochemical and biological characteristics of wastewater effluent and sludge. Int. J. Environ. Sci. Technol. 2020, 17, 1021–1034. [Google Scholar]
- de Farias, V.L.; Monteiro, K.X.; Rodrigues, S.; Narciso Fernandes, F.A.; Saavedra Pinto, G.A. Comparison of Aspergillus niger spore production on potato dextrose agar (PDA) and crushed corncob medium. J. Gen. Appl. Microbiol. 2010, 56, 399–402. [Google Scholar]
- Zhang, W.; Alvarez-Gaitan, J.P.; Dastyar, W.; Saint, C.P.; Zhao, M.; Short, M.D. Value-added products derived from waste activated sludge: A biorefinery perspective. Water 2018, 10, 545–564. [Google Scholar]
- Goñi, I.; Garcia-Alonso, A.; Saura-Calixto, F. A starch hydrolysis procedure to estimate glycemic index. Nutr. Res. 1997, 17, 427–437. [Google Scholar]
- Butterworth, P.J.; Warren, F.J.; Grassby, T.; Patel, H.; Ellis, P.R. Analysis of starch amylolysis using plots for first-order kinetics. Carbohydr. Polym. 2012, 87, 2189–2197. [Google Scholar]
- Yao, Y. Use of Carbohydrate, Protein and Fat to Characterise Wastewater in Terms of Its Major Elemental Constituents and Energy. Master’s Thesis, The University of Manchester, Manchester, UK, 2014. [Google Scholar]
- Deepa, G.; Singh, V.; Naidu, K.A. A comparative study on starch digestibility, glycemic index and resistant starch of pigmented (‘Njavara’ and ‘Jyothi’) and a non-pigmented (‘IR 64’) rice varieties. J. Food Sci. Technol. 2010, 47, 644–649. [Google Scholar]
- Siddiqi, R.A.; Singh, T.P.; Rani, M.; Sogi, D.S.; Bhat, M.A. Diversity in grain, flour, amino acid composition, protein profiling, and proportion of total flour proteins of different wheat cultivars of north India. Front. Nutr. 2020, 7, 141. [Google Scholar] [PubMed]
- Vriens, L.; Nihoul, R.; Verachtert, H. Activated sludges as animal feed: A review. Biol. Wastes 1989, 27, 161–207. [Google Scholar]
- Schoevaart, R.; Wolbers, M.W.; Golubovic, M.; Ottens, M.; Kieboom, A.P.G.; van Rantwijk, F.; van der Wielen, L.A.M.; Sheldon, R.A. Preparation, optimization, and structures of cross-linked enzyme aggregates (CLEAs). Biotechnol. Bioeng. 2004, 87, 754–762. [Google Scholar]
- Nadar, S.S.; Muley, A.B.; Ladole, M.R.; Joshi, P.U. Macromolecular cross-linked enzyme aggregates (M-CLEAs) of α-amylase. Int. J. Biol. Macromol. 2016, 84, 69–78. [Google Scholar] [PubMed]
- Akyon, B.; Lipus, D.; Bibby, K. Glutaraldehyde inhibits biological treatment of organic additives in hydraulic fracturing produced water. Sci. Total Environ. 2019, 666, 1161–1168. [Google Scholar] [PubMed]
- Rupley, J.A.; Gratton, E.; Careri, G. Water and globular proteins. Trends Biochem. Sci. 1983, 8, 18–22. [Google Scholar] [CrossRef] [Green Version]
- Camisasca, G.; Iorio, A.; De Marzio, M.; Gallo, P. Structure and slow dynamics of protein hydration water. J. Mol. Liq. 2018, 268, 903–910. [Google Scholar] [CrossRef]
- Chaudhari, S.A.; Singhal, R.S. A strategic approach for direct recovery and stabilization of Fusarium sp. ICT SAC1 cutinase from solid state fermented broth by carrier free cross-linked enzyme aggregates. Int. J. Biol. Macromol. 2017, 98, 610–621. [Google Scholar] [PubMed]
