

The Anti-Amoebic Activity of a Peptidomimetic against Acanthamoeba castellanii



 , ,
, ,  and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Acanthamoeba Culturing and Test Compounds
2.2. Amoebicidal Assay
2.3. Amoebistatic Assay
2.4. Encystation Assay
2.5. Excystment Assay
2.6. Lysis of Horse Red Blood Cells (RBCs)
2.7. Statistical Analyses
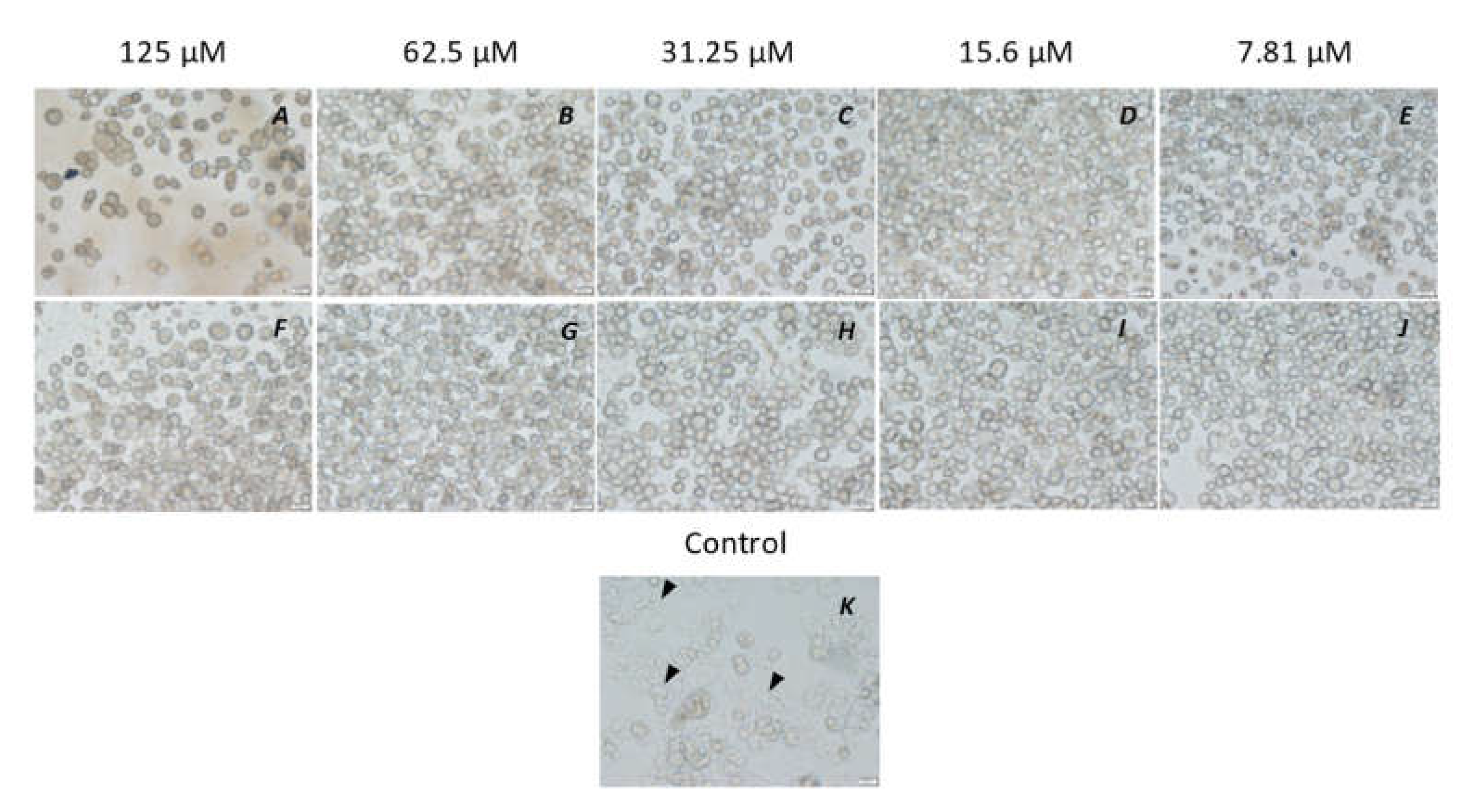
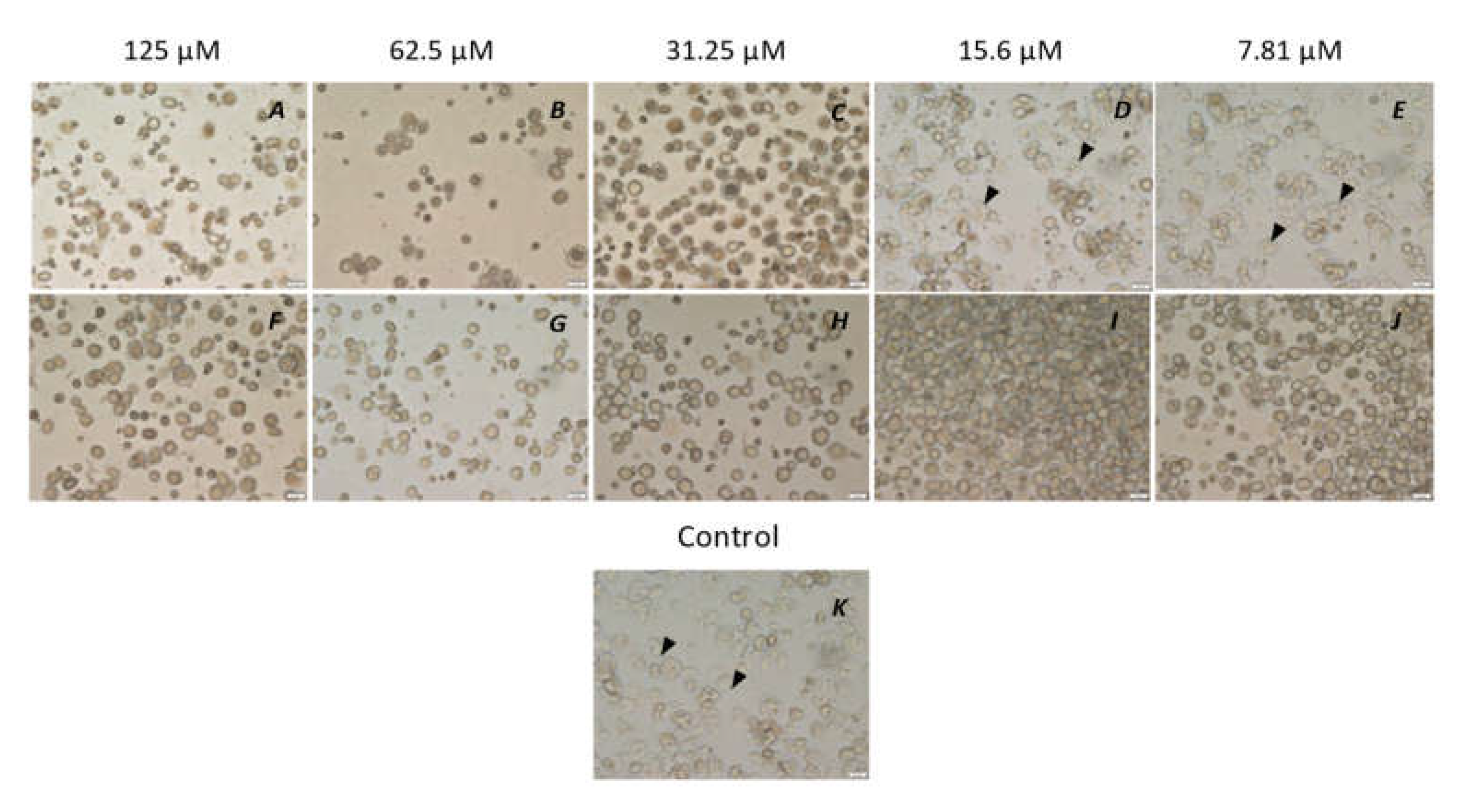
3. Results
3.1. Amoebicidal Assay
3.2. Amoebistatic Assay
3.3. Encystation Assay
3.4. Excystation Assay
3.5. Haemolysis Assay
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Siddiqui, R.; Khan, N.A. Biology and pathogenesis of Acanthamoeba. Parasites Vectors 2012, 5, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carnt, N.; Hoffman, J.M.; Verma, S.; Hau, S.; Radford, C.F.; Minassian, D.C.; Dart, J.K.G. Acanthamoeba keratitis: Confirmation of the UK outbreak and a prospective case-control study identifying contributing risk factors. Br. J. Ophthalmol. 