

Efficacy of Trichoderma longibrachiatum Trichogin GA IV Peptaibol analogs against the Black Rot Pathogen Xanthomonas campestris pv. campestris and other Phytopathogenic Bacteria

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Trichogin GA IV Synthetic Analogs Used in This Work
2.2. Xcc Isolation and Identification
2.3. Antimicrobial Activity of Peptaibols
2.4. MIC and MBC Determinations
2.5. Peptide 4r Activity against Gram-Negative Bacterial Plant Pathogens and the Gram-Positive Bacterium B. subtilis
2.6. Tobacco Cell Culture Maintenance and Evans Blue Assay
2.7. Xcc Inoculation Assays
2.8. Statistical Analysis
3. Results
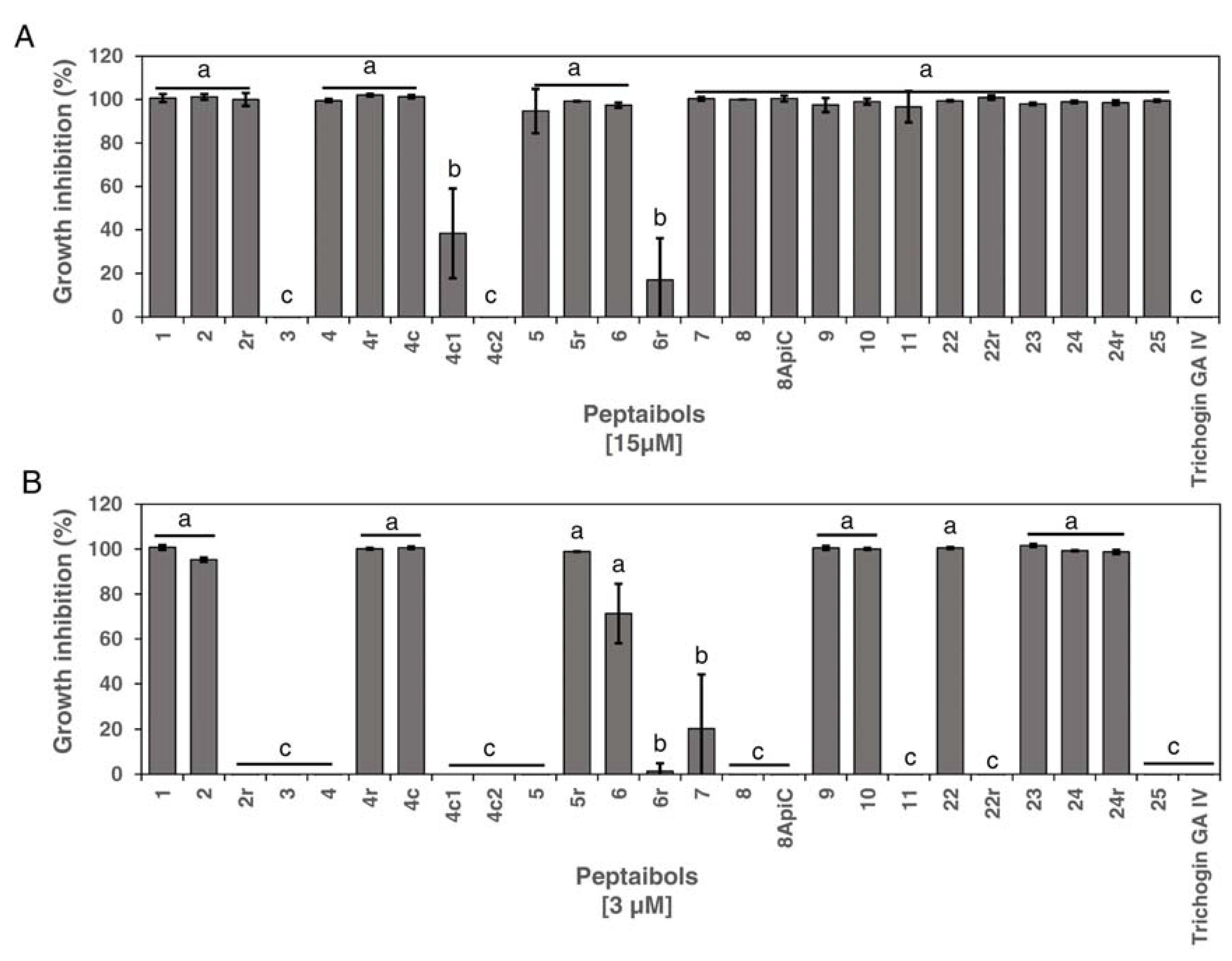
3.1. Antimicrobial Activity of Peptides against Xcc
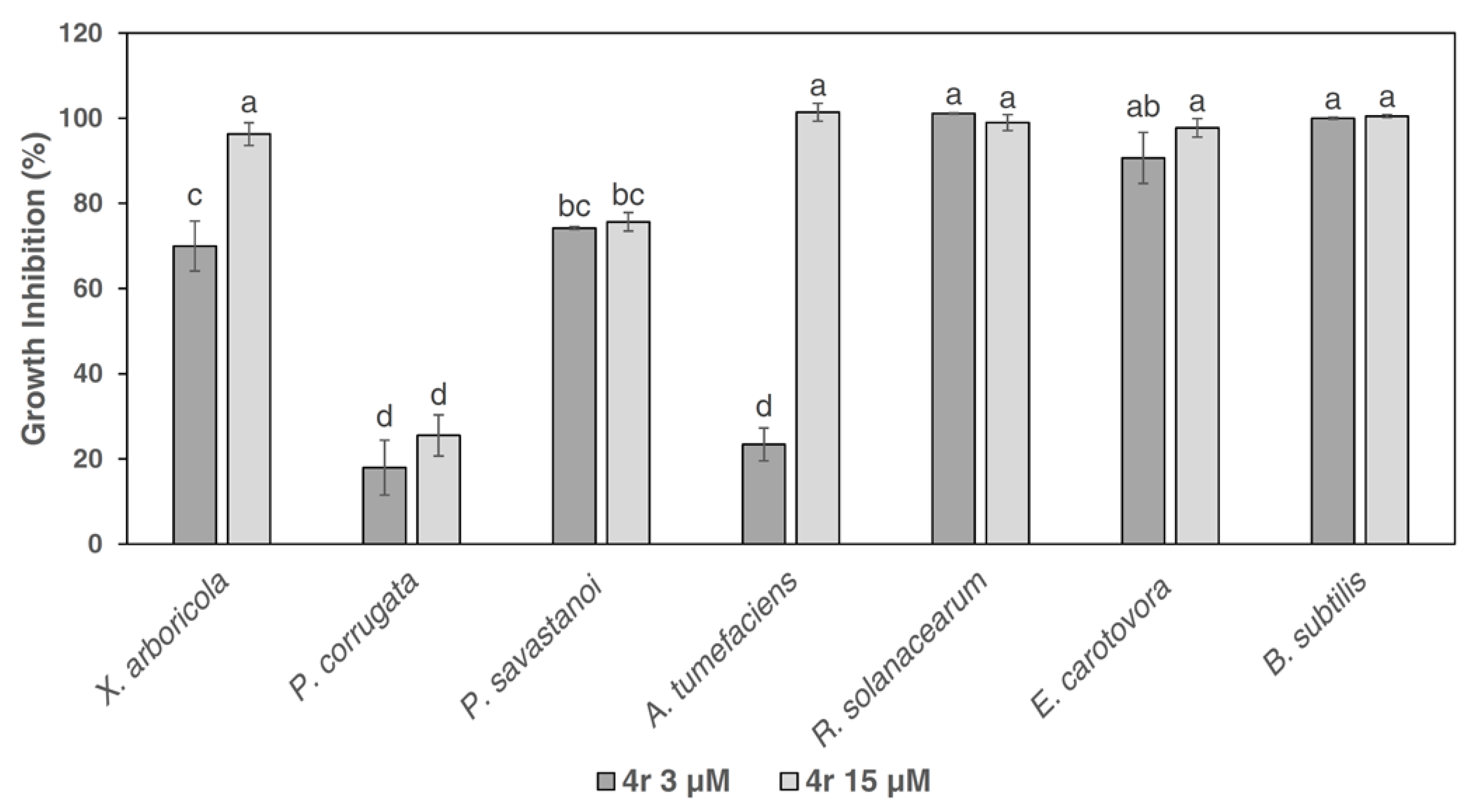
3.2. Growth Inhibition Activity of Peptide 4r towards Gram-Negative Bacterial Plant Pathogens and the Gram-Positive Bacterium B. subtilis
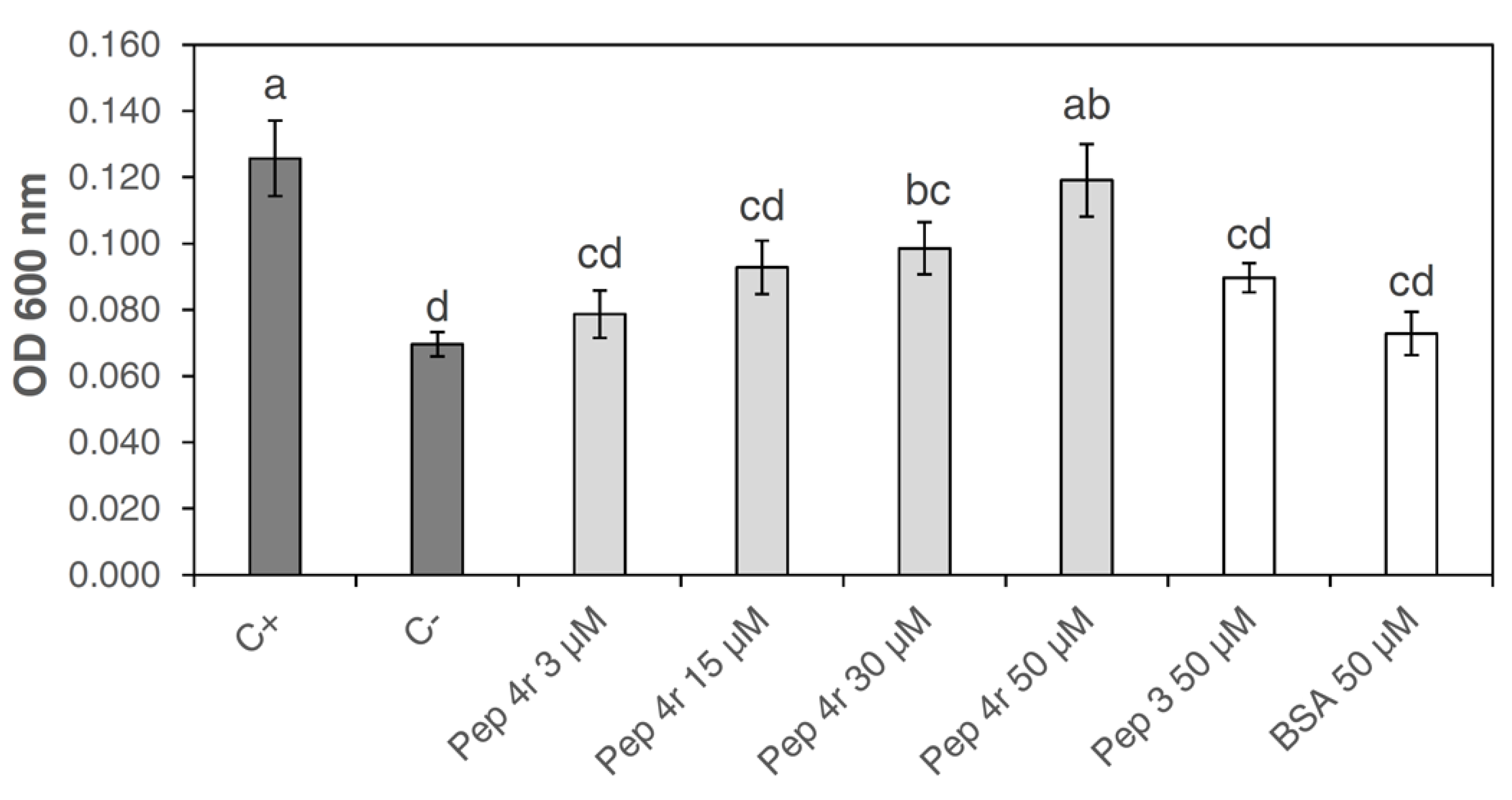
3.3. Cytotoxicity of Peptide 4r on Tobacco Cells
3.4. Effect of Peptide Treatment on Cauliflower Plants against Xcc Infection
4. Discussion
5. Patents
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Martins, P.M.M.; Merfa, M.V.; Takita, M.A.; de Souza, A.A. Persistence in Phytopathogenic Bacteria: Do We Know Enough? Front. Microbiol. 2018, 9, 1099. [Google Scholar] [CrossRef] [PubMed]
- Mansfield, J.; Genin, S.; Magori, S.; Citovsky, V.; Sriariyanum, M.; Ronald, P.; Dow, M.; Verdier, V.; Beer, S.V.; Machado, M.A.; et al. Top 10 plant pathogenic bacteria in molecular plant pathology. Mol. Plant Pathol. 2012, 13, 614–629. [Google Scholar] [CrossRef] [PubMed]
- Gupta, M.; Vikram, A.; Bharat, N. Black rot-A devastating disease of crucifers: A review. Agric. Rev. 2013, 34, 269. [Google Scholar] [CrossRef]
- Eichmeier, A.; Peňázová, E.; Pokluda, R.; Vicente, J.G. Detection of Xanthomonas campestris pv. campestris through a real-time PCR assay targeting the Zur gene and comparison with detection targeting the hrpF gene. Eur. J. Plant Pathol. 2019, 155, 891–902. [Google Scholar] [CrossRef]