- Kong, J.; Yu, S. Fourier transform infrared spectroscopic analysis of protein secondary structures. Acta Biochim. Biophys. Sin. 2007, 39, 549–559. [Google Scholar] [PubMed] [Green Version]
- Kristoffersen, K.A.; van Amerongen, A.; Böcker, U.; Lindberg, D.; Wubshet, S.G.; de Vogel-van den Bosch, H.; Horn, S.J.; Afseth, N.K. Fourier-transform infrared spectroscopy for monitoring proteolytic reactions using dry-films treated with trifluoroacetic acid. Sci. Rep. 2020, 10, 7844–7853. [Google Scholar]
- Li, B.; Shan, C.; Zhou, Q.; Fang, Y.; Wang, Y.; Xu, F.; Han, L.; Ibrahim, M.; Guo, L.; Xie, G.; et al. Synthesis, characterization, and antibacterial activity of cross-linked chitosan-glutaraldehyde. Mar. Drugs 2013, 11, 1534–1552. [Google Scholar]
- Hero, J.S.; Romero, C.M.; Pisa, J.H.; Perotti, N.I.; Olivaro, C.; Martinez, M.A. Designing cross-linked xylanase aggregates for bioconversion of agroindustrial waste biomass towards potential production of nutraceuticals. Int. J. Biol. Macromol. 2018, 111, 229–236. [Google Scholar]
- Durica-Mitic, S.; Göpel, Y.; Görke, B. Carbohydrate utilization in bacteria: Making the most out of sugars with the help of small regulatory RNAs. Microbiol. Spectr. 2018, 6. [Google Scholar] [CrossRef]
- Ma, Z.; Boye, J.I.; Simpson, B.K.; Prasher, S.O.; Monpetit, D.; Malcolmson, L. Thermal processing effects on the functional properties and microstructure of lentil, chickpea, and pea flours. Food Res. Int. 2011, 44, 2534–2544. [Google Scholar]
- Hill, G.A.; Macdonald, D.G.; Lang, X. a-Amylase inhibition and inactivation in barley malt during cold starch hydrolysis. Biotechnol. Lett. 1997, 19, 1139–1141. [Google Scholar]
- de la Rosa-Millan, J.; Orona-Padilla, J.L.; Flores-Moreno, V.M.; Serna-Saldivar, S.O. Effect of jet-cooking and hydrolyses with amylases on the physicochemical and in vitro digestion performance of whole chickpea flours. Int. J. Food Sci. Technol. 2020, 55, 690–701. [Google Scholar]
- Molobela, I.P.; Cloete, T.E.; Beukes, M. Protease and amylase enzymes for biofilm removal and degradation of extracellular polymeric substances (EPS) produced by Pseudomonas fluorescens bacteria. Afr. J. Microbiol. Res. 2010, 4, 1515–1524. [Google Scholar]
- Yu, G.H.; He, P.J.; Shao, L.M.; Lee, D. Extracellular enzymes in sludge flocs collected at 14 full-scale wastewater treatment plants. J. Chem. Technol. Biotechnol. 2008, 83, 1717–1725. [Google Scholar]
- Ahmed, R.Z.; Siddiqui, K.; Arman, M.; Ahmed, N. Characterization of high molecular weight dextran produced by Weissella cibaria CMGDEX3. Carbohydr. Polym. 2012, 90, 441–446. [Google Scholar] [PubMed]
- Merck KGaA. IR Spectrum Table & Chart. Available online: https://www.sigmaaldrich.com/GB/en/technical-documents/technical-article/analytical-chemistry/photometry-and-reflectometry/ir-spectrum-table#table-by-group (accessed on 27 May 2023).