2018, 102, 1621–1628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clarke, B.; Sinha, A.; Parmar, D.N.; Sykakis, E. Advances in the diagnosis and treatment of Acanthamoeba keratitis. J. Ophthalmol. 2012, 2012, 484892. [Google Scholar] [CrossRef] [Green Version]
- Szentmary, N.; Daas, L.; Shi, L.; Laurik, K.L.; Lepper, S.; Milioti, G.; Seitz, B. Acanthamoeba keratitis—Clinical signs, differential diagnosis and treatment. J. Curr. Ophthalmol. 2019, 31, 16–23. [Google Scholar] [CrossRef] [PubMed]
- List, W.; Glatz, W.; Riedl, R.; Mossboeck, G.; Steinwender, G.; Wedrich, A. Evaluation of Acanthamoeba keratitis cases in a tertiary medical care centre over 21 years. Sci. Rep. 2021, 11, 1036. [Google Scholar] [CrossRef] [PubMed]
- Naranjo, A.; Martinez, J.D.; Miller, D.; Tonk, R.; Amescua, G. Systemic miltefosine as an adjunct treatment of progressive Acanthamoeba keratitis. Ocul. Immunol. Inflamm. 2021, 29, 1576–1584. [Google Scholar] [CrossRef]
- Kosrirukvongs, P.; Wanachiwanawin, D.; Visvesvara, G.S. Treatment of Acanthamoeba keratitis with chlorhexidine. Ophthalmology 1999, 106, 798–802. [Google Scholar] [CrossRef]
- Fanselow, N.; Sirajuddin, N.; Yin, X.T.; Huang, A.J.W.; Stuart, P.M. Acanthamoeba keratitis, pathology, diagnosis and treatment. Pathogens 2021, 10, 323. [Google Scholar] [CrossRef]
- Dart, J.K.; Saw, V.P.; Kilvington, S. Acanthamoeba keratitis: Diagnosis and treatment update 2009. Am. J. Ophthalmol. 2009, 148, 487–499.e482. [Google Scholar] [CrossRef]
- Dutta, D.; Cole, N.; Kumar, N.; Willcox, M.D. Broad spectrum antimicrobial activity of melimine covalently bound to contact lenses. Investig. Ophthalmol. Vis. Sci. 2013, 54, 175–182. [Google Scholar] [CrossRef]
- Huan, Y.; Kong, Q.; Mou, H.; Yi, H. Antimicrobial peptides: Classification, design, application and research progress in multiple fields. Front. Microbiol. 2020, 11, 2559. [Google Scholar] [CrossRef]
- Antimicrobial Peptide Database. Available online: https://aps.unmc.edu/ (accessed on 4 November 2022).
