
Epidemiology, Evolution of Antimicrobial Profile and Genomic Fingerprints of Pseudomonas aeruginosa before and during COVID-19: Transition from Resistance to Susceptibility
 ,
,  , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
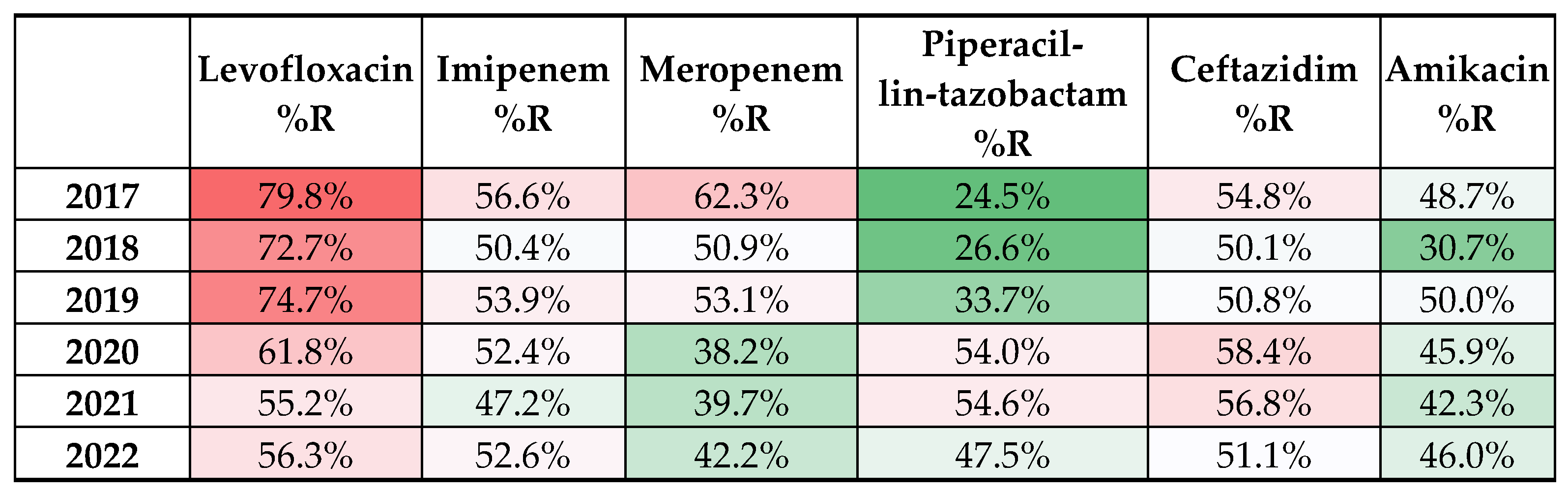
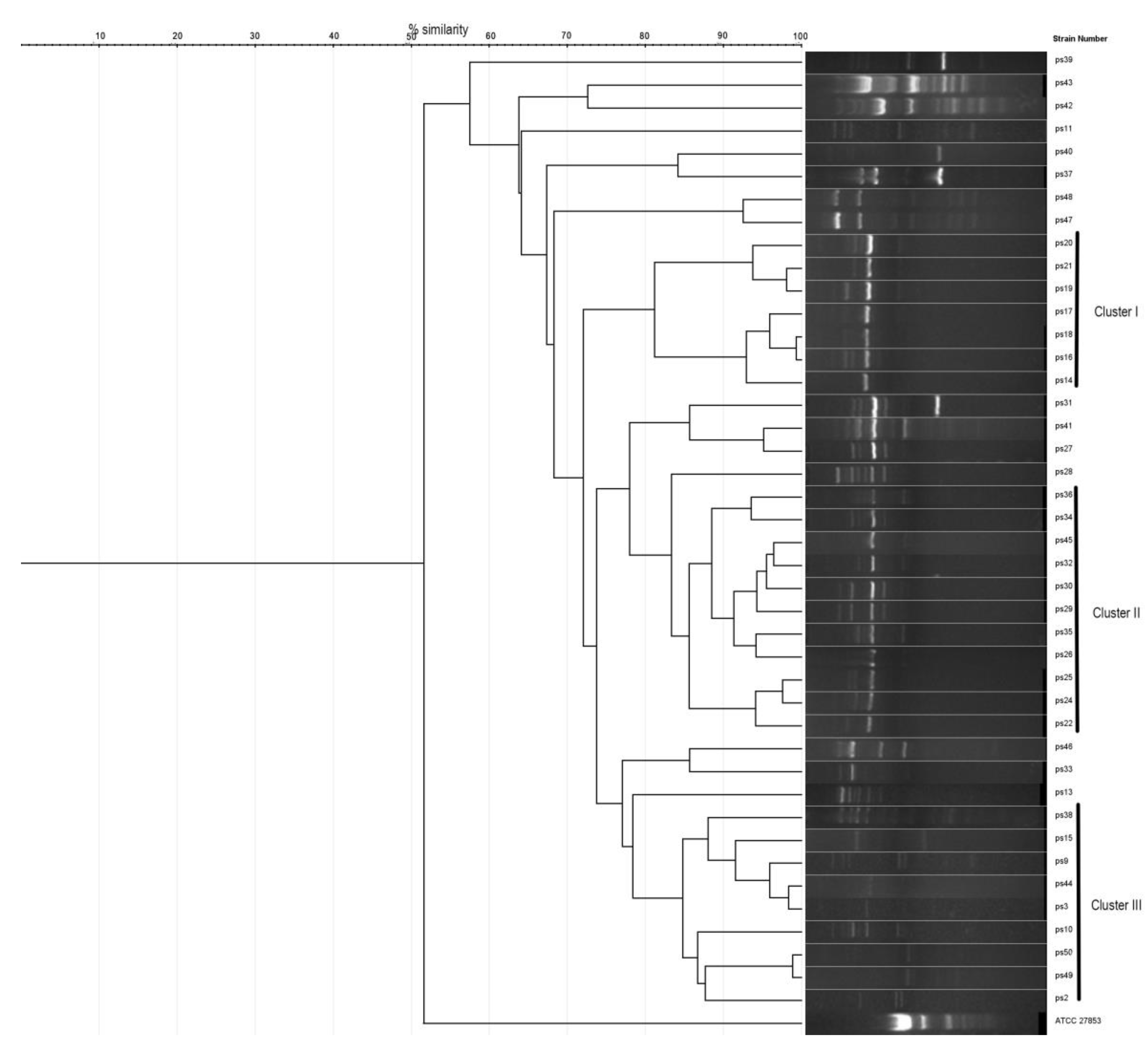
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mita, Y.; Shigemura, K.; Osawa, K.; Kitagawa, K.; Kotaki, T.; Shirakawa, T.; Miyara, T.; Fujisawa, M. Clinical Risk Factors for Death Caused by Extended-Spectrum Beta-Lactamase: Producing Bacteria. Urol. Int. 2019, 102, 205–211. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention (U.S.). Antibiotic Resistance Threats in the United States, 2019; Centers for Disease Control and Prevention (U.S.): Atlanta, GA, USA, 2019.
- Infectious Diseases Kill over 17 Million People a Year: WHO Warns of Global Crisis. Indian Pediatr. 1996, 33, 617–623.