- Baldino, L.; Concilio, S.; Cardea, S.; De Marco, I.; Reverchon, E. Complete glutaraldehyde elimination during chitosan hydrogel drying by SC-CO2 processing. J. Supercrit. Fluids 2015, 103, 70–76. [Google Scholar]
- Parida, U.K.; Nayak, A.K.; Binhani, B.K.; Nayak, P.L. Synthesis and characterization of chitosan-polyvinyl alcohol blended with Cloisite 30B for controlled release of the anticancer drug curcumin. J. Biomater. Nanobiotechnol. 2011, 2, 12. [Google Scholar]
- Motulsky, H. Analysis Checklist: Fitting a Model. Available online: https://www.graphpad.com/guides/prism/7/curve-fitting/reg_analysischeck_nonlin_completed.htm (accessed on 5 March 2023).







| Chemical Parameter, Model Coefficient and Statistics | CLEA Dose Rate (CLEA Suspension: Substrate) | |
|---|---|---|
| 1:100 v/v | 1:10 v/v | |
| sTOC: | ||
| C0 (mg/g TS) | 13.6 ± 0.22 | 15.5 ±2.16 |
| C∞ (mg/g TS) | 58.0 ±3.78 | 66.9 ± 5.08 |
| k (h−1) | 0.114 ± 0.014 | 0.263 ± 0.059 |
| Equation | C = 13.60 + 44.42 (1-e−0.114t) | C = 15.51 + 51.39 (1-e−0.263t) |
| R2 | >0.99 | 0.96 |
| p-value for normality test | >0.05 | >0.05 |
| Pass normality test (alpha = 0.05) | Yes | Yes |
| Reducing sugar: | ||
| C0 (mg/g TS) | 7.19 ± 0.12 | 4.05 ± 0.45 |
| C∞ (mg/g TS) | 42.4 ± 2.31 | 59.9 ± 9.39 |
| k (h−1) | 0.086 ± 0.008 | 0.102 ± 0.024 |
| Equation | C = 7.19 + 35.24 (1-e−0.086t) * | C = 4.05 + 55.82 (1-e−0.102t) |
| R2 | >0.99 | 0.99 |
| p-value for normality test | >0.05 | >0.05 |
| Pass normality test (alpha = 0.05) | Yes | Yes |
| Tyrosine: | ||
| C0 (mg/g TS) | 0.92 ± 0.02 | 1.34 ± 0.02 |
| C∞ (mg/g TS) | 4.47 ± 0.34 | 5.97 ± 0.08 |
| k (h−1) | 0.125 ± 0.018 | 0.287 ± 0.010 |
| Equation | C = 0.92 + 3.55 (1-e−0.125t) | C = 1.34 + 4.63 (1-e−0.287t) ** |
| R2 | >0.99 | >0.99 |
| p-value for normality test | <0.001 | >0.05 |
| Pass normality test (alpha = 0.05) | No *** | Yes |
| Sludge Type | γ-Irradiated WAS | Sonicated WAS |
|---|---|---|
| C0 (mg/g TS) | 5.61 ± 0.18 | 7.08 ± 0.06 |
| C∞ (mg/g TS) | 8.21 ± 0.23 | 10.27 ± 0.12 |
| k (h−1) | 0.521 ± 0.131 | 0.342 ± 0.031 |
| Equation | C = 5.61 + 2.60 (1-e−0.521t) * | C = 7.08 + 3.19 (1-e−0.342t) ** |
| R2 | 0.91 | >0.99 |
| p-value for normality test | >0.05 | >0.05 |
| Pass normality test (alpha = 0.05) | Yes | Yes |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Z.; Smith, S.R. Cross-Linked Enzyme Aggregate (CLEA) Preparation from Waste Activated Sludge. Microorganisms 2023, 11, 1902. https://doi.org/10.3390/microorganisms11081902
Liu Z, Smith SR. Cross-Linked Enzyme Aggregate (CLEA) Preparation from Waste Activated Sludge. Microorganisms. 2023; 11(8):1902. https://doi.org/10.3390/microorganisms11081902
Chicago/Turabian StyleLiu, Ziyi, and Stephen R. Smith. 2023. "Cross-Linked Enzyme Aggregate (CLEA) Preparation from Waste Activated Sludge" Microorganisms 11, no. 8: 1902. https://doi.org/10.3390/microorganisms11081902