- Mishra, S.; Arora, N.K. Evaluation of rhizospheric Pseudomonas and Bacillus as biocontrol tool for Xanthomonas campestris pv campestris. World J. Microbiol. Biotechnol. 2012, 28, 693–702. [Google Scholar] [CrossRef]
- Monteiro, L.; de Lima Ramos Mariano, R.; Souto-Maior, A.M. Antagonism of Bacillus spp. against Xanthomonas campestris pv. campestris. Braz. Arch. Biol. Technol. 2005, 48, 23–29. [Google Scholar] [CrossRef]
- Nuñez, A.M.P.; Rodríguez, G.A.A.; Monteiro, F.P.; Faria, A.F.; Silva, J.C.P.; Monteiro, A.C.A.; Carvalho, C.V.; Gomes, L.A.A.; Souza, R.M.; de Souza, J.T.; et al. Bio-based products control black rot (Xanthomonas campestris pv. campestris) and increase the nutraceutical and antioxidant components in kale. Sci. Rep. 2018, 8, 10199. [Google Scholar] [CrossRef]
- Montesinos, E. Antimicrobial peptides and plant disease control. FEMS Microbiol. Lett. 2007, 270, 1–11. [Google Scholar] [CrossRef]
- Breen, S.; Solomon, P.S.; Bedon, F.; Vincent, D. Surveying the potential of secreted antimicrobial peptides to enhance plant disease resistance. Front. Plant Sci. 2015, 6, 900. [Google Scholar] [CrossRef]
- Camó, C.; Bonaterra, A.; Badosa, E.; Baró, A.; Montesinos, L.; Montesinos, E.; Planas, M.; Feliu, L. Antimicrobial peptide KSL-W and analogues: Promising agents to control plant diseases. Peptides 2019, 112, 85–95. [Google Scholar] [CrossRef]
- Mariz-Ponte, N.; Regalado, L.; Gimranov, E.; Tassi, N.; Moura, L.; Gomes, P.; Tavares, F.; Santos, C.; Teixeira, C. A synergic potential of antimicrobial peptides against Pseudomonas syringae pv. actinidiae. Molecules 2021, 26, 1461. [Google Scholar] [CrossRef] [PubMed]