- Akkerman, A.E.; Kuyvenhoven, M.M.; van der Wouden, J.C.; Verheij, T.J. Prescribing Antibiotics for Respiratory Tract Infections by GPs: Management and Prescriber Characteristics. Br. J. Gen. Pract. 2005, 55, 114–118. [Google Scholar] [PubMed]
- Schroeck, J.L.; Ruh, C.A.; Sellick, J.A.; Ott, M.C.; Mattappallil, A.; Mergenhagen, K.A. Factors Associated with Antibiotic Misuse in Outpatient Treatment for Upper Respiratory Tract Infections. Antimicrob. Agents Chemother. 2015, 59, 3848–3852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cepas, V.; Soto, S.M. Relationship between Virulence and Resistance among Gram-Negative Bacteria. Antibiotics 2020, 9, 719. [Google Scholar] [CrossRef]
- Liu, J.-Y.; Dickter, J.K. Nosocomial Infections: A History of Hospital-Acquired Infections. Gastrointest. Endosc. Clin. 2020, 30, 637–652. [Google Scholar] [CrossRef]
- Laxminarayan, R.; Duse, A.; Wattal, C.; Zaidi, A.K.M.; Wertheim, H.F.L.; Sumpradit, N.; Vlieghe, E.; Hara, G.L.; Gould, I.M.; Goossens, H.; et al. Antibiotic Resistance—The Need for Global Solutions. Lancet Infect. Dis. 2013, 13, 1057–1098. [Google Scholar] [CrossRef] [Green Version]
- Samir, A.; Abdel-Moein, K.A.; Zaher, H.M. Emergence of Penicillin-Macrolide-Resistant Streptococcus Pyogenes among Pet Animals: An Ongoing Public Health Threat. Comp. Immunol. Microbiol. Infect. Dis. 2020, 68, 101390. [Google Scholar] [CrossRef]
- Köck, R.; Becker, K.; Cookson, B.; van Gemert-Pijnen, J.E.; Harbarth, S.; Kluytmans, J.; Mielke, M.; Peters, G.; Skov, R.L.; Struelens, M.J.; et al. Methicillin-Resistant Staphylococcus Aureus (MRSA): Burden of Disease and Control Challenges in Europe. Eurosurveillance 2010, 15, 19688. [Google Scholar] [CrossRef] [Green Version]
- ECDC/EMEA Joint Technical Report: The Bacterial Challenge: Time to React. Available online: https://www.ecdc.europa.eu/en/publications-data/ecdcemea-joint-technical-report-bacterial-challenge-time-react (accessed on 24 October 2020).
- Sader, H.S.; Castanheira, M.; Mendes, R.E.; Flamm, R.K. Frequency and Antimicrobial Susceptibility of Gram-Negative Bacteria Isolated from Patients with Pneumonia Hospitalized in ICUs of US Medical Centres (2015–2017). J. Antimicrob. Chemother. 2018, 73, 3053–3059. [Google Scholar] [CrossRef]
- Perovic, O.; Singh-Moodley, A.; Duse, A.; Bamford, C.; Elliott, G.; Swe Swe, K.; Kularatne, R.; Lowman, W.; Whitelaw, A.; Nana, T.; et al. National Sentinel Site Surveillance for Antimicrobial Resistance in Klebsiella Pneumoniae Isolates in South Africa, 2010–2012. S. Afr. Med. J. Suid-Afr. Tydskr. Vir Geneeskd. 2014, 104, 563–568. [Google Scholar] [CrossRef]
- Saleem, A.F.; Ahmed, I.; Mir, F.; Ali, S.R.; Zaidi, A.K. Pan-Resistant Acinetobacter Infection in Neonates in Karachi, Pakistan. J. Infect. Dev. Ctries. 2009, 4, 30–37. [Google Scholar] [CrossRef] [Green Version]
- Thakare, R.; Kesharwani, P.; Dasgupta, A.; Srinivas, N.; Chopra, S. Chapter 1—Antibiotics: Past, Present, and Future. In Drug Discovery Targeting Drug-Resistant Bacteria; Kesharwani, P., Chopra, S., Dasgupta, A., Eds.; Academic Press: Cambridge, MA, USA, 2020; pp. 1–8. ISBN 978-0-12-818480-6. [Google Scholar]
- Hematzadeh, A.; Haghkhah, M. Biotyping of Isolates of Pseudomonas aeruginosa Isolated from Human Infections by RAPD and ERIC-PCR. Heliyon 2021, 7, e07967. [Google Scholar] [CrossRef]