- Wang, G.; Li, X.; Wang, Z. APD3: The antimicrobial peptide database as a tool for research and education. Nucleic Acids Res. 2016, 44, D1087–D1093. [Google Scholar] [CrossRef] [Green Version]
- Kuppusamy, R.; Willcox, M.; Black, D.S.; Kumar, N. Short cationic peptidomimetic antimicrobials. Antibiotics 2019, 8, 44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Helmerhorst, E.J.; Reijnders, I.M.; van’t Hof, W.; Simoons-Smit, I.; Veerman, E.C.; Amerongen, A.V. Amphotericin B- and fluconazole-resistant Candida spp., Aspergillus fumigatus, and other newly emerging pathogenic fungi are susceptible to basic antifungal peptides. Antimicrob. Agents Chemother. 1999, 43, 702–704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feder, R.; Dagan, A.; Mor, A. Structure-activity relationship study of antimicrobial dermaseptin S4 showing the consequences of peptide oligomerization on selective cytotoxicity. J. Biol. Chem. 2000, 275, 4230–4238. [Google Scholar] [CrossRef] [Green Version]
- Meiller, T.F.; Hube, B.; Schild, L.; Shirtliff, M.E.; Scheper, M.A.; Winkler, R.; Ton, A.; Jabra-Rizk, M.A. A novel immune evasion strategy of Candida albicans: Proteolytic cleavage of a salivary antimicrobial peptide. PLoS ONE 2009, 4, e5039. [Google Scholar] [CrossRef] [Green Version]
- Potempa, J.; Pike, R.N. Corruption of innate immunity by bacterial proteases. J. Innate Immun. 2009, 1, 70–87. [Google Scholar] [CrossRef] [Green Version]
- Vijay, A.K.; Bandara, M.; Zhu, H.; Willcox, M.D. Protamine as a potential amoebicidal agent for contact lens disinfection. Optom. Vis. Sci. 2013, 90, 119–124. [Google Scholar] [CrossRef]
- Schuster, F.L.; Jacob, L.S. Effects of magainins on ameba and cyst stages of Acanthamoeba polyphaga. Antimicrob. Agents Chemother. 1992, 36, 1263–1271. [Google Scholar] [CrossRef] [Green Version]
- Feldman, S.T.; Speaker, M.; Cleveland, P. Effect of magainins on Acanthamoeba castellanii. Rev. Infect. Dis. 1991, 13 (Suppl. S5), S439. [Google Scholar] [CrossRef]
- Polat, Z.A.; Savage, P.B.; Genberg, C. In vitro amoebicidal activity of a ceragenin, cationic steroid antibiotic-13, against Acanthamoeba castellanii and its cytotoxic potential. J. Ocul. Pharm. 2011, 27, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Sacramento, R.S.; Martins, R.M.; Miranda, A.; Dobroff, A.S.; Daffre, S.; Foronda, A.S.; De Freitas, D.; Schenkman, S. Differential effects of alpha-helical and beta-hairpin antimicrobial peptides against Acanthamoeba castellanii. Parasitology 2009, 136, 813–821. [Google Scholar] [CrossRef] [PubMed]
- Schlusselhuber, M.; Humblot, V.; Casale, S.; Methivier, C.; Verdon, J.; Leippe, M.; Berjeaud, J.M. Potent antimicrobial peptides against Legionella pneumophila and its environmental host, Acanthamoeba castellanii. Appl. Microbiol. Biotechnol. 2015, 99, 4879–4891. [Google Scholar] [CrossRef]
- Kennedy, S.M.; Deshpande, P.; Gallagher, A.G.; Horsburgh, M.J.; Allison, H.E.; Kaye, S.B.; Wellings, D.A.; Williams, R.L. Amoebicidal activity of poly-epsilon-lysine functionalized hydrogels. Investig. Ophthalmol. Vis. Sci. 2022, 63, 11. [Google Scholar] [CrossRef] [PubMed]