- Mendes, R.J.; Sario, S.; Luz, J.P.; Tassi, N.; Teixeira, C.; Gomes, P.; Tavares, F.; Santos, C. Evaluation of three antimicrobial peptides mixtures to control the phytopathogen responsible for fire blight disease. Plants 2021, 10, 2637. [Google Scholar] [CrossRef] [PubMed]
- Mendes, R.J.; Regalado, L.; Luz, J.P.; Tassi, N.; Teixeira, C.; Gomes, P.; Tavares, F.; Santos, C. In vitro evaluation of five antimicrobial peptides against the plant pathogen Erwinia amylovora. Biomolecules 2021, 11, 554. [Google Scholar] [CrossRef] [PubMed]
- Spohn, R.; Daruka, L.; Lázár, V.; Martins, A.; Vidovics, F.; Grézal, G.; Méhi, O.; Kintses, B.; Számel, M.; Jangir, P.K.; et al. Integrated evolutionary analysis reveals antimicrobial peptides with limited resistance. Nat. Commun. 2019, 10, 4538. [Google Scholar] [CrossRef]
- Zeilinger, S.; Gruber, S.; Bansal, R.; Mukherjee, P.K. Secondary metabolism in Trichoderma—Chemistry meets genomics. Fungal Biol. Rev. 2016, 30, 74–90. [Google Scholar] [CrossRef]
- Stoppacher, N.; Neumann, N.K.N.; Burgstaller, L.; Zeilinger, S.; Degenkolb, T.; Brückner, H.; Schuhmacher, R. The comprehensive peptaibiotics database. Chem. Biodivers. 2013, 10, 734–743. [Google Scholar] [CrossRef]
- Toniolo, C.; Crisma, M.; Formaggio, F.; Peggion, C.; Epand, R.F.; Epand, R.M. Lipopeptaibols, a novel family of membrane active, antimicrobial peptides. Cell. Mol. Life Sci. 2001, 58, 1179–1188. [Google Scholar] [CrossRef]
- Szekeres, A.; Leitgeb, B.; Kredics, L.; Antal, Z.; Hatvani, L.; Manczinger, L.; Vágvölgyi, C. Peptaibols and related peptaibiotics of Trichoderma. A review. Acta Microbiol. Immunol. Hung. 2005, 52, 137–168. [Google Scholar] [CrossRef]
- De Zotti, M.; Biondi, B.; Peggion, C.; Formaggio, F.; Park, Y.; Hahm, K.S.; Toniolo, C. Trichogin GA IV: A versatile template for the synthesis of novel peptaibiotics. Org. Biomol. Chem. 2012, 10, 1285–1299. [Google Scholar] [CrossRef]