- de Bentzmann, S.; Plésiat, P. The Pseudomonas aeruginosa Opportunistic Pathogen and Human Infections. Environ. Microbiol. 2011, 13, 1655–1665. [Google Scholar] [CrossRef]
- Esposito, S.; De Simone, G. Update on the Main MDR Pathogens: Prevalence and Treatment Options. Infez. Med. 2017, 25, 301–310. [Google Scholar]
- Behzadi, P.; Baráth, Z.; Gajdács, M. It’s Not Easy Being Green: A Narrative Review on the Microbiology, Virulence and Therapeutic Prospects of Multidrug-Resistant Pseudomonas aeruginosa. Antibiotics 2021, 10, 42. [Google Scholar] [CrossRef]
- Zeeshane, M. Diabetic Foot Infection Due to Pseudomonas aeruginosa, Peshawar. J. Wazir Muhammad Inst. Paramed. Technol. 2021, 1, 7–10. [Google Scholar] [CrossRef]
- Botelho, J.; Grosso, F.; Peixe, L. Antibiotic Resistance in Pseudomonas aeruginosa–Mechanisms, Epidemiology and Evolution. Drug Resist. Updat. 2019, 44, 100640. [Google Scholar] [CrossRef]
- Streeter, K.; Katouli, M. Pseudomonas aeruginosa: A Review of Their Pathogenesis and Prevalence in Clinical Settings and the Environment. Infect. Epidemiol. Microbiol. 2016, 2, 25–32. [Google Scholar] [CrossRef]
- Multidrug-Resistant Pseudomonas aeruginosa|A.R. & Patient Safety Portal. Available online: https://arpsp.cdc.gov/profile/antibiotic-resistance/mdr-pseudomonas-aeruginosa?hai-select-resistance-by-state=hai33 (accessed on 28 September 2022).
- European Centre for Disease Prevention and Control; World Health Organization. Antimicrobial Resistance Surveillance in Europe: 2022: 2020 Data; Publications Office: Luxembourg, 2022. [Google Scholar]
- Ali, M.M.; Zahra, Q.U.A.; Khan, M.I. Frequency and Antimicrobial Susceptibility Pattern of Pseudomonas aeruginosa in Human Pus Samples at Holy Family Hospital Rawalpindi. J. Microbiol. Biotechnol. 2021, 6, 000189. [Google Scholar]
- Magill, S.S.; Edwards, J.R.; Bamberg, W.; Beldavs, Z.G.; Dumyati, G.; Kainer, M.A.; Lynfield, R.; Maloney, M.; McAllister-Hollod, L.; Nadle, J.; et al. Multistate Point-Prevalence Survey of Health Care-Associated Infections. N. Engl. J. Med. 2014, 370, 1198–1208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Álvarez-Lerma, F.; Palomar, M.; Olaechea, P.; Otal, J.J.; Insausti, J.; Cerdá, E. Estudio Nacional de Vigilancia de Infección Nosocomial En Unidades de Cuidados Intensivos: Informe Evolutivo de Los Años 2003–2005. Med. Intensiv. 2007, 31, 6–17. [Google Scholar] [CrossRef] [PubMed]
- Vaez, H.; Salehi-Abargouei, A.; Ghalehnoo, Z.R.; Khademi, F. Multidrug Resistant Pseudomonas aeruginosa in Iran: A Systematic Review and Metaanalysis. J. Glob. Infect. Dis. 2018, 10, 212–217. [Google Scholar] [CrossRef] [PubMed]
- Prevalence and Antimicrobial Susceptibility of Pseudomonas aeruginosa Mucoid and Non-Mucoid Type–PubMed. Available online: https://pubmed.ncbi.nlm.nih.gov/15916087/ (accessed on 28 September 2022).
- Ahmed, O.B. Incidence and Antibiotic Susceptibility Pattern of Pseudomonas aeruginosa Isolated from Inpatients in Two Tertiary Hospitals. Clin. Microbiol. Open Access 2016, 5, 1–4. [Google Scholar] [CrossRef] [Green Version]
- Fahim, N.A.E. Prevalence and Antimicrobial Susceptibility Profile of Multidrug-Resistant Bacteria among Intensive Care Units Patients at Ain Shams University Hospitals in Egypt—A Retrospective Study. J. Egypt. Public Health Assoc. 2021, 96, 7. [Google Scholar] [CrossRef]
- Eucast: Expected Phenotypes. Available online: https://www.eucast.org/expert_rules_and_expected_phenotypes/expected_phenotypes (accessed on 6 October 2022).