- Akbar, N.; Kaman, W.E.; Sarink, M.; Nazmi, K.; Bikker, F.J.; Khan, N.A.; Siddiqui, R. Novel Antiamoebic tyrocidine-derived peptide against brain-eating amoebae. ACS Omega 2022, 7, 28797–28805. [Google Scholar] [CrossRef] [PubMed]
- Fiori, P.L.; Mattana, A.; Dessi, D.; Conti, S.; Magliani, W.; Polonelli, L. In vitro acanthamoebicidal activity of a killer monoclonal antibody and a synthetic peptide. J. Antimicrob. Chemother. 2006, 57, 891–898. [Google Scholar] [CrossRef] [Green Version]
- De Carvalho Climaco, M.; de Oliveira, Y.L.M.; Ramos, A.C.S.; Ramos-de-Souza, J.; Silva, A.; Jain, S.; Rott, M.B.; Scher, R.; Correa, C.B.; Barbosa, A.A.T.; et al. Nisin induces cell-cycle arrest in free-living amoebae Acanthamoeba castellanii. Acta Parasitol. 2022, 67, 511–517. [Google Scholar] [CrossRef]
- Otri, A.M.; Mohammed, I.; Abedin, A.; Cao, Z.; Hopkinson, A.; Panjwani, N.; Dua, H.S. Antimicrobial peptides expression by ocular surface cells in response to Acanthamoeba castellanii: An in vitro study. Br. J. Ophthalmol. 2010, 94, 1523–1527. [Google Scholar] [CrossRef] [Green Version]
- Méndez-Samperio, P. Peptidomimetics as a new generation of antimicrobial agents: Current progress. Infect. Drug Resist. 2014, 7, 229–237. [Google Scholar] [CrossRef] [Green Version]
- Luo, G.; Zhang, J.; Wang, H.; Sun, Y.; Cheng, B.; Xu, Z.; Zhang, Y.; Li, H.; Lu, W.; Nemeth, E.; et al. Human defensin-inspired discovery of peptidomimetic antibiotics. Proc. Natl. Acad. Sci. USA 2022, 119, e2117283119. [Google Scholar] [CrossRef]
- Ibrahim, M.A.; Panda, S.S.; Oliferenko, A.A.; Oliferenko, P.V.; Girgis, A.S.; Elagawany, M.; Küçükbay, F.Z.; Panda, C.S.; Pillai, G.G.; Samir, A.; et al. Macrocyclic peptidomimetics with antimicrobial activity: Synthesis, bioassay, and molecular modeling studies. Org. Biomol. Chem. 2015, 13, 9492–9503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Chio, C.; Previti, S.; Amendola, G.; Cosconati, S.; Schirmeister, T.; Zappalà, M.; Ettari, R. Development of novel benzodiazepine-based peptidomimetics as inhibitors of rhodesain from Trypanosoma brucei rhodesiense. ChemMedChem 2020, 15, 995–1001. [Google Scholar] [CrossRef]
- Kuppusamy, R.; Yasir, M.; Yee, E.; Willcox, M.; Black, D.S.; Kumar, N. Guanidine functionalized anthranilamides as effective antibacterials with biofilm disruption activity. Org. Biomol. Chem. 2018, 16, 5871–5888. [Google Scholar] [CrossRef] [PubMed]
- Browne, K.; Kuppusamy, R.; Chen, R.; Willcox, M.D.P.; Walsh, W.R.; Black, D.S.; Kumar, N. Bioinspired polydopamine coatings facilitate attachment of antimicrobial peptidomimetics with broad-spectrum antibacterial activity. Int. J. Mol. Sci. 2022, 23, 2952. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Yu, T.T.; Kuppusamy, R.; Hassan, M.M.; Alghalayini, A.; Cranfield, C.G.; Willcox, M.D.P.; Black, D.S.; Kumar, N. Cholic acid-based antimicrobial peptide mimics as antibacterial agents. Int. J. Mol. Sci. 2022, 23, 4623. [Google Scholar] [CrossRef] [PubMed]
- Benjamin, A.B.; Moule, M.G.; Didwania, M.K.; Hardy, J.; Saenkham-Huntsinger, P.; Sule, P.; Nielsen, J.E.; Lin, J.S.; Contag, C.H.; Barron, A.E.; et al. Efficacy of cathelicidin-mimetic antimicrobial peptoids against Staphylococcus aureus. Microbiol. Spectr. 2022, 10, e0053422. [Google Scholar] [CrossRef]