- De Zotti, M.; Sella, L.; Bolzonello, A.; Gabbatore, L.; Peggion, C.; Bortolotto, A.; Elmaghraby, I.; Tundo, S.; Favaron, F. Targeted amino acid substitutions in a Trichoderma peptaibol confer activity against fungal plant pathogens and protect host tissues from Botrytis cinerea infection. Int. J. Mol. Sci. 2020, 21, 7521. [Google Scholar] [CrossRef]
- Baccelli, I.; Luti, S.; Bernardi, R.; Favaron, F.; de Zotti, M.; Sella, L. Water-soluble Trichogin GA IV-derived peptaibols protect tomato plants from Botrytis cinerea infection with limited impact on plant defenses. Front. Plant Sci. 2022, 13, 881961. [Google Scholar] [CrossRef] [PubMed]
- Sella, L.; Govind, R.; Caracciolo, R.; Quarantin, A.; Vu, V.V.; Tundo, S.; Nguyen, H.M.; Favaron, F.; Musetti, R.; de Zotti, M. Transcriptomic and ultrastructural analyses of Pyricularia oryzae treated with fungicidal peptaibol analogs of Trichoderma trichogin. Front. Microbiol. 2021, 12, 3083. [Google Scholar] [CrossRef] [PubMed]
- De Zotti, M.; Biondi, B.; Formaggio, F.; Toniolo, C.; Stella, L.; Park, Y.; Hahm, K.S. Trichogin GA IV: An antibacterial and protease-resistant peptide. J. Pept. Sci. 2009, 15, 615–619. [Google Scholar] [CrossRef] [PubMed]
- Jelušić, A.; Berić, T.; Mitrović, P.; Dimkić, I.; Stanković, S.; Marjanović-Jeromela, A.; Popović, T. New insights into the genetic diversity of Xanthomonas campestris pv. campestris isolates from winter oilseed rape in Serbia. Plant Pathol. 2021, 70, 35–49. [Google Scholar] [CrossRef]
- Chen, Y.L.; Lee, C.C.; Lin, Y.L.; Yin, K.M.; Ho, C.L.; Liu, T. Obtaining long 16S rDNA sequences using multiple primers and its application on dioxin-containing samples. BMC Bioinform. 2015, 16 (Suppl. 18), S13. [Google Scholar] [CrossRef]
- Park, Y.J.; Lee, B.M.; Ho-Hahn, J.; Lee, G.B.; Park, D.S. Sensitive and specific detection of Xanthomonas campestris pv campestris by PCR using species-specific primers based on hrpF gene sequences. Microbiol. Res. 2004, 159, 419–423. [Google Scholar] [CrossRef]