- Botelho, J.; Roberts, A.P.; León-Sampedro, R.; Grosso, F.; Peixe, L. Carbapenemases on the Move: It’s Good to Be on ICEs. Mob. DNA 2018, 9, 37. [Google Scholar] [CrossRef] [Green Version]
- Thi, M.T.T.; Wibowo, D.; Rehm, B.H.A. Pseudomonas aeruginosa Biofilms. Int. J. Mol. Sci. 2020, 21, 8671. [Google Scholar] [CrossRef]
- Holmes, C.L.; Anderson, M.T.; Mobley, H.L.T.; Bachman, M.A. Pathogenesis of Gram-Negative Bacteremia. Clin. Microbiol. Rev. 2021, 34, e00234-20. [Google Scholar] [CrossRef]
- Kwok, W.C.; Ho, J.C.M.; Tam, T.C.C.; Ip, M.S.M.; Lam, D.C.L. Risk Factors for Pseudomonas aeruginosa Colonization in Non-Cystic Fibrosis Bronchiectasis and Clinical Implications. Respir. Res. 2021, 22, 132. [Google Scholar] [CrossRef]
- Kunadharaju, R.; Rudraraju, A.; Sethi, S. Pseudomonas aeruginosa Colonization and COPD: The Chicken or the Egg? Arch. Bronconeumol. 2022, 58, 539–541. [Google Scholar] [CrossRef]
- Wolska, K.; Szweda, P. A Comparative Evaluation of PCR Ribotyping and ERIC PCR for Determining the Diversity of Clinical Pseudomonas aeruginosa Isolates. Pol. J. Microbiol. 2008, 57, 157–163. [Google Scholar]
- Zarei, O.; Shokoohizadeh, L.; Hossainpour, H.; Alikhani, M.Y. Molecular Analysis of Pseudomonas aeruginosa Isolated from Clinical, Environmental and Cockroach Sources by ERIC-PCR. BMC Res. Notes 2018, 11, 668. [Google Scholar] [CrossRef] [Green Version]
- Mkhize, S.; Amoako, D.G.; Shobo, C.O.; Zishiri, O.T.; Bester, L.A. Genotypic and Phenotypic Characterizations of Methicillin-Resistant Staphylococcus Aureus (MRSA) on Frequently Touched Sites from Public Hospitals in South Africa. Int. J. Microbiol. 2021, 2021, 6011045. [Google Scholar] [CrossRef]
- Jalalifar, S.; Motallebirad, T.; Dashtbin, S.; Mirzaei, R.; Khorshidi, M.; Esfahani, B.N. Molecular Typing of Streptococcus Agalactiae- CMLSB Phenotype Isolates by Enterobacterial Repetitive Intergenic Consensus-PCR (ERIC-PCR) in Isfahan, Iran. Infect. Epidemiol. Microbiol. 2022, 8, 139–147. [Google Scholar] [CrossRef]