- Patel, A.; Hammersmith, K. Contact lens-related microbial keratitis: Recent outbreaks. Curr. Opin. Ophthalmol. 2008, 19, 302–306. [Google Scholar] [CrossRef]
- Verani, J.R.; Lorick, S.A.; Yoder, J.S.; Beach, M.J.; Braden, C.R.; Roberts, J.M.; Conover, C.S.; Chen, S.; McConnell, K.A.; Chang, D.C.; et al. National outbreak of Acanthamoeba keratitis associated with use of a contact lens solution, United States. Emerg. Infect. Dis. 2009, 15, 1236–1242. [Google Scholar] [CrossRef]
- Joslin, C.E.; Tu, E.Y.; Shoff, M.E.; Booton, G.C.; Fuerst, P.A.; McMahon, T.T.; Anderson, R.J.; Dworkin, M.S.; Sugar, J.; Davis, F.G.; et al. The association of contact lens solution use and Acanthamoeba keratitis. Am. J. Ophthalmol. 2007, 144, 169–180. [Google Scholar] [CrossRef] [Green Version]
- Kilvington, S.; Heaselgrave, W.; Lally, J.M.; Ambrus, K.; Powell, H. Encystment of Acanthamoeba during incubation in multipurpose contact lens disinfectant solutions and experimental formulations. Eye Contact Lens. 2008, 34, 133–139. [Google Scholar] [CrossRef]
- Radford, C.F.; Lehmann, O.J.; Dart, J.K.; for the National Acanthamoeba Keratitis Study Group. Acanthamoeba keratitis: Multicentre survey in England 1992-6. Br. J. Ophthalmol. 1998, 82, 1387–1392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsu, C.-C.; Kuo, Y.-S.; Lin, P.-Y.; Chen, K.-H. Overnight orthokeratology-associated Acanthamoeba keratitis at a tertiary referral hospital in Taiwan: A retrospective case-control study. J. Chin. Med. Assoc. 2022, 85, 381–387. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Wang, Z.; Qu, J.; Zhang, Y.; Sun, X. Acanthamoeba keratitis related to contact lens use in a tertiary hospital in China. BMC Ophthalmol. 2019, 19, 202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Narasimhan, S.; Madhavan, H.N.; Therese, K.L. Development and application of an in vitro susceptibility test for Acanthamoeba species isolated from keratitis to polyhexamethylene biguanide and chlorhexidine. Cornea 2002, 21, 203–205. [Google Scholar] [CrossRef]
- Baig, A.M.; Iqbal, J.; Khan, N.A. In vitro efficacies of clinically available drugs against growth and viability of an Acanthamoeba castellanii keratitis isolate belonging to the T4 genotype. Antimicrob. Agents Chemother. 2013, 57, 3561–3567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anwar, A.; Chi Fung, L.; Anwar, A.; Jagadish, P.; Numan, A.; Khalid, M.; Shahabuddin, S.; Siddiqui, R.; Khan, N.A. Effects of shape and size of cobalt phosphate nanoparticles against Acanthamoeba castellanii. Pathogens 2019, 8, 260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aqeel, Y.; Siddiqui, R.; Iftikhar, H.; Khan, N.A. The effect of different environmental conditions on the encystation of Acanthamoeba castellanii belonging to the T4 genotype. Exp. Parasitol. 2013, 135, 30–35. [Google Scholar] [CrossRef]
- Sabir, S.; Yu, T.T.; Kuppusamy, R.; Almohaywi, B.; Iskander, G.; Das, T.; Willcox, M.D.P.; Black, D.S.; Kumar, N. Novel seleno- and thio-urea containing dihydropyrrol-2-one analogues as antibacterial agents. Antibiotics 2021, 10, 321. [Google Scholar] [CrossRef]
- Yasir, M.; Dutta, D.; Willcox, M.D.P. Mode of action of the antimicrobial peptide Mel4 is independent of Staphylococcus aureus cell membrane permeability. PLoS ONE 2019, 14, e0215703. [Google Scholar] [CrossRef] [Green Version]