- Ruiz, C.; Nadal, A.; Montesinos, E.; Pla, M. Novel Rosaceae plant elicitor peptides as sustainable tools to control Xanthomonas arboricola pv. pruni in Prunus spp. Mol. Plant Pathol. 2018, 19, 418–431. [Google Scholar] [CrossRef]
- Mustafa, N.R.; de Winter, W.; van Iren, F.; Verpoorte, R. Initiation, growth and cryopreservation of plant cell suspension cultures. Nat. Protoc. 2011, 6, 715–742. [Google Scholar] [CrossRef]
- Castro-Concha, L.A.; Escobedo, R.M.; de Miranda-Ham, M.L. Measurement of cell viability in in vitro cultures. Methods Mol. Biol. 2006, 318, 71–76. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022. [Google Scholar]
- Mohamed-Benkada, M.; François Pouchus, Y.; Vérité, P.; Pagniez, F.; Caroff, N.; Ruiz, N. Identification and biological activities of long-chain peptaibols produced by a marine-derived strain of Trichoderma longibrachiatum. Chem. Biodivers. 2016, 13, 521–530. [Google Scholar] [CrossRef]
- Zhang, Y.Q.; Zhang, S.; Sun, M.L.; Su, H.N.; Li, H.Y.; Kun, L.; Zhang, Y.Z.; Chen, X.L.; Cao, H.Y.; Song, X.Y. Antibacterial activity of peptaibols from Trichoderma longibrachiatum SMF2 against gram-negative Xanthomonas oryzae pv. oryzae, the causal agent of bacterial leaf blight on rice. Front. Microbiol. 2022, 13, 4032. [Google Scholar] [CrossRef] [PubMed]
- Heredia-Ponce, Z.; de Vicente, A.; Cazorla, F.M.; Gutiérrez-Barranquero, J.A. Beyond the wall: Exopolysaccharides in the biofilm lifestyle of pathogenic and beneficial plant-associated Pseudomonas. Microorganisms 2021, 9, 445. [Google Scholar] [CrossRef] [PubMed]
- Licciardello, G.; Caruso, A.; Bella, P.; Gheleri, R.; Strano, C.P.; Anzalone, A.; Trantas, E.A.; Sarris, P.F.; Almeida, N.F.; Catara, V. The LuxR regulators PcoR and RfiA co-regulate antimicrobial peptide and alginate production in Pseudomonas corrugata. Front. Microbiol. 2018, 9, 521. [Google Scholar] [CrossRef]