- Syrmis, M.W.; O’Carroll, M.R.; Sloots, T.P.; Coulter, C.; Wainwright, C.E.; Bell, S.C.; Nissen, M.D. Rapid Genotyping of Pseudomonas aeruginosa Isolates Harboured by Adult and Paediatric Patients with Cystic Fibrosis Using Repetitive-Element-Based PCR Assays. J. Med. Microbiol. 2004, 53, 1089–1096. [Google Scholar] [CrossRef] [Green Version]
- Parkins, M.D.; Somayaji, R.; Waters, V.J. Epidemiology, Biology, and Impact of Clonal Pseudomonas aeruginosa Infections in Cystic Fibrosis. Clin. Microbiol. Rev. 2018, 31, e00019-18. [Google Scholar] [CrossRef] [Green Version]
- Pirzadian, J.; Persoon, M.C.; Severin, J.A.; Klaassen, C.H.W.; de Greeff, S.C.; Mennen, M.G.; Schoffelen, A.F.; Wielders, C.C.H.; Witteveen, S.; van Santen-Verheuvel, M.; et al. National Surveillance Pilot Study Unveils a Multicenter, Clonal Outbreak of VIM-2-Producing Pseudomonas aeruginosa ST111 in the Netherlands between 2015 and 2017. Sci. Rep. 2021, 11, 21015. [Google Scholar] [CrossRef]
- Opperman, C.J.; Moodley, C.; Lennard, K.; Smith, M.; Ncayiyana, J.; Vulindlu, M.; Gafoor, M.; Govender, N.; Ismail, H.; Bamford, C.; et al. A Citywide, Clonal Outbreak of Pseudomonas aeruginosa. Int. J. Infect. Dis. 2022, 117, 74–86. [Google Scholar] [CrossRef]
- Bertrand, X.; Bailly, P.; Blasco, G.; Balvay, P.; Boillot, A.; Talon, D. Large Outbreak in a Surgical Intensive Care Unit of Colonization or Infection with Pseudomonas aeruginosa That Overexpressed an Active Efflux Pump. Clin. Infect. Dis. 2000, 31, e9–e14. [Google Scholar] [CrossRef]
- ORDIN 1101 30/09/2016–Portal Legislativ. Available online: https://legislatie.just.ro/Public/DetaliiDocument/182388 (accessed on 28 September 2022).
- Surveillance Atlas of Infectious Diseases. Available online: https://www.ecdc.europa.eu/en/surveillance-atlas-infectious-diseases (accessed on 28 September 2022).




{kind=link}
{kind=link}
{kind=link}
| Year | Total Number of Bacterial Isolates in MCCH | Total Number of Pseudomonas spp. Isolated (n, %) | Males (n, %) | Females (n, %) | Average Age (Years) (SD-Standard Deviation) |
|---|---|---|---|---|---|
| 2017 | 2556 | 547 (21.4%) | 289 (52.83%) | 258 (47.17%) | 68 (SD = 14.1) |
| 2018 | 4243 | 456 (10.74%) | 252 (55.26%) | 204 (44.74%) | 67 (SD = 15.65) |