- Schuster, F.L.; Visvesvara, G.S. Opportunistic amoebae: Challenges in prophylaxis and treatment. Drug Resist. Updates 2004, 7, 41–51. [Google Scholar] [CrossRef]
- Elder, M.; Kilvington, S.; Dart, J. A clinicopathologic study of in vivo sensitivity testing and Acanthamoeba keratitis. Investig. Ophthalmol. Vis. Sci. 1994, 35, 1059–1064. [Google Scholar]
- Illingworth, C.D.; Cook, S.D. Acanthamoeba keratitis. Surv. Ophthalmol. 1998, 42, 493–508. [Google Scholar] [CrossRef] [PubMed]
- Zasloff, M. Magainins, a class of antimicrobial peptides from Xenopus skin: Isolation, characterization of two active forms, and partial cDNA sequence of a precursor. Proc. Natl. Acad. Sci. USA 1987, 84, 5449–5453. [Google Scholar] [CrossRef] [Green Version]
- Mookherjee, N.; Anderson, M.A.; Haagsman, H.P.; Davidson, D.J. Antimicrobial host defence peptides: Functions and clinical potential. Nat. Rev. Drug Discov. 2020, 19, 311–332. [Google Scholar] [CrossRef] [PubMed]
- Redd, T.K.; Talbott, M.; Cevallos, V.; Lalitha, P.; Seitzman, G.D.; Lietman, T.M.; Keenan, J.D. In vitro comparison of the Acanthamoeba cysticidal activity of povidone iodine, natamycin, and chlorhexidine. Ophthalmol. Sci. 2021, 1, 100025. [Google Scholar] [CrossRef] [PubMed]
- Mathers, W.D.; Sutphin, J.E.; Folberg, R.; Meier, P.A.; Wenzel, R.P.; Elgin, R.G. Outbreak of keratitis presumed to Be caused by Acanthamoeba. Am. J. Ophthalmol. 1996, 121, 129–142. [Google Scholar] [CrossRef]
- Lim, N.; Goh, D.; Bunce, C.; Xing, W.; Fraenkel, G.; Poole, T.R.; Ficker, L. Comparison of polyhexamethylene biguanide and chlorhexidine as monotherapy agents in the treatment of Acanthamoeba keratitis. Am. J. Ophthalmol. 2008, 145, 130–135. [Google Scholar] [CrossRef]
- Asiedu-Gyekye, I.J.; Mahmood, A.S.; Awortwe, C.; Nyarko, A.K. Toxicological assessment of polyhexamethylene biguanide for water treatment. Interdiscip. Toxicol. 2015, 8, 193–202. [Google Scholar] [CrossRef] [Green Version]
- Ansel, H.C. Hemolysis of erythrocytes by antibacterial preservatives IV. Hemolytic activity of chlorhexidine diacetate. J. Pharm. Sci. 1967, 56, 616–619. [Google Scholar] [CrossRef]
- Kovacs, C.J.; Lynch, S.C.; Rah, M.J.; Millard, K.A.; Morris, T.W. Acanthamoeba encystment: Multifactorial effects of buffers, biocides, and demulcents present in contact lens care solutions. Clin. Ophthalmol. 2015, 9, 1905–1913. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Peguda, H.K.; Carnt, N.A.; Gu, Z.; Kumar, N.; Willcox, M.D.P.; Kuppusamy, R. The Anti-Amoebic Activity of a Peptidomimetic against Acanthamoeba castellanii. Microorganisms 2022, 10, 2377. https://doi.org/10.3390/microorganisms10122377
Peguda HK, Carnt NA, Gu Z, Kumar N, Willcox MDP, Kuppusamy R. The Anti-Amoebic Activity of a Peptidomimetic against Acanthamoeba castellanii. Microorganisms. 2022; 10(12):2377. https://doi.org/10.3390/microorganisms10122377
Chicago/Turabian StylePeguda, Hari Kumar, Nicole A. Carnt, Zi Gu, Naresh Kumar, Mark D. P. Willcox, and Rajesh Kuppusamy. 2022. "The Anti-Amoebic Activity of a Peptidomimetic against Acanthamoeba castellanii" Microorganisms 10, no. 12: 2377. https://doi.org/10.3390/microorganisms10122377