- Hentzer, M.; Teitzel, G.M.; Balzer, G.J.; Heydorn, A.; Molin, S.; Givskov, M.; Parsek, M.R. Alginate overproduction affects Pseudomonas aeruginosa biofilm structure and function. J. Bacteriol. 2001, 183, 5395–5401. [Google Scholar] [CrossRef] [PubMed]
- Badosa, E.; Ferre, R.; Planas, M.; Feliu, L.; Besalú, E.; Cabrefiga, J.; Bardají, E.; Montesinos, E. A library of linear undecapeptides with bactericidal activity against phytopathogenic bacteria. Peptides 2007, 28, 2276–2285. [Google Scholar] [CrossRef] [PubMed]
- Montesinos, E.; Badosa, E.; Cabrefiga, J.; Planas, M.; Feliu, L.; Bardají, E. Antimicrobial Peptides for Plant Disease Control. From Discovery to Application; Rajasekaran, K., Cary, J., Jaynes, J., Montesinos, E., Eds.; American Chemical Society: Washington, DC, USA, 2012; Volume 1095, ISBN 9780841227484. [Google Scholar]
- Makovitzki, A.; Viterbo, A.; Brotman, Y.; Chet, I.; Shai, Y. Inhibition of fungal and bacterial plant pathogens in vitro and in planta with ultrashort cationic lipopeptides. Appl. Environ. Microbiol. 2007, 73, 6629–6636. [Google Scholar] [CrossRef] [PubMed]
- Baró, A.; Mora, I.; Montesinos, L.; Montesinos, E. Differential susceptibility of Xylella fastidiosa strains to synthetic bactericidal peptides. Phytopathology 2020, 110, 1018–1026. [Google Scholar] [CrossRef]
- Auten, R.L.; Davis, J.M. Oxygen toxicity and reactive oxygen species: The devil is in the details. Pediatr. Res. 2009, 66, 121–127. [Google Scholar] [CrossRef]
- Jelušić, A.; Popović, T.; Dimkić, I.; Mitrović, P.; Peeters, K.; Višnjevec, A.M.; Tavzes, Č.; Berić, T. Changes in the winter oilseed rape microbiome afected by Xanthomonas campestris pv. campestris and biocontrol potential of the indigenous Bacillus and Pseudomonas isolates. Biol. Control 2021, 160, 104695. [Google Scholar] [CrossRef]