| 2019 | 3559 | 434 (12.19%) | 223 (51.38%) | 211 (48.62%) | 66 (SD = 16.21) |
| 2020 | 658 | 117 (17.78%) | 66 (56.41%) | 51 (43.59%) | 65 (SD = 17.33) |
| 2021 | 1879 | 299 (15.91%) | 163 (54.51%) | 136 (45.49%) | 66 (SD = 15.68) |
| 2022 | 1509 | 141 (9.43%) | 80 (56.73%) | 61 (43.27%) | 65 (SD = 16.19) |
| Year | 2018 | 2019 | 2020 | 2021 | 2022 |
|---|---|---|---|---|---|
| 2017 | p = 0.82 OR: 0.93 | p = 0.43 OR: 0.94 | p= 0.0041 OR: 1.37 | p < 0.0001 OR: 1.23 | p = 0.0013 OR: 0.72 |
| 2018 | - | p = 0.94 OR: 1.00 | p = 0.0007 OR: 1.46 | p = 0.0007 OR: 1.31 | p = 0.01 OR: 0.77 |
| 2019 | - | - | p = 0.0009 OR: 1.45 | p = 0.001 OR: 1.3 | p = 0.01 OR: 0.76 |
| 2020 | - | - | - | p = 0.37 OR: 0.52 | p < 0.0001 OR: 0.52 |
| 2021 | - | - | - | - | p < 0.001 OR: 0.58 |
| Number of Pseudomonas Isolates (n = 1994) from 2017–2022 | |||||||
|---|---|---|---|---|---|---|---|
| Species | P. aeruginosa | P. alcaligenes | P. fluorescens | P. putida | P. stutzerii | Pseudomonas spp. | |
| 2017 | 98.35% (n = 538) | - | 0.18% (n = 1) | 0.18% (n = 1) | - | 1.27% (n = 7) | |
| 2018 | 98.02% (n = 447) | - | 0.21% (n = 1) | - | - | 1.75% (n = 8) | |
| 2019 | 96.08% (n = 417) | - | 0.46% (n = 2) | - | - | 3.45% (n = 15) | |
| 2020 | 100% (n = 117) | - | - | - | - | - | |
| 2021 | 97.99% (n = 293) | - | 0.66% (n = 2) | 1.00% (n = 3) | - | 0.33% (n = 1) | |
| 2022 | 92.19 (n = 130) | 0.7% (n = 1) | 2.12% (n = 3) | 1.41% (n = 2) | 3.54% (n = 5) | - | |
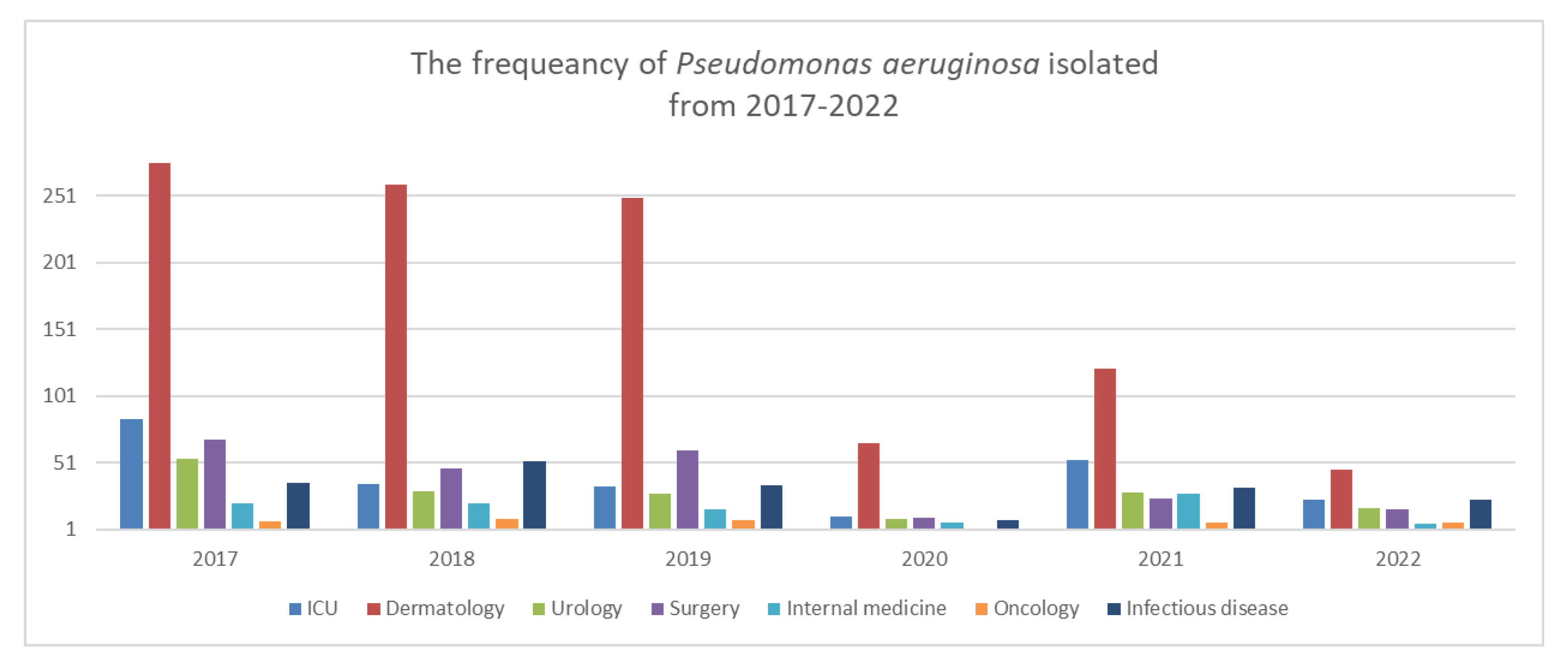
| Department | |||||||
| Years | 2017 | 2018 | 2019 | 2020 | 2021 | 2022 | |
| ICU | 15.35% (n = 84) | 7.67% (n = 35) | 7.6% (n = 33) | 9.4% (n = 11) | 17.72% (n = 53) | 16.31% (n = 23) | |
| Surgical | 12.43% (n = 68) | 10.3% (n = 47) | 13.82% (n = 60) | 0.85% (n = 10) | 8.02% (n = 24) | 11.34% (n = 16) | |
| Medical | 3.83% (n = 21) | 4.6% (n = 21) | 3.68% (n = 16) | 5.12% (n = 6) | 9.36% (n = 28) | 3.54% (n = 5) | |
| Infectious diseases | 6.58% (n = 36) | 11.40% (n = 52) | 7.83% (n = 34) | 6.83% (n = 8) | 10.7% (n = 32) | 16.31% (n = 23) | |
| Dermatology | 50.63% (n = 277) | 57.45% (n = 262) | 58.75% (n = 255) | 60.68% (n = 71) | 42.47% (n = 127) | 36.17% (n = 51) | |
| Oncology | 1.27% (n = 7) | 1.97% (n = 9) | 1.84% (n = 8) | 1.7% (n = 2) | 2.00% (n = 6) | 4.25% (n = 6) | |
| Urology | 9.87% (n = 54) | 6.57% (n = 30) | 6.45% (n = 28) | 7.69% (n = 9) | 9.69% (n = 29) | 12.05% (n = 17) | |
| Infection site/Pathological products | |||||||
| 2017 | 2018 | 2019 | 2020 | 2021 | 2022 | Total | |
| LRT 1 | 8.75% (n = 48) | 9.21% (n = 42) | 2.76% (n = 12) | 11.11% (n = 13) | 16.72% (n = 50) | 4.96% (n = 7) | 172 |
| Pus | 74.77% (n = 409) | 74.56% (n = 342) | 83.17% (n = 361) | 76.06% (n = 89) | 60.86% (n = 182) | 60.28% (n = 85) | 1468 |
| Urine | 12.61% (n = 69) | 12.93% (n = 59) | 9.9% (n = 43) | 11.11% (n = 13) | 15.38% (n = 46) | 21.27% (n = 30) | 260 |
| Blood | 0.91% (n = 5) | 0.87% (n = 4) | 0.46% (n = 2) | 0.85% (n = 1) | 2.00% (n = 6) | 1.41% (n = 2) | 20 |
| Others | 2.92% (n = 16) | 1.97% (n = 9) | 3.68% (n = 16) | 0.85 %(n = 1) | 5.01% (n = 15) | 12.05% (n = 17) | 74 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Coșeriu, R.L.; Vintilă, C.; Mare, A.D.; Ciurea, C.N.; Togănel, R.O.; Cighir, A.; Simion, A.; Man, A. Epidemiology, Evolution of Antimicrobial Profile and Genomic Fingerprints of Pseudomonas aeruginosa before and during COVID-19: Transition from Resistance to Susceptibility. Life 2022, 12, 2049. https://doi.org/10.3390/life12122049
Coșeriu RL, Vintilă C, Mare AD, Ciurea CN, Togănel RO, Cighir A, Simion A, Man A. Epidemiology, Evolution of Antimicrobial Profile and Genomic Fingerprints of Pseudomonas aeruginosa before and during COVID-19: Transition from Resistance to Susceptibility. Life. 2022; 12(12):2049. https://doi.org/10.3390/life12122049
Chicago/Turabian StyleCoșeriu, Răzvan Lucian, Camelia Vintilă, Anca Delia Mare, Cristina Nicoleta Ciurea, Radu Ovidiu Togănel, Anca Cighir, Anastasia Simion, and Adrian Man. 2022. "Epidemiology, Evolution of Antimicrobial Profile and Genomic Fingerprints of Pseudomonas aeruginosa before and during COVID-19: Transition from Resistance to Susceptibility" Life 12, no. 12: 2049. https://doi.org/10.3390/life12122049