- Papaianni, M.; Ricciardelli, A.; Fulgione, A.; D’Errico, G.; Zoina, A.; Lorito, M.; Woo, S.L.; Vinale, F.; Capparelli, R. Antibiofilm activity of a Trichoderma metabolite against Xanthomonas campestris pv. campestris, alone and in association with a phage. Microorganisms 2020, 8, 620. [Google Scholar] [CrossRef]
- Cerutti, A.; Jauneau, A.; Auriac, M.C.; Lauber, E.; Martinez, Y.; Chiarenza, S.; Leonhardt, N.; Berthomé, R.; Noël, L.D. Immunity at cauliflower hydathodes controls systemic infection by Xanthomonas campestris pv campestris. Plant Physiol. 2017, 174, 700–716. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peptide | Sequence | Synthesis |
|---|---|---|
| Trichogin GA IV | nOct-Aib-Gly-Leu-Aib-Gly-Gly-Leu-Aib-Gly-Ile-Lol | |
| 1 | nOct-Aib-Lys-Leu-Aib-Lys-Gly-Leu-Aib-Gly-Ile-Lol | [20] |
| 2 | nOct-Aib-Lys-Leu-Aib-Gly-Gly-Leu-Aib-Gly-Ile-Lol | [20] |
| 2r | nOct-Aib-Lys-Leu-Aib-Gly-Gly-Leu-Aib-Gly-Ile-Leu-NH2 | [22] |
| 3 | nOct-Aib-Gly-Leu-Aib-Lys-Gly-Leu-Aib-Gly-Ile-Lol | [20] |
| 4 | nOct-Aib-Gly-Leu-Aib-Lys-Lys-Leu-Aib-Gly-Ile-Lol | [20] |
| 4r | nOct-Aib-Gly-Leu-Aib-Lys-Lys-Leu-Aib-Gly-Ile-Leu-NH2 | [20] |
| 4c | nOct-Aib-Lys-Lys-Leu-Aib-Gly-Ile-Lol | [20] |
| 4c1 | nOct-Aib-Gly-Leu-Aib-Lys-Lys-Leu-Leu-NH2 | [20] |
| 4c2 | nOct-Aib-Lys-Lys-Leu-Aib-Gly-Ile-Leu-NH2 | [20] |
| 5 | nOct-Aib-Lys-Leu-Aib-Lys-Gly-Leu-Aib-Lys-Ile-Lol | [22] |
| 5r | nOct-Aib-Lys-Leu-Aib-Lys-Gly-Leu-Aib-Lys-Ile-Leu-NH2 | This work |
| 6 | nOct-Aib-Gly-Leu-Aib-Lys-Aib-Leu-Aib-Gly-Ile-Lol | [20] |
| 6r | nOct-Aib-Gly-Leu-Aib-Lys-Aib-Leu-Aib-Gly-Ile-Leu-NH2 | [21] |
| 7 | nOct-Aib-Lys-Leu-Aib-Gly-Lys-Leu-Aib-Gly-Ile-Lol | [20] |
| 8 | nOct-Aib-Gly-Leu-Aib-Gly-Gly-Leu-Api-Gly-Ile-Lol | [20] |
| 8ApiC | nOct-Aib-Gly-Leu-Aib-Gly-Gly-Leu-Api-Ile-Lol | [22] |
| 9 | N3Ac-Aib-Lys-Leu-Aib-Lys-Lys-Leu-Aib-Lys-Ile-Lol | This work |
| 10 | nOct-Aib-Lys-Leu-Aib-Lys-Lys-Leu-Aib-Lys-Ile-Leu-Aib-Lys-Leu-Aib-Lys-Lys-Leu-Aib-Lys-Ile-Lol | This work |
| 11 | nOct-Toac-Gly-Leu-Aib-Gly-Gly-Leu-Aib-Arg-Ile-Lol | This work |
| 22 | nOct-Aib-Gly-Leu-Aib-Gly-Lys-Leu-Aib-Gly-Ile-Lol | [22] |
| 22r | nOct-Aib-Gly-Leu-Aib-Gly-Lys-Leu-Aib-Gly-Ile-Leu-NH2 | [22] |
| 23 | nOct-Aib-Lys-Leu-Aib-Lys-Lys-Leu-Aib-Lys-Ile-Lol | This work |
| 24 | nOct-Aib-Lys-Leu-Aib-Lys-Lys-Leu-Aib-Gly-Ile-Lol | This work |
| 24r | nOct-Aib-Lys-Leu-Aib-Lys-Lys-Leu-Aib-Gly-Ile-Leu-NH2 | This work |
| 25 | nOct-Aib-Gly-Leu-Aib-Gly-Gly-Leu-Aib-Lys-Ile-Lol | [21] |
| Peptides | 0.5 µM | 1 µM | 2 µM | 3 µM |
|---|---|---|---|---|
| 2 | - | - | - | MIC |
| 3 | - | - | - | - |
| 4r | - | - | MIC | MBC |
| 4c | - | - | - | MIC |
| 5r | - | - | MIC and MBC | |
| 9 | - | MIC | MBC | |
| 10 | - | - | MIC and MBC | |
| 22 | - | - | MIC | - |
| 23 | - | MIC and MBC | ||
| 24 | - | MIC | MBC | |
| 24r | - | MIC | MBC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Caracciolo, R.; Sella, L.; De Zotti, M.; Bolzonello, A.; Armellin, M.; Trainotti, L.; Favaron, F.; Tundo, S. Efficacy of Trichoderma longibrachiatum Trichogin GA IV Peptaibol analogs against the Black Rot Pathogen Xanthomonas campestris pv. campestris and other Phytopathogenic Bacteria. Microorganisms 2023, 11, 480. https://doi.org/10.3390/microorganisms11020480
Caracciolo R, Sella L, De Zotti M, Bolzonello A, Armellin M, Trainotti L, Favaron F, Tundo S. Efficacy of Trichoderma longibrachiatum Trichogin GA IV Peptaibol analogs against the Black Rot Pathogen Xanthomonas campestris pv. campestris and other Phytopathogenic Bacteria. Microorganisms. 2023; 11(2):480. https://doi.org/10.3390/microorganisms11020480
Chicago/Turabian StyleCaracciolo, Rocco, Luca Sella, Marta De Zotti, Angela Bolzonello, Marco Armellin, Livio Trainotti, Francesco Favaron, and Silvio Tundo. 2023. "Efficacy of Trichoderma longibrachiatum Trichogin GA IV Peptaibol analogs against the Black Rot Pathogen Xanthomonas campestris pv. campestris and other Phytopathogenic Bacteria" Microorganisms 11, no. 2: 480. https://doi.org/10.3390/microorganisms